
Home » Posts tagged 'Vitamin D'
Tag Archives: Vitamin D
Vitamin D, Physiology, and the Cold
1200 words
I’ve been chronicling the VDH recently since it has great explanatory—and predictive—power. Light skin is a clear adaptation to low UVR, while dark skin is a clear adaptation to high UVR. Dark, highly melanized skin confers advantages in high UVR environments, such as protection against DNA damage, and also absorbs sufficient UV for vitamin D production while also protecting against folate depletion. However, when our ancestors migrated out of Africa, dark skin would not cut it in temperate environments with highly variable UV rays. This is where our highly adaptive physiology came into play, ensuring that we survived in highly variable environments. Light skin was important in low UVR environments in order to synthesize ample vitamin D, however, that synthesized vitamin D then conferred numerous other physiological advantages to the cold.
Eighty to ninety percent of the vitamin D required for humans comes from the sun, whereas ten to twenty percent comes from the diet, such as fatty fish, eggs, and dairy products (fortified with vitamin D, of course) (Ajabshir, Asif, and Nayer, 2014). Humans need to rely on high amounts of UV rays for vitamin D synthesis (Carlberg, 2014) other than Arctic peoples. Since dark skin does not synthesize vitamin D as well as light skin, skin gradually lightened as our ancestors migrated out of Africa (Juzeniene et al, 2009). This was then imperative to the physiologic adaptations that then occurred as our physiology had to adapt to novel, colder environments with fewer UV rays.
Sufficient amounts of vitamin D are highly important for the human musculoskeletal system (Wintermeyer et al, 2016), which is extremely important for birthing mothers. Along with the increased vitamin D synthesis in low UV environments, the heightened production of vitamin D conferred numerous other physiologic benefits which then helped humans adapt to colder environments with more varying UVR.
Vasoconstriction occurs when the blood vessels constrict which leads to heightened blood pressure, whereas vasodilation is the dilation of blood vessels which decreases blood pressure. So evolutionarily speaking, we had to have adaptive physiology in order to be able to “switch” back and forth between vasoconstriction and vasodilation, depending on what the current environment needed. Vasodilation, though, most likely had no advantage in high UV environments, and thus must have been an advantage in low UV environments, where it was more likely to be colder and so, when the blood vessels constrict, blood pressure increases and thus, heat loss could be considerably slowed in these environments due to these physiologic adaptations.
The races also differ, along with many other physiologic abilities, in nitric oxide-mediated vasodilation. Vasodilation is the dilation of blood vessels, which increases blood pressure. Mata-Greenwood and Chen (2008) reviewed the relevant literature regarding black/white differences in nitric oxide-dependent vasorelaxation and concluded that nitric oxide vasodilation is reduced in darker-skinned populations. Thus, we can infer that in lighter-skinned populations nitric oxide vasodilation is increased in lighter-skinned populations, which would have conferred a great physiological advantage when it came to colonizing environments with lower UV rays.
VDR and vitamin D metabolizing enzymes are present in adipose tissue. Tetrahydrobiopterin; which acts as a cofactor in the synthesis of nitric oxide and its primary function is as a vasodilator in the blood vessels (meaning that blood pressure is increased, to keep more heat in the cold) (Chalupsky and Cai, 2005). Since vasodilation is the body’s primary response to heat stress, blood flow increases which allows heat to leave the body. Therefore, the human body’s ability regarding vasodilation and vasoconstriction mechanisms were important in surviving areas with varying UVR.
One function of our adipose tissue is the storage of vitamin D, while vitamin D metabolizing enzymes and VDR are also expressed in the adipocyte (Abbas, 2017). With these known actions of vitamin D on adipose tissue, we can speculate that since vitamin D and the VDR are expressed in adipose tissue, it may have exerted a role in the adipose tissue which may have been important for surviving in cold, low UV environments (see below).
Furthermore, since these mechanisms are brought on by short-term changes, we can infer that it would hardly be of any use in high UVR environments and would be critical in temperate environments. So, vasodilation and vasoconstriction have little to no benefit in high UVR environments but seem to be imperative in temperate environments where UVR varies. It’s also likely that vitamin D influences vasodilation by influential nitric oxide synthesis (see Andrukhova et al, 2014) and vasoconstriction by influencing the renin-angiotensin system (Ajabshir, Asig, and Nayer, 2014).
This would have conferred great benefit to our ancestors as they migrated into more temperate and colder climates. You can read this for information on how adaptive our physiology is and why it’s like that. Because we went into numerous new environments and natural selection couldn’t act quickly enough, therefore the human body’s physiology is extremely adaptive.
What this suggests is that as skin lightened and adapted to low UV, the increased synthesis in vitamin D influenced vasodilation by a strong influence on nitric oxide synthase, along with vasoconstriction, implies that it would have been easier to survive in novel environments due to adaptive physiology and skin color, along with body fat reserves and the physiologic effects of vitamin D on adipose tissue. These physiologic adaptations would have been of no to little use in Africa. Thus, they must have been useful after we migrated out of Africa and experienced wildly varying environments—the whole reason why our physiology evolved (Richardson, 2017: chapter 5).
When the human body is exposed to cold, a few things occur: cutaneous vasoconstriction, shivering (Castellani and Young, 2016), “behavioral thermoregulation” (Young, Sawka, and Pandolf, 1996), while the human body can adapt physiologically to the cold (Young, 1994). The physiologic functions that vitamin D and folate in regard to vasodilation and vasoconstriction, there is a great chance that these effects were important in maintaining energy homeostasis in colder climates.
In sum, the evolution of light skin conferred a great survival advantage to our ancestors. This then upped the production of vitamin D synthesis in the body, which where then of utmost importance in regard to the adaptation of the human physiology to colder, lower-UV environments. Without our adaptive physiological systems, we would not have been able to leave Africa into novel environments. We need both behavioral thermoregulation as well as adaptive physiology to be able to survive in novel environments. Thus, the importance of skin lightening in our evolution becomes clearer:
As humans migrated out of Africa, lighter skin was needed to synthesize vitamin D. This was especially important to women, who needed higher amounts of vitamin D, in order to produce enough calcium for lactation and pregnancy—so the babe had enough calcium to grow its skeleton in the womb. With the uptake in vitamin D synthesis, this then allowed more adaptive physiologic changes that occurred due to the cold, and along with vasodilation and vasoconstriction, along with shivering and adapting behaviorally to the new environments, were our ancestors able to survive. Dark skin cannot synthesize vitamin D as well as light skin in low UV environments; this also can be seen with the lowered production of nitric oxide-dependent vasodilation in dark-skinned populations. Thus, vasoconstriction conferred no physiologic benefit in high UV environments, but almost certainly conferred a physiologic benefit in low UV environments.
Why Are Women Lighter than Men? Skin Color and Sexual Selection
1550 words
Skin color differences between the sexes are always discussed in terms of women being lighter than men, but never men being darker than women. This is seen in numerous animal studies (some reviewed by Rushton and Templer, 2012; read rebuttal here; also see Ducrest, Keller, and Roulin, 2008). Though, the colors that evolved on the animal’s fur due to whatever mate choices are irrelevant to the survival capabilities that the fur, feathers etc give to the organism in question. So, when we look at humans, we lost our protective body hair millions of years ago (Lieberman, 2015), and with that, we could then sweat. So since furlessness evolved in the lineage Homo, there was little flexibility in what could occur due to environmental pressures on skin color in Africa. It should be further noted that, as Nina Jablonski writes in her book Living Color: The Biological and Social Meaning of Skin Color (2012, pg 74)
No researchers, by the way, have explored the opposite possibility, that women deliberately selected darker men!
One hypothesis proposes that lighter skin in women first arose as a byproduct due to the actions of differing levels of hormones in the sexes—with men obviously having higher levels of testosterone, making them darker them women. So according to this hypothesis, light-skinned women evolved since men could tell high-quality from low-quality mates as well as measure hormonal status and childbearing potential, which was much easier to do with lighter- than darker-skinned women.
Another hypothesis put forth is that further from the equator, sexual competition between women would have increased for mates since mates were depleted, and so light skin evolved since men found it more beautiful. Thus, women living at higher latitudes were lighter than women living at lower latitudes because men had to go further to hunt which meant they were more likely to die which caused even greater competition between females, lightening their skin even more. And another, related, argument, proposed that light skin in women evolved due to a complex of childlike traits which includes a higher voice, smoother skin and childlike facial features, which then reduced male competition and aggressiveness. But women did not stay around waiting to be provisioned and they got out and gathered, and hunted sometimes, too.
Harris (2005) proposes that light skin evolved due to parental selection—mothers choosing the lightest daughters to survive, killing off the darker ones. All babies are born pale—or at least lacking the amount of pigment they have later in life. So how would parental—mostly maternal—selection have caused selection for lighter skin in girls as Harris (2005) proposes? It’d be a pretty large guessing game.
The role of sexual selection in regard to human skin color, though, has been tested and falsified. Madrigal and Kelly (2007a) tested the hypothesis that skin reflectance should be positively correlated with distance from the equator. It was proposed by other authors that as our ancestors migrated out of Africa, environmental selection relaxed and sexual selection took over. Their data did not lend credence to the hypothesis and falsified it.
Madrigal and Kelly (2007a: 475) write (emphasis mine):
We tested the hypothesis that human sexual dimorphism in skin color should be positively correlated with distance from the equator, a proposal generated by the sexual selection hypothesis. We found no support for that proposition. Before this paper was written, the sexual selection hypothesis was based on stated male preference data in a number of human groups. Here, we focused on the actual pattern of sexual dimorphism. We report that the distribution of human sexual dimorphism in relation to latitude is not that which is predicted by the sexual selection hypothesis. According to that hypothesis, in areas of low solar radiation, there should be greater sexual dimorphism, because sexual selection for lighter females is not counterbalanced by natural selection for dark skin. Our data analysis does not support this prediction.
Though Frost (2007) replied, stating that Madrigal and Kelly (2007a) presumed that sexual selection was equal in all areas. Madrigal and Kelly (2007b) responded, stating that they tested one specific hypothesis regarding sexual selection and found it to be false. Frost (2007) proposed two hypotheses in order to test his version, but, again, no one has proposed that women select darker men, which could be a cause of lighter-skinned women (though sexual selection does not—and cannot—explain the observed gradation in skin color between men and women).
Skin color differences between men and women first arose to ensure women enough calcium for lactation and pregnancies. Since skin pigmentation protects against UVR but also must generate vitamin D, it must be light or dark enough to ensure ample vitamin D production in that certain climate, along with protecting against the UVR in that climate. So women needed sufficient vitamin D, which meant they needed sufficient calcium to ensure a strong skeleton for the fetus, for breastfeeding and for the mother’s own overall health.
However, breastfeeding new babes is demanding on the mother’s body (calcium reserves are depleted four times quicker), and the calcium the babe needs to grow its skeleton comes directly from the mother’s bones. Even a mother deficient in vitamin D will still give calcium to the babe at the expense of her own health. But she then needs to increase her reserves of calcium in order to ensure future pregnancies aren’t fatal for her or her offspring.
Though, at the moment to the best of my knowledge, there are no studies on calcium absorption, vitamin D levels and the recovery of the female skeleton after breastfeeding. (Though n3 fatty acids are paramount as well, and so a mother must have sufficient fat stores; see Lassek and Gaulin, 2008.) Thus, light-skinned women are most likely at an advantage when it comes to vitamin D production: The lighter they are, the more vitamin D and calcium they can produce for more pregnancies. Since light skin synthesizes vitamin D more efficiently, the body could then synthesize and use calcium more efficiently. The body cannot use and absorb calcium unless vitamin D is present. Since the fetus takes calcium from the mother’s skeleton, ample amounts of vitamin D must be present. For ample amounts of vitamin D to be present, the skin must be light enough to ensure vitamin D synthesis which would be needed for calcium absorption (Cashman, 2007; Gallagher, Yalamanchili, and Smith, 2012; Aloia et al, 2013).
Nina Jablonski writes in her book (2012, 77):
Women who are chronically deficient in vitamin D because of successive pregnancies and periods of breastfeeding experience a form of bone degeneration called osteomalacia. This has serious consequences for infants born of later pregnancies and for mothers themselves, who are at greater risk of breaking bones. It makes sense that protection of female health during the reproductive years would be a top evolutionary priority, so we are now investigating whether, in fact, slightly lighter skin in women might be a fairly simple way of ensuring that women get enough vitamin D after pregnancy and breastfeeding to enable their bodies to recover quickly. The need for maintaining strong female skeletons through multiple pregnancies may have been the ultimate evolutionary reason for the origin of differences in skin color between men and women.
While Jablonski and Chaplin (2000: 78) write:
We suggest that lighter pigmentation in human females began as a trait directly tied to increased fitness and was subsequently reinforced and enhanced in many human populations by sexual selection.
It is obvious that skin color in women represents a complex balancing act between giving the body the ability to synthesize ample vitamin D and protect from UVR. Skin coloration in humans is very clearly highly adaptive to UVR, and so, with differing average levels of UVR in certain geographic locales, skin color would have evolved to accommodate the human body to whichever climate it found itself in—because human physiology is perhaps the ultimate adaptation.
Sexual selection for skin color played a secondary, not primary role (Jablonski, 2004: 609) in the evolution of skin color differences between men and women. There is a delicate balancing act between skin color, vitamin D synthesis, and UVR protection. Women need to produce enough vitamin D in order to ensure enough calcium and its absorption to the baby and then ensure there are ample amounts to replace what the baby took while in the womb in order for future pregnancies to be successful. Sexual selection cannot explain the observed gradation in skin color between the races and ethnies of the human race. In my opinion, the only explanation for the observed explanation is the fact that skin color evolved due to climatic demands, while independent justification exists for the hypothesis as a whole (Jablonski and Chaplin, 2010).
I don’t see any way that sexual selection can explain the observed gradation in skin color around the world. Skin color is very clearly an adaptation to climate, though of course, cultural customs could widen the skin color differences between the sexes, and make women lighter over time. Nevertheless, what explains the observed skin gradation is adaptation to climate to ensure vitamin D synthesis among a slew of other factors (Jones et al, 2018). Sexual selection, while it may explain small differences between the sexes, cannot explain the differences noted between the native human races.
The Vitamin D Receptor and the Updated VDH
2850 words
The relationship between vitamin D and the vitamin D receptor (VDR) has been found to be of recent importance in explaining the modulation of gene expression. The VDR helps us adapt to the climate, is epistatic with skin color genes, and so on. Due to the importance of the VDR, vitamin D, and another nutrient I’ve discussed in the past—folate—this drives the argument that the need to produce vitamin D was an important factor in the evolution of skin colors around the world as migrations out of Africa took place. It is also important to note that other competing hypotheses are not necessarily alternative hypotheses to the VDH (which is short for vitamin D-folate hypothesis), since there is significant overlap between them due to what we now know about the roles of vitamin D, folate (especially due to what we know now about how vitamin D, folate and the VDR regulate gene expression),the VDR, and skin color genes. Thus, the theories have been integrated and the updated hypothesis takes into account the other theories which has significant overlap with the VDH.
Jones et al (2018) is the most recent review of the VDH; in the review, they integrate new findings of folate, vitamin D, the VDR, and skin color genes with other supposedly competing hypotheses into a new and improved VDH model which will be discussed at the end of this article.
Vitamin D
Vitamin D is an important hormone (since it is a steroid, not a vitamin), which is the only one that is produced exogenously (from UV rays). Vitamin D is responsible for many physiologic functions including: regulating calcium levels by increasing calcium absorption, stimulates intestinal absorption of phosphate, stimulates osteoblasts which then produce receptor activator nuclear factor (RANKL) which then stimulates osteoclastogenesis which then activates osteoclasts for bone reabsorption (DeLuca, 2004). It has been further noted that around 5 percent of the human genome is under the influence of vitamin D (Jones et al, 2018).
Folate
Folate is an important water-soluble B vitamin. Since vitamin D and folate are linked by their sensitivities to UVR, then we must look at them independently and see what they do. In the case of folate, UVR causes folate degradation through the absorption of UVRs or, on the other hand, when folate oxidizes through free radicals after UVR exposure (Jones et al, 2018). So the hypothesis proposes that skin color in high UV areas evolved due to the need for protection of folate levels due to UVR degradation. On the other hand, depigmentation occurred in order for the body to produce adequate vitamin D in low UV areas.
Folate is needed to synthesize and repair DNA (Mahmood, 2014). Though common rebuttals to the VDH include supposed lack of evidence for the VDH, Jones et al (2018) write:
The potential impacts of a deficiency of these nutrients on natural selection is an ongoing debate and is a common argument raised against the vitamin D–folate hypothesis. However, these arguments often do not consider that the benefits of an adequate vitamin D and folate status on reproductive success extend far beyond their roles in maintaining reproductive health.
Vitamin D receptor
In recent years, it has been found that the VDR has had a profound influence on our adaptation to local climates our ancestors found themselves in after the trek out of Africa. Most cells and organs of the body have a vitamin D receptor (Wacker and Holick, 2013), so the importance of the VDR and certain genes involved in the production of skin color, vitamin D, and folate can be seen. Thus, evidence for the hypothesis would be differential expression of certain genes that are related to the VDR. Jones et al (2018) report on a few common VDR variants and ethnicity: FOK1 which has a lower frequency in African than European and East Asian populations, and Cdx2 which was highest in Africans and lowest in Europeans. Tiosano et al (2016) reported that multiple loci which are involved with the VDR gene display strong latitudinal clines, which is evidence for the hypothesis.
The VDR helps humans adapt to changes in UV radiation, it is “part of an evolutionary complex that adapts humans to changing UV radiation” (Hochberg and Templeton, 2010: 310). This is further corroborated by the fact that the VDR promoter and skin color genes are epistatic (Popsiech et al, 2014; Tiosano et al, 2016). Skin pigmentation levels, furthermore, determine plasma vitamin D levels and VDR autoregulation (Saccone, Asani, and Bornman, 2015).
The VDR works in concert with retinoic acid receptors (Schrader et al, 1993) which then bind to nucleotide base pairs called the vitamin D-responsive elements (VDRE) which then exert their effects on gene expression (Kato, 2000; Pike and Meyer, 2010; Janik et al, 2017).
Gene expression
Vitamin D elicits numerous functions on gene expression through the VDR, by binding elements of vitamin D to the target genes. Since the VDR works together with other receptors that bind to the VDRE, they can have strong effects on gene expression. Now, we know that vitamin D and folate are important for humans. We know that the VDR gene appears to be under strong selection, though only in the context of other genes (Tiosano et al, 2016). Thus, the VDR—along with folate and vitamin D—are extremely important for gene expression and the adaptation of the human body to differing climates.
Competing hypotheses
Skin barrier hypothesis
The skin barrier hypothesis (SBH) proposes that dark skin color arose to protect against environmental damage. This hypothesis is based on the fact that darker-pigmented peoples posess an enchanced barrier function in comparison to ligher-pigmented people, which is mainly due to the role of melanin in the scattering of UVR across the skin (Jones et al, 2018). Jones et al state that this hypothesis is “proposed as a discrete theory to the vitamin D-folate hypothesis“, but since both vitamin D and folate both have other responsibilities in the human body such as the development of skin structure, and the development of defense mechanisms that protect against UV radiation including heat and microbial stressors.
Folate may also have another important role in the human body: regulating the production, and stabilizing tetrahydrobiopterin. Melanin supports folate from UVR degradation, which then supports folate’s influence on melanin. But, as Jones et al write, tetrahydrobiopterin also acts as a cofactor in the synthesis of nitric oxide which is important in regard to vasoconstriction (blood vessel constriction). Vasoconstriction is related to increased heat flow since blood vessels are constricted, along with an increase in heart rate. As I have noted in the past, shivering revs the body’s metabolism in cold clmates in order to produce ample heat. Jones et al (2018) write:
From an evolutionary perspective, our ability to maintain vasodilation/vasoconstriction mechanisms would have been important in surviving varying UVR environments. As these mechanisms may been seen as relatively short-term responses to temperature changes, they are likely to be of greater importance in temperate UVR environments rather than environments of high UVR. This is supported by nitric oxide dependent vasodilation shown to be reduced in darkly skinned populations [59]. This suggests that vasodilation processes offer no advantage in extreme UVR environments but may be important in temperate UVR environments, where seasonal and daily temperature fluctuations are seen.
Thus, since there would be no advantage for this mechanism in equatorial climates, it must be for more colder, Arctic climates which further lends credence to the VDH. (Since vitamin D and folate play many roles in regard to human physiologic adaptation to climate, along with the VDR.)
Metabolic conservation hypothesis
This hypothesis proposes that our ancestors became depigmented after the migrations out of Africa since there was a need to draw energetic resources away from melanin production and move that energy that would have been for melanin production for other metabolic processes that a population would need in a colder environment. Thus, it is argued that the lighter skin of European and East Asian populations can be explained by the need energetic resources being moved away from pigmenting the skin to other, more important, metabolic processes that the ancestors of Europeans and East Asians experienced. But this hypothesis has numerous premises of the VDH, including the main premise: that human skin depigmented as we migrated into areas with fewer UV rays (Jones et al, 2018). Thus, vitamin D was extremely important in driving the effects of vasodilation/vasoconstriction.
Clearly, the role of vitamin D in the adipose tissue was important for human adaptation to colder climates. Since lighter skin can produce more vitamin D in low UV climates, this was another factor that helped when we left Africa: skin lightened for better vitamin D synthesis. Since vitamin D synthesis is related to gene expression and expression of about 5 percent of our genomes, the production of more vitamin D was beneficial. So depigmentation, while being primarily due to low UV radiation, can also be seen to allow for more efficient physiologic responses and adaptations to the newer, colder climates.
Skin mutagenesis hypothesis
The last competing theory is the skin mutagenesis hypothesis. This hypothesis proposes that skin pigmentation arose as a mechanism to protect against various skin cancers. The hypothesis is based on the fact that darker-pigmented individuals are at lower risk of developing skin cancers since their skin pigmentation can fight off UV radiation. Of course, knowing what we know about vitamin D and folate, these two agents would be involved regarding this hypothesis, since both agents have photoprotective effects. Vitamin D is extremely important to DNA repair (Graziano et al, 2016), as vitamin D reduces cell and DNA damage.
Though many authors dispute the claims of this hypothesis since the effects of skin cancer would occur after the reproductive years and would thusly not have an effect on natural selection for skin color. Though those who argue for the validity of the hypothesis propose that it would help in hunter-gatherer peoples whose old train their young their ways of life.
Since these interactions have between these variables have been verified at the molecular genetic level, this lends even more credence to the VDH. (The findings inclue the frequency of common VDR variants between different ethnic groups, to UVR and folate metabolism genes which were found to be significantly associated with the frequency of 16 common folate variants and skin pigmentation in a genomic analysis of 30,000 people which were novel relationships; Jones et al, 2018a). These findings discussed by Jones et al (2018b) “indicate the existence of interactions between UVR, skin type, and vitamin D and folate genes, and they provide supporting molecular evidence for the vitamin D–folate hypothesis.”
Sexual selection
Madrigal and Kelly (2007a) tested a sexual selection hypothesis proposed by a few proponents of the sexual selection hypithesis. Madrigal and Kelly (2007a) tested the hypothesis that skin color reflectance should be positively correlated with distance from the equator. They, however, showed that the pattern in skin color dimorphism seen around the globe was not consistent with the sexual selection hypothesis, and thus their data did not lend credence to the sexual selection hypothesis. The hypothesis states that in areas with low UV radiation, environmental selection for skin color should be relaxed and there should be a higher rate of sexual dimorphism in peoples from northerly climates due to sexual selection for lighter-skinned women. Nevertheless, the data compiled by Madrigal and Kelly (2007a) do not lend credence to the hypothesis.
Frost (2007) responded that Madrigal and Kelly (2007a) presumed that sexual selection was equal in all areas, but was constrained by natural selection for dark skin. Frost (2007) also states that sexual dimorphism in human skin color may not be able to be expressed in lighter-skinned populations at higher latitudes. Frost’s objections stem from the fact that Madrigal and Kelly tested a specific hypothesis proposed by proponents of the sexual selection hypothesis, though Madrigal and Kelly hope that Frost can test his hypotheses. However, I think it’s a moot issue. Sexual selection for women occurred after selection for light skin due to vitamin D synthesis which ensured more calcium for pregnancy and lactation.
Thusly, sexual selection for lighter skin would continue to ensure ample vitamins for women and their pregnancies and lactation to feed their babies. This would further be butressed by the fact that vitamin D exerts effects on the adipocites which lends even more credence to the claim that light skin evolved first for vitamin D synthesis. Vitamin D then exerted effects on the adipocite since more vitamin D could be produced in the absence of high levels of UV, which then aided in human physiologic adaptations to climate.
Integration of current skin color theories
As can be seen from the competing theories, they are not necessarily explaining different things, and each supposed competing theory has an aspect from the VDH in it. Thusly, it is possible to integrate the so-called competing theories into a larger explanatory framework.
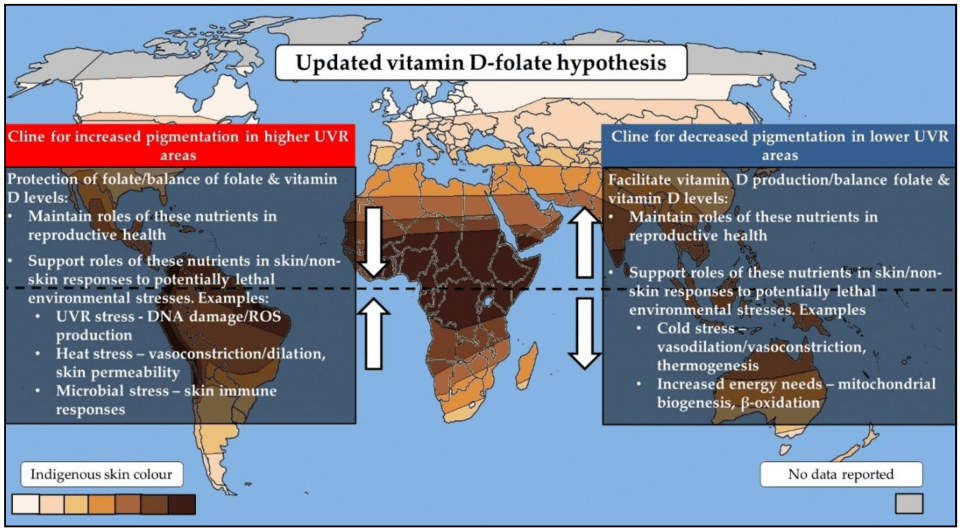
Jones et al (2018b) update the VDH by integrating the other theories into it, since they are similar and do not contradict the VDH (since aspects of each one can be used to explain different aspects of the VDH). The updated hypothesis is thus:
Vitamin D and folate have differing sensitivities to UVR. Vitamin D can be synthesized following UVR exposure, folate may be degraded. So the VDH proposes that the two differing skin colors (light and dark) evolved at differing latitudes as a “balancing mechanism” to maintain adequate levels of the two agents vitamin D and folate. Since adequate levels of vitamin D and folate were maintained, there would be no ill health effects after migrating into colder climates. Vitamin D and folate both act as photoprotectors of the skin and can decrease environmental stressors. Vitamin D also exerts important effects on adipocites—both types—which then further aid in human physiologic adaptations to the cold. Perhaps most importantly, the VDR and skin color genes are epistatic—the VDR is imperative in the human body’s adaptation to new climates.
The latest research (reviewed by Jones et al, 2018b) show strong support for the interaction between genes and folate/vitamin D processes with skin pigmentation and UVR.
Health Disparities
Health disparities due to vitamin D deficiency are well-noted in the literature. Human migrations over the recent centuries and decades have caused environmental mismatches between a population’s adapted skin type and current UV level in the location the population migrated to. Many darkly-pigmented people now live in areas with low UVR, and thusly suffer from health consequences. This leads to them either not having an adequate vitamin D-folate balance along with the risk of not having the adequate skin protections for a given environment, since UV rays influence folate and vitamin D production and so, a mismatched skin color to UV environment would cause problems for skin protection since the environment is not ancestral to that certain skin color.
As I have previously noted, it has been argued that blacks are not vitamin D deficient, and thusly not vitamin D deficient. Though, these claims rest on a slew of false arguments that have since been rebutted. It has been argued that since blacks are deficient in vitamin D, which begins in the womb, and vitamin D deficiencies cause changes in large and small arteries and arterials, that vitamin D deficiency could be the cause of higher rates of hypertension in black Americans (Rostand, 2010).
Conclusion
The role of vitamin D, folate, the VDR, and certain genes is under further invesitgation. This group of agents exert powerful effects on human physiology which then help with the adaptation of humans to differing climates. Folate and the vitamin D receptor play a crucial role in protecting the skin from environmental and microbial stressors. Vitamin D and the VDR are expressed in the adipose tissue, while vitamin D regulates adipogenesis and adipocite apoptosis (Abbas, 2017). Further evidence shows that there are multiple loci that are involved in the VDR that show latitudinal clines (Tiosano et al, 2016). One of the most things that lends credence to the VDH is the fact that the VDR and skin color genes are epistatic and help humans adapt to climates.
The VDH is in great shape, contrary to popular belief (Elias, Williams, and Bikle, 2016). The VDH is one of the only games in town to explain the skin color gradient noticed around the world, with vitamin D being the only agent that accounts for skin color differences. The VDH explains how and why human skin color is vastly different, and the main reason is adaptation to UV rays—or lack thereof.
Grant (2018) concludes that:
The UVB–vitamin D–cancer hypothesis has considerable supporting scientific evidence from a variety of study types: geographical ecological, observational, and laboratory studies of mechanisms, as well as several clinical trials.
Clearly, the VDH explains the incidence of the observed skin gradiation around the world the best out of the so-called competing hypotheses (which are similar enough to the VDH to where they can be absorbed into the VDH). Most importantly, the VDH predicted a novel fact—that molecular genetic evidence would show that light skin evolved independently numerous times in our lineage (Jablonksi and Chaplin, 2009).
Arctic Skin Color and the Vitamin D Hypothesis
1400 words
The Vitamin D Hypothesis (VDH) purports to explain the range of skin colors observed between races/ethnies around the world. Since there are little UVR and even less vitamin-D-producing UVB in the northern hemisphere, other ways of producing/getting ample amounts of vitamin D were imperative for survival. Locations such as the far north were uninhabited up until 12,000 years ago—the explanation being that populations didn’t have the culture to survive such harsh conditions (see Goebel, 1999; Bergman et al, 2004). However, a more likely reason was that there were biological limits on the production of vitamin D due to the lack of UVB rays for most of the year. In this article, I will discuss the skin color of Arctic peoples and why it does not follow the simple gradient of UVB around the world.
To overcome the biological limitations of little to no UVB throughout the year, they needed to supplement with foods to get ample amounts of vitamin D—to cover what they did not get from the weak UVB rays. To overcome the limitation of their environment and vitamin D production, they had to consume fatty animals who had ample stores of vitamin D in their systems. The types of foods allowed peoples to live so far north, since there were little vitamin-D-producing UVB rays, lifestyle and culture is how we conquered the unforgiving far north.
Peoples like the Inuit and Saami eat a diet that is high in vitamin D. Inuits, for example, eat a diet high in vitamin D and n-3 fatty acids (Schaebel et al, 2015). Due to the high vitamin D intake from their diet, they were able to supplement what they did not get from the sun in their diet and thusly were able to live in the unforgiving cold north due to their diet high in vitamin D (Deng and Xu, 2018). Their dark skin color can be explained in a few ways: their diet (high in vitamin-D-rich marine mammals), UVB rays bouncing off ice, snow, and water, and they are recent migrants to those climes, which would explain their darker skin color compared to other populations that have evolved for a longer time in these climates (Jablonski and Chaplin, 2002).
When people look at Arctic people such as the Inuit, they look at their skin color and see the amount of UVB rays they receive during the year and presume that the VDH is wrong because, according to the VDH, Arctic peoples should have the lightest skin but have dark skin—compared to others who evolved recently in those latitudes—but they have dark-ish skin for that latitude. The answer is simple: they were able to consume enough vitamin D in their diet—a lack of vitamin D production/consumption was one barrier to living in the far north which was then overcome with culture and the foods peoples eat.
The environment of the Arctic is dim and dark for most of the year, though during the summer, of course—when they are most active—they are bathed in solar radiation which is then reflected by the snow, ice, and water. Fresh white snow reflects 94 percent UVA rays and 88 percent of UVB rays. Chadysiene and Girgzdys (2008; 87) write:
The average data of experimental measurements show that maximum albedo of UVA radiation (of about 94%) was at 1 p.m. in comparison with albedo of UVB radiation of about 88% at 2 p.m. The measurements of albedo were performed on fresh snow with big crystals.”
For example, Inuit populations in northern Greenland report spending up to 16 hours outdoors in the spring and summer months, and would be exposed to UV rays bouncing from ice, snow, and water (Andersen, Jakobsen, and Laurberg, 2012). Exposure to UV rays for this extended period of time—along with eating a diet high in vitamin D—is enough to explain their skin color.
Clearly, Arctic people get bathed in UVB and UVA rays from being reflected off the snow and ice, which gives them their darker skin color. They have the ability to tan (which is distinct from the American term “tanning”) and their tanning ability protects them from high doses of UVR that are reflected from the snow whereas their diet high in vitamin D gives them their darkish skin color and allows them to remain healthy in such a harsh, unforgiving environment.
Nina Jablonski has been writing about the VDH for about 30 years. Jablonski writes in her book Living Color: The Biological and Social Meaning of Skin Color (2012: 68):
Traditional cultures of the Inuit and the Saami center on harvesting vitamin-D-rich foods. The dietary focus for both groups has compensated for the vitamin D they cannot produce in their skin. Both peoples remain healthy when they stick to their traditional diets but suffer badly from vitamin D deficiencies when they switch to Western diets that are lower in vitamin D.
Here’s the thing: when these populations move away from their natural, vitamin-D-rich diet, they suffer from many deficiencies regarding vitamin D, even today many Inuit populations suffer from vitamin D deficiency, both children, and adults (Hayek, 2011). So the change in the Inuit diet is the cause of these deficiencies—their traditional diet was high in vitamin D, but their new diet (the Western diet) is low in vitamin D; since they have dark skin and the UVB is so variable throughout the year, they then suffer from vitamin D deficiencies (Sharma et al, 2011). Sharma et al (2011: 475) conclude that Arctic people are at-risk for vitamin D deficiency due to lack of UVB exposure, moving away from a traditional diet high in vitamin D to a Western diet low in vitamin D, combined with their dark skin.
Frost (2012) claims that while the explosion of rickets in Arctic populations is due to a change in diet (shifting away from a high meat diet) and “increased consumption of certain reactive substances: phytic acids in commercially processed cereals; sodium bicarbonate in baking soda; and aluminum hydroxide in antacids” (Frost, 2012). The dominant source of vitamin D for the Inuit is their diet (Schaebel et al, 2015), and so, due to their shift away from their natural diet high in fatty fish and vitamin D, once they began eating a diet not ancestral to them, then the maladies began. We can see this with every country/population that begins to eat a new diet full of processed foods.
Since the frequency of rickets has exploded in populations that eat a Western-like diet and not their traditional diet, this implies that the traditional diet provided enough vitamin D, and when they began eating a new diet with less vitamin D, then these problems such as rickets occurred.
To end these implications, the Inuit need to return to consuming a traditional diet, since their traditional diets have the adequate vitamins and minerals needed to survive in the environment they are currently in (Koladhooz et al, 2013). Higher BMI (body mass index), their skin color, and the latitude of where they live contribute to low vitamin D production. Inuits who consumed a low number of traditional food items were more likely to be deficient in vitamin D (Anderson et al, 2013) while this deficiency is seen even in Inuit school children (Hayek, Egeland, and Weiler, 2010; Singleton et al, 2015).
In sum, there is no anomaly regarding the skin color of Arctic peoples; the hypothesis is called “the vitamin D hypothesis”, and so they get ample vitamin D from the reflection of UV rays from the snow, ice, and water. Reasons for the darkness of their skin include the fact that they are recent migrants to those locations, they consume a diet high in vitamin D, and the reflection of UV rays from albedo surfaces.
The hypothesis that UVB exposure explains the observed skin gradients predicted a novel fact—that populations that migrated out of Africa would be seen to have light skin. This occurred multiple times through three different molecular pathways, in the Neanderthals (Lalueza-Fox et al, 2007) and Europeans and East Asians (different molecular mechanisms for them; Norton et al, 2007). This was a risky, successful and novel prediction made by Jablonski and Chaplin (2000). That this does not hold for Arctic people is not a blow to the hypothesis; it is perfectly explained by the bouncing of UVR off of albedo surfaces and a high vitamin D diet. Skin color is an adaptation to UV rays.
Cold Winter Theory, the Vitamin D Hypothesis and the Prediction of Novel Facts
2400 words
HBDers purport that as one moves further north from Africa that IQ raises as a function of how the population in question needed to survive. The explanation is that as our species migrated out of Africa, more “intelligence” was needed and this is what explains the current IQ disparities across the world: the ancestors of populations evolving in different areas with different demands then changed their “IQs” and this then is responsible for differential national development between nations. Cold winter theory (CWT) explains these disparities.
On the other hand is the vitamin D hypothesis (VDH). The VDH purports to explain why populations have light skin at northern latitudes. As the migration north out of Africa occurred, peoples needed to get progressively lighter in order to synthesize vitamin D. The observation here is that as light skin is selected for in locations where UVB is absent, seasonal or more variable whereas dark skin is selected for where UVB is stronger. So we have two hypotheses: but there is a problem. Only one of these hypotheses makes novel predictions. Predictions of novel predictions are what science truly is. A predicted fact is a novel fact for a hypothesis if it wasn’t used in the construction of the hypothesis (Musgrave, 1988). In this article, I will cover both the CWT and VDH, predictions of facts that each made (or didn’t make) and which can be called “science”.
Cold winter theory
The cold winter theory, formulated by Lynn and Rushton, purports to give an evolutionary explanation for differences in national IQs: certain populations evolved in areas with deathly cold winters in the north, while those who lived in tropical climes had, in comparison to those who evolved in the north, an “easier time to live”. Over time as populations adapted to their environments, differences in ‘intelligence’ (whatever that is) evolved due to the different demands of each environment, or so the HBDers say.
Put simply, the CWT states that IQ differences exist due to different evolutionary pressures. Since our species migrated into cold, novel environments, this was the selective pressure needed for higher levels of ‘intelligence’. On the other hand, humans who remained in Africa and other tropical locations did experience these novel, cold environments and so their ‘intelligence’ stayed at around the same level as it was 70,000 years ago. Many authors hold this theory, including Rushton (1997), Lynn (2006), Hart, (2007) Kanazawa (2008), Rushton and Templer (2012; see my thoughts on their hypothesis here) and Wade (2014). Lynn (2013) even spoke of a “widespreadonsensus” on the CWT, writing:
“There is widespread consensus on this thesis, e.g. Kanazawa (2008), Lynn (1991, 2006), and Templer and Arikawa (2006).”
So this “consensus” seems to be a group of his friends and his own publications. We can change this sentence to ““There is widespread consensus on this thesis, including two of my publications, a paper where the author assumes that the earth is flat: “First, Kanazawa’s (2008) computations of geographic distance used Pythagoras’ theorem and so the paper assumed that the earth is flat (Gelade, 2008).” (Wicherts et al, 2012) and another publication where the authors assume hot weather leads to lower intelligence. Oh yea, they’re all PF members. Weird.” That Lynn (2013) calls this “consensus” is a joke.
What caused higher levels of ‘intelligence’ in those that migrated out of Africa? Well, according to those who push the CWT, finding food and shelter. Kanazawa, Lynn, and Rushton all argue that finding food, making shelter and hunting animals were all harder in Eurasia than in Africa.
One explanation for high IQs of people who evolved recently in northern climes is their brain size. Lynn (2006: 139) cites data showing the average brain sizes of populations, along with the temperatures in that location:
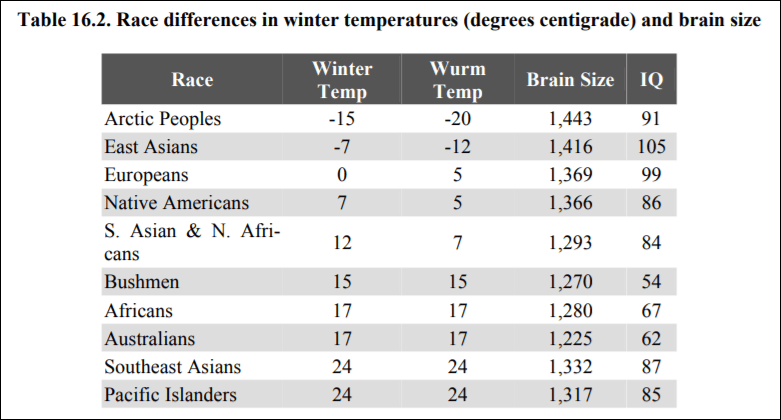
Do note the anomaly with the Arctic peoples. To explain this away in an ad-hoc manner, Lynn (2006: 156-7) writes:
These severe winters would be expected to have acted as a strong selection for increased intelligence, but this evidently failed to occur because their IQ is only 91. The explanation for this must lie in the small numbers of the Arctic Peoples whose population at the end of the twentieth century was only approximately 56,000 as compared with approximately 1.4 billion East Asians.
This is completely ad-hoc. There is no independent verifier for the claim. That the Arcitic don’t have the highest IQs but experienced the harshest temperatures and therefore have the biggest brain size is a huge anomaly, which Lynn (2006) attempts to explain away by population size.
He does not explain why natural selection among Arctic peoples would result in larger brain sizes or enhanced visual memory yet the same evolutionary pressures associated with a cold environment would not also produce higher intelligence. Arctic peoples have clear physical adaptations to the cold, such as short, stocky bodies well-suited to conserving heat.
Furthermore, the argument that Lynn attempts is on the mutations/population size is special pleading—he is ignoring anomalies in his theory that don’t fit it. However, “evolution is not necessary for temperature and IQ to co-vary across geographic space” (Pesta and Poznanski, 2014).
If high ‘intelligence’ is supposedly an adaptation to cold temperatures, then what is the observation that disconfirms a byproduct hypothesis? On the other hand, if ‘intelligence’ is a byproduct, which observation would disconfirm an adaptationist hypothesis? No possible observation can confirm or disconfirm either hypothesis, therefore they are just-so stories. Since a byproduct explanation would explain the same phenomena since byproducts are also inherited, then just saying that ‘intelligence’ is a byproduct of, say, needing larger heads to dissipate heat (Lieberman, 2015). One can make any story they want to fit the data, but if there is no prediction of novel facts then how useful is the hypothesis if it explains the data it purports to explain and only the data it purports to explain?
It is indeed possible to argue that hotter climates need higher levels of intelligence than colder climates, which has been argued in the past (see Anderson, 1991; Graves, 2002; Sternberg, Grigorenko, and Kidd, 2005). Indeed, Sternberg, Grigorenko, and Kidd (2005: 50) write: “post hoc evolutionary arguments … can have the character of ad hoc “just so” stories designed to support, in retrospect, whatever point the author wishes to make about present-day people.” One can think up any “just-so” story to explain any data. But if the “just-so” story doesn’t make any risky predictions of novel facts, then it’s not science, but pseudoscience.
Vitamin D hypothesis
The VDH is simple: those populations that evolved in areas with seasonal, absent, or more variable levels of UVB have lighter skin than populations that evolved in areas with strong UVB levels year-round (Chaplan and Jablonksi, 2009: 458). Robins (2009) is a huge critic of the VDH, though her objections to the VDH have been answered (and will be discussed below).
The VDH is similar to the CWT in that it postulates that the adaptations in question only arose due to migrations out of our ancestral lands. We can see a very strong relationship between high UVB rays and dark skin and conversely with low UVB rays and light skin. Like with the CWT, the VDH has an anomaly and, coincidentally, the anomaly has to do with the same population involved in the CWT anomaly.
Arctic people have dark-ish skin for living in the climate that they do. But since they live in very cold climates then we have a strange anomaly here that needs explaining. We only need to look at the environment around them. They are surrounded by ice. Ice reflects UVB rays. UVB rays hit the skin. Arctic people consume a diet high in vitamin D (from fish). Therefore what explains Arctic skin color is UVB rays bouncing off the ice along with their high vitamin D diet. The sun’s rays are, actually, more dangerous in the snow than on the beach, with UVB rays being 2.5 more times dangerous in the snow than beach.
Evolution in different geographic locations over tens of thousands of years caused skin color differences. Thus, we can expect that, if peoples are out of the conditions where their ancestors evolved their skin color, that there would then be expected complications. For example, if human skin pigmentation is an adaptation to UV rays (Jablonski and Chaplan, 2010), we should expect that, when populations are removed from their ancestral lands and are in new locations with differing levels of UV rays, that there would be a subsequent uptick in diseases caused by vitamin D deficiencies.
This is what we find. We find significant differences in circulating serum vitamin D levels, and these circulating serum vitamin D levels then predict health outcomes in certain populations. This would only be true if sunlight influenced vitamin D production and that skin progressively gets lighter as one moves away from Africa and other tropical locations.
Skin pigmentation regulates vitamin D production (Neer, 1975). This is due to the fact that when UVB rays strike the skin, we synthesize vitamin D, and the lighter one’s skin is, the more vitamin D can be synthesized in areas with fewer UVB rays. (Also see Daraghmeh et al, 2016 for more evidence for the vitamin D hypothesis.)
P1) UV rays generate vitamin D in human skin
P2) Human populations that migrate to climates with less sunlight get fewer UV rays
P3) To produce more vitamin D, the skin needs to get progressively lighter
C) Therefore, what explains human skin variation is climate and UV rays linked to vitamin D production in the skin.
Novel predictions
Science is the generation of novel facts from risky predictions (Musgrave, 1988; Winther, 2009). And so, hypotheses that predict novel facts from risky predictions are scientific hypotheses, whereas those hypotheses that need to continuously backtrack and think up ad-hoc hypotheses are then pseudoscientific. Pseudoscience is simple enough to define. The Stanford Encyclopedia of Philosophy defines it as:
“A pretended or spurious science; a collection of related beliefs about the world mistakenly regarded as being based on scientific method or as having the status that scientific truths now have.”
All theories have a protective belt of ad hoc hypotheses. Theories become pseudoscientific when they fail to make new predictions and must take on more and more ad-hoc hypotheses that have no predictive value. If the ad-hoc hypotheses that are added to the main hypothesis have no predictive value then the new explanations for whichever hypothesis that is in danger of being falsified are just used to save the hypothesis from being refuted and it thus becomes pseudoscience.
In the case of CWT, it makes no prediction of novel facts; it only explains the data that it purports to explain. What is so great about the CWT if it makes no predictions of novel facts and only explains what it purports to explain? One may attempt to argue that it has made some ‘novel’ predictions but the ‘predictions’ that are proposed are not risky at all.
For example, Hart (2007: 417) makes a few “predictions”, but whether or not they’re “risky” or “novel” I’ll let you decide (I think they’re neither, of course). He writes that very few accomplishments will be made by Africans, or Australian or New Guinean Aborigines; members of those groups will not be highly represented in chess; and that major advances in scientific fields will come from those of European ancestry or the “Monglids”, Koreans, Chinese or Japanese.
On the other hand, Hart (2007: 417) makes two more “predictions”: he says that IQ data for Congoid Pygmies, Andaman Islanders, and Bantu-speaking people are few and far between and he believes that when enough IQ testing is undertaken there he expects IQ values between 60 and 85. Conversely, for the Lapps, Siberians, Eskimoes, Mongols and Tibetans, he predicts that IQ values should be between 85-105. He then states that if these “predictions” turn out to be wrong then he would have to admit that his hypothesis is wrong. But the thing is, he chose “predictions” that he knew would come to pass and therefore these are not novel, risky predictions but are predictions that Hart (2007) knows would come to pass.
What novel predictions has the VDH made? This is very simple. The convergent evolution of light skin was predicted in all hominids that trekked out of Africa and into colder lands. This occurred “because of the importance of maintaining the potential for producing pre-vitamin D3 in the skin under conditions of low annual UVB (Jablonski and Chaplin, 2000; Jablonski, 2004)” while these predictions “have been borne out by recent genetic studies, which have demonstrated that depigmented skin evolved independently by different molecular mechanisms multiple times in the history of the human lineage” (Chaplan and Jablonksi, 2009: 452). This was successfully predicted by Chaplan and Jablonski (2000).
The VDH still holds explanatory scope and predictive success; no other agent other than vitamin D can explain the observation that light skin is selected for in areas where there is low, absent or seasonal UVB. Conversely, in areas where there is a strong, year-round presence of UVB rays, dark skin is selected for.
Conclusion
Scientific hypotheses predict novel facts not known before the formulation of the hypothesis. The VDT has successfully predicted novel facts, whereas I am at a loss thinking of a novel fact that the CWT predicted.
In order to push an adaptationist hypothesis for CWT and ‘intelligence’, one must propose an observation that would confirm the adaptationist hypothesis while at the same time disconfirming the byproduct hypothesis. Since byproducts are inherited to, the byproduct hypothesis would predict the same things that an adaptationist hypothesis would. Thus, the CWT is a just-so story since no observation would confirm or disconfirm either hypothesis. On the other hand, the CWT doesn’t make predictions of novel facts, it makes “predictions” that are already known and would not undermine the hypothesis if disproved (but there would always be a proponent of the CWT waiting in the wings to propose an ad-hoc hypothesis in order to save the CWT, but I have already established that it isn’t science).
On the other hand, the VDT has successfully predicted that hominins that trekked out of Africa would have light skin which was then subsequently confirmed by genomic evidence. The fact that strong UVB rays year-round predict dark skin whereas seasonal, absent, or low levels of UVB predict light skin has been proved to be true. With the advent of genomic testing, it has been shown that hominids that migrated out of Africa did indeed have lighter skin. This is independent verification for the VDH; the VDH has predicted a novel fact whereas the CWT has not.
Race and Vitamin D Deficiency
1600 words
Vitamin D is an important “vitamin” (it is really a steroid hormone). It is produced when the skin (the largest organ in the body) is exposed to the sun’s UVB rays (Nair and Maseeh, 2012). So this is one of the only ways to get natural levels of UVB. We can then think that, if a population is outside of its natural evolutionary habitat (the habitat where that skin color evolved), then we should note numerous problems caused by the lack of vitamin D in whichever population is studied outside of a location that doesn’t get the correct amount of UVB rays from the sun.
Black Americans are more likely than other ethnies to be deficient in vitamin D (Harris, 2006; Cosman et al, 2007; Nair, 2012; Forest and Stuhldreher, 2014; Taksler et al, 2014). But, paradoxically, low vitamin D levels don’t cause weaker bones in black Americans (O’Conner et al, 2014). However, like with all hypotheses, there are naysayers. For example. Powe et al (2013) argue that vitamin D tests misdiagnose blacks, that blacks have a form of the vitamin that cells can use called 25-hydroxyvitamin D. They conclude: “Community-dwelling black Americans, as compared with whites, had low levels of total 25-hydroxyvitamin D and vitamin D–binding protein, resulting in similar concentrations of estimated bioavailable 25-hydroxyvitamin D. Racial differences in the prevalence of common genetic polymorphisms provide a likely explanation for this observation.” Though there are a whole host of problems here.
The limitations of Powe et al (2013) striking: it was cross-sectional and observational (like most nutrition studies) so they were unable to predict effects of vitamin-D binding protein on bone fractures; no data on the consumption of vitamin D supplements; measurement of bone turnover markers, urinary calcium excretion and levels of 1,25-dihydroxyvitamin D may explain the effect of VDBP (vitamin D-binding protein) on mineral metabolism; and they relied on a calculation, rather than a measurement of 25-hydroxyvitamin D levels.
Powe et al’s (2013) findings, though, have been disputed. Using different measurement tools from Powe et al (2013), Henderson et al (2015) conclude that “Counter to prior observations by immunoassay, VDBG concentrations did not vary by race.” While Bouillon (2014) writes: In our view, black Americans, as compared with white Americans, have lower levels of not only total 25-hydroxyvitamin D but also free or bioavailable 25-hydroxyvitamin D.” And finally, Hollis and Bikle (2014) write: “Specifically, for any given physically measured level of bio-available 25-hydroxyvitamin D, the authors are overestimating bio-available 25-hydroxyvitamin D by 2 to 2.5 times owing to underestimation of vitamin D–binding protein in blacks.”
Either way, even if what Powe et al (2013) conclude is true, that would not mean that black Americans should not supplement with vitamin D, since many diseases and health problems are associated with low vitamin D intake in blacks, including osteoporosis, cardiovascular disease, cancer, diabetes, and other serious conditions (Harris, 2006). An indirect relationship between low levels of vitamin D and hypertension is also noted (Mehta and Agarwal, 2017). Since there is an indirect relationship between vitamin D levels and hypertension, then we should keep an eye on this because black Americans have some of the highest levels of hypertension in the world (Ferdinand and Armani, 2007; see also Fuchs, 2011).
Vitamin D is, of course, important for skeletal and nonskeletal health (Kennel et al, 2010). So if vitamin D is important for skeletal and nonskeletal health, we should see more diseases in black Americans that imply a lack of this steroid in the body. Although blacks have stronger bones even when deficient in vitamin D, it is still observed that black children who break their forearms have less vitamin D circulating in their blood (Ryan et al, 2011). This observation is borne out by the data, since black children are more likely to be deficient in vitamin D compared to other ethnies (Moore, Murphy, and Hollick, 2005). Since black skin predicts vitamin D deficiency (Thomas and Demay, 2000), it seems logical to give vitamin D supplements to children, especially black children, on the basis that it would help lower incidences of bone fractures, even though blacks have stronger bones than whites.
Furthermore, physiologically “normal” levels of vitamin D differ in blacks compared to whites (Wright et al, 2012). They showed that it is indeed a strong possibility that both whites and blacks have different levels of optimum vitamin D. Wright et al (2012) showed that there is a relationship between 25(OH)D levels and intact parathyroid hormone (iPth); for blacks, the threshold in which there was no change was 20 ng/ml whereas for whites it was 30 ng/ml which suggests that there are different levels of optimal vitamin D for each race, and the cause is due to skin color. Thus, physiologically “normal” levels of vitamin D differ for blacks and whites.
There is also a high prevalence of vitamin D deficiency/insufficiency and asthma in black inner-city youth in Washington DC (Freishtat et al, 2010). We can clearly see that, even though black Americans have stronger bones than white Americans and vitamin D predicts bone strength, the fact that blacks have stronger bones than whites even while being deficient in vitamin D on average does not mean that black Americans should not supplement with vitamin D, since it would ameliorate many other problems they have that are related to vitamin D deficiency.
There are also racial differences in prostate cancer (PCa) acquisition too, and vitamin D deficiency may also explain this disparity (Khan and Partin, 2004; Bhardwaj et al, 2017). I have heavily criticized the explanations that testosterone influences PCa, while having indicated that environmental factors such as diet and vitamin D deficiency may explain a large amount of the gap (Batai et al, 2017; but see Stranaland et al, 2017 for a contrary view). Since low vitamin D is related to prostate cancer, by supplementing with vitamin D, it is possible that levels of PCa may decrease. Kristal et al (2014) show that both high and low levels of vitamin D are associated with PCa.
Evidence also exists that vitamin D levels and hypertension are related. Rostand (2010) proposes a unified hypothesis: an important role exists in vitamin D deficiency and the pathogenesis and maintenance of hypertension in blacks (Rostand, 2010).
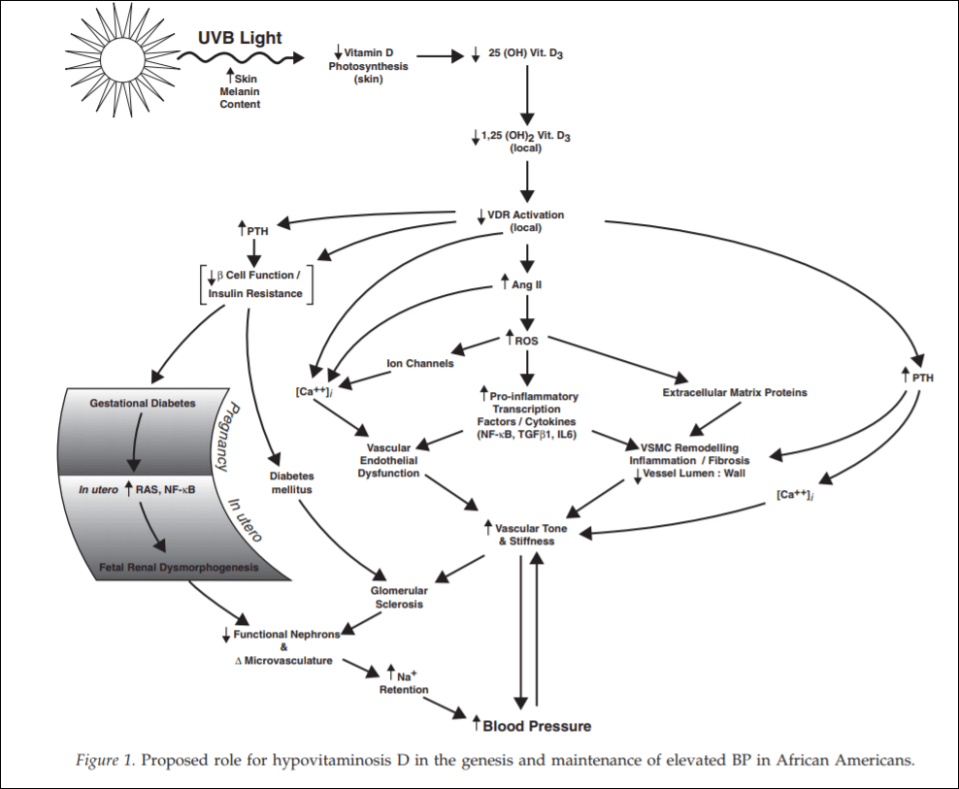
(From Rostand, 2010)
Since black Americans are no longer near the equator, their ability to synthesize vitamin D from UVB rays is diminished. This then probably leads the RAS (renin-angiotensin system) and inflammatory cytokine activation which then leads to vascular endothelial dysfunction along with structural changes to the microvasculature, which have been linked to vascular (arterial) stiffness along with increased vascular resistance, and these changes are shown to precede hypertension, which also occurs early in life. So since blacks are deficient in vitamin D, which even starts in the womb (Bodnar et al, 2007; Dawodu and Wagner, 2007; Lee et al, 2007; Khalessi et al, 2015; Seto et al, 2016), and this vitamin D deficiency most likely produces changes in large and small arteries and arterials, this could be the explanation for higher hypertension in black Americans (Rostand, 2010: 1701).
This would be a large environmental mismatch: since the population is displaced from its ancestral homeland, then this causes problems since it is not the environment where their ancestors evolved. So in this case, since black Americans are concentrated in the southeast corner of the United States, this may explain the high rates of vitamin D deficiency and hypertension in the black American community.
People whose ancestors evolved in locations with fewer UVB rays have lighter skin, whereas people whose ancestors evolved in locations with more UVB rays have darker skin. Thus, by placing populations in their opposite evolutionary environment, we can see how and why deleterious effects would occur in the population that is in the mismatched environment. For whites, skin cancer would occur, whereas for blacks, higher rates of hypertension and low birth weights occur.
Looking at levels of vitamin D deficiency in races is a great way to understand the evolution of certain populations. Because if the vitamin D hypothesis is correct, if skin color is an adaptation to UVB rays, with light skin being an adaptation to low UVB while dark skin is an adaptation to high UVB, then we can safely hypothesize about certain problems that would arise in races that are outside of their natural habitats. We have confirmed these hypotheses—black Americans who are outside of the location that their ancestors evolved in are more likely to have deleterious symptoms, and the symptoms are due to differences in vitamin D production, which come down to differences in skin color and how the skin synthesizes vitamin D in low-light environments.
Even though blacks have stronger bones than whites, this does not mean that they do not experience fractures at a high rate—especially children—and since the association was noticed, then by supplementing with vitamin D, this may lower the disparity of these types of injuries.
Since black Americans, compared to their evolutionary history, live in low-light environments, this then explains the how and why of vitamin D deficiency and why blacks need to supplement with vitamin D; no matter if certain studies show that blacks are ‘healthy’ even though they have low levels of vitamin D. If true (which I strongly doubt), that does not mean that black Americans should not supplement with vitamin D, because numerous other maladies are associated with vitamin D intake. This is one aspect where understanding the evolution of our species and the different races in it would lead to better medical care for individuals and ancestral groups that may need special treatment.
It is clear that race and geography should inform vitamin D intake, for if we do this, many diseases that arise can be ameliorated and quality of life can increase for everyone.
