
Biases and Political Beliefs
2150 words
The study of political bias is very important. Once the source of what motivates political bias—which no doubt would translate to other facets of life—is found, individual action can be taken to minimize any future bias. Two recent studies found that contrary to other studies showing that conservatives are more biased than liberals, both groups were equally as biased.
Everyone is biased—even physicians (Cain and Detsky, 2008). When beliefs we hold to be true are questioned, we do anything we can to shield ourselves from conflicting information. Numerous studies have looked into biases in politics, with some studies showing that conservatives are more likely to be biased towards their views more than liberals. However, recent research has shown that this is not true.
Frimer, Skitka, and Motyl, (2017) showed there were similar motives to shield one’s self from contradictory information. Hearing opposite viewpoints—especially for staunch conservatives and liberals—clearly leads to them doing anything possible to, in their heads, defend their dearly held beliefs. In four studies (1: people would forgo the chance to win money if they didn’t have to hear the opposite sides’ opinions on the same-sex marriage debate; 2: thinking back to the 2012 election; 3: upcoming elections in the US and Canada; “a range of other Culture War Issues” (Frimer, Skitka, and Motyle, 2017); and 4: both groups reported similar diversions towards hearing the opposite group’s beliefs), both groups reported that hearing the other side’s beliefs would induce cognitive dissonance (Frimer, Skitka, and Motyle, 2017). They meta-analyzed all of their studies and still found that both groups would “rather remain in their ideological bubbles”.
Ditto et al (2017) also had similar findings. They meta-analyzed 41 studies with over 12,000 participants, testing two hypotheses: 1) conservatives would be more biased than liberals and 2) there would be equal amounts of bias. They discovered that the correlation for partisan bias was “robust”, with a correlation of .254. They showed that “liberals (r = .248) and conservatives (r = .247) showed nearly identical levels of bias across studies” (Ditto et al, 2017).
These two studies show what we know is true: it’s extremely hard/damn near impossible to change one’s view. Someone can be dead wrong, yet attempt to gather up whatever kind of data they possibly can to shield themselves from the truth.
This all comes down to one thing: the backfire effect. When we are presented with contradictory information, we immediately reject it. Everyone is affected by this bias. One study showed that corrections frequently failed to correct political misconceptions, with these attempted corrections actually doing the opposite, people increased their misconception of the group in question (Nyhan and Riefler, 2010). The thing is, people lack the knowledge about political matters which then affects their opinions. These studies show why it’s next to impossible to change one’s view in regards to anything, especially political matters.
New York University’s Professor of Ethical Leadership and social psychologist with a specialty in morality Jonathan Haidt also talks about partisan bias in his outstanding book on religion and politics The Righteous Mind: Why Good People are Divided by Politics and Religion (Haidt, 2013). This book is outstanding and I highly recommend it. I’ve written about some of his thoughts in his book, his theory on the evolution of morality is very well argued. Moral reasoning is just a post-hoc search for reasons to justify the judgments that people have already made. When asked why people are so averse to questions they find morally wrong, they cannot give good reasons to why they find the scenarios morally wrong (Haidt, 2001). More specifically, people couldn’t say why it was morally wrong to have sex with a sibling even though they were told that they used birth control and both enjoyed the act, suffering no emotional damage. This is direct evidence for Haidt’s ‘wag-the-dog’ illusion.
Haidt (2001: 13) writes:
If moral reasoning is generally a post-hoc construction intended to justify automatic moral intuitions, then our moral life is plagued by two illusions. The first illusion can be called the “wag-the-dog” illusion: we believe that our own moral judgment (the dog) is driven by our own moral reasoning (the tail). The second illusion can be called the “wag-the-otherdog’s-tail” illusion: in a moral argument, we expect the successful rebuttal of an opponent’s arguments to change the opponent’s mind. Such a belief is like thinking that forcing a dog’s tail to wag by moving it with your hand should make the dog happy.
Except the opponent’s mind is never changed. People always search for things to affirm their worldviews.
In his book, Haidt cites a study done on 14 liberals and conservatives who were stuck into an fMRI machine to scan their brains when shown 18 slides to see how their brain changed when viewing them (Weston et al, 2006). The first of which slide one set was George W. Bush praiding Ken Lay, the CEO of Enron. After, they were shown a slide in which the former President avoided mentioning Lay’s name. “At this point, Republicans were squirming” (Haidt, 2013: 101). Then they were finally shown a slide that said that Bush “felt betrayed” by the CEO’s actions and was shocked to find out that he was corrupt. There was a set of similar slides showing similar contradictory statements from John Kerry. The researchers had engineered situations that made the individual uncomfortable when shown their candidate contradicted themselves, while at the same time not showing any signs of being uncomfortable when it was shown their ideological opposite was caught being a hypocrite (Haidt, 2013: 101).
This study shows that emotional and intuitive processes are the causes for such extreme biases, with one only employing reasoning when it supports their own conclusions. Weston et al (2006) saw that when the individuals looked at the final slides, they had a sense of ‘escape’ and ‘release’. They cite further studies showing that this sense of escape and release is associated with the release of dopamine in the nucleus accumbens and dorsal striatum in other animals (Weston et al, 2006). So the subjects experienced this small hit of dopamine when they saw the final slide that showed everything was “OK”. If this is true, then this explains why we engage in these ‘addictive behaviors’—believing things with such conviction, even when shown contradictory information.
Like rats that cannot stop pressing a button, partisans may be simply unable to stop believing weird things. The partisan brain has been reinforced so many times for performing mental contortions that free it from unwanted beliefs. Even partisanship may be literally addictive. (Haidt, 2013: 103)
Haidt has also been covering the recent University protests that have been occurring around the country. About fifty years ago, a judge predicted the political turmoil we see in Universities today, writing:
No one can be expected to accept an inferior status willingly. The black students, unable to compete on even terms in the study of law, inevitably will seek other means to achieve recognition and self-expression. This is likely to take two forms. First, agitation to change the environment from one in which they are unable to compete to one in which they can. Demands will be made for elimination of competition, reduction in standards of performance, adoption of courses of study which do not require intensive legal analysis, and recognition for academic credit of sociological activities which have only an indirect relationship to legal training. Second, it seems probable that this group will seek personal satisfaction and public recognition by aggressive conduct, which, although ostensibly directed at external injustices and problems, will in fact be primarily motivated by the psychological needs of the members of the group to overcome feelings of inferiority caused by lack of success in their studies. Since the common denominator of the group of students with lower qualifications is one of race this aggressive expression will undoubtedly take the form of racial demands–the employment of faculty on the basis of race, a marking system based on race, the establishment of a black curriculum and a black law journal, an increase in black financial aid, and a rule against expulsion of black students who fail to satisfy minimum academic standards.
This seems to have come true today, seeing as political diversity has decreased in psychology, for instance, in the past fifty years (Duarte et al, 2015). In America, they found that 58-66 percent of social science professors identified as liberals, whereas only 5-8 percent identified as conservatives. Self-identified Democrats also outnumbered Conservatives by almost 8 to 1. Other researchers found that 52 to 77 percent of humanities professors were liberal with only 4-8 percent identifying as Conservative, for a ratio of about 5 to 1, favoring liberals. Finally, 84 percent of psychologists identified as liberal, with only 8 percent identifying as conservative for a 10.5 to 1 ratio (Duarte et al, 2015). However, this skew has only existed for about fifty years. When our institutions show this heavy skew in political beliefs, self-affirming, self-fulfilling prophecies will affect the quality of what is taught to students which will have a negative effect on the type of education received.
Finally, when talking about political biases, one cannot go without mentioning Stephen Jay Gould. Although I’ve come to love his work on evolutionary theory, he was horribly wrong on human differences and let his motivations, biases and political views cloud his judgement and drive him to be grossly dishonest in his posthumous attacks of a man long dead who could no longer defend himself in one Samuel Morton, which first appeared in 1978. This culminated in his widely acclaimed (and, as fas as I can tell, still given to college students to read) book Mismeasure of Man (Gould, 1981). In the book, he attacked Morton for being biased in his measurements of his skull collection. However, in 2011, an anthropology team lead by Jason Lewis remeasured Morton’s skulls and found that Morton was not biased and his measurements were correct (Lewis et al, 2011). Gould was the one who ended up showing the huge bias that he accused Morton of and, ironically for Gould, he was the case study in avoiding bias in scholarship and science, not Morton.
However, as is usually the case, long debates such as this are not so easily settled. Philosopher Michael Weisberg (Weisberg, 2014) argued that Gould’s arguments against Morton were sound and that “Although Gould made some errors and overstated his case in a number of places, he provided prima facie evidence, as yet unrefuted, that Morton did indeed mismeasure his skulls in ways that conformed to 19th century racial biases.” Further, Kaplan, Pigliucci and Banta (2015) argue that Gould’s problem with Morton’s measurements came down to how the measurements should have been done (lead shot or seed). They contend that many of Lewis et al’s (2011) claims against Gould were “misleading” and “had no relevance to Gould’s published analysis.” They also argue that both Gould’s and Morton’s methods (inclusion/exclusion of skulls, how to compute averages, etc) were “inappropriate”. Nevertheless, the point is, this debate seems to be far from over and I await the next chapter. Whatever the case may be, Gould vs. Morton is a perfect case of politics and bias in science.
Everyone is biased. Researchers, physicians, normal everyday people, etc. But where we become most biased is when politics comes into play. To become better, well-rounded people with a myriad of knowledge, we need to listen to other’s viewpoints without immediately rejecting them. But, first, we must recognize the cognitive bias and attempt to correct it. Political differences begin in the brain and then are shaped by experience. These political differences then lead to feelings of disgust when hearing of the views of the ‘opposite team’. Both sides of the political spectrum are equally as biased, contrary to each groups’ perception of this particular issue. There are differences in the brain between Conservatives and Liberals, and when they see their ‘enemy’ engage in contradictory behavior they get joy, whereas when they see their guy engage in the same contradictory behavior they show disgust.
The long debate on Morton’s skulls that’s been raging for over forty years is the perfect look into how politics, motivation, and bias comes into effect in science, no matter which camp ultimately ends up being right (I’m in the Morton camp, obviously). Studying the causes and effects of why we have such strong biases can lead to a better understanding of the causes of these underlying defense mechanisms—the causes of the backfire effect and similar cognitive biases. Everyone and anyone—from the scientist to the layman—should always let what the facts say guide their points of view and not their emotions.
When you are studying any matter, or considering any philosophy, ask yourself only what are the facts and what is the truth that the facts bear out. Never let yourself be diverted either by what you wish to believe, or by what you think would have beneficent social effects if it were believed. But look only, and solely, at what are the facts. That is the intellectual thing that I should wish to say. —Bertrand Russel, 1959
Chewing the Cud
by Scott Jameson
RaceRealist and I have been ruminating on a lot of stuff lately. Here’s a fun one: what economic system works best relative to what we know about human health? In my mind there are two approaches: the libertarian approach, and quasi-fascism.
In the libertarian approach, there’s no regulation of sugar placed in our food. That’s already the case. But here’s an improvement: you don’t have to pay for anyone’s gastric bypass after they overeat that sugar.
In the fascist approach, there is regulation of sugar, because a fascist state does not allow people to poison each other for profit. You still have to pay for others’ medical expenses, but those expenses will be lower.
Here’s an advantage to the libertarian approach. In that society, the people who stuff their faces and refuse to get off the couch- who are dumber and lazier on average, probably- will have a higher mortality rate on average. Eugenics need not cost a dime.
But you run into a snag, sand in the gears of your hands-off system, when Big Food kicks out a whole bunch of crappy dietary advice, at which point a minority of reasonably intelligent people will be led astray, perhaps to the grave. How could a libertarian society stop that from taking place? Would it even bother? Could the system broadly work in spite of this snag?
A libertarian society doesn’t pay for idiots to have children. That’s good, but half of your population (women) are unlikely to ever support it. Women don’t do libertarianism; observe Rand Paul’s demographic Achilles Heel on page 25. When women asked men what to do about so-and-so’s eighth unpaid for child, we’d have to look them in the eyes and give a deadpan “let’s hope private charity can handle it.” There was a time, before FDR, when women would’ve accepted that answer. They were still in the kitchen back then, and I don’t know how to put them back there.
A fascist society has more hands-on eugenics, possibly genome editing or embryo selection. Also good. Expensive, but obviously worth it.
We welcome your input on these issues.
*****
As an aside, White men are well-known as the most conservative, small government, nationalist group out there in our current political atmosphere. I always hear people spewing the schmaltziest nonsense about the values of the Founding Fathers. They were, relative to our political compass, nationalist libertarians. Accordingly, modern nationalists and libertarians do best with the exact same demographics that used to vote on candidates back then: property-owning White men. The sole reason that Ron and Rand Paul couldn’t get elected is that they are too similar to the Founding Fathers. Any other candidate who blathers on about the Founding values is simply a liar, and their obvious lies show a disrespect of your intelligence.
If you’re a libertarian, but not an ethno-nationalistic and patriarchal thinker, then you simply haven’t gotten the memo: women and minorities do not want to create the same world that you do, nor will they ever. Evolution gave us women who want social safety nets and other races which are better off if they parasitize off of your tax dollars. All of the most libertarian societies that ever existed (early US, ancient Athens, Roman Republic) were entirely run by White men, and adding women to the electorate gave us the welfare state. Aristophanes was right.
*****
We’re also ruminating on the difference between IQ and expertise. I know of no mentally complicated task of which one can be a master without being intelligent. Take the IQs of chess grandmasters and you will find no morons.
Contrast that with purely physical activities. I bet you there are some really stupid people out there who are great at dancing for example. A prodigiously capable cerebellum may not predict an equally capable frontal lobe.
Discounting tasks which exclusively require things like simple physical coordination, muscle memory, etc, I ought to think that IQ is the biggest component of expertise.
The Evolutionary Puzzle of Floresiensis
1600 words
Back in October, I wrote that floresiensis is either descended from Erectus or habilis, since those were the only two hominins in the region. Yesterday a study was published titled The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters (Argue et al, 2017), in which Argue et al argue that floresiensis was not descended from a shipwrecked Erectus, as is a popular view. Another theory is that floresiensis is descended from habilis. The third theory is that floresiensis is Homo sapiens with a pathology, but that has been disproven (Falk et al, 2009). It was commonly thought that the LB1 floresiensis specimen was a pathological human inflicted with Laron syndrome which is a type of growth hormone deficiency (Laron and Klinger, 1994).
Argue et al found that floresiensis and erectus had completely different bone structures, particularly in the pelvis and jaw. They now believe that the theory that floresiensis is a derived form of an erectus that swam or rafted to Flores has been defintively refuted. They found that floresiensis was a sister species to habilis. So either a common ancestor of floresiensis or habilis swam to Flores from Africa, or floresiensis evolved in Africa and swam to Flores. They used new phylogenetic techniques to ascertain that floresiensis is stil a part of our lineage, but shows no phylogenetic relationship to erectus on the tree.
According to Baab (2016), biogeography shows that Indonesian erectus is the best fit with what is currently known. She says if floresiensis was derived from erectus that it “implies some degree of body size reduction and more marked brain size reduction.”
Kubo, Kono, and Kaifu (2013) conclude that the evolution of floresiensis from early Javanese erectus is possible when comparing the brain cases of both specimens. However, if floresiensis descended from habilis, then the brain size reduction wouldn’t be as marked (and is still due to island dwarfism, just not on as large of a scale as it would be if floresiensis were descended from erectus). The LB1 specimen also shows the closest neural affinities to early Asian erectus (Baab, Mcnulty, and Harvati, 2013; but see Vannuci, Barron, and Holloway, 2013 for the microcephalic view). Weston and Lister, (2009) showed that there was a 30 percent reduction in brain size in Magalasy hippos, which lends credence to the insular dwarfism hypothesis for floresiensis. Craniofacial morphology also shows that floresiensis evolved from Asian erectus (Kaifu et al, 2011).
The teeth of unknown hominin found at Mata Menge are intermediate between floresiensis and erectus, being 600,000 years older than where floresiensis was found (van den Bergh et al, 2016). This lends credence to the hypothesis that floresiensis is derived from erectus. Furthermore, insular dwarfism is seen in primate species isolated on islands, with changes in body size seen in child populations even on large islands not far from the mainland (Bromham and Cardillo, 2007, Welch, 2009). Genetically isolated on islands, primates can become bigger if the parent population was smaller, or smaller if the parent population was bigger. This is due to differing energy demands relative to the parent population, along with differing predators/prey.
The island rule even holds in the deep sea. As is the case with islands, the deep sea is also associated with decreased food availability. Looking at several species of gastropods, McClain, Boyer, and Rosenberg (2006) found that the island rule held in small-bodied shallow species. They were found to have larger bodied deep-sea representatives, with the same being true for large bodied deep-sea gastropods. Further, island dwarfism in elephants on the islands Sicily, Malta, Cyprus; mammoths on the California channel islands; and red deer on the island Jersey involved body mass changes of 5- to 100-fold over 2,300 to 120,000 generations (Evans et al, 2012).
So the overall hypothesis that island dwarfism is still intact, albeit if floresiensis is derived from habilis, the reduction in brain/body size would be smaller than if floresiensis evolved from early Asian erectus.
Further evidence for brain/body size reduction due to less food availability is noted by Daniel Lieberman in his book The Story of the Human Body: Evolution, Health, and Disease (Lieberman, 2013). While talking about the evolution of floresiensis on page 123 he writes:
The same energetic constraints and processes also affect hunter-gatherers . 62
And in the 62nd footnote on page 391 he writes:
Several human “pygmy” populations (people whose height does not exceed 150 centimeters, or 4.9 feet) have evolved in energy limited places like rain forests or islands. Perhaps the small size of the Dmansi hominins from Georgia also reflected selection to save energy among the first colonists of Eurasia.
Either way, if floresiensis evolved from erectus or habilis, considerable reductions in brain size have to be explained, since the smallest erectus brain ever found is 600 cubic centimeters while the smallest habilis brain ever found is 510 cubic centimeters (Lieberman, 2013: 124), with floresiensis having a brain 417 cubic centimeters (Falk et al, 2007).
What is most important about the insular dwarfism hypothesis in regards to the evolution of floresiensis is the effect of energy reduction/food availability and quality in regards to populations isolated on islands from parent populations. Floresiensis was able to survive on about 1200 kcal by shrinking, needing to consume about 1400 kcal during lactation compared to 1800 kcal for an erectus female who needed about 2500 kcal during lactation (Lieberman, 2013: 125). The cognitive price for the reduction in the brain size of floresiensis is not known, but since brains are so energy expensive (Aiello and Wheeler, 1995; Herculano-Houzel and Kaas, 2011; Fonseca-Azevedo and Herculano-Houzel, 2012), the reduction seen in floresiensis is no surprise.
Energy is one of the most important drivers for the evolution of a species, the evolution of floresiensis is one major example of this. Whether floresiensis evolved from habilis or erectus, reduced energy on the island caused the brain and body size of floresiensis to get smaller to cope with fewer things to eat. Keep in mind that habilis was a meat-eater as well, and with lower-quality energy on the island, the brain would have to reduce in size as it’s one of the most expensive organs in the body. As I’ve been saying for a long time now, the quality of energy is most important to the evolution of a species—especially Man. Cooking was imperative to our evolution, and with a lower-quality diet, we, too, would evolve smaller brains and bodies to compensate for reduced energy consumption since our brains take 25 percent of our daily energy requirements to power despite being 2 percent of our overall body weight.
The evolution of floresiensis shows how important energy is in the evolution of species. Its biggest implication—no matter if floresiensis evolved from habilis or erectus—is how important diet quality is to evolution, as I’ve noted here, here, here, here, here, and here. Without our high-quality diet, we, too, would suffer the same body/brain size reductions that floresiensis did.
References
Aiello, L. C., & Wheeler, P. (1995). The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution. Current Anthropology,36(2), 199-221. doi:10.1086/204350
Argue, D., Groves, C. P., Lee, M. S., & Jungers, W. L. (2017). The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters. Journal of Human Evolution. doi:10.1016/j.jhevol.2017.02.006
Baab, K. L., Mcnulty, K. P., & Harvati, K. (2013). Homo floresiensis Contextualized: A Geometric Morphometric Comparative Analysis of Fossil and Pathological Human Samples. PLoS ONE,8(7). doi:10.1371/journal.pone.0069119
Baab, K.L. (2016). The place of Homo floresiensis in human evolution. Journal of Anthropological Sciences, 94, 5-18.
Bergh, G. D., Kaifu, Y., Kurniawan, I., Kono, R. T., Brumm, A., Setiyabudi, E., . . . Morwood, M. J. (2016). Homo floresiensis-like fossils from the early Middle Pleistocene of Flores. Nature,534(7606), 245-248. doi:10.1038/nature17999
Bromham, L., & Cardillo, M. (2007). Primates follow the ‘island rule’: implications for interpreting Homo floresiensis. Biology Letters,3(4), 398-400. doi:10.1098/rsbl.2007.0113
Evans AR, Jones D, Boyer AG, Brown JH, Costa DP, et al. (2012) The maximum rate of mammal evolution. Proc Natl Acad Sci USA 109: 4187–4190.
Falk, D., Hildebolt, C., Smith, K., Morwood, M. J., Sutikna, T., Jatmiko, … Prior, F. (2007). Brain shape in human microcephalics and Homo floresiensis. Proceedings of the National Academy of Sciences of the United States of America, 104(7), 2513–2518. http://doi.org/10.1073/pnas.0609185104
Falk, D., Hildebolt, C., Smith, K., Jungers, W., Larson, S., Morwood, M., . . . Prior, F. (2009). The type specimen (LB1) of Homo floresiensis did not have Laron Syndrome. American Journal of Physical Anthropology,140(1), 52-63. doi:10.1002/ajpa.21035
Fonseca-Azevedo, K., & Herculano-Houzel, S. (2012). Metabolic constraint imposes tradeoff between body size and number of brain neurons in human evolution. Proceedings of the National Academy of Sciences,109(45), 18571-18576. doi:10.1073/pnas.1206390109
Herculano-Houzel, S., & Kaas, J. H. (2011). Gorilla and Orangutan Brains Conform to the Primate Cellular Scaling Rules: Implications for Human Evolution. Brain, Behavior and Evolution, 77(1), 33–44. http://doi.org/10.1159/000322729
Kaifu, Y., Baba, H., Sutikna, T., Morwood, M. J., Kubo, D., Saptomo, E. W., . . . Djubiantono, T. (2011). Craniofacial morphology of Homo floresiensis: Description, taxonomic affinities, and evolutionary implication. Journal of Human Evolution,61(6), 644-682. doi:10.1016/j.jhevol.2011.08.008
Kubo, D., Kono, R. T., & Kaifu, Y. (2013). Brain size of Homo floresiensis and its evolutionary implications. Proceedings of the Royal Society B: Biological Sciences,280(1760), 20130338-20130338. doi:10.1098/rspb.2013.0338
Laron, Z., & Klinger, B. (1994). Laron Syndrome: Clinical Features, Molecular Pathology and Treatment. Hormone Research,42(4-5), 198-202. doi:10.1159/00018419
3
Lieberman, D. (2013). The Story of the human body – evolution, health and disease. Penguin.
Mcclain, C. R., Boyer, A. G., & Rosenberg, G. (2006). The island rule and the evolution of body size in the deep sea. Journal of Biogeography,33(9), 1578-1584. doi:10.1111/j.1365-2699.2006.01545.x
Vannucci, R. C., Barron, T. F., & Holloway, R. L. (2011). Craniometric ratios of microcephaly and LB1, Homo floresiensis, using MRI and endocasts. Proceedings of the National Academy of Sciences of the United States of America, 108(34), 14043–14048. http://doi.org/10.1073/pnas.1105585108
Welch, J. J. (2009). Testing the island rule: primates as a case study. Proceedings of the Royal Society B: Biological Sciences,276(1657), 675-682. doi:10.1098/rspb.2008.1180
Weston, E. M., & Lister, A. M. (2009). Insular dwarfism in hippos and a model for brain size reduction in Homo floresiensis. Nature, 459(7243), 85–88. http://doi.org/10.1038/nature07922
Evolution and IQ Linkfest IV
1050 words
A decorated raven bone discovered in Crimea may provide insight into Neanderthal cognition (A raven bone discovered in Crimea shows two extra notches, which the researchers say they did intentionally to create a consistent—perhaps symbolic—pattern.)
Just like humans, artificial intelligence can be sexist and racist (Just like Tay. RIP in peace, Tay. Princeton University academics have shown, using a database of over 840 billion words from the Internet, that AI mimics humans and, they too, can apparently become ‘racist and sexist’, just like their creators. A “bundle of names” being associated with European Americans were more likely to be associated with pleasant terms, compared to African Americans who were associated with unpleasant terms. Male names were associated with careers while female names were associated with family. Seems like the AI just gets information and makes a conclusion based on the information, learning as it goes along. I don’t think putting human buzzwords onto AI is practical.)
Childhood lead exposure linked to lower adult IQ (This has been known for, literally, decades. In the new study, every 5 micrograms/dl increase in blood lead levels early in life was shown to decrease IQ by 1.61 points by the time the subjects reached 38 years of age along with reductions in working memory and perceptual reasoning. More shockingly, children who had blood lead levels over 10 micrograms/dl had IQs 4.25 points lower than their peers with low blood lead levels. Some research has shown that for each decrease in 1 point decrease in IQ annual salary decreases by about 200 to 600 dollars. This also shows that high blood-lead levels during childhood cannot be recovered from. There is currently an epidemic of lead poisoning in the Detroit Metropolitan area affecting Arab and African American children (Nriagu et al, 2014), as well as in low-income blacks in Saginaw, Michigan (Smith and Nriagu, 2010) and immigrant children in NYC (Tehranifar et al, 2008). However, see Koller et al (2004) for a different view, showing that blood-lead levels during childhood only account for 4 percent of the variation in cognition whereas parental and social factors account for more than 40 percent. High blood lead levels also lead to an increase in detrimental behavior (Sciarillo, Alexander, and Farrell, 1992). Clearly, measures need to be taken to reduce lead in water around the country as quality of life will improve for everyone.)
Molecular clocks track human evolution (Scientists estimate that we emerged around 200kya in Africa and spread throughout the world 50-100 kya through looking at the ‘molecular clock’—mutation and recombination. Some geneticists estimate that there are between 1.5 and 2 million mutational changes between humans and Neanderthals—estimating a splitting date at 550-750kya.)
How Ants Figured Out Farming Millions of Years Before Humans (Ants have probably been farming since the Chicxulub meteor impact that killed the dinosaurs in one South American rainforest. The researchers found that all fungus-farming ants came from the same ancestor around 60mya. Thirty million years later, another farming species emerged. One of the species was a “more complex agriculturalist” which probably transported its fungus to different locations while the other species were “lower, less complex agriculturalists” which grew fungus capable of “escaping its garden and living independently”. The dry climate allowed the more complex agriculturalists to domesticate the fungus and control the temperature by digging holes for storage.
Mimicking an impact on Earth’s early atmosphere yields all 4 RNA bases (Researchers showed that mimicking an asteroid crash resulted in the formation of the production of all four bases of RNA, the molecules essential for life. Take that, Creationists.)
Study finds some significant differences in brains of men and women (Adjusting for age, researchers found that women had significantly thicker cortices than men while men had higher brain volumes in every area they looked at, including the hippocampus (which plays a part in memory and spatial awareness), the amygdala (plays a role in memory, emotions, and decision-making), striatum (learning, inhibition, and reward-processing), and the thalamus (processing and relaying sensory information to other parts of the body). Men also varied more in cortical thickness and volume much more than women, which supports other studies showing that men only have higher IQ distributions than women, not higher average intelligence. There are, of course, differences between the male and female brain, but I’m now rethinking my position on male/female IQ differences (leaning towards no). Read the preprint here: Sex Differences In The Adult Human Brain: Evidence From 5,216 UK Biobank Participants
Infants show racial bias toward members of own ethnicity, against those of others (Now the study I’ve seen a buzz about the past few days. In one of the studies, researchers found that babies associated other races’ faces with sad music while associating their own races’ face with upbeat, happy music. The second study showed that infants were more likely to learn from their own race than another, which relied on gaze cues. Both studies tested infants between the ages of 6 to 9 months with both studies finding no racial bias in infants less than 6 months of age. The researchers, of course, conclude that this occurs because of lack of contact with other race babies/people. In study one, infants aged 3 to 10 months watched videos with a female adult with a neutral facial expression. The babies heard music before viewing each clip; happy music and same race face; happy music and opposite race face; sad music and same race face; and sad music and opposite race face. They found that babies looked longer at same-race faces with happy music while looking longer at other-race faces with sad music. The second study looked at whether or not babies would be willing to learn from their own race or a different race. Babies aged 6 to 9 months were shown a video in which a female adult appeared and looked at one of the four corners of the screen. In some of the videos, an animal appeared where the person on the screen looked while in other videos, an animal appeared at a non-looked at location. They showed that babies followed the gaze of their own race more often than the other race, showing that infants are “biased” towards learning from their own race. This is clearly evidence for Genetic Similarity Theory. Expect a research article on this soon. Read the abstracts here: Infants Rely More on Gaze Cues From Own-Race Than Other-Race Adults for Learning Under Uncertainty and Older but not younger infants associate own-race faces with happy music and other-race faces with sad music)
Race, Testosterone, and Honor Culture
2300 words
Misinformation about testosterone and strength in regards to race is rampant in the HBD-o-sphere. One of the most oft-repeated phrases is that “Blacks have higher levels of testosterone than whites”, even after controlling for numerous confounds. However, the people who believe this literally only cite one singular study with 50 blacks and 50 whites. Looking at more robust data with higher ns shows a completely different story. Tonight I will, again, go through the race/testosterone conundrum (again).
Type I fibers fire first when heavy lifting. Whites have more type I fibers. Powerlifters and Olympic lifters have a greater amount type IIa fibers, with fewer type IIx fibers (like whites). This explains why blacks are hardly represented in powerlifting and strongman competitions.
Somatype, too, also plays a role. Whites are more endo than blacks who are more meso. Endomorphic individuals are stronger, on average, than mesomorphic and ectomorphic individuals.
Blacks have narrower hips and pelves. This morphological trait further explains why blacks dominate sports. Some people may attempt to pick out one variable that I speak about (fiber type, morphology, somatype, fat mass, etc) and attempt to disprove it, thinking that disproving that variable will discredit my whole argument. However, fiber typing is set by the second trimester, with no change in fiber type from age 6 to adulthood (Bell et al, 1980).
It is commonly believed that blacks have higher levels of testosterone than whites. However, this claim is literally based off of one study (Ross et al, 1986) when other studies have shown low to no difference in T levels (Richards et al, 1992; Gapstur et al, 2002; Rohrmann et al, 2007; Mazur, 2009; Lopez et al, 2013; Richard et al 2014). People who still push the “blacks-have-higher-T-card” in the face of this evidence are, clearly, ideologues who want to cushion their beliefs when presented with contradictory evidence (Nyhan and Reifler, 2010).
‘Honor Culture’ and testosterone
In all of my articles on this subject, I have stated—extensively—that testosterone is mediated by the environment. That is, certain social situations can increase testosterone. This is a viewpoint that I’ve emphatically stated. I came across a paper while back that talks about a sociological perspective (I have huge problems with social ‘science’, [more on that soon] but this study was very well done) in regards to the testosterone difference between blacks and whites.
Some people when they read this, however, may go immediately to the part of the paper that says what they want it to say without fully assessing the paper. In this section, I will explain the paper and how it confirms my assertions/arguments.
Mazur (2016) begins the paper talking about ‘honor culture‘, which is a culture where people avoid intentionally offending others while also maintaining a status for not backing down from a confrontation. This theory was proposed by Richard Nisbett in 1993 to explain why the South had higher rates of violence—particularly the Scotch-Irish.
However parsimonious the theory may sound, despite its outstanding explanatory power, it doesn’t hold while analyzing white male homicides in the South. It also doesn’t hold analyzing within-county homicide rates either, since apparently poverty better explains higher homicide rates.
But let’s assume it’s true for blacks. Let’s assume the contention to be true that there is an ‘honor culture’ that people take part in.
Young black men with no education had higher levels of testosterone than educated whites and blacks. Looking at this at face value—literally going right to the section of the paper that says that poor blacks had higher testosterone, nearly 100 ng/ml higher than the mean testosterone of whites. As Mazur (2016) notes, this contradicts his earlier 2009 study in which he found no difference in testosterone between the races.
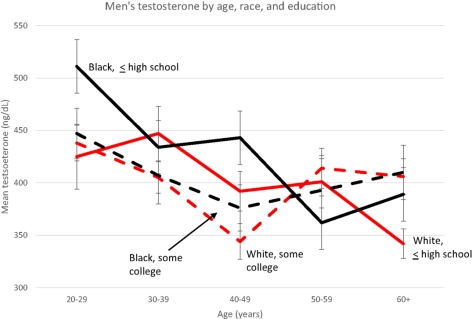
Note the low testosterone for both races at age 20-29—ranging from about 515 to 425—why such low testosterone levels for young men? Anyway, the cause for the higher levels is due to the type of honor culture that blacks participate in, according to Mazur (which is consistent with the data showing that testosterone rises during conflict/aggressive situations).
Mazur cites Elijah Anderson, saying that most youths have a “code of the streets” they take part in, which have to do with interpersonal communication such as “gait and verbal expressions” to deter aggressive behavior.
Testosterone is not a causal variable in regards to violent behavior. But it does rise during conflicts with others, watching a favorite sports team, asserting dominance, and even how you carry yourself (especially your posture). Since low-class blacks participate in these types of behaviors, then they would have higher levels of testosterone due to needing to “keep their status.”
When testosterone rises in these situations, it increases the response threat in mens’ brains, most notably showing increased activity in the amygdala. Further, dominant behavior and posture also increase testosterone levels. Putting this all together, since blacks with only a high school education have higher testosterone levels and are more likely to participate in honor culture compared to whites and blacks with higher educational achievement, then they would have higher testosterone levels than whites and blacks with a high school education who do not participate in honor culture.
Further, as contrary to what I have written in the past (and have since rescinded), there is no indication of higher testosterone levels in black women with low education. It seems this ‘honor culture’ effect on testosterone only holds for black men with only a high school education.
Mazur’s (2016) most significant finding was that black men aged 20-29 with only a high school education had 91 ng/ml higher testosterone than whites. Among older and/or educated men, testosterone did not vary. This indicates that since they have attained higher levels of educational success, there is no need to participate in ‘honor culture’.
This is yet further evidence for my assertion that environmental variables such as posture, dominance, and aggressive behavior raise testosterone levels.
The honor culture hypothesis is found to hold in Brazil in a comparative study of 160 inmates and non-inmates (De Souza et al, 2016). As Mazur (2016) notes, the honor culture hypothesis could explain the high murder rate for black Americans—the need to ‘keep their status’. It’s important to note that this increase in testosterone was not noticed in teenage or female blacks (because they don’t participate in honor culture).
There is a perfectly good environmental—not genetic—reason for this increase in testosterone in young blacks with only a high school education. Now that we know this, back to race and strength.
Mazur (2009) found that black men in the age range of 20-69, they averaged .39 ng/ml higher testosterone than whites, which is partly explained by lower marriage rates and low adiposity. White men are more likely to be obese than black men, since black men with more African ancestry are less likely to be obese. When controlling for BMI, blacks are found to have 2.5-4.9 percent more testosterone than whites (Gapstur et al, 2002, Rohrmann et al, 2007, Richard et al, 2014). There is little evidence for the assertion that blacks have higher levels of testosterone without environmental triggers.
Blacks between the age of 12 and 15 average lower levels of testosterone than whites. However, after the age of 15, “testosterone levels increase rapidly” with blacks having higher peak levels than whites (seen in table 2 below). After adjusting for the usual confounds (BMI, smoking, age, physical activity, and waist circumference), blacks still had higher levels of testosterone—which is attributed to higher levels of lean mass.
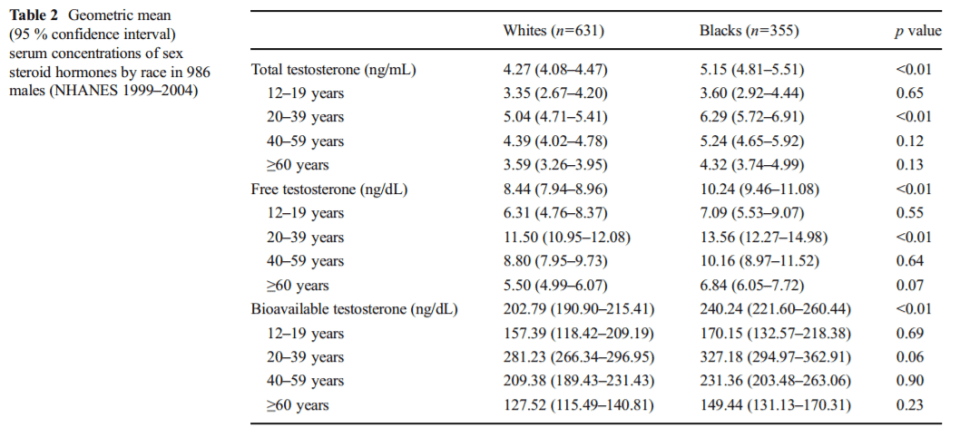
As seen above in table 2 from Hu et al (2014), the difference in total testosterone between blacks and whites aged 20-39 was 6.29 ng/ml and 5.04 ng/ml respectively, with free testosterone for whites being 11.50 and 13.56 for blacks and finally bioavailable testosterone for whites and blacks aged 20-39 was 281.23 and 327.18 ng/ml respectively. These small differences in testosterone cannot account for racial disparities in violence nor prostate cancer—since there is no relationship between prostate cancer and testosterone (Stattin et al, 2003; Michaud, Billups, and Partin, 2015).
In regards to Africans, the best studies I can find comparing some African countries with the West study salivary testosterone. However, there is a direct correlation between salivary testosterone and free serum testosterone (Wang et al, 1981; Johnson, Joplin, and Burrin, 1987). Of the studies I could find, Kenyan pastoralists called the Ariaal have lower levels of testosterone than Western men (Campbell, O’Rourke, and Lipson, 2003; Campbell, Gray, and Ellison, 2006) while men in Zimbabwe had levels “much lower” compared to Western populations (Lukas, Campbell, and Ellison, 2004). Lastly, among men aged 15 to 30, salivary testosterone levels in an American sample was 335 pmol//l compared to 286 pmol/l in men from the Congo (Elisson et al, 2002). Even certain African populations don’t have higher testosterone levels than Western peoples.
Conclusion
The meme that blacks have higher rates of testosterone in comparison to whites needs to be put to rest. This is only seen in blacks who participate in ‘honor culture’, which is an environmental variable. This is in contrast to people who believe that it is genetic in nature—environmental variables can and do drive hormones. Mazur (2016) is proof of that. Mazur (2016) also shows that the honor culture hypothesis doesn’t hold for teens or black males—so they don’t have elevated levels of testosterone. Certain studies of African populations, however, do not show higher levels of testosterone than Western populations.
Looking at the complete literature—rather than a select few studies— we can see that testosterone levels between white and black Americans are not as high as is commonly stated (Richards et al, 1992; Gapstur et al, 2002; Rohrmann et al, 2007; Mazur, 2009; Lopez et al, 2013; Hu et al, 2014; Richard et al, 2014). Further, even if blacks did have higher levels of testosterone than whites—across the board (sans honor culture), it still wouldn’t explain higher rates of black violence when compared to whites, nor would it explain higher prostate cancer rates (Stattin et al, 2003; Michaud, Billups, and Partin, 2015).
Only blacks with low educational achievement have higher levels of testosterone—which, even then is not enough to explain higher rates of violence or prostate cancer acquisition. Other factors explain the higher murder rate (i.e., honor culture, which increases testosterone, the environmental trigger matters first and foremost) and violent crime that blacks commit. But attempting to explain it with 30-year-old studies (Ross et al, 1986) and studies that show that environmental factors increase testosterone (Mazur, 2016) don’t lend credence to that hypothesis.
References
Bell, R. D., Macdougall, J. D., Billeter, R., & Howald, H. (1980). Muscle fiber types and morphometric analysis of skeletal muscle in six-year-old children. Medicine & Science in Sports & Exercise,12(1). doi:10.1249/00005768-198021000-00007
Campbell, B., O’rourke, M. T., & Lipson, S. F. (2003). Salivary testosterone and body composition among Ariaal males. American Journal of Human Biology,15(5), 697-708. doi:10.1002/ajhb.10203
Campbell, B. C., Gray, P. B., & Ellison, P. T. (2006). Age-related patterns of body composition and salivary testosterone among Ariaal men of Northern Kenya. Aging Clinical and Experimental Research,18(6), 470-476. doi:10.1007/bf03324846
De Souza, Souza, B. C., Bilsky, W., & Roazzi, A. (2016). The culture of honor as the best explanation for the high rates of criminal homicide in Pernambuco: A comparative study with 160 convicts and non-convicts. Anuario de Psicología Jurídica,26(1), 114-121. doi:10.1016/j.apj.2015.03.001
Ellison, P. T., Bribiescas, R. G., Bentley, G. R., Campbell, B. C., Lipson, S. F., Panter-Brick, C., & Hill, K. (2002). Population variation in age-related decline in male salivary testosterone. Human Reproduction,17(12), 3251-3253. doi:10.1093/humrep/17.12.3251
, , , , , . Serum androgen concentrations in young men: a longitudinal analysis of associations with age, obesity, and race—the CARDIA male hormone study. Cancer Epidemiol Biomarkers Prev 2002; 11: 1041–7
Hu, H., Odedina, F. T., Reams, R. R., Lissaker, C. T., & Xu, X. (2014). Racial Differences in Age-Related Variations of Testosterone Levels Among US Males: Potential Implications for Prostate Cancer and Personalized Medication. Journal of Racial and Ethnic Health Disparities,2(1), 69-76. doi:10.1007/s40615-014-0049-8
Johnson, S. G., Joplin, G. F., & Burrin, J. M. (1987). Direct assay for testosterone in saliva: Relationship with a direct serum free testosterone assay. Clinica Chimica Acta,163(3), 309-318. doi:10.1016/0009-8981(87)90249-x
Lopez, D. S., Peskoe, S. B., Joshu, C. E., Dobs, A., Feinleib, M., Kanarek, N., . . . Platz, E. A. (2013). Racial/ethnic differences in serum sex steroid hormone concentrations in US adolescent males. Cancer Causes & Control,24(4), 817-826. doi:10.1007/s10552-013-0154-8
Lukas, W. D., Campbell, B. C., & Ellison, P. T. (2004). Testosterone, aging, and body composition in men from Harare, Zimbabwe. American Journal of Human Biology,16(6), 704-712. doi:10.1002/ajhb.20083
Mazur, A. (2009). The age-testosterone relationship in black, white, and Mexican-American men, and reasons for ethnic differences. The Aging Male,12(2-3), 66-76. doi:10.1080/13685530903071802
Mazur, A. (2016). Testosterone Is High among Young Black Men with Little Education. Frontiers in Sociology,1. doi:10.3389/fsoc.2016.00001
Michaud, J. E., Billups, K. L., & Partin, A. W. (2015). Testosterone and prostate cancer: an evidence-based review of pathogenesis and oncologic risk. Therapeutic Advances in Urology,7(6), 378-387. doi:10.1177/1756287215597633
Nyhan, B., & Reifler, J. (2010). When Corrections Fail: The Persistence of Political Misperceptions. Political Behavior,32(2), 303-330. doi:10.1007/s11109-010-9112-2
Richard, A., Rohrmann, S., Zhang, L., Eichholzer, M., Basaria, S., Selvin, E., . . . Platz, E. A. (2014). Racial variation in sex steroid hormone concentration in black and white men: a meta-analysis. Andrology,2(3), 428-435. doi:10.1111/j.2047-2927.2014.00206.x
Richards, R. J., Svec, F., Bao, W., Srinivasan, S. R., & Berenson, G. S. (1992). Steroid hormones during puberty: racial (black-white) differences in androstenedione and estradiol–the Bogalusa Heart Study. The Journal of Clinical Endocrinology & Metabolism,75(2), 624-631. doi:10.1210/jcem.75.2.1639961
Rohrmann, S., Nelson, W. G., Rifai, N., Brown, T. R., Dobs, A., Kanarek, N., . . . Platz, E. A. (2007). Serum Estrogen, But Not Testosterone, Levels Differ between Black and White Men in a Nationally Representative Sample of Americans. The Journal of Clinical Endocrinology & Metabolism,92(7), 2519-2525. doi:10.1210/jc.2007-0028
Ross R, Bernstein L, Judd H, Hanisch R, Pike M, Henderson B. Serum testosterone levels in healthy young black and white men. J Natl Cancer Inst. 1986 Jan;76(1):45–48
Stattin, P., Lumme, S., Tenkanen, L., Alfthan, H., Jellum, E., Hallmans, G., . . . Hakama, M. (2003). High levels of circulating testosterone are not associated with increased prostate cancer risk: A pooled prospective study. International Journal of Cancer,108(3), 418-424. doi:10.1002/ijc.11572
Wang, C., Plymate, S., Nieschlag, E., & Paulsen, C. A. (1981). Salivary Testosterone in Men: Further Evidence of a Direct Correlation with Free Serum Testosterone. The Journal of Clinical Endocrinology & Metabolism,53(5), 1021-1024. doi:10.1210/jcem-53-5-1021
Evolution Denial Part II
1450 words
Why do people deny evolution? Not just evolution from single-celled organisms to multicellular lifeforms, but human evolution as well? Most people who deny evolution don’t have the knowledge to assess it correctly. They fall back on the Bible and say “God did this, the Bible says…. God says…” all the while looking at you as a heathen when you attempt to talk some basic biology or, God forbid, the process of evolution.
I met a woman the other day and I asked her what she was studying in school. She tells me anatomy and physiology (right up my alley). So we start talking about some basic anatomy and physiology before I ask the question: “Do you believe in evolution?” She gave me a blank stare and said no.
“Humans as we know them have always existed in this form,” she said. I just started laughing at her ignorance and then she said “Evolution at the macro level is not possible but it is at the micro level”, repeating the same old and tired Creationist talking points. I said to her that there is no evidence for creation and that the evidence we do have points to evolution. I said that the theory of evolution has so much backing, so much evidence, that to believe otherwise you’d have to purposefully close your mind to the truth, to shut out any and all contradictory information.
One of the funniest things she said to me was that she wants to cure diseases. To that, I said if she wants to do that then she must look at diseases from an evolutionary perspective (Gluckman et al, 2011). She said that she doesn’t need to know how diseases were in the past, just how they are today. I also said that if she is studying anatomy and physiology then she must understand that many of our appendages are derived from our hominin ancestors, which began with Erectus as I’ve covered in my article Man the Athlete. Diseases also must be looked at through an evolutionary lens, so if anyone wants to cure diseases, then they must first understand and accept that things are constantly changing and evolving to better survive in that environment.
When I said that there is no evidence for Creation she got really mad. She said that there is no evidence that “we evolved from monkeys” which gave me a good laugh. Even people who believe in evolution still make that mistake of believing that we evolved from monkeys. One of the most common statements from Creationists is “If humans evolved from monkeys then why are monkeys still around?”, wrongly assuming that we literally evolved from monkeys, incorrectly misinterpreting that we share a common ancestor with monkeys 6-12 mya.
About 6mya, there was a chromosomal fusion on chromosome two; two ancestral ape chromosomes fused to make chromosome two (Idjo et al, 1991). That is some nice chromosomal evidence for common descent from our ape cousins. Creationists, however, purport that a gene in chromosome 2, DDX11L2, writing that the “alleged fusion site is not a degenerate fusion sequence but is and, since creation, has been a functional feature in an important gene.” Further, Tomkins’ claim that the fusion site is actually a gene is wrong since the fusion site is more than 1300 bases away from the gene.
The ancestral equivalents of chromosome 2—2p and 2q—fused together in a fusion event some 6mya. This precise fusion site is on chromosome 2 (Hellier et al, 2004). Creationists will say and do anything to attempt to ‘rebut’ this contention. Genetic evidence is the best evidence we have (due to Punctuated Equilibria, which causes the spottiness in the fossil record), and still, these ‘Creationist geneticists’ will do anything they can to attempt to have Evolutionists go on the defensive. However, the onus is on them to disprove the mountains of evidence.
One of the funniest things this woman said to me is that man has always been in this form and that we didn’t evolve from “monkeys”, which is when I said that it’s more complicated than that: we have fish ancestors, named Tiktaalik who had the beginnings of the human arm and hand, along with Pikaia Gracilens—our oldest ancestor. If Pikaia would have died out in the Cambrian explosion some 550 mya, we wouldn’t be here today. We are here today due to the happenstance of numerous accidents of history—contingencies of “just history” to quote Stephen Jay Gould.
Nevertheless, Creationists will always attempt to distort evolutionary science to fit their agendas. Stephen Jay Gould battled Creationists throughout his career. Creationists would quote mine his books to show that Evolutionists do show evidence of “Creation”. One of his most quote mined works is his and Eldredge’s theory of Punctuated Equilibria (1972). Just because a look at the whole fossil record shows species remaining in stasis for most of their history before a short burst of evolutionary change then that must mean that there was a guiding hand involved in the process. Here is a full list of quote mines that Creationists use from Eldredge and Gould.
As you can see, Creationists use any kind of mental gymnastics to disprove evolution. However, no matter how hard you try with Creationists, you can’t educate people into believing in evolution. This is mainly due to the backfire effect which occurs when you show people contradictory information to a dearly held belief and they frantically attempt to gather evidence to shield themselves from contradictory evidence (Nyhan and Reifler, 2010). This cognitive bias holds for more than political debates, though it’s most often seen there. Showing people any kind of contradictory information will have them search and search for anything to shield themselves from the truth. However, no amount of ‘information’ provided by Creationists will disprove evolutionary theory.
Gould and Eldredge aren’t the only Evolutionist that Creationists quote mine–one of the most famous quote mines is from Darwin’s The Descent of Man in which he talks about defending his theory from detractors, mainly the spottiness of the fossil record (which Eldredge and Gould’s Punctuated Equilibria explains). However, this doesn’t stop Creationists—and even some Evolutionists who fall for Creationist trickery—to believe that Darwin was talking about something completely different, in that Darwin was ‘racist’ talking about the ‘superior races’ exterminating the ‘inferior races’. Reading the quote in its entirety, however, shows something completely different. Alas, some people don’t care about facts antruthut and only care about their agenda they attempt to push.
Even setting evolutionary theory aside, basic geology disproves Creationism. The author of the piece, geologist David Montgomery, says that there is a rock outside of his office that proves Creationism wrong. The rock shows that there is more to the geologic record that could be explained by a single grand flood. Now that geologists now have the tools and data to infer that the earth is billions of years old—not thousands as Young Earth Creationists (YECs) claim—YECs change up their interpretation of the Creation story in Genesis to go from literal days to “days in Genesis refer to geological ages”. Clear mental gymnastics in the face of contradictory evidence.
There are five mass extinctions that are accepted in the scientific community (Jablonski, 2001) (though I am reading a book at the moment that talks about nine mass extinction events with Man pushing the tenth, I will return to this in the future). After these contingencies of ‘just history’, we can see that we are incredibly lucky that our ancestors did not die out. From a Pikaia Gracilens surviving the Cambrian radiation, to Tiktaalik and its venturing onto land from the sea and finally the survival of a shrew-like ancestor during the extinction of the dinosaurs, we should thank our lucky stars that these things went our way, because if not, I wouldn’t be sitting here writing this at the moment and you would not be reading this. Evolutionary history is littered with these events—events that, if they went the other way would not lead to the evolution of Man again.
In sum, people who do not believe in evolutionary theory clearly are emotionally invested in believing in a story of Creation—sans evidence, only their belief. On the other hand, evolutionists such as we have all the data on our side when it comes to this debate. Creationists have to use any kind of warped logic to not believe the mountains of evidence that have piled up since Darwin wrote On the Origin. However, as everyone knows, reality isn’t what just what you believe. Just because Creationists handwave away the data that people like us provide to them doesn’t mean that evolution isn’t true.
The Evolution of Human Skin Variation
4050 words
Human skin variation comes down to how much UV radiation a population is exposed to. Over time, this leads to changes in genetic expression. If that new genotype is advantageous in that environment, it will get selected for. To see how human skin variation evolved, we must first look to chimpanzees since they are our closest relative.
The evolution of black skin
Humans and chimps diverged around 6-12 mya. Since we share 99.8 percent of our genome with them, it’s safe to say that when we diverged, we had pale skin and a lot of fur on our bodies (Jablonski and Chaplin, 2000). After we lost the fur on our bodies, we were better able to thermoregulate, which then primed Erectus for running (Liberman, 2015). The advent of fur loss coincides with the appearance of sweat glands in Erectus, which would have been paramount for persistence hunting in the African savanna 1.9 mya, when a modern pelvis—and most likely a modern gluteus maximus—emerged in the fossil record (Lieberman et al, 2006). This sets the stage for one of the most important factors in regards to the ability to persistence hunt—mainly, the evolution of dark skin to protect against high amounts of UV radiation.
After Erectus lost his fur, the unforgiving UV radiation beamed down on him. Selection would have then occurred for darker skin, as darker skin protects against UV radiation. Dark skin in our genus also evolved between 1 and 2 mya. We know this since the melanocortin 1 receptor promoting black skin arose 1-2 mya, right around the time Erectus appeared and lost its fur (Lieberman, 2015).
However, other researchers reject Greaves’ explanation for skin cancer being a driver for skin color (Jablonksi and Chaplin, 2014). They cite Blum (1961) showing that skin cancer is acquired too late in life to have any kind of effect on reproductive success. Skin cancer rates in black Americans are low compared to white Americans in a survey from 1977-8 showing that 30 percent of blacks had basal cell carcinoma while 80 percent of whites did (Moon et al, 1987). This is some good evidence for Greaves’ hypothesis; that blacks have less of a rate of one type of skin cancer shows its adaptive benefits. Black skin evolved due to the need for protection from high levels of UVB radiation and skin cancers.
Highly melanized skin also protects against folate destruction (Jablonksi and Chaplin, 2000). As populations move away from high UV areas, the selective constraint to maintain high levels of folate by blocking high levels of UV is removed, whereas selection for less melanin prevails to allow enough radiation to synthesize vitamin D. Black skin is important near the equator to protect against folate deficiency. (Also see Nina Jablonski’s Ted Talk Skin color is an illusion.)
The evolution of white skin
The evolution of white skin, of course, is much debated as well. Theories range from sexual selection, to diet, to less UV radiation. All three have great explanatory power, and I believe that all of them did drive the evolution of white skin, but with different percentages.
The main driver of white skin is living in colder environments with fewer UV rays. The body needs to synthesize vitamin D, so the only way this would occur in areas with low UV rays.
White skin is a recent trait in humans, appearing only 8kya. A myriad of theories have been proposed to explain this, from sexual selection (Frost, 2007), which include better vitamin D synthesis to ensure more calcium for pregnancy and lactation (which would then benefit the intelligence of the babes) (Jablonski and Chaplin, 2000); others see light skin as the beginnings of more childlike traits such as smoother skin, a higher pitched voice and a more childlike face which would then facilitate less aggressiveness in men and more provisioning (Guthrie, 1970; from Frost, 2007); finally, van den Berghe and Frost (1986) proposed that selection for white skin involved unconscious selection by men for lighter-skinned women which is used “as a measure of hormonal status and thus childbearing potential” (Frost, 2007). The three aforementioned hypotheses have sexual selection for lighter skin as a proximate cause, but the ultimate cause is something completely different.
The hypothesis that white skin evolved to better facilitate vitamin D synthesis to ensure more calcium for pregnancy and lactation makes the most sense. Darker-skinned individuals have a myriad of health problems outside of their ancestral climate, one of which is higher rates of prostate cancer due to lack of vitamin D. If darker skin is a problem in cooler climates with fewer UV rays, then lighter skin, since it ensures better vitamin D synthesis, will be selected for. White skin ensures better and more vitamin D absorption in colder climates with fewer UV rays, therefore, the ultimate cause of the evolution of white skin is a lack of sunlight and therefore fewer UV rays. This is because white skin absorbs more UV rays which is better vitamin D synthesis.
Peter Frost believes that Europeans became white 11,000 years ago. However, as shown above, white skin evolved around 8kya. Further, contrary to popular belief, Europeans did not gain the alleles for white skin from Neanderthals (Beleza et al, 2012). European populations did not lose their dark skin immediately upon entering Europe—and Neanderthal interbreeding didn’t immediately confer the advantageous white skin alleles. There was interbreeding between AMH and Neanderthals (Sankararaman et al, 2014). So if interbreeding with Neanderthals didn’t infer white skin to proto-Europeans, then what did?
A few alleles spreading into Europe that only reached fixation a few thousand years ago. White skin is a relatively recent trait in Man (Beleza et al, 2012). People assume that white skin has been around for a long time, and that Europeans 40,000 ya are the ancestors of Europeans alive today. That, however, is not true. Modern-day European genetic history began about 6,500 ya. That is when the modern-day European phenotype arose—along with white skin.
Furthermore, Eurasians were still a single breeding population 40 kya, and only diverged recently, about 25,000 to 40,000 ya (Tateno et al, 2014). The alleles that code for light skin evolved after the Eurasian divergence. Polymorphisms in the genes ASIP and OCA2 may code for dark and light skin all throughout the world, whereas SLC24A5, MATP, and TYR have a predominant role in the evolution of light skin in Europeans but not East Asians, which suggests recent convergent evolution of a lighter pigmentation phenotype in European and East Asian populations (Norton et al, 2006). Since SLC24A5, MATP, and TYR are absent in East Asian populations, then that means that East Asians evolved light skin through completely different mechanisms than Europeans. So after the divergence of East Asians and Europeans from a single breeding population 25-40kya, there was convergent evolution for light pigmentation in both populations with the same selection pressure (low UV).
Some populations, such as Arctic peoples, don’t have the skin color one would predict they should have based on their ancestral environment. However, their diets are high in shellfish which is high in vitamin D, which means they can afford to remain darker-skinned in low UV areas. UV rays reflect off of the snow and ice in the summer and their dark skin protects them from UV light.
Black-white differences in UV absorption
If white skin evolved to better synthesize vitamin D with fewer (and less intense) UV rays, then those with blacker skin would need to spend a longer time in UV light to synthesize the same amount of vitamin D. Skin pigmentation, however, is negatively correlated with vitamin D synthesis (Libon, Cavalier, and Nikkels, 2013). Black skin is less capable of vitamin D synthesis. Furthermore, blacks’ skin color leads to an evolutionary environmental mismatch. Black skin in low UV areas is correlated with rickets (Holick, 2006), higher rates of prostate cancer due to lower levels of vitamin D (Gupta et al, 2009; vitamin D supplements may also keep low-grade prostate cancer at bay).
Libon, Cavalier, and Nikkels, (2013) looked at a few different phototypes (skin colors) of black and white subjects. The phototypes they looked at were II (n=19), III (n=1), and VI (n-11; whites and blacks respectively). Phototypes are shown in the image below.
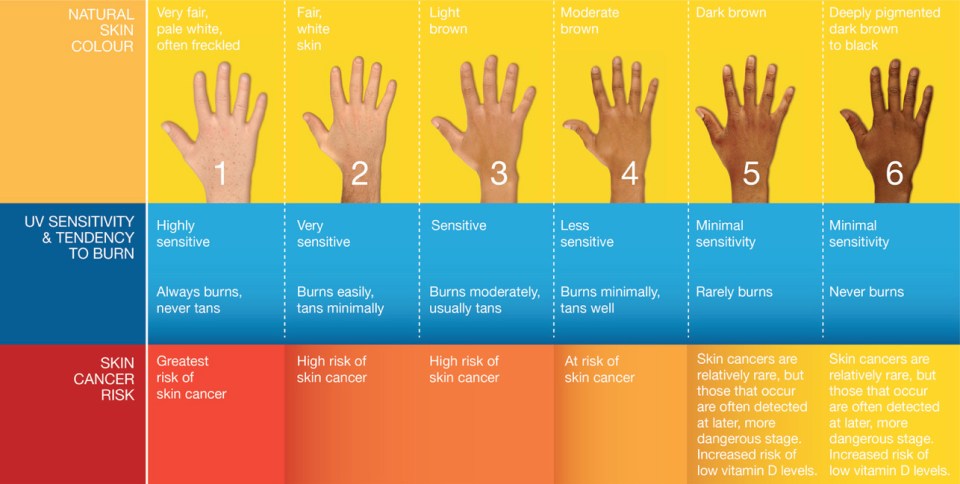

To avoid the influence of solar UVB exposure, this study was conducted in February. On day 0, both the black and white subjects were vitamin D deficient. The median levels of vitamin D in the white subjects was 11.9 ng/ml whereas for the black subjects it was 8.6 ng/ml—a non-statistically significant difference. On day two, however, concentrations of vitamin D in the blood rose from 11.9 to 13.3 ng/ml—a statistically significant difference. For the black cohort, however, there was no statistically significant difference in vitamin D levels. On day 6, levels in the white subjects rose from 11.6 to 14.3 ng/ml whereas for the black subjects it was 8.6 to 9.57 ng/ml. At the end of day 6, there was a statistically significant difference in circulating vitamin D levels between the white and black subjects (14.3 ng/ml compared to 9.57 ng/ml).
Different phototypes absorb different amounts of UV rays and, therefore, peoples with different skin color absorb different levels of vitamin D. Lighter-skinned people absorb more UV rays than darker-skinned people, showing that white skin’s primary cause is to synthesize vitamin D.
UVB exposure increases vitamin D production in white skin, but not in black skin. Pigmented skin, on the other hand, hinders the transformation of 7-dehydrocholesterol to vitamin D. This is why blacks have higher rates of prostate cancer—they are outside of their ancestral environment and what comes with being outside of one’s ancestral environment are evolutionary mismatches. We have now spread throughout the world, and people with certain skin colors may not be adapted for their current environment. This is what we see with black Americans as well as white Americans who spend too much time in climes that are not ancestral to them. Nevertheless, different-colored skin does synthesize vitamin D differently, and knowledge of this will increase the quality of life for everyone.
Even the great Darwin wrote about differences in human skin color. He didn’t touch human evolution in On the Origin of Species (Darwin, 1859), but he did in his book Descent of Man (Darwin, 1871). Darwin talks about the effects of climate on skin color and hair, writing:
It was formerly thought that the colour of the skin and the character of the hair were determined by light or heat; and although it can hardly be denied that some effect is thus produced, almost all observers now agree that the effect has been very small, even after exposure during many ages. (Darwin, 1871: 115-116)
Darwin, of course, championed sexual selection as the cause for human skin variation (Darwin, 1871: 241-250). Jared Diamond has the same view, believing that natural selection couldn’t account for hair loss, black skin and white skin weren’t products of natural selection, but female mate preference and sexual selection (Greaves, 2014).
Parental selection for white skin
Judith Rich Harris, author of the book The Nurture Assumption: Why Kids Turn Out the Way They Do (Harris, 2009), posits another hypothesis for the evolution of light skin for those living in northern latitudes—parental selection. This hypothesis may be controversial to some, as it states that dark skin is not beautiful and that white skin is.
Harris posits that selection for lighter skin was driven by sexual selection, but states that parental selection for lighter skin further helped the fixation of the alleles for white skin in northern populations. Neanderthals were a furry population, as they had no clothes, so, logic dictates that if they didn’t have clothes then they must have had some sort of protection against the cold Ice Age climate, therefore they must have had fur.
Harris states that since lighter skin is seen as more beautiful than darker skin, then if a woman birthed a darker/furrier babe than the mother would have committed infanticide. Women who birth at younger ages are more likely to commit infanticide, as they still have about twenty years to birth a babe. On the other hand, infanticide rates for mothers decrease as she gets older—because it’s harder to have children the older you get.
Harris states that Erectus may have been furry up until 2 mya, however, as I’ve shown, Erectus was furless and had the ability to thermoregulate—something that a hairy hominin was not able to do (Lieberman, 2015).
There is a preference for lighter-skinned females all throughout the world, in Africa (Coetzee et al, 2012); China and India (Naidoo et al, 2016; Dixson et al, 2007); and Latin America and the Philipines (Kiang and Takeuchi, 2009). Light skin is seen as attractive all throughout the world. Thus, since light skin allows better synthesize of vitamin D in colder climes with fewer UV rays, then there would have been a myriad of selective pressures to push that along—parental selection for lighter-skinned babes being one of them. This isn’t talked about often, but infanticide and rape have both driven our evolution (more on both in the future).
Harris’ parental selection hypothesis is plausible, and she does use the right dates for fur loss which coincides with the endurance running of Erectus and how he was able to thermoregulate body heat due to lack of fur and more sweat glands. This is when black skin began to evolve. So with migration into more northerly climes, lighter-skinned people would have more of an advantage than darker-skinned people. Infanticide is practiced all over the world, and is caused—partly—by a mother’s unconscious preferences.
Skin color and attractiveness
Lighter skin is seen as attractive all throughout the world. College-aged black women find lighter skin more attractive (Stephens and Thomas, 2012). It is no surprise that due to this, a lot of black women lighten their skin with chemicals.
In a sample of black men, lighter-skinned blacks were more likely to perceive discrimination than their darker-skinned counterparts (Uzogara et al, 2014). Further, in appraising skin color’s effect on in-group discrimination, medium-skinned black men perceived less discrimination than lighter- and darker-skinned black men. Lastly—as is the case with most studies—this effect was particularly pronounced for those in lower SES brackets. Speaking of SES, lighter-skinned blacks with higher income had lower blood pressure than darker-skinned blacks with higher income (Sweet et al, 2007). The authors conclude that a variety of psychosocial stress due to discrimination must be part of the reason why darker-skinned blacks with a high SES have worse blood pressure—but I think there is something else at work here. Darker skin on its own is associated with high blood pressure (Mosley et al, 2000). I don’t deny that (perceived) discrimination can and does heighten blood pressure—but the first thing that needs to be looked at is skin color.
Lighter-skinned women are seen as more attractive (Stephen et al, 2009). This is because it signals fertility, femininity, and youth. One more important thing it signals is the ability to carry a healthy child to term since lighter skin in women is associated with better vitamin D synthesis which is important for a growing babe.
Skin color and intelligence
There is a high negative correlation between skin color and intelligence, about –.92 (Templer and Arikawa, 2006). They used the data from Lynn and Vanhanen’s 2002 book IQ and the Wealth of Nations and found that there was an extremely strong negative correlation between skin color and IQ. However, data wasn’t collected for all countries tested and for half of the countries the IQs were ‘estimated’ from other surrounding countries’ IQs.
Jensen (2006) states that the main limitation in the study design of Arikawa and Templer (2006) is that “correlations obtained from this type of analysis are completely non-informative regarding any causal or functional connection between individual differences in skin pigmentation and individual differences in IQ, nor are they informative regarding the causal basis of the correlation, e.g., simple genetic association due to cross-assortative mating for skin color and IQ versus a pleiotropic correlation in which both of the phenotypically distinct but correlated traits are manifested by one and the same gene.”
Lynn (2002) purported to find a correlation of .14 in a representative sample of American blacks (n=430), concluding that the proportion of European genes in African Americans dictates how intelligent that individual black is. However, Hill (2002) showed that when controlling for childhood environmental factors such as SES, the correlation disappears and therefore, a genetic causality cannot be inferred from the data that Lynn (2002) used.
Since Lynn found a .14 correlation between skin color and IQ in black Americans, that means that only .0196 percent of the variation in IQ within black American adults can be explained by skin color. This is hardly anything to look at and keep in mind when thinking about racial differences in IQ.
However, other people have different ideas. Others may say that since animal studies find that lighter animals are less sexually active, are less aggressive, have a larger body mass, and greater stress resistance. So since this is seen in over 40 species of vertebrate, some fish species, and over 30 bird species (Rushton and Templer, 2012) that means that it should be a good predictor for human populations. Except it isn’t.
we know the genetic architecture of pigmentation. that is, we know all the genes (~10, usually less than 6 in pairwise between population comparisons). skin color varies via a small number of large effect trait loci. in contrast, I.Q. varies by a huge number of small effect loci. so logically the correlation is obviously just a correlation. to give you an example, SLC45A2 explains 25-40% of the variance between africans and europeans.
long story short: it’s stupid to keep repeating the correlation between skin color and I.Q. as if it’s a novel genetic story. it’s not. i hope don’t have to keep repeating this for too many years.
Finally, variation in skin color between human populations are primarily due to mutations on the genes MC1R, TYR, MATP (Graf, Hodgson, and Daal, 2005), and SLC24A5 (also see Lopez and Alonso, 2014 for a review of genes that account for skin color) so human populations aren’t “expected to consistently exhibit the associations between melanin-based coloration and the physiological and behavioural traits reported in our study” (Ducrest, Keller, and Roulin, 2008). Talking about just correlations is useless until causality is established (if it ever is).
Conclusion
The evolution of human skin variation is complex and is driven by more than one variable, but some are stronger than others. The evolution of black skin evolved—in part—due to skin cancer after we lost our fur. White skin evolved due to sexual selection (proximate cause) and to better absorb UV rays for vitamin D synthesis in colder climes (the true need for light skin in cold climates). Eurasians split around 40kya, and after this split both evolved light skin pigmentation independently. As I’ve shown, the alleles that code for skin color between blacks and whites don’t account for differences in aggression, nor do they account for differences in IQ. The genes that control skin color (about a dozen) pale in comparison to the genes that control intelligence (thousands of genes with small effects). Some other hypotheses for the evolution of white skin are on par with being as controversial as the hypothesis that skin color and intelligence co-evolved—mainly that mothers would kill darker-skinned babies because they weren’t seen as beautiful as lighter-skinned babies.
The evolution of human skin variation is extremely interesting with many competing hypotheses, however, to draw wild conclusions based on just correlations in regards to human skin color and intelligence and aggression, you’re going to need more evidence than just correlations.
References
Bang KM, Halder RM, White JE, Sampson CC, Wilson J. 1987. Skin cancer in black Americans: A review of 126 cases. J Natl Med Assoc 79:51–58
Beleza, S., Santos, A. M., Mcevoy, B., Alves, I., Martinho, C., Cameron, E., . . . Rocha, J. (2012). The Timing of Pigmentation Lightening in Europeans. Molecular Biology and Evolution,30(1), 24-35. doi:10.1093/molbev/mss207
Blum, H. F. (1961). Does the Melanin Pigment of Human Skin Have Adaptive Value?: An Essay in Human Ecology and the Evolution of Race. The Quarterly Review of Biology,36(1), 50-63. doi:10.1086/403275
Coetzee V, Faerber SJ, Greeff JM, Lefevre CE, Re DE, et al. (2012) African perceptions of female attractiveness. PLOS ONE 7: e48116.
Darwin, C. (1859). On the origin of species by means of natural selection, or, the preservation of favoured races in the struggle for life. London: J. Murray.
Darwin, C. (1871). The descent of man, and selection in relation to sex. London: John Murray, Albemarle Street.
Dixson, B. J., Dixson, A. F., Li, B., & Anderson, M. (2006). Studies of human physique and sexual attractiveness: Sexual preferences of men and women in China. American Journal of Human Biology,19(1), 88-95. doi:10.1002/ajhb.20584
Ducrest, A., Keller, L., & Roulin, A. (2008). Pleiotropy in the melanocortin system, coloration and behavioural syndromes. Trends in Ecology & Evolution,23(9), 502-510. doi:10.1016/j.tree.2008.06.001
Frost, P. (2007). Human skin-color sexual dimorphism: A test of the sexual selection hypothesis. American Journal of Physical Anthropology,133(1), 779-780. doi:10.1002/ajpa.20555
Graf, J., Hodgson, R., & Daal, A. V. (2005). Single nucleotide polymorphisms in theMATP gene are associated with normal human pigmentation variation. Human Mutation,25(3), 278-284. doi:10.1002/humu.20143
Greaves, M. (2014). Was skin cancer a selective force for black pigmentation in early hominin evolution? Proceedings of the Royal Society B: Biological Sciences,281(1781), 20132955-20132955. doi:10.1098/rspb.2013.2955
Gupta, D., Lammersfeld, C. A., Trukova, K., & Lis, C. G. (2009). Vitamin D and prostate cancer risk: a review of the epidemiological literature. Prostate Cancer and Prostatic Diseases,12(3), 215-226. doi:10.1038/pcan.2009.7
Guthrie RD. 1970. Evolution of human threat display organs. Evol Biol 4:257–302.
Harris, J. R. (2006). Parental selection: A third selection process in the evolution of human hairlessness and skin color. Medical Hypotheses,66(6), 1053-1059. doi:10.1016/j.mehy.2006.01.027
Harris, J. R. (2009). The nurture assumption: why children turn out the way they do. New York: Free Press.
Hill, Mark E. 2002. Skin color and intelligence in African Americans: A reanalysis of Lynn’s data. Population and Environment 24, no. 2:209–14
Holick, M. F. (2006). Resurrection of vitamin D deficiency and rickets. Journal of Clinical Investigation,116(8), 2062-2072. doi:10.1172/jci29449
Jablonski, N. G., & Chaplin, G. (2000). The evolution of human skin coloration. Journal of Human Evolution,39(1), 57-106. doi:10.1006/jhev.2000.0403
Jablonski, N. G., & Chaplin, G. (2014). Skin cancer was not a potent selective force in the evolution of protective pigmentation in early hominins. Proceedings of the Royal Society B: Biological Sciences,281(1789), 20140517-20140517. doi:10.1098/rspb.2014.0517
Jensen, A. R. (2006). Comments on correlations of IQ with skin color and geographic–demographic variables. Intelligence,34(2), 128-131. doi:10.1016/j.intell.2005.04.003
Kiang, L., & Takeuchi, D. T. (2009). Phenotypic Bias and Ethnic Identity in Filipino Americans. Social Science Quarterly,90(2), 428-445. doi:10.1111/j.1540-6237.2009.00625.x
Libon, F., Cavalier, E., & Nikkels, A. (2013). Skin Color Is Relevant to Vitamin D Synthesis. Dermatology,227(3), 250-254. doi:10.1159/000354750
Lieberman, D. E. (2015). Human Locomotion and Heat Loss: An Evolutionary Perspective. Comprehensive Physiology, 99-117. doi:10.1002/cphy.c140011
Lieberman, D. E., Raichlen, D. A., Pontzer, H., Bramble, D. M., & Cutright-Smith, E. (2006). The human gluteus maximus and its role in running. Journal of Experimental Biology,209(11), 2143-2155. doi:10.1242/jeb.02255
López, S., & Alonso, S. (2014). Evolution of Skin Pigmentation Differences in Humans. ELS. doi:10.1002/9780470015902.a0021001.pub2
Lynn, R. (2002). Skin color and intelligence in African Americans. Population and Environment, 23, 365–375.
Mosley, J. D., Appel, L. J., Ashour, Z., Coresh, J., Whelton, P. K., & Ibrahim, M. M. (2000). Relationship Between Skin Color and Blood Pressure in Egyptian Adults : Results From the National Hypertension Project. Hypertension,36(2), 296-302. doi:10.1161/01.hyp.36.2.296
Naidoo, L.; Khoza, N.; Dlova, N.C. A fairer face, a fairer tomorrow? A review of skin lighteners. Cosmetics 2016, 3, 33.
Norton, H. L., Kittles, R. A., Parra, E., Mckeigue, P., Mao, X., Cheng, K., . . . Shriver, M. D. (2006). Genetic Evidence for the Convergent Evolution of Light Skin in Europeans and East Asians. Molecular Biology and Evolution,24(3), 710-722. doi:10.1093/molbev/msl203
Rushton, J. P., & Templer, D. I. (2012). Do pigmentation and the melanocortin system modulate aggression and sexuality in humans as they do in other animals? Personality and Individual Differences,53(1), 4-8. doi:10.1016/j.paid.2012.02.015
Sankararaman, S., Mallick, S., Dannemann, M., Prüfer, K., Kelso, J., Pääbo, S., . . . Reich, D. (2014). The genomic landscape of Neanderthal ancestry in present-day humans. Nature,507(7492), 354-357. doi:10.1038/nature12961
Stephen, I. D., Smith, M. J., Stirrat, M. R., & Perrett, D. I. (2009). Facial Skin Coloration Affects Perceived Health of Human Faces. International Journal of Primatology,30(6), 845-857. doi:10.1007/s10764-009-9380-z
Stephens, D., & Thomas, T. L. (2012). The Influence of Skin Color on Heterosexual Black College Women’s Dating Beliefs. Journal of Feminist Family Therapy,24(4), 291-315. doi:10.1080/08952833.2012.710815
Sweet, E., Mcdade, T. W., Kiefe, C. I., & Liu, K. (2007). Relationships Between Skin Color, Income, and Blood Pressure Among African Americans in the CARDIA Study. American Journal of Public Health,97(12), 2253-2259. doi:10.2105/ajph.2006.088799
Tateno, Y., Komiyama, T., Katoh, T., Munkhbat, B., Oka, A., Haida, Y., . . . Inoko, H. (2014). Divergence of East Asians and Europeans Estimated Using Male- and Female-Specific Genetic Markers. Genome Biology and Evolution,6(3), 466-473. doi:10.1093/gbe/evu027
Templer, D. I., & Arikawa, H. (2006). Temperature, skin color, per capita income, and IQ: An international perspective. Intelligence,34(2), 121-139. doi:10.1016/j.intell.2005.04.002
Uzogara, E. E., Lee, H., Abdou, C. M., & Jackson, J. S. (2014). A comparison of skin tone discrimination among African American men: 1995 and 2003. Psychology of Men & Masculinity, 15(2), 201–212. http://doi.org/10.1037/a0033479
van den Berghe PL, Frost P. 1986. Skin color preference, sexual dimorphism and sexual selection: a case of gene-culture coevolution? Ethn Racial Stud 9:87–113.
Problems With Forensic Facial “Reconstruction”: Implications for the Facial “Reconstruction” of Ancient Hominin
1050 words
Forensic facial reconstruction is the process by which a face is reconstructed from a deceased individual’s “skeletal remains through an amalgamation of artistry, forensic science, anthropology, osteology, and anatomy.” This technique is used largely only when skeletal remains are the only evidence at the scene of a crime. Perhaps most famously, it was used in the facial reconstruction of Mitochondrial Eve—but just how accurate is forensic facial reconstruction?
The average person may believe that forensic facial reconstruction is successful and a good proxy for what an individual may have looked like. This is bolstered by the fact that people hear of stories in which facial reconstruction are successful, never hearing of the countless number of cases in which the “reconstruction” fails to identify anyone.
Numerous papers in the literature, though, do show that forensic facial reconstruction does have a high success rate. For instance, Lee et al (2011) and Wilkinson et al (2006) show that this method has a considerable chance of getting facial morphology in the ballpark of how they look. However, this was done on live subjects and is of no use for Mitochondrial Eve/deceased individuals. Facial reconstructions of the deceased are, for the purpose of this article, what we need to look at, not studies looking at live people. (There are also hurdles for facial recognition systems.)
One of the biggest hurdles for the accuracy of forensic facial reconstruction is that average facial tissue thickness cannot be inferred (most importantly, the lips, cartilage, skin and fat). Due to this, with no prior information to look at, the look of the skull will be subjective with the “reconstruction” looking somewhat similar due to chance. Further, the main facial features are largely determined by the shape of the skull.
Now to the fun part: Is this what Mitochondrial Eve really looked like?

There are a number of features that are problematic to infer from these facial reconstructions. Lips, ears, skin, craniofacial muscles—all are extremely hard, or next to impossible, to predict if the only thing we had was a skeleton. Further, since there are few tested relationships between soft and hard tissue for modern humans “it is clear that the use of facial approximation techniques on ancestral skulls of modern Homo are fundamentally flawed, as previously reported by Montagu” (Stephan, 2003). Since soft tissue quickly decays, it is left up to artistic interpretation. Further, attempting to map ape morphology since we diverged a few million years before is misleading, due to the fact that hard and soft tissue relationships are not likely to be the same for apes and our hominin ancestors.
Now that we know the so-called “reconstruction” of Mitochondrial Eve is not what she really looked like, there are a few more problems with this method I’d like to go over.
Forensic facial reconstruction is used when the remains of an unidentified individual are discovered. If the bones of the deceased are all that forensic artists have to go off of, the finished product may be extremely subjective/biased. People who believe that forensic facial reconstruction truly works may say “It works all the time. If it didn’t, how would it be able to solve crimes?” Success rates for the identification of individuals ranges from 50 to 100 percent (Stephan, 2003: 196), and so, the belief that “reconstructions” are largely accurate continue to persist.
However, like with the case of the famous stereotype threat with a modicum of unpublished studies, the “success” of forensic facial reconstruction is also skewed by non-reporting of unsuccessful cases
It is also rare for forensic facial approximation to be to be better than chance, with 403 incorrect facial identifications out of 592 identification scenarios in one study (Stephan and Henneberg, 2001). Facial reconstruction has the greatest accuracy if there is any knowledge of past injuries for the individual, a photo, and soft tissue. Obviously, in the case of Mitochondrial Eve, we don’t have a photo nor do we have soft tissue and knowledge of past injuries are a non-factor. So it seems that if some people claim to know what the first AMH looked like, it’s probably “just a guess”, and a pretty bad one at that due to the no knowledge of hard and soft facial tissue in the hominin lineage.
The hardest part about facial reconstruction is reconstructing soft tissues accurately since they quickly decompose. This is even more of a problem for people who lived hundreds of thousands of years ago. We’ve never seen any alive so we don’t know what they may have looked like to be able to infer what Mitochondrial Eve would have looked like. Things like lips/mouth, skin, hair, and ears are largely up to artistic interpretation, which are subjective in nature. Craniofacial morphology has also changed in the past 200 thousand years, which may be due to a decrease in testosterone/androgen receptors.
If we can’t identify humans with facial recognition better than chance, what makes anyone think that we can even have the slightest idea of how Mitochondrial Eve looked—when some of the most important parts of the phenotype aren’t around to observe and thus subjectivity then comes into play. Any “reconstructions” you come across, you should take with a grain of salt. It’s next to impossible to know what ancient hominins may have looked like due to the absence of soft tissue, and so any phenotype that a so-called “reconstruction” may give is, largely, up to the interpretation of the individual artist.
With our current technology, it’s next to impossible to ascertain what Mitochondrial Eve—or any other ancient hominin for that matter—may have looked like.
References
Lee, W., Wilkinson, C. M., & Hwang, H. (2011). An Accuracy Assessment of Forensic Computerized Facial Reconstruction Employing Cone-Beam Computed Tomography from Live Subjects. Journal of Forensic Sciences,57(2), 318-327. doi:10.1111/j.1556-4029.2011.01971.x
Stephan, C. (2003). Anthropological facial ‘reconstruction’ – recognizing the fallacies, ‘unembracing’ the errors, and realizing method limits. Science & Justice,43(4), 193-200. doi:10.1016/s1355-0306(03)71776-6
Stephan, C. N., & Henneberg, M. (2001). Building Faces from Dry Skulls: Are They Recognized Above Chance Rates? Journal of Forensic Sciences,46(3). doi:10.1520/jfs14993j
Wilkinson, C., Rynn, C., Peters, H., Taister, M., Kau, C. H., & Richmond, S. (2006). A Blind Accuracy Assessment of Computer-Modeled Forensic Facial Reconstruction Using Computed Tomography Data From Live Subjects. Forensic Science, Medicine and Pathology,2(3), 179-188. doi:10.1385/fsmp:2:3:179
Wilkinson, C. (2010). Facial reconstruction—anatomical art or artistic anatomy? Journal of Anatomy,216(2), 235-250. doi:10.1111/j.1469-7580.2009.01182.x
Reading Wrongthought Books In Public
750 words.
How many of you have read wrongthought books in public? Books like Race, Evolution, and Behavior, The Blank Slate, The Nurture Assumption, or any other kind of wrongthought literature? I’ve had two specific run-ins this week reading a wrongthought book in public, both people giving me the expected reaction “Why are you reading that?” I bring a few books with me in my daily travels, one of them being Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid To Talk About It.
I was in the bookstore the other day reading Taboo while drinking some coffee and someone came up to me and said, “Why are you reading that?” I responded, “Because I’m interested in the material, why else why I read it?” The person scowled at me and walked away. I don’t understand why you need to ask dumb questions like that. Clearly, if I’m reading something I find an interest in it. People are clearly scared of acknowledging any kind of racial differences in public—even if they cheer for their favorite sports teams a few times a week, not realizing that they’re cheering on people who have inherent genetic advantages due to their morphology. Ironically enough, the book I was reading was talking about how and why blacks dominate sports. That’s ‘racist’ though, so people don’t want to talk about it specifically in public, but they know the truth unconsciously.
As I wrote the other day, people are scared of talking about things that are “natural” in human populations. Because the admission of one population being inherently better, on average than another would then open the gates to talking about other more uncomfortable things, such as racial differences in intelligence. I bet seeing someone read a book like Taboo in public without having knowledge of population differences is pretty jarring for someone who is not privy to the information. However, as we all know, reality isn’t based on how we wish it to be. These taboo subjects that people are scared to approach don’t go away if they don’t acknowledge them.
I had another run-in the other day while drinking some coffee, reading the same wrongthought book. The person asked me “Why are you reading that in public?” with a surprised look on his face. I laughed and said, “Because I can,” and sipped my coffee. People don’t want their preconceived notions challenged (the notion of ‘equality’ of human races). To see someone reading a wrongthought book in public is heresy, and people will always give you an odd look if you’re reading one in public.
I also saw someone reading Outliers by Malcolm Gladwell the other day and I just had to tell them that Gladwell is wrong in his contention that practicing something for 10,000 hours is something that Gladwell has been rebutted on and continues to perpetuate. People should always know there are two sides to every story, whether they want to hear it or not.
People change their minds through reading. So if people who’ve not been exposed to wrongthought books see others reading them, they may be compelled to look into it themselves. That’s the best part about reading those types of books in public. A person may look into it themselves and change their views. I’ve personally found it much easier to change my views on something when I’ve read it myself in comparison to if I’m debating someone.
There is a cognitive bias known as the ‘backfire effect’, where correcting someone actually increases their misperceptions. Engaging people’s beliefs, in the end, just makes them hold onto them more strongly and has them search for evidence to confirm their beliefs. So by people being exposed to wrongthought books they can see them and look into it for themselves and weigh both sides on their own without the possibility of the backfire effect.
So just by exposing others to the covers of these wrongthought books may have them look into something themselves and, possibly, someone else will accept genetic causes for human differences. If you do read a wrongthought book in public and someone attempts to engage you due to what you’re reading, remember the backfire effect. This is why debating people doesn’t change their views because showing people they are wrong (with scholarly sources) has them hold onto their views more strongly. To avoid this, just read a wrongthought book in public so people can see it and look into it themselves.
