
Home » Race Realism (Page 13)
Category Archives: Race Realism
Just-so Stories: Christianity
1200 words
Storytelling in religions is ubiquitous. Storytelling can be found in all religions for myriad reasons, including—but not limited to—(1) the passing of traditions; (2) how the mores of the society in question should be structured (Wuthnow, 2011); (3) bringing people into the religion by creating origin myths, and so on. In short, the function of religious storytelling has numerous primary directives, but one of the most important is answering why questions (Braustein, 2012) (i.e., “Why are we here?”, “Why do we do X?”), not how questions (i.e., “How did we get here?”). Thus, the importance of religious storytelling seems to be obvious: religious storytelling serves its function in virtue of explaining why questions which were formulated by a human mind. It also, for billions of people around the world, grounds morality in an objective reality given to Man by God.
If religious storytelling can answer how questions, then, it is proposed, we can believe these stories as evidence that the religions in question are true. However, there is much pushback in believing these religious stories. Religious stories are what can be termed “just-so stories”, which amount to hypotheses which offer “little in the way of independent evidence to suggest that it is actually true” (Law, 2016). It is for this reason that we cannot—nor should we—accept any religious just-so stories: they lack sufficient evidence for belief, other than what they purport to explain. In this article, I will discuss certain just-so stories and Christianity—how they fail to successfully argue what they purport to but still serve important (moralistic) functions. Religion and language/writing are deeply intertwined, and so, understanding how and why we speak and write along with the history of civilization will help us to better understand how and why we tell these kinds of stories.
Religion has been around as long as agriculture (Peoples, Duda, and Marlowe, 2016), and so, if we understand the co-evolution of both of these variables, then we may understand how and why these types of stories have persisted through the ages. Storytelling—in a religious manner—is a way to ground a certain group’s morals in something objective. This is one very important reason that religious storytelling has persisted. For example, Christians argue that Jesus rose from the dead in 3 days and that God created the universe we live in, and so on. These two claims, though, are just-so stories since there is no independent evidence for the claims. There are other more important functions to Christian storytelling other than believing in the ultimate truths of the purported claims.
Stories have many functions—one of the many functions is to give purpose to one’s life. Yet another function is to guide how one lives their lives using stories that were created thousands of years ago. Storytelling in Christianity is a large topic—one that is held dear to many people. What are some of the stories and are there any independent reasons to believe them?
Creationists—those who believe that a Supreme Being created everything we see around us—use the Bible and its stories as evidence that evolution is false and Creationism is true. One prominent Creationist is Dr. Bo Kirkwood who wrote The Evolution Delusion: A Scientific Study of Creation and Evolution (Kirkwood, 2016). In the book, he attempts to use Christian storytelling to attempt to discredit the fact of evolution. He makes many questionable claims including “God’s fingerprints are seen by observing his finely tuned universe” (Kirkwood, 2016: 210). This is a perfect example of a just-so story—make a claim that has no evidence to support it, the only evidence to support it is the claim (that “God’s fingerprints are seen by observing his finely tuned universe”).
No evidence exists that the universe is “God’s fingerprints … finely tuned” the universe, and so it is a just-so story. The arguments pushed by Creationists such as Kirkwood (2016) do not make any testable predictions. The Big Bang theory, however, does make testable predictions and is, therefore, not a just-so story. Note how Kirkwood’s (2016) claim have no independent evidence to back them—that God “finely tuned” the universe is one of the ultimate just-so stories. It’s literally based off of faith and no evidence. Nothing can falsify the claim that God “finely tuned” the universe, and so we should disregard claims like this from Creationists. Kirkwood’s (2016) claims that the universe is “finely tuned” by God is an attempt to argue that there is a Creator of the universe we live in, and so we should dispense with evolutionary thinking and embrace the Creator, God. Kirkwood (2016) believes that, if evolutionary theory can be disproven by appealing to claims that God “finely tuned” the universe, then more people would believe God’s Word and society would start trending in a more religious, Christian trajectory.
Just because claims from Creationists are false does not mean that religious storytelling has no use—no function—in the modern day (or even thousands of years ago when they were first formulated) One of the most important functions for storytelling in Christianity is the grounding of objective morality. Most of the stories told in the Bible have a moralistic message that it wants to convey. Do this, not that, because God is watching and this objective morality is grounded by God. These stories, then, got passed down through each successive generation and eventually became got sorted into what is now the Bible during the Council of Nicaea.
One of the best examples of morality in the Bible—and the stories that accompany it—is Romans 13: 8-10, where it is stated that, many of the Commandments can be summed up succinctly as “Love does no harm to a neighbor. Therefore, love is the fulfillment of the law” (New International Version). The Ten Commandments were given to Moses by God – they were given to Moses to give to the Israelites in order to guide them to live good, moral lives. So, if “… whoever loves others has fulfilled the law” (New International Version), and those who follow the Ten Commandments have fulfilled the law by loving—not harming—their neighbors, then those who follow the story that Moses was given the Commandments by God are living a moral life and therefore these stories help people in our societies live moral lives. That these stories help billions of Christians live moral lives does not speak to the truth of the claim that these Commandments were given to Moses by God, though. That is irrelevant. The most important thing here is that these stories help others live moral lives. (Quite obviously, the stories are not sufficient to live a moral life, since there are non-religious people who do so.)
Religious storytelling—irrespective of the truth of the stories—is clearly important in our societies. They help people with certain moral quandaries, to live their lives a certain way because they believe that someone is watching them. If these stories help people live good, moral lives, then it does not matter if the ultimate claims of the stories are true or not. Therefore, Christian storytelling has very important functions for how our current societies function which is why Christian storytelling has persisted through the ages, even if the claims are false.
Strength and Neuromuscular Coordination
2250 words
PumpkinPerson (PP) has some weird—and uneducated—views regarding strength and coordination, which, of course, implies that he has no understanding of what “coordination” truly is. He seems to have convinced himself that coordination weightlifting does not require coordination (neuromuscular coordination; hereafter NMC—the ability of the central nervous system—CNS—to control muscles). That view is patently ridiculous. In this article, I will explain the logic behind the fact that strength and power exercises, in particular, NEED a high NMC, and without a high NMC, the athlete in question cannot perform to their highest potential.
PP wrote about an “athletic g factor” to attempt to liken it to the “g factor” regarding “intelligence” tests, but I’m not worried about that comparison (IQ is boring to me now). What I am worried about are his outlandish claims regarding what he believes regarding strength and NMC. PP cited Jensen’s Bias in Mental Testing where Jensen cited a correlation matrix in which “all of [the] correlations were positive“, writing that he’s “not sure why some commenters think weight lifting requires coordination when the correlation between strength (hand grip, chinning) and coordination (Pursuit rotor tacking, Mirror star tracing) is zero” (PP; Physical Coordination).
Well, “some commenters” have actual experience in what he is talking about, so, forgive me if I don’t believe the claims that, in my opinion, he pulled out of thin air. Take chin-ups. Imagine a case of someone attempting to chin-up that does not have high NMC. Since they were not coordinated, do you think they would be able to do a controlled rep in order to complete one rep? Or would their body be all over the place, flailing around since they do not have the mind-muscle (MMC) connection required to complete the lift. Now take his other example, hand grip. On its face, one might assume that this requires no NMC. But think about the process of gripping something tightly. If the muscles in the forearm, for example, are not adequately trained, then, in all lifts involving forearm strength (a great majority of which involve at least some type of forearm strength) will not be able to be performed properly, since the individual in question does not have the NMC required to properly do the exercise in question.
PP then says that when he “lift weights, [he doesn’t] feel like [he’s] using coordination.” This proves two things to me: (1) PP does not know how to lift properly, and then (2) follows that he does not know about the MMC.
The MMC is where the mind and the body “meet.” Acetylcholine functions as a neurotransmitter. This neurotransmitter “communicates” with the muscles in the body to cause a contraction. This contraction, then, causes the action of voluntary muscle movement. (I had an A&P professor explain to me that, out of the whole textbook he taught out of, one of the only things in the textbook that we could choose to do was move the body—contract muscles and cause movement). So when acetylcholine is released, it latches onto muscle fibers and causes muscle contractions.
We can put the MMC in this way: imagine doing a movement such as a bicep curl. One is not actively attempting to use the proper levers in order to properly lift the weight. On the other hand, if one is actively thinking about the muscles being used in the movement, then they are using the connection—they are strengthing their MMC and, in turn, developing the proper NMC which is required in order to properly lift weights and get the most returns possible from your time spent lifting.
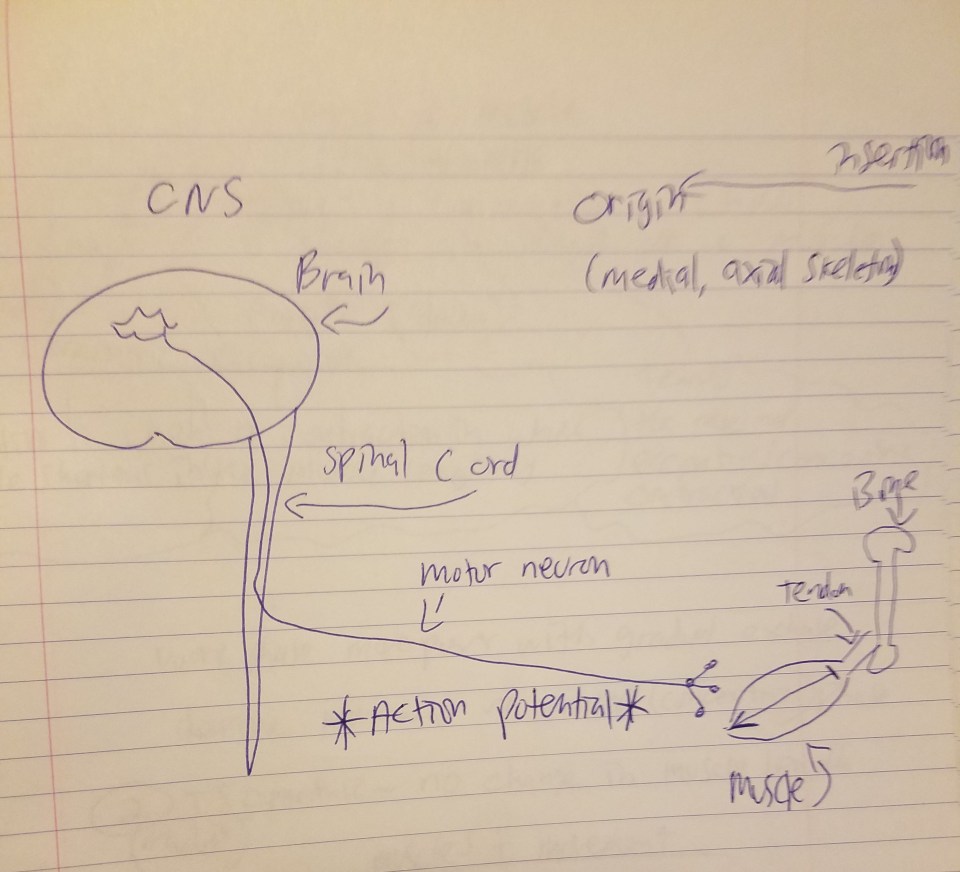
The above diagram I drew is the process by which muscle action occurs. In my recent article on fiber typing and metabolic disease, I explained the process by which muscles contract:
But the skeletal muscle will not contract unless the skeletal muscles are stimulated. The nervous system and the muscular system communicate, which is called neural activiation—defined as the contraction of muscle generated by neural stimulation. We have what are called “motor neurons”—neurons located in the CNS (central nervous system) which can send impulses to muscles to move them. This is done through a special synapse called the neuromuscular junction. A motor neuron that connects with muscle fibers is called a motor unit and the point where the muscle fiber and motor unit meet is callled the neuromuscular junction. It is a small gap between the nerve and muscle fiber called a synapse. Action potentials (electrical impulses) are sent down the axon of the motor neuron from the CNS and when the action potential reaches the end of the axon, hormones called neurotransmitters are then released. Neurotransmitters transport the electrical signal from the nerve to the muscle.
So action potentials (APs) are carried out at the junction between synapses. So, regarding acetylcholine, when it is released, it binds to the synapses (a small space which separates the muscle from the nerve) and it then binds onto the receptors of the muscle fibers. Now we know that, in order for a muscle to contract, the brain sends the chemical message (acetylcholine) across synapses which then initiates movement. So, as can be seen from the diagram above, the MMC refers to the chemo-electric connection between the motor cortex, the cortico-spinal column, peripheral nerves and the neuromuscular junction. A neuromuscular junction is a synapse formed by the contact between a motor neuron and a muscle fiber. This is why beginners in the gym get stronger in the first 8 weeks or so of training—there has not been enough time for muscle to adequately grow in that time span. Thus, when people lift weights correctly, what they are doing is training their NMC—and their mind—to be able to adequately perform these types of actions in a safe, controlled manner.
How is NMC measured? It’s not simple to measure it, and in reality, the most feasible way to “measure it” in real life situations without the use of a lab is to just see one’s progress while they progress through higher and higher weights from their starting weights and they learn to perform the exercise in question safely. But a more empirical measure used in order to measure NMC are electromyography (EMG) tests. In fact, this test is THE MEASURE used to measure NMC, since all of the relevant variables in question (some seen in the above diagram) are tested. EMGs are used for numerous reasons, mostly in order to test for types of motor diseases which affect muscle action. There is also a related measure here: a nerve conduction study. This measures the speed and strength of signals traveling between two synapses, and so, the better one’s nerve conduction is in regard to muscle action, the higher their NMC is and, therefore, the better they will be able to perform any certain lift. So, for example, we can say that one’s NMC increased and the cause was resistance training if their EMG tests increase.
Imagine an Olympic lifter going to snatch 400 pounds. Would any sane person bet that they have low NMC (i.e., a low rate of firing between synapses as measured by an EMG)? A claim such as this would be quite preposterous—individuals like Olympic lifters clearly have trained both their bodies and minds in order to lift to the best of their abilities. And if they did NOT have high NMC (i.e., a higher rate of firing between synapses), then the weight would wobble and ultimately fall, causing the lifter serious injury. But, of course, we do not see that, since strength and NMC are closely related.
I now have some examples of studies which looked into this matter (that thinking about the action one is performing activates the primary muscles used in the movement in question), which will definitely put PP’s claims to rest for good.
Neuromuscular coordination is needed, for example, to be able to “squat lift” correctly (meaning, pick up a load from a squatting start and lift it; Scholz, Millford, and McMillan, 1995). Our understanding of how this occurs has greatly increased in 30 some-odd years since our technology has improved.
Now, take the MMC. We can simply define it as “One focusing on using the muscles in question to perform the lift.” Calatayud et al (2016) studied 18 resistance-trained men on a 1RM (one-rep max) bench press. Each individual in the study participated in 2 sessions: one to determine their 1RM and another experimental session. Calatayud et al (2016) attempted to control for as many factors as possible in order to attempt to see if the baseline changed at all. For example, all measures were made by the same two investigators; all measures were taken in the same facility; all participants participated in the same warm-up mobility drills prior to performing the lift; all participants performed the lift in the exact same manner they performed the two aforementioned sessions (same technique and body position, i.e., suicide grip and powerlifting technique).
They found that (1) higher levels of EMG activity lead to moving more weight; (2) the men could “selectively activate pectoralis and triceps muscles during the
bench press when this exercise is performed at low intensities” (Calatayud et al, 2016), at moderate intensities; (3) that focusing on one muscle (i.e., triceps brachii over pec major) did not hamper activation in one over the other; and (4) a threshold exists between 60-80 percent existed for muscle activation. Thus, experienced resistance-trained men can actively increase activity in certain muscles when cued to focus on those certain muscles.
Snyder and Fry (2012) studied 11 D-III football players on the bench press while recording EMG activity. They found that, when verbal cues were given to focus on the chest muscles, EMG increased by 22 percent, but when verbally cued to focus on the triceps, the pec major returned to baseline (though this does not mean, of course, that performance was hampered), while EMG activity increased by 26 percent. However, in-line with the findings from Calatayud et al (2016), when 80% 1RM were tested, EMG activity in the triceps remained unchanged, implying that there is a threshold.
The results of this study show that trained subjects can alter the participation of muscles in both moderate and higher-intensity multijoint resistance training exercises in response to verbal instructions, because both TB and PM activities were increased selectively in response to 2 different sets of instructions at 50% 1RM and 80% 1RM. This indicates that verbal instructions from trainers, therapists, and coaches are likely to have a measurable effect on muscle involvement, although it is unclear how generalizable this effect might be to all training exercises. Previous research from our laboratory (23) indicated that untrained subjects performing a lat pull-down at 30% max isometric load could respond to verbal instructions to increase back muscle involvement by increasing latissimus dorsi activity while maintaining proper form and similar speed of movement. The subjects in that study increased latissimus dorsi activity by 17.6%, whereas in the current study, verbal instruction resulted in a 22.3% increase from baseline at 50% 1RM for PM and a 25.6% increase for TB. However, antagonist activity was not measured by Snyder and Leech (23), and it was possible that the subjects activated antagonist muscles to offset additional force produced by agonist muscles. This study addressed this possibility, but no changes were seen in antagonist muscle activity with verbal instructions. The question of the effect of higher testing loads was also addressed by this study, and it was found that at 50% 1RM, the subjects were capable of altering muscle participation of both the horizontal adductors and the elbow extensors, but at 80% 1RM, only the horizontal adductors were affected. (Snyder and Fry, 2012)
If the activity of a muscle as measured by EMG is increased, then we can say that, for all intents and purposed, that NMC is high. One who is not familiar with a lift will have low NMC, that is, the firing will be low compared to someone with high NMC. Quite clearly, verbal instruction to focus on certain muscles can better activate them, and, using EMG, we can say that they have high NMC if the firing between synapses is fast.
Rutherford and Jones (1986) write that “It is concluded that a large part of the improvement in the ability to lift weights was due to an increased ability to coordinate other muscle groups involved in the movement such as those used to stabilise the body.” How weird is that… While Kim, Lockhart, and Roberto (2009) in their sample of elderly individuals found that “Strength gain by exercise training plays a role in the improved coordination of other fixator muscles necessary for body support while performing daily tasks such as cooking, gardening, reaching for an object, and walking, and in gaining more coordinated contractions between agonist and antagonist muscle groups leading to greater net force in the imposing movements.” Finally, Dahab and McCambridge (2009) found that strength training in kids improves the number and coordination of active neurons along with the firing rate pattern. This is important because the number and coordination of active neurons along with the rate of firing pattern influences—very strongly—NMC and how coordinated they will be.
In conclusion, it is quite obvious that PP does not know what he is talking about and only writes what sounds good in his head without having an adequate understanding of anatomy and physiology, NMC, MMC, APs and the like. These types of confusions can be cleared up by having an adequate understanding of anatomy and physiology and knowing how and why muscle actions are done, where they begin and where they end. Clearly, the claim that weight lifting requires no coordination is false.
Dorothy Roberts on Race as a Political Entity
2050 words
I recently bought Dorothy Roberts’ Fatal Invention: How Science, Politics and Big Business Re-create Race in the Twenty-First Century (2011) (it was $1.99 in the nook store, couldn’t pass it up), which, how the title of the book explains, discusses how race is recreated today using methods of the past as well as methods of the future. One of her main claims is that race is a political entity. Now, while I don’t disagree here (there are of course social aspects to what we call “races”), she completely rides against biological racial realism (eg Spencer, 2014; Hardimon, 2017). Her concept, though, is similar to Hardimon’s (2017) socialrace concept, and it is already a part of Spencer’s Blumenbachian partitions (since race is both biologically and socially constructed in the American view of race). While I do not believe that you need genes to delineate race, Roberts also goes on the attack on Rosenberg et al (2002), who both Hardimon and Spencer cite to buttress their arguments on the reality of biological races.
Race is not a biological category that is politcally charged. It is a political category that has been disguised as a biological one. (Roberts, 2011: 14)
Note how this is extremely similar to Hardimon’s socialrace concept. In Hardimon’s concept, socialraces have a biological correlate: minimalist races. Hardimon’s concept says, for example, that “Hispanics/Latinos” are socialraces but they are a group that do not have a corresponding minimalist race—because “Hispanics/Latinos” are a mixture of different races. Race IS a biological category that has been politically charged: Groups look different; groups that look different share geographic ancestry; groups that look different that share geographic ancestry are derived from the same geographic location; therefore race is a biological category and is therefore politically charged (one reason) since people do not like the out-group—people that look different from themselves.
This distinction is important because many people misinterpret the phrase “race is socially constructed” to mean that the biological category of race has a social meaning, so that each society interprets differently what is means to belong to a biological race. According to this view, first we are born into a race, and then our society determines the consequences of this natural inheritance. There is, then, no contradiction between seeing race as both biological and socially constructed. (Roberts, 2011: 14)
There, actually, IS NO CONTRADICTION between seeing race as socially and biologically constructed. Racial categories pick out real kinds in nature—which is what “biological racial realism” means. Since our racial categories pick out real kinds in nature, then, when it comes to society and social construction, whatever is believed about certain races in that society will be socially constructed. You can’t, for example, call a Nigerian Caucasian (see more on this below) because it does not make any sense.
Roberts then goes on (p. 14-15) about how “human beings do not fit the zoological definition of race” since a “biological race is a population of organisms that can be distinguished from other populations in the same species based on differences in inherited traits.” And so, since no human groups have this high degree of genetic differentiation, there are no human races, but only one human race.
Though Hardimon (2017: 99) articulates the best definition of race I have come across:
A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.
So we know that (1) populations exhibit distinctive features; (2) these populations that exhibit these distinctive features correspond to that population’s geographic ancestry, (3) these populations that exhibit these distinctive features which correspond to geographic ancestry belong to a biological line of descent which was initiated by reproductive isolated and geographically separated founding populations; so (4) race exists.
We know race is a political grouping because it has its political roots in slavery and colonialism, it has served its political function over the four hundred years since its inceptio, and its boundary lines—how many there are and who belongs to each one—have shifted over time and across nations to suit those political purposes. Who qualifiies as white, black, and Indian has been the matter of countless rule changes and judicial decisions. These racial reclassifications did not occur in response to scientific advances in human biology, but in response to sociopolitical imperatives. They reveal that was is being defined, orgainzed, and interpreted is a political relationship and not an innate classification. (Roberts, 2011: 15)
We can take this two ways here: (1) point out that Roberts is conflating minimalist/populationist races with socialraces (which is exactly what she is describing to the tee). Yes, since race is partly social, then, based on the social attitudes of people which do change over time. Then, in that society, certain groups who were barred from being in another group may be allowed “into” the group. This does not mean that race is not biological. “Oh the Irish were considered “not white” at one point in time, therefore race doesn’t exist since groups can exit group A and become group B based on sociopolitical inclinations.” This, of course, goes over the distinctive phenotypic differences between groups with peculiar geographic ancestry. THAT is what defines race; what Roberts is discussing is important, since race is partly political, but it is not the whole story.
In addition to the grotesque lynchings that terrorized blacks throughout the South, an especially brutal form of reenslavement was the false imprisonment of thousands of black men who were then leased to white farmers, entrepreneurs, and corporations as a source of cheap labor.
It is in this accute distinction that between the political status of whites and blacks, this way of governing the power relationship between them, that we find the origins of race. Colonial landowners inherited slavery as an ancient practive, but they invented race as a modern system of power. They employed Aristotle’s concept of natural slaves and natural rulers to define permanent features of black and white people. Race separated human beings into two fundamentally distinct groups: those who were indelibly born to be lifelong servants and those who were born to be their masters. Race radically transformed not only what it meant to be enslaved, but what it meant to be free. (Roberts, 2011: 23)
Let’s accept Roberts’ argument here that the political status of whites and blacks was a way to govern the power relationship between them: so what? That group A subjugated group B and attempted to justify it with X, Y, and Z doesn’t mean that group A and group B are not biological races—it just means that group A subjugated group B and, in the future, there were social repercussions (which is also a part of the phenomenon of race as a partly social construct).
Roberts then discusses the Census (p. 31-35) and how ever-changing racial definitions undermine the claim that biological racial realism is true. In Spencer’s argument, the US meaning of “race” is just a referent, “specifically the referent of US racial discourse” (Spencer, 2014: 1027). This is because, in America, race-talk is tied to the census. We Americans are familiar with the racial groupings on the census since they are not only in use on the census but numerous other institutions. Spencer (2014) then discusses how we can use “phonetic cues alone (e.g., African American Vernacular English), surnames alone (e.g., “Chen”), first names alone (e.g., “Lakisha”), and visual cues alone (e.g., a person’s face)” (Spencer, 2014: 1027) to know someone’s race. Therefore, according to Spencer, the discourse used in the census is the discourse used nation-wide.
But the census does not set what “race” means on these forms: the OMB (Office of Management Budgeting) does. The OMB refers to race as a “set” of populations, and so this leads Spencer to believe that the “sets” of populations that the OMB is referring to are whites, blacks, Asians, Pacific Islanders, and American Indians. So race is a particular, not a kind as Hardimon argues.
Roberts then argues against the “new racial science”, most forcefully, against Rosenberg et al (2002). She brings up the usual discourse “…the number of genetic clusters is dictated by the computer user, not the computer program” (Roberts, 2011: 74). Roberts says that the clusters are “arbitrary.” Roberts says that Rosenberg et al’s (2002) study failed to verify 18th-century racial typology, but it did confirm what we have known since Lewontin’s (1972) analysis: that there is more genetic variation within races than between them. About 93-95 percent of human genetic variation was found to be within race whereas 5-7 percent of human genetic variation was found to be between groups.
Roberts says that the clusters are “arbitrary.” This is a common critique, but it is irrelevant. The five populations found by Structure are genetically structured—they are meaningfully demarcated on the basis of genetic markers (Hardimon, 2017: 88). Roberts also discusses the K = 6 run, which identified the Kalash people.
The fact that structure represents a population as genetically distinct does not entail that the population is a race. Nor is the idea that populations corresponding to the five major geographic areas are minimalist races undercut by the fact that structure picks out the Kalash as a genetically distinct group. Like the K=5 graph, the K=6 graph shows that modulo our assumption, continental-level races are genetically structured” (Hardimon, 2017: 88).
The five clusters identified by Rosenberg et al (2002) represent continental-level minimalist races so the five populations which correspond to the major geographic locations throughout the world are continental-level minimalist races. So it is, in fact, possible to place individuals into different their continental-level minimalist race without knowing anything about the race or ancestry of the individuals from which the microsatellites were drawn. Rosenberg et al (2002) studied the populations based on language, culture, and geography, not skin color or race.
It is true that Rosenberg et al (2002) found 4.3 percent of the overall human genetic variation to be between races—but this does not ride against claims from biological racial realists. The genetic variation is enough to say that we have partitions at K = 5.
“People are born with ancestry that comes from their parents but are assigned a race” is how Camara Jones, a research director at the Centers for Disease Control (CDC) explains it. (Roberts, 2011: 77)
People are assigned races based on the ethnicity/ancestry of the parents. A Nigerian would not be assigned to the Asian race, since the Nigerian has none of the features which make “Asians” Asian.
This is very simple: if both parents belong to race R, then the child will be race R as well. If parent 1 belongs to R1 and so does parent 2, then the child will belong to R1 as well (since the parents have distinct physical features which correspond with geographic ancestry and their ancestors derived from a distinct geographic location. So, since people are born with ancestry that comes from their parents, then they are assigned their PARENT’S race; they are not assigned A race, as if one can assign any individual to any race. But what if one parent belongs to R1 and the other belongs to R2? Hardimon’s minimalist concept is vague here; it only shows that races exist, it does not say which populations are races. If an individual’s parents belong to R1 and R2, then that individual is mixed race. The existence of mixed race people, of course, does not rail against the existence of biological races.
In sum, Roberts does make some good points (in what I have read of the book so far), but she gets it wrong on race. Hardimon and Spencer have both defended the methodology/concepts used by Rosenberg et al (2002) and in doing so, they successfully argued for the existence of biological races—though their two viewpoints differ. That race is, in part, socially (politically) constructed is irrelevant. What Roberts does not understand is that these socially constructed groups (“white”, “black”) still, very much so, capture biological differences between them. That they are socialraces does not mean that they DO NOT have different physical features which correspond to geographic ancestry. The socialrace concept (which Roberts espouses in her book) is separate from Hardimon’s other scientific race concepts. But it is already inherent in Spencer’s, since his Blumenbachian partitions are social constructs of a biological reality. You don’t need genes to delineate races and minimalist races exist and are biologically real.
(I will cover other things from her book as I get to them. I will discuss race and medicine at length.)
Just-So Stories: The Slavery Hypertension Hypothesis
1800 words
Blacks have higher BP on average than whites. Why? One popular explanation is the Slavery Hypertension Hypothesis (SHH). The SHH is a hypothesis which posits 2 things: (1) that those living in the African climate were subject to limited water and salt, and dehydration so, a higher sodium-retention mechanism evolved in those populations to retain salt, which also leads to hypertension; and (2) during the Middle Passage there were high amounts of vomiting, diarrhea, heat, and little salt and so surviving slaves were “selected for” salt conserving water and salt. Then, when they reached the plantations, due to low water, copious sweating, and intense work, there were additional selective pressures which “selected for” water and salt conservation.
This hypothesis is so popular, that it was even pushed by Oprah, when Dr. Mehmet Oz asked Oprah why blacks have higher BP than whites. Lujan and DiCarlo (2018) write:
During a May 2007 Oprah show, Dr. Mehmet Oz asked Oprah, “Do you know why African-Americans have high blood pressure?” Oprah promptly replied that Africans who survived the slave trade’s Middle Passage “were those who could hold more salt in their bodies.” To which Dr. Oz exclaimed, “That’s perfect!” (64, 71). According to Dr. Oz and Oprah, African-Americans today are afflicted by hypertension at higher rate than whites because of genes passed on by their ancestors, genes that favored salt retention and that, in turn, cause high blood pressure (Fig. 1) (71). [They are implying that genetic ancestry is associated with BP; see below.]
Lujan and DiCarlo (2018) state that when individuals were “salt-loaded”, normal salt-resistant individuals retained just as much sodium in their bodies as salt-sensitive individuals. Salt-resistant individuals retain as much salt as salt-sensitive individuals—but they did not develop hypertension.
Furthermore, available evidence suggests that the difference in salt-sensitivity between African-Americans and Caucasians (European-Americans) is significantly smaller than what the Slavery Hypertension Hypothesis suggests. In fact, Chrysant and colleagues (14) were unable to find differences in the blood pressure response to salt by race, age, sex, or body weight. Thus salt sensitivity is not a racial problem, but rather a human problem, and the generalization that blacks are salt sensitive and whites are not should be discarded (14). It is important to note that measurements of salt retention in humans have come into serious question (50).
The hypothesis, as explained above, explains the data it purports to explain and only the data it purports to explain and is, therefore, a just-so story. Using the definition from Sterelny and Griffiths (1999: 61), a just-so story is “an adaptive scenario, a hypothesis about what a trait’s selective history might have been and hence what its function may be.”
So, the just-so story goes, that Africans in Africa—and those who survived the Middle Passage—had genes which favored better salt retention, and so, they were “selected for” which lead to an increased chance of survival in the low-salt, low-water, high-heat environment. The hypothesis is clearly ad hoc – notice that African-descended people have higher rates of blood pressure and then work backward. What in their recent or past history, could have lead to these high rates of hypertension in today’s societies.
This method is the usual EP reverse engineering method—strongly criticized by philosophers of science Robert Richardson (2007) and David Buller (2005)—which is “the inference from function to cause” (Richardson, 2007: 51). The just-so storytellers then work backward from a data point and “reason” how the trait became fixated in a particular population. So the formulators of the SHH wanted to infer function from cause—what the function of higher African BP was.
So the just-so story in question was formulated, which leads to genetic essentialist and determinist views—that genes are “causing” and were “selected for”—to explain the data they wanted to explain. But it makes no testable predictions, so it’s a just-so story. The hypothesis is inherently ad hoc—the “justification” for the hypothesis was reasoned backward from a fact we know today—that blacks have higher BP—and the “speculation” was provided as if it were true—which has permeated into the media, as can be seen above.
There are more sensible explanations for differences in hypertension between blacks and whites (I use those terms since they are socialraces). Genetic determinists would always go to the genes as an explanation for differences in any trait X. However, there is no reason to posit genetic differences between population groups as evidence for the differences in the causes of the trait in question. There are more sensible explanations for the BP disparity between blacks and whites.
Williams (1992) cites social factors as much more important than genetic factors in the etiology of hypertension – stress, social support, coping patterns, health behavior, sodium, calcium, and potassium consumption, alcohol consumption, and obesity. Citing these environmental factors that raise BP is critical—the human body’s physiology is adaptive and so, it can adapt to differing environments based on the reactions of the individual in that environment. This, of course, holds for nutrition as well. Nutrition most definitely affects BP – nutrition also affects rates of obesity (obviously). Blacks are more likely to be lower SES. Since blacks are more likely to be lower SES, they have higher rates of obesity which lead to higher rates of BP, too.
But one of the most important factors here is education. If people don’t know something, then they won’t do it. If they are taught ways to reduce symptom X, there is a higher chance of them reducing symptom X because they are better-armed with the knowledge against it. Knowing that all of these different environmental factors influence BP, then this points to a main culprit: education. Non, Gravlee, and Mulligan (2012) argue that it’s not ancestry that explains hypertension, but differences in education.
Non, Gravlee, and Mulligan (2012) analyzed both environmental and genetic factors which lead to hypertension. Thy found that in the black sample, systolic BP and mean arterial pressure (MAP) were higher among those who had a HS diploma or lower, but found no differences by education in the white sample.
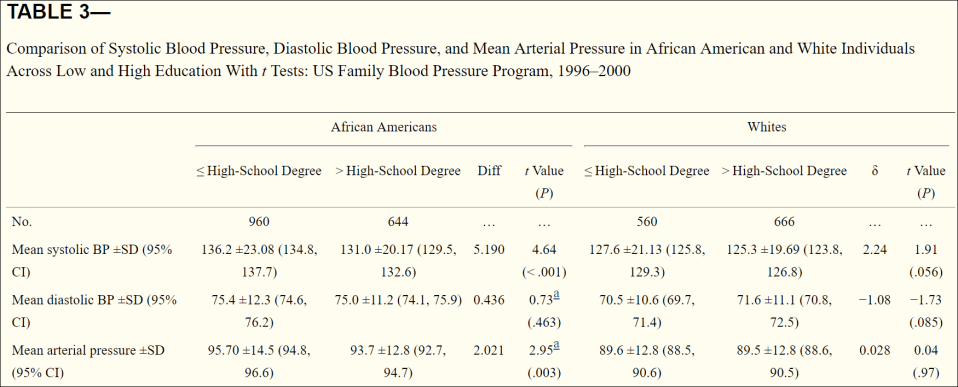
So black men were predicted to have a higher SBP, then white men, black women and finally white women, across all levels of education. SBP declined most sharply in black men and women compared to white men and women.
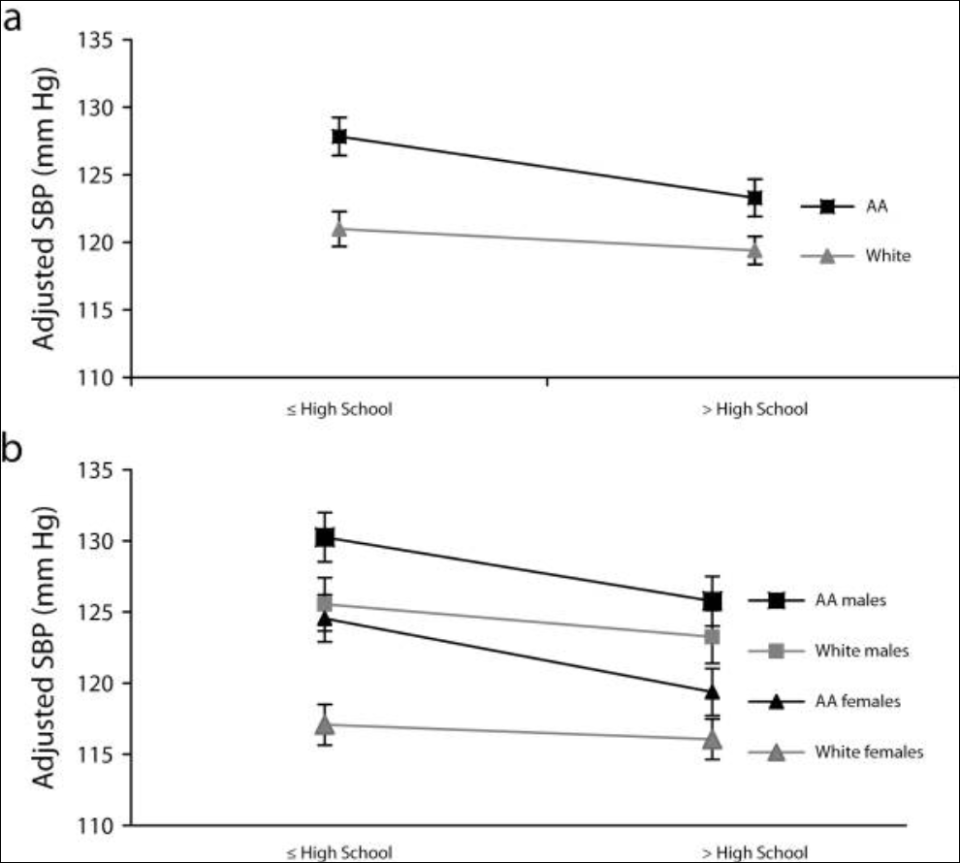
Genetic ancestry was not associated with BP among black Americans, but there was a significant association between education and BP. Education is, of course, not too good a measure of the social environment. Even using this measure, significant reductions in BP were found. Genetic ancestry is supposed to be associated BP in virtue of the ancestral environment of black Americans, along with supposed selection pressures which occurred on the Middle Passage. So if genetic ancestry isn’t associated, then the hypothesis is discarded.
Non, Gravlee, and Mulligan’s (2012) results support the “minority poverty hypothesis” because “the worst blood pressures were predicted for people who faced the double burden of being less educated and identifying as African American.” The minority poverty hypothesis is “The idea is that black people who live in poverty are uniquely disadvantaged in attaining good health because of the combination of poverty and race” (Hall, Humphreys, and Ruseski, 2015: 5).
Because genetic ancestry was estimated from only 294 loci, and a large set of populations across Africa, which may not be best for representing the West African ancestry of black Americans (Note how this is the population in question in the theory we are discussing). So an analysis focusing just on West African populations may change the relationship. Education was their only measure of the social environment, but other measures of the social environment, like “residential segregation, psychosocial stress, and everyday discrimination” may fully account for higher levels of BP in black Americans. Of course, further there needs to be further study to see whether it is the education per se that causes the differences in BP or if education serves only as a marker for other aspects of the social environment.
The evidence that education accounts for a lot of the variation in differences in BP between blacks and whites is strong. If it is other aspects of the social environment, and not education per se, then there is something in that environment that does not elicit the physiological response that leads to higher BP. We can also, of course, liken this to the Mazur’s (2016) honor culture hypothesis—the hypothesis “that young men’s participation in the honor culture of poor black neighborhoods has the effect of elevating T.” This is due to the adaptiveness of our physiological systems and how it adapts to the environment based on environmental cues.
There was one recent study where they found that “Among black male barbershop patrons with uncontrolled hypertension, health promotion by barbers resulted in larger blood-pressure reduction when coupled with medication management in barbershops by specialty-trained pharmacists” (Victor et al, 2018). This, of course, makes sense. If one is made aware of anything wrong with them, then they will be more likely to seek help for their ailments.
Victor et al (2018) write:
Because black men with hypertension often have multiple cardiovascular risk factors,37 marked reductions in blood pressure — if sustained with the use of our approach and then initiated more widely — might reduce the high rates of hypertension-related disability and death among black men with hypertension in the United States.11
Since three out of four black men have high blood pressure by the time they are 55, then if this can and does hold for the long-term, then this would help many individuals.
Seventy-eight barber shops enrolled in the program. The n was 319 men who had a SBP of 140 mm or higher from 52 black-owned barber shops. The intervention increased doctor visits and anti-hypertensive medications (which I disagree with). Pharmacists were placed in the shop and checked the BP of black men who entered (barbers were also trained to measure BP). Reductions of 21.6 and 14.9 SBP and DBP respectively were seen. 63 percent of those who participated achieved a normal BP whereas 12 percent of those in the control group did so. This, clearly, is another way in which education can lower BP in this population.

This is a great idea—and if further study confirms that this works, it should begin to be implemented elsewhere. The most important factor is outreach—getting the information to people and teaching them how to reduce it on their own through lifestyle modifications. And since outreach is related to educating people on a certain topic, then this, too, falls under the—somewhat large—umbrella of “education.”
In sum, the SHH is a just-so story and doesn’t explain why blacks have higher rates of BP than whites. Genetic ancestry seems to not explain hypertension rates between blacks and whites. Social environment changes and outreach can lower BP disparities between populations. If one understands the intricacies of physiology, then they would understand the physiological responses to different environmental/social stimuli.
Muscle Fibers, Obesity, Cardiometabolic Disorders, and Race
2650 words
The association between muscle fiber typing obesity and race is striking. It is well-established that blacks have a higher proportion of type II skeletal muscle fibers than whites and these higher proportions of these specific types of muscle fibers lead to physiological differences between the two races which then lead to differing health outcomes between them—along with differences in athletic competition. Racial differences in health are no doubt complex, but there are certain differences between the races that we can look at and say that there is a relationship here that warrants further scrutiny.
Why is there an association between negative health outcomes and muscle phsyiology? The answer is very simple if one knows the basics of muscle physiology and how and why muscles contract (it is worth noting that out of a slew of anatomic and phsyiologic factors, movement is the only thing we can consciously control, compare to menstration and other similar physiologic processes which are beyond our control). In this article, I will describe what muscles do, how they are controlled, muscle physiology, the differences in fiber typing between the races and what it means for health outcomes between them.
Muscle anatomy and physiology
Muscle fiber number is determined by the second trimester. Bell (1980) noted that skeletal muscle fiber in 6 year olds is not different from normal adult tissue, and so, we can say that between the time in the womb and age 6, muscle fiber type is set and cannot be changed (though training can change how certain fibers respond, see below).
Muscle anatomy and physiology is interesting because it shows us how and why we move the way we do. Tendons attach muscle to bone. Attached to the tendon is the muscle belly. The muscle belly is made up of facsicles and the fascicles are made up of muscle fibers. Muscle fibers are made up of myofibrils and myofibrils are made up of myofilaments. Finally, myofilaments are made up of proteins—specifically actin and myosin, this is what makes up our muscles.

(Image from here.)
Muscle fibers are encased by sarcolemma which contains cell components such as sarcoplasm, nuclei, and mitochondria. They also have other cells called myofibrils which contain myofilaments which are then made up of actin (thin filaments) and mysoin (thick filaments). These two types of filaments form numerous repeating sections within a myofibril and each repeating section is known as a sarcomere. Sarcomeres are the “functional” unit of the muscle, like the neuron is for the nervous system. Each ‘z-line’ denotes another sarcomere across a myofibril (Franzini-Armstrong, 1973; Luther, 2009).
Other than actin and myosin, there are two more proteins important for muscle contraction: tropomyosin and troponin. Tropomyosin is found on the actin filament and it blocks myosin binding sites which are located on the actin filament, and so it keeps myosin from attaching to muscle while it is in a relaxed state. On the other hand, troponin is also located on the actin filament but troponin’s job is to provide binding sites for calcium and tropomyosin when a muscle needs to contract.
So the structure of skeletal muscle can be broken down like so: epymyseum > muscle belly > perimyseum > fascicle > endomyseum > muscle fibers > myofibrils > myofilaments > myosin and actin. Note diagram (C) from above; the sarcomere is the smallest contractile unit in the myofibril. According to sliding filament theory (see Cook, 2004 for a review), a sarcomere shortens as a result of the ‘z-lines’ moving closer together. The reason these ‘z-lines’ converge is because myosin heads attach to the actin filament which asynchronistically pulls the actin filament across the myosin, which then results in the shortening of the muscle fiber. Sarcomeres are the basic unit controlling changes in muscle length, so the faster or slower they fire depends on the majority type of fiber in that specific area.
But the skeletal muscle will not contract unless the skeletal muscles are stimulated. The nervous system and the muscular system communicate, which is called neural activiation—defined as the contraction of muscle generated by neural stimulation. We have what are called “motor neurons”—neurons located in the CNS (central nervous system) which can send impulses to muscles to move them. This is done through a special synapse called the neuromuscular junction. A motor neuron that connects with muscle fibers is called a motor unit and the point where the muscle fiber and motor unit meet is callled the neuromuscular junction. It is a small gap between the nerve and muscle fiber called a synapse. Action potentials (electrical impulses) are sent down the axon of the motor neuron from the CNS and when the action potential reaches the end of the axon, hormones called neurotransmitters are then released. Neurotransmitters transport the electrical signal from the nerve to the muscle.
Muscle fiber types
The two main categories of muscle fiber are type I and type II—‘slow’ and ‘fast’ twitch, respectively. Type I fibers contain more blood cappilaries, higher levels of mitochondria (which transforms food into ATP) and myoglobin which allows for an improved delivery of oxygen. Since myoglobin is similar to hemoglobin (the red pigment which is found in red blood cells), type I fibers are also known as ‘red fibers.’ Type I fibers are also smaller in diameter and slower to produce maximal tension, but are also the most fatigue-resistant type of fiber.
Type II fibers have two subdivisions—IIa and IIx—based on their mechanical and chemical properties. Type II fibers are in many ways the opposite of type I fibers—they contain far fewer blood cappilaries, mitochondria and myoglobin. Since they have less myoglobin, they are not red, but white, which is why they are known as ‘white fibers.’ IIx fibers have a lower oxidative capacity and thusly tire out quicker. IIa, on the other hand, have a higher oxidative capacity and fatigue slower than IIx fibers (Herbison, Jaweed, and Ditunno, 1982; Tellis et al, 2012). IIa fibers are also known as intermediate fast twitch fibers since they can use both anarobic and aerobic metabolism equally to produce energy. So IIx fibers are a combo of I and II fibers. Type II fibers are bigger, quicker to produce maximal tension, and tire out quicker.
Now, when it comes to fiber typing between the races, blacks have a higher proportion of type II fibers compared to whites who have a higher proportion of type I fibers (Ama et al, 1986; Ceaser and Hunter, 2015; see Entine, 2000 and Epstein, 2014 for reviews). Higher proportions of type I fibers are associated with lower chance of cardiovascular events, whereas type II fibers are associated with a higher risk. Thus, “Skeletal muscle fibre composition may be a mediator of the protective effects of exercise against cardiovascular disease” (Andersen et al, 2015).
Now that the basics of muscle anatomy and physiology are apparent, hopefully the hows and whys of muscle contraction and what different muscle fibers do are becoming clear, because these different fibers are distributed between the races in uneven frequencies, which then leads to differences in sporting performance but also differents in health outcomes.
Muscle fibers and health outcomes
We now know the physiology and anatomy of muscle and muscle fiber typing. We also know the differences between each type of skeletal muscle fiber. Since the two races do indeed differ in the percentage of skeletal muscle fiber possessed on average, we then should find stark differences in health outcomes, part of the reason being these differences in muscle fiber typing.
While blacks on average have a higher proportion of type II muscle fibers, whites have a higher proportion of type I muscle fibers. Noting what I wrote above about the differences between the fiber types, and knowing what we know about racial differences in disease outcomes, we can draw some inferences on how differences in muscle fiber typing between races/individuals can then affect disease seriousness/acquisition.
In their review of black-white differences in muscle fiber typing, Ceaser and Hunter (2015) write that “The longitudinal data regarding the rise in obesity indicates obesity rates have been highest among non-Hispanic Black women and Hispanic women.” And so, knowing what we know about fiber type differences between races and how these fibers act when they fire, we can see how muscle fiber typing would contribute to differences in disease acquisition between groups.
Tanner et al (2001) studied 53 women (n=28, lean women; and n=25, obese women) who were undergoing an elective abdominal surgery (either a hysterectomy or gastric bypass). Their physiologic/anatomic measures were taken and they were divided into races: blacks and whites, along with their obesity status. Tanner et al found that the lean subjects had a higher proportion of type I fibers and a lower proportion of type IIx fibers whereas those who were obese were more likely to have a higher proportion of type IIb muscle fibers.
Like other analyses on this matter, Tanner et al (2001) showed that the black subjects had a higher proportion of type II fibers in comparison to whites who had a higher proportion of type I fibers (adiposity was not taken into account). Fifty-one percent of the fiber typing from whites was type I whereas for blacks it was 43.7 pervent. Blacks had a higher proportion of type IIx fibers than whites (16.3 percent for whites and 23.4 for blacks). Lean blacks and lean whites, though, had a similar percentage of type IIx fibers (13.8 percent for whites and 15 percent for blacks). It is interesting to note that there was no difference in type I fibers between lean whites and blacks (55.1 percent for whites and 54.1 percent for blacks), though muscle fibers from obese blacks contained far fewer type I fibers compared to their white counterparts (48.6 percent for whites and 34.5 for blacks). Obese blacks’ muscle fiber had a higher proportion of type IIx fibers than obese whites’ fiber typing (19.2 percent for whites and 31 percent for blacks). Lean blacks and lean whites had a higher proportion of type I fibers than obese blacks and obese whites. Obese whites and obese blacks had more type IIx fibers than lean whites and lean blacks.
So, since type II fibers are insulin resistant (Jensen et al, 2007), then they should be related to glucose intloerance—type II diabetes—and blacks with ancestry from West Africa should be most affected. Fung (2016, 2018) shows that obesity is a disease of insulin resistance, and so, we can bring that same rationale to racial differences in obesity. Indeed, Nielsen and Christensen (2011) hypothesize that the higher prevalence of glucose intolerance in blacks is related to their lower percentage of type I fibers and their higher percentage of type II fibers.
Nielsen and Christensen (2011) hypothesize that since blacks have a lower percentage of type I fibers (the oxidative type), this explains the lower fat oxidation along with lower resting metabolic rate, sleeping metabolic rate, resting energy expenditure and Vo2 max in comparison to whites. Since type I fibers are more oxidative over the glycolitic type II fibers, the lower oxidative capacity in these fibers “may cause a higher fat storage at lower levels of energy intake than in individuals with a higher oxidative capacity” (Nielsen and Christensen, 2011: 611). Though the ratio of IIx and IIa fibers are extremely plastic and affected by lifestyle, Nielsen and Christensen do note that individuals with different fiber typings had similar oxidative capacity if they engaged in physical activity. Recall back to Caesar and Hunter (2015) who note that blacks have a lower maximal aerobic capacity and higher proportion of type II fibers. They note that lack of physical activity exacerbates the negative effects that a majority type II fibers has over majority type I. And so, some of these differences can be ameliorated between these two racial groups.
The point is, individuals/groups with a higher percentage of type II fibers who do not engage in physical activity have an even higher risk of lower oxidative capacity. Furthermore, a higher proportion of type II fibers implies a higher percentage of IIx fibers, “which are the least oxidative fibres and are positively associated with T2D and obesity” (Nielsen and Christensen, 2011: 612). They also note that this may explain the rural-urban difference in diabetes prevalance, with urban populations having a higher proportion of type II diabetics. They also note that this may explain the difference in type II diabetes in US blacks and West African natives—but the reverse is true for West Africans in the US. There is a higher rate of modernization and, with that, a higher chance to be less physically active and if the individual in question is less physically active and has a higher proportion of type II fibers then they will have a higher chance of acquiring metabolic diseases (obesity is also a metabolic disease). Since whites have a higher proportion of type I fibers, they can increase their fat intake—and with it, their fat oxidation—but this does not hold for blacks who “may not adjust well to changes in fat intake” (Nielsen and Christensen, 2011: 612).
Nielsen and Christensen end their paper writing:
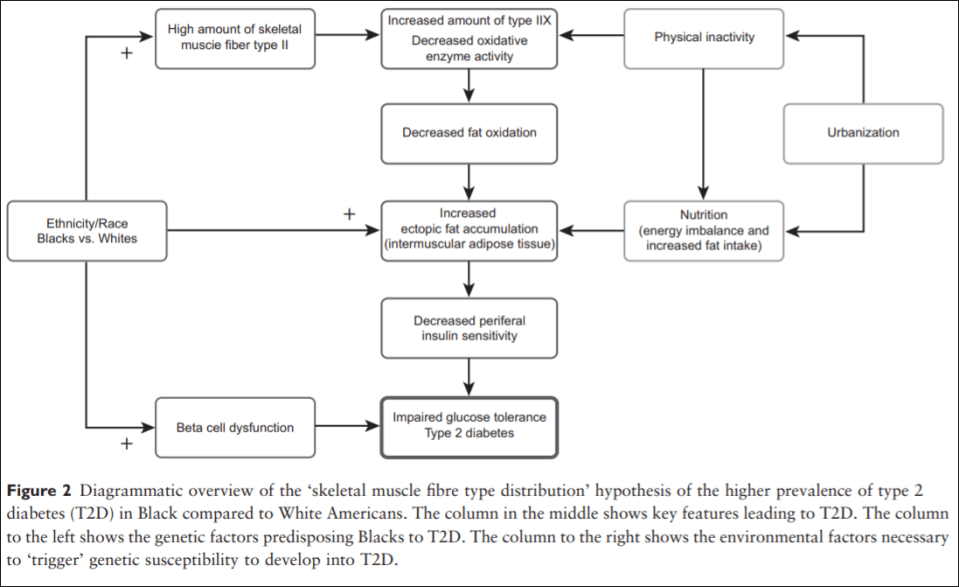
Thus, Blacks of West African ancestry might be genetically predisposed to T2D because of an inherited lower amount of skeletal muscle fibre type I, whereby the oxidative capacity and fat oxidation is reduced, causing increased muscular tissue fat accumulation. This might induce skeletal muscle insulin resistance followed by an induced stress on the insulin-producing beta cells. Together with higher beta-cell dysfunction in the West African Diaspora compared to Whites, this will eventually lead to T2D (an overview of the ‘skeletal muscle distribution hypothesis’ can be seen in Figure 2).
Lambernd et al (2012) show that muscle contractions eliminated insuin resistance by blocking pro-inflammatory signalling pathways: this is the mechanism by which physical activity decreases glucose intolerance and thusly improves health outcomes—especially for those with a higher proportion of type II fibers. Thus, it is important for individuals with type II fibers to exercise, since sedentariness is associated with an age-related insulin resistance due to impaired GLUT4 utilization (Bunprajun et al, 2013).
(Also see Morrison and Cooper’s (2006) hypothesis that “reduced oxygen-carrying capacity induced a shift to more explosive muscle properties” (Epstein, 2014: 179). Epstein notes that the only science there is on this hypothesis is one mouse and rat study showing that low hemoglobin can “induce a switch to more explosive muscle fibers” (Epstein, 2014: 178), but this has not been tested on humans to see if it would hold. If this is tested on humans and if it does hold, then that would lend credence to Morrison’s and Cooper’s (2006) hypothesis.)
Conclusion
Knowing what we know about muscle anatomy and physiology and how muscles act we can understand the influence the different muscle types have on disease and how they contribute to disease variation between race, sex and the individual level. Especially knowing how type II fibers act when the individual in question is insulin resistant is extremely important—though it has been noted that individuals who participate in aerobic exercise decrease their risk for cardiometabolic diseases and can change the fiber distribution difference between IIx and IIa fibers, lowering their risk for acquiring cardiometabolic diseases (Ceaser and Hunter, 2015).
Thinking back to sarcomeres (the smallest contractile unit in the muscle) and how they would act in type II fibers: they would obviously contract much faster in type II muscles over type I muscles; they would then obviously tear faster than type I muscles; since type II muscles are more likely to be insulin resistant, then those with a higher proportion of type II fibers need to focus more on aerobic activity to “balance out” type IIx and IIa fibers and decrease the risk of cardiometabolic disease due to more muscle contractions (Lambernd et al, 2012). Since blacks have a higher proportion of type II fibers and are more likely to be sedentary than whites, and since those who have a higher proportion of type II fibers are more likely to be obese, then it is clear that exercise can and will ameliorate some of the disparity in cardiometabolic diseases between blacks and whites.
Why Biological Racial Realism is True
1650 words
To be a realist about race is to hold that racial categories pick out real kinds in nature. (Smith, 2015: 43; Nature, Human Nature, and Human Difference Race in Early Modern Philosophy)
Claims from biological racial realists are simple: Racial categories pick out real kinds in nature. If racial categories pick out real kinds in nature, then race surely exists.
Racial groups—or groups taken to be racial groups—characterize three conditions from Hardimon (2017): C1: they are distinguished from other groups by patterns of visible physical features; C2: the members are linked by common ancestry which is peculiar to that group; and C3: they derive from a distinct geographic location.
Justifying C1 is simple: groups taken to be ‘racial’ are distinguished from other groups on the basis of physical characters. Someone from Europe looks different than someone from Africa; someone from Africa looks different than someone from Asia; someone from Asia looks different than someone from the Pacific Islands; someone from the Pacific Islands looks different than the Natives of America. Groups taken to be ‘racial’ have different facial features; they have different morphology. Thus, since there are heritable differences between groups taken to be ‘racial’, then this is evidence that race does indeed exist.
It’s important to also discuss what C1 does not demand: it does not demand that racial groups be distinguished by each of their visible physical features; it does not demand that each visible physical features of members of a race be identical; it allows skin color to vary just as much within race as it does between race; finally, it also allows great variation in hair color, skull morphology and skin color. Thus, since Hardimon’s concept is ‘vague’, then one might be able to say that it is “clinal” (that is, these differences vary by geography). But “Physical anthropologist Frank Livingstone’s well-known adage “There are no races, only clines” overlooks the possibility that, logically speaking, races might be clines” (Hardimon, 2017: 38). The claim “There are no races, only clines” is one that is oft-repeated against the reality of biological races.
C2, very simply, shows that differences in visible physical features are not the only things that delineate race: race is also defined in terms of ancestry and is therefore essential to the concept of race (I’d argue that ancestry is essential to any argument that attempts to establish races as biologically real). Races are, clearly, morphologically demarcated ancestry groups. The justification for C2 is thus: it is intuitive. Examples of race articulated in the past also bore this very basic concept: Linneus’ europeaus, asiaticus, afer, and americanus; Blumenbach’s Mongoloid, Caucasoid, Ethiopian, Malay and American; UNESCO’s Negroid, Mongoloid, and Caucasoid (deployed most famously by JP Rushton); and the Office of Management and Budget’s American Indian (or Alaskan native), black, Asian, whites, native Hawaiians (Pacific Islanders) (see Spencer, 2014 for a treatment of the OMB’s views on race and his ‘radical solution to the race problem’).
Now, finally, C3: the condition that groups taken to be ‘racial’ must derive from a distinct geographic location. Race, and the names used to refer to race, and so “The use of typonyms in the naming of racial groups suggests that the thinkers who chose these names were thinking of race as a geographical grouping” (Hardimon, 2017: 50). So, C1 and C2 have been established. This leaves us with C3. Races differ in patterns of visible physical features; these differences are explained by differences in geographic location. If race R1 derives from geographic location G1, and G1 is distinct from G2 which race R2 inhabits, then races R1 and R2 will look physically different.
Thus the groups that we think of when we think about race are groups that genetically transmit heritable characters to their offspring which then correspond to differences in geographic ancestry. So groups that satisfy C1-C3 are ‘races’, in the normal sense of the word. Groups that satisfy C1-C3 are articulated in Hardimon’s (2017) populationist race concept using Rosenberg et al’s (2002) data, and these are, largely, the same groups that Blumenbach pointed out centuries ago (Spencer, 2014).
The Visible Physical Features of Minimalist Race Are Racial
The visible physical features of minimalist race that correspond to geographical ancestry count as “racial” because they are defining features of minimalist races. They no more need to be correlated with normatively important features to be properly counted as racial then minimalist races need to be characterized by normatively important features to be properly counted as races. Just as the concept of minimalist race deflates the concept of RACE, so too it deflates the concept of RACIAL. Visible physical features that correspond to geographical ancestry are eo ipso racial. (Hardimon, 2017: 52)
Hardimon (2017: 99) also articulates one of the best definitions of race I have come across:
A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.
Since Hardimon’s views are new (published in 2017), there are no replies to his argument—excpet one, by Spencer (2018). Spencer doesn’t take to two of Hardimon’s claims: that (1) that the minimalist concept of race is the ordinary concept of race and (2) that minimalist races are biologically real. He grants (1) until Hardimon provides evidence that the minimalist concept of race is the ordinary concept of race. (2), on the other hand, Spencer attacks.
His objection to (2) comes down to the simple fact that 13/17 of the different conceptions of racial groups discussed by Linnaeus, Blumenbach, the OMB, and UNESCO do not fit C1-C3 (Blumenbach’s races do fit C1-C3). Spencer (2018) states that “there’s no ancestor that Eurasians share that’s not also shared by East Asians, Oceanians, and Native Americans […] there’s no ancestor that East Asians share that’s not also shared by Oceanians and Native Americans.” (See Duda and Zrzavy, 2016.) We need to be clear on what Hardimon means by “ancestry.” The dictionary definition of “ancestry” is thus: “one’s family or ethnic descent.” On this definition of “ancestry”, groups taken to be races do have “distinct ancestry“, so defined, and so, Hardimon’s (2017) C2 does indeed hold.
Biological racial realism “should” mean “race is a geniuine kind in biology.” Take the argument from Spencer (2011: 24):
(1) The meaning of ‘biological racial realism’ in the race debate should be a metaphysically minimal interpretation of important scientific kindhood that also does the most justice to what counts as an important scientific kind.
(2) A “metaphysically minimal” interpretation of important scientific kindhood is one that does not adopt unnecessary and contentious metaphysical assumptions.
(3) The interpretation of important scientific kindhood that does the most justice to what counts as an important scientific kind is the one that best captures epistemically important scientific kinds—or ‘EIS kinds’ for short.
(4) The candidates for important scientific kindhood in the race debate are natural kinds, naturali kinds, naturalu kinds, naturalp kinds, realp biological kinds, reali biological kinds, and geniuine kinds.
(5) No kind of kind in the race debate is both metaphysically minimal and does a better job of capturing EIS kindhood than genuine kinds.
(6) Therefore, the meaning of ‘biological racial realism’ in the race debate should be ‘race is a genuine kind in biology’.
Spencer has good critiques of Hardimon’s minimalist/populationist race view, but it does not hold.
Even if we allow Spencer’s views on Hardimon’s arguments for the existence of race to hold, Spencer himself has articulated a sound argument for the existence of race. In his 2014 paper A Radical Solution to the Race Problem, Spencer (2014) shows that Americans defer to the US Census on matters of race; the US Census defers to the OMB; the OMB refers to “sets of” populations—blacks, whites, Asians, Native Americans and Pacific Islanders; these “sets of” populations are not kinds (like what Hardimon argues his racial classifications are); therefore, races are not ‘kinds’, the term ‘race’ refers to sets of population groups. Thus, according to Spencer (2014), race refers to “proper names” for population groups, not “kinds”.
Both of Hardimon’s and Spencer’s arguments show that race is a biological reality; they both show that biological racial realism is true. Their concepts pick out real kinds in nature (Smith, 2016: 43).
Philosopher Justin Smith, in his book 2015 book Nature, Human Nature, and Human Difference Race in Early Modern Philosophy articulates a Hardimonian argument for the existence our habits of distinguishing between human populations:
Now if scientific taxonomy builds on folk taxonomy, and if racial classification builds on this in turn, there might be some basis for supposing that something about the modern habit of distinguishing between human groups on racial grounds is more deep-seated than we have acknowledged it to be. (Smith, 2015: 47)
The reason why there “might be some basis for supposing that something about the modern habit [which is not truly modern; Sarich and Miele, 2004; which I am sure that Smith knows due to the content of his book] of distinguishing between human groups on racial grounds is more deep-seated” than we have acknowledged because we are picking out real kinds that exist in nature.
Conclusion
Race, as a concept, is biologically real. Racial categories pick out real kinds in nature, as argued by Spencer (2014) and Hardimon (2017). Criticisms on Hardimon from Spencer or Spencer from Hardimon do not take away from this one fact: that race exists and is biologically real. Groups taken to be ‘racial’ look different from each other; they look different from each other due to their geographic locations and their ancestry (C1-C3). Since this is true, then race exists. We can argue this view simply:
P1. If groups of people look different from each other depending on where their ancestors evolved, then race exists.
P2. Groups of people look different from each other depending on where their ancestors evolved.
C. Therefore, race exists since people look different depending on where their ancestors evolved (modus ponens, P1, P2).
Biological racial realism is true.
Arguments For and Against Preimplantation Genetic Diagnosis
1500 words
Like abortion, preimplantation genetic diagnosis (PGD) is feared. This is due, in part, to fears of eugenics coming back through a “backdoor” with the advent of new technology such as CRISPR/CAS9 and other types of tools we can use to genetically modify ourselves. The case of PGD—just like abortion—has been heavily debated in recent times, more so due to the recent strides in genomics we have made since the advent of the Human Genome Project.
PGD offers us a method to identify embryos with genetic diseases. Understandably, this has raised caution with some, due to the strong link with eugenic thinking/policies. See The Ethical Implications of Preimplantation Genetic Diagnosis. Thus, by scanning the genomes of fetuses, we can then see if they have a higher chance of acquiring any disease and select fetuses which have a lower to nonexistent chance of acquiring said disease.
An argument against PGD
In his paper Just diagnosis? Preimplantation genetic diagnosis and injustices to disabled people, Peterson (2005) presents one slippery slope argument against PGD (Freeman, 1996) (and later provides a refutation). The argument that Peterson (2005) presents is a “slippery slope” argument—that is, it’s an argument which argues that if we allow X, then since we allowed X, then horrible thing Y can and will follow. Peterson (2005) articulates the argument thus:
As situation A (the use of PGD to select against severe genetic diseases) is refined, “it will be difficult, if not impossible, to contain the uses of such research”. A will therefore bring about situation B, where PGD will be used to select against mild or perhaps non-medical conditions.
Besides the refinement of A, B will be brought about because “There will likely be an increasing pressure … on people to take advantage of these techniques, and not bring even a mildly disabled child into the world …”.
Finally, we could reach a morally abhorrent outcome Z, which is disturbingly close to eugenics, where our notion of the moral equality of all human beings, including those with disabilities, is undermined.
Z is so morally bad, that it outweighs the benefits of undertaking A.
Therefore, A should not be undertaken.
This argument, in my view, seems to be appealing to emotion by saying that since we can reach morally abhorrent outcome Z (a type of eugenics), then we should not continue with this practice. However, others argue that this discriminates against people with disabilities (see Katthab, 2009). Peterson (2005) argues that Freeman’s (1996) argument “lacks empirical support” and so it makes the conclusion difficult to assess; technology can and will be regulated which would quell any fears of possible use of this technology for any eugenic ideals; and, through using PGD, we can use it to “fight the obvious causes of discrimination, such as intolerance and egoism“, which would, in turn, reduce discrimination. Lastly, addressing Freeman’s (1996) concerns that PGD would lead to the discrimination of currently disabled persons, Peterson (2005) claims that “even if we accept that PGD will generate discrimination against disabled people, it is far from obvious that this is sufficient to warrant its moral condemnation.” Thus, Peterson (2005) concludes that Freeman’s (1996) argument is not sufficient to end the use of PGD technology. (Also see Robertson, 2003 for the view that “except for sex selection of the first child, most current extensions of PGD are ethically acceptable“.)
Many arguments against PGD rely on the concept of a fetus as a person and terminating any fetus with any prospective disease is paramount to killing a person. Others, of course, hinge on the fact that PGD does help reduce the risk of a babe being born with deleterious diseases, it does not completely ameliorate any generic risk for disease and so the fetus must be monitored through conception up until pregnancy to be sure that no disease appears during conception. And, of course, certain diseases that may hamper one’s quality of life may not appear until one reaches adolescence, adulthood, middle or old age. This is another fact against PGD: that even selecting embryos that apparently have no risk for disease, they may acquire diseases in older age which would not be seen since some diseases only generate symptoms at certain stages of life.
One final objection to PGD is also moral: it could, and will, send a message to any individuals currently alive that their lives are somehow “less” than others, since individuals with a chance to acquire said disease are selected against, as McConachy (2010) argues.
Lastly, Richardson (2017: 155-157) argues that the selection of embryos with so-called “potential” is ill-founded since they talk about vague concepts such as “egg quality”. Differences in potential lie, supposedly, in the “genetic blueprint” (see my article DNA Is not a Blueprint for arguments against that notion), though “That view implies that differences in individuals in important functions are largely due to differences in genes. As we have seen, though, things are far from being so simple” (Richardson, 2017: 156).
An argument for PGD
PGD can be used for many things; most importantly, screening the genome of a perspective fetus before IVF. Though, this has led some to worry that this could be a way in which eugenics can “sneak in through the backdoor” by virtue of making people with diseases more likely to be discriminated against since “disabled phenotypes” would slowly be phased out by PGD. One argument for could be:
Parents have rights; if parents have rights, then they have the right to do what they want with their children, and they want to do what is best for their children; therefore a parent should have the right to use PGD to select the best-possible embryo for implantation.
This is where we think of the implications of aborting a fetus, or not implanting a fetus that has a higher chance of acquiring any disease. There are, of course, certain people who would willingly select embryos which have a high chance to be disabled, because they themselves are disabled or they believe they “should be” disabled themselves and so want disabled children. Since parents have rights, as can be seen in the reasoning chain above, then parents should be able to choose the status of their babe. But if the babe’s quality of life is low, then is it ethical for that person to select an embryo with a high chance of acquiring a disease?
Another argument “for” PGD can be:
Humans should not suffer; if we can prevent human suffering with our current technology, then we have a moral imperative to do whatever is in our power to do so; if we can prevent low qualities of life for any embryo E, then we should do so; therefore, we should screen embryos for diseases that can and will lower their qualities of life and select against these embryos.
One may argue that a fetus may not have a “moral right” to life (see Tooley, 1972), though, if we know that a fetus has a high chance to have such a debilitating disease such that it lowers its quality of life, then it should be aborted/not implanted in the womb. Religious views, of course, come into play here but I am not worried about them; I am worried only about sound arguments for them. (See Fasoulioutis and Schenker, 1997 for these views.)
PGD may, of course, prevent abortions of said fetus since we know that the fetus in question may have a higher chance of acquiring a certain disease, so if one is against abortion, then they may be for the use of PGD to screen the fetuses’ genome to scan for any readily apparent problems in their genome in regard to the acquiring of certain genetic diseases.
Arguments for PGD hinge on parents wanting the best possible lives for the children they conceive and the arguments really rely on parental autonomy, the parent’s want to choose how their kid is born, if their chance for disease is high or not (which also would turn to “designer babies”; an argument against “designer babies” will be erected soon. If parents can do what’s best for an unborn child then, most would argue they have a moral imperative to give the babe the best possible life and so they should abort/select against certain embryos which have a high chance of acquiring diseases.
Conclusion
There do not seem to be as many strong arguments against PGD compared to abortion. Though one can use the basic blueprint of the argument against abortion and liken it to PGD. The PGD debate is similar to the abortion debate. One can use similar arguments against abortion to argue against PGD.
These debates are both ethical scientific: we have the ability to now do X which would stop suffering Y in embryo E. Just because we can do something, does that mean we should do so? Like with the editing of the germline, we don’t know what types of consequences would occur since we have, pretty much, no experience in editing the germline/genes of humans in a large-scale way.
An Argument For and Against Abortion
1350 words
Abortion is a touchy subject for many people. There are many different arguments for and against abortion, including, but not limited to, the woman’s right to do what she wants with her body on the pro-abortion side, to the right of a fetus to live a good life if there is little chance of the fetus developing a serious disease. In this article, I will provide two arguments: one for and one against abortion. The abortion debate is an ethical, not scientific, one, and so, we must use argumentation to see the best way to move forward in this debate.
An argument for abortion
Michael Tooley, in his paper Abortion and Infanticide, provides an argument not only for the abortion of fetuses, but the killing of infants and animals since they cannot conceive of continuing their selves. He argues that an organism only has a right o life of they can conceive of that right to life. His conclusion is that it should be morally permissible to end a baby’s life shortly after birth since it cannot conceive of wanting to live. The conclusion of the argument also includes—quite controversially, in fact—young infants and (nonhuman) animals. Ben Saunders articulates Tooley’s argument in Just the Arguments: 100 of the Most Important Arguments in Western Philosophy (2011: 284-286):
P1. If A has a morally serious right to X, then A must be able to want X.
P2. If A is able to want X, then A must be able to conceive if X.
C1. If A has a morally serious right to X, then A must be able to conceive of X (hypothetical syllogism, P1, P2).
P3. Fetuses, young infants, and animals cannot conceive of their continuing as subjects of mental states.
C2. Fetuses, young infants, and animals cannot want their continuance as subjects of mental states (modus tollens, P2, P3).
C3. Fetuses, young infants, and animals do not have morally serious rights to continue as subjects of mental states (modus tollens, P1, C2).
P4. If something does not have a morally serious right to life, then it is not morally wrong to kill it painlessly.
C4. It is not wrong to kill fetuses, young infants, or animals painlessly (modus ponens, C3, P4).
Of course, most people would seriously disagree with C4, since a babe’s life is one of the most precious things in the world— the protection of said babes is how we continue our species. However, the argument is deductively valid, and so one must show which premise is wrong and why. This argument—along with the one that will be presented below against abortion (of healthy fetuses)—is very strong. Thus, if a woman so pleases (along with her autonomy), she can choose to abort her fetus since it is not wrong to kill a fetus painlessly. (I am not aware if fetuses can feel pain or not, however. If they can, then the conclusion of this argument does not hold.)
Tooley’s argument regarding the killing of infants is similar to an argument made by Gibiulini and Minerva (2013) who argue that since fetuses and newborns don’t have the same moral status as actual persons, fetuses and infants can eventually become persons, and since adoption is not always in the best interests 9f people, then “‘after-birth abortion’ (killing a newborn) should be permissible in all the cases where abortion is, including cases where the newborn is not disabled” (Giubilini and Minerva, 2013).
An argument against abortion
One strong argument against abortion exists: Marquis’ (1989) argument in his paper Why Abortion Is Immoral. Women may want an abortion for many reasons: such as not wanting to carry a babe to term, to finding out that the babe has a serious genetic disorder. Though, what matters to this argument is not the latter, but the former: the mother wanting an abortion of a healthy fetus. Marquis’ argument is simple: killing is wrong; killing is wrong since killing ends one’s life, and ending one’s life means they won’t experience anything anymore, they won’t be happy anymore, they won’t be able to accomplish things, and this is one of the greatest losses that can be suffered; abortions of a healthy fetus cause the loss of experiences, activities, and enjoyment to the fetus; thus, the abortion of a healthy fetus is not only ethically wrong, but seriously wrong. Marquis’ (1989) argument is put succinctly by Leslie Burkholder in the book Just the Arguments: 100 of the Most Important Arguments in Western Philosophy (2011: 282-283):
P1. Killing this particular adult human being or child would be seriously wrong.
P2. What makes it so wrong is that it causes the loss of this individual’s future experiences, activities, projects, and enjoyments, and this loss is one of the greatest losses that can be suffered.
C1. Killing this adult human being or child would be seriously wrong, and what makes it so wrong is that it causes the loss of this individual’s future experiences, activities, projects, and enjoyments, and this loss is one of the greatest losses that can be suffered (conjunction, P1, P2).
P3. If killing this particular adult human being or child would be seriously wrong and what makes it so wrong is that it causes the loss of all this individual’s experiences, activities, projects, and enjoyments, and this loss is one of the greatest losses that can be suffered, then anything that causes to any individual the loss of all future experiences, activities, projects, and enjoyments is seriously wrong.
C2. Anything that causes to any individual the loss of all future experiences, activities, projects, and enjoyments is seriously wrong (modus ponens, C1, P3).
P4. All aborting of any healthy fetus would cause the loss to that individual of all its future experiences, activities, projects, and enjoyments.
C3. If A causes to individual F the loss of all future experiences, activities, projects, and enjoyments, then A is seriously wrong (particular instantiation, C2).
C4. If A is an abortion of healthy fetus F, then A causes to individual F the loss of all future experiences, activities, projects, and enjoyments (particular instantiation, P4).
C5. If A is an abortion of a healthy fetus F, then A is seriously wrong (hypothetical syllogism, C3, C4).
C6. All aborting of any healthy fetus is seriously wrong (universal generalization, C5).
In this case, the argument is about abortion in regard to healthy fetuses. This argument, like the one for abortion, is also deductively valid. (Arguments for and against the abortion of unhealthy fetuses will be covered in the future.) Thus, if a fetus is healthy then it should not be aborted since doing so would cause the individual to lose their future experiences, enjoyments, activities, and projects. Thus, the abortion of a healthy fetus is seriously and morally wrong. This argument clearly establishes the fetuses’ right to life if it is healthy.
Conclusion
Both of these arguments for and against abortion are strong; on the “for” side, we have the apparent facts that fetuses, infants, and (nonhuman) animals cannot want their continuance of their mental states since they cannot conceive of their continuance and want of mental states, so if they cannot want their continuance of their mental states they do not have a morally serious right to life and it is, therefore, morally right to kill them painlessly. On the “against” side, we have the facts that aborting healthy fetuses will cause the loss of all future experiences, enjoyments, activities, and projects, and so, the abortion of these healthy fetuses is both seriously and morally wrong.
I will cover these types of arguments—and more—in the future. However, if one is against genetic modification, embryo selection, preimplantation genetic diagnosis (PGD) and ‘eugenics’, then one must, logically, be against the abortion of healthy fetuses as well. These two arguments, of course, have implications for any looming eugenic policies as well, which I will cover in the future.
(I, personally, lean toward the “against” side in this debate; though, of course, the argument presented in this article on the “for” side is strong as well.)
The Physical Reality of Race
1600 words
Rational people can just look at people of different ancestries and see that there is something to what we call “race.” We notice that others look different based on where their ancestors came from and we classify people into different races on the basis of their physical appearance. Anti-biological racial realists may point to the fact that there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016; Hardimon, 2017). While this is true, this does not mean that race is “just a social construct” (a phrase used to deflate the meaning of “race”); it is both a social construct and a biological reality.
The definition of race is simple—a group of populations which genetically transmit heritable characteristics which correspond to that group’s geographic ancestry who also belong to a biological line of descent which was initiated by a geographically isolated and reproductively isolated founding populations (Hardimon, 2017). Note how this definition says nothing about differences in allele frequencies between populations between populations—because, for these purposes, they’re irrelevant for the argument being made. The fact of the matter is, the reality of race hinges on two things: (1) the heritable differences between population groups which were geographically/reproductively isolated and (2) our ability to discern these population groups by their phenotype.
A great book on the history of race, its meaning and how the term was used over the ages is Race: The Reality of Human Differences by Sarich and Miele (2004). For the purposes of this piece, the first two chapters are the most important, since they touch on aspects of race that I have in the past—mainly the fact that we only need phenotype to discern one’s race. People from Europe look phenotypically different from people from Africa who look phenotypically different from people from Asia etc. These differences between these groups are evidence that race exists—these racial differences in phenotype are due, in part, to the climate they evolved in while geographically and reproductively isolated (two conditions for racehood).
Sarich and Miele (2004: 29) write:
Vince [Sarich; one of the authors of the book] naively asked for the legal definition of “race” and was told there wasn’t one.
[…]
As we began working on this book, we discussed the issue of the legal definition of “race” … He informed us that there is still no legal definition of “race”; nor, as far as we know, does it appear that the legal system feels the need for one. Thus, it appears that the most adversarial part of our complex society, the legal system, not only continues to accept the existence of “race” but also relies on the ability of the average individual to sort people into races. Our legal system treats “racial identification” as self-evident …
[…]
The courts have come to accept the commonsense definition of race, and it is this commonsense view that, as we show, best conforms to reality. A look at two recent (2000) cases is illustrative. In both Rice v. Office of Hawaiian Affairs and in Hank v. Rochester School District, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race (even if the racial categories are euphemistically termed “peoples” or “populations”). No special expertise was assumed or granted in defining or recognizing race other than the everyday commonsense usage, as given in the Oxford English Dictionary, that a race is “a group of persons connected by common descent” or “a tribe, nation, or people, regarded as common stock.” The courts and the contending parties, in effect, accepted the existence of race and the ability of the ordinary person to distinguish between races based on a set of physical features.
In Rice v. Office of Hawaiian Affairs, Rice challenged the state of Hawaii since they did not allow him to vote—on the basis that he was not a native Hawaiian, and that the electoral system of Hawaii is for the benefit of Hawaiians and Hawaiians only. Everyone agreed that Rice was a Hawaiian citizen—but he did not have Hawaiian ancestry, so he could not be recognized as “Hawaiian” under state law. However, the SCOTUS overturned the ruling (that Rice should not be allowed to vote on the basis of not having Hawaiian ancestry) 7-2, citing the 15th amendment: “The right of the citizens of the United States to vote shall not be denied or abridged by the United States or any State on account of race, color, or previous condition of servitude.” Sarich and Miele (2004: 31) write “The 15th amendment is explicit—race means what the average person thinks it means—and the majority of the Supreme Court read it that way.” (Also see Hong, 2008 for an overview of the case.)
On the other hand, in Haak v. Rochester School District, the Second Circuit Court of Appeals ruled that a white fourth-grade student named Jessica Haak could transfer from her current district to another district (full of whites) since the transfer program was initiated with the idea of lessening the racial isolation of the adjoining districts. Jessica’s mother cited the 14th amendement, and a district court ruled in their favor but the Second Circuit Court of Appeals overturned the decision. “A “minority pupil” was defined as “a pupil who is of Black or Hispanic origin or is a member of another minority group that historically has been the subject of discrimination” (Sarich and Miele, 2004: 31).
The critical points here are that in both Rice and Haak, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race. No special expertise was assumed or granted in defining or recognizing race other than the everyday usage of the term. In Rice, the court, in effect, took judicial notice of the commonsense definition of race. In Haak, the court accepted physical appearance as a valid means by which the average citizen can recognize races and distinguish among them.
In short, the courts accepted the existence of race, even if the legislature was afraid to use the offending word.
Despite the fact that Sarich and Miele (2004) claim that there is no legal definition of race, Cornell Law School has one definition stating that “the term “racial group” means a set of individuals whose identity as such is distinctive in terms of physical characteristics or biological descent.” While the Law Dictionary, citing the 15th amendment writes that race is “A tribe, people, or nation, belonging or supposed to belong to the same stock or lineage. “Race, color, or previous condition of servitude.” Const U. S., Am. XV.” (Also see Hoffman, 2004 who argues that “race” should not be used in the legal system.)
Notice how Sarich and Miele’s (2004) description of “race” and what “race” is almost—word-for-word—like Spencer’s Blumenbachian partitions (Spencer, 2014). Americans defer to the US Census Bureau on matters of race; the US Census Bureau defers to the Office of Management and Budget (OMB) who speak of sets of populations; these sets of populations correspond to geographic clusters who have distinct phenotypes based on their geographic ancestry, which the average American can discern; therefore race exists. Spencer states that when Americans refer to “race” that Americans refer to both a social construct and a biological reality—that is, Americans socially construct race (think of how Hardimon’s minimalist concept of race is related to the concept of socialrace) but these social constructs do have biological underpinnings which can be discerned in two ways: (1) just observation of phenotypes and (2) looking into the genomes of genetically related individuals who make up these population groups.
Even the ancients distinguished races and sorted them on the basis of hair color/type, skin color, physiognomy etc. “[The Egyptians, Greeks, Romans, Indians, and Chinese] sorted [broad racial groups] based upon the same set of characteristics—skin color, hair form, and head shape” while “it is evident that they relied upon a set of observable features (skin color and form, body build, facial features) quite similar to those used in the commonsense notion of race and the racial classifications of nineteenth-century anthropology to sort the many diverse groups they encountered into a smaller number of categories” (Sarcih and Miele, 2004: 42).
It is very clear that, ever since antiquity at the very least, we have been classifying racial groups on the basis of phenotype—and, come to find out, this is one of the best ways to sort people—and you don’t even need to look at genetic differences between groups. Phenotype is clearly enough to delineate racial groupings, you don’t need genes to delineate race. We only need to recognize that (1) people look different on the basis of where they (or their ancestors) came from; (2) observe that these physical differences between people who come from different places are between real and existing groups; (3) people have common ancestry with others; (4) people derive from distinct geographic locations; so (5) we can infer that race exists.
Race is very clearly a reality—both biologically and socially. At least three sound arguments exist for the existence of race (Sarich and Miele, 2004; Spencer, 2014; Hardimon, 2017; see Hardimon’s and Spencer’s arguments at length). Even those in antiquity delineated races on the basis of physical features—exactly what has been argued by Spencer and Hardimon. Race is physically real—people look different from each other individually, ethnically, and racially.
Biological racial realism is true, and if biological racial realism is true then race exists.
(1) If groups of people look different from each other depending on where their ancestors evolved, then race exists.
(2) Groups of people look different from each other depending on where their ancestors evolved.
(3) Therefore, race exists since people look different depending on where their ancestors evolved.
