
Height and IQ Genes
1100 words
Genes account for about 80 percent of the variation in height and IQ, with both height and IQ correlating at .2. Therefore, genes must contribute largely to population variances in height. However, finding certain genes that contribute largely to these two traits is a problem, largely because both traits are polygenic in nature. Recent research has shown that most—or all–genes are height genes. If this is the case, are most—or all—genes IQ genes?
Height is around 80-90 percent heritable (Peeters et al, 2009). What this means is that the difference between the tallest and shortest 5 percent of the population is 11 inches, with 10 inches being accounted for by genes and 1 inch being accounted for by environment (Heine, 2017: 30). The gene that contributes the most to human height has been found to give 1/6th of an inch (Weedon et al, 2007). However, a recent meta-analysis shows that certain rare alleles give as much as 8/10ths of an inch (Hirschhorn, Deloukas, and Lettre, 2017). Furthermore, thousands of gene variants combined explain about 50 percent of human height (Yang et al, 2010). Yang et al (2010) also found 294,831 SNPs related to people’s height, which is—more or less—12 times the number of genes in our genome (Heine, 2017: 30; the number of genes in our genome is in the range of 19,000-20,000; Ezkurdia et al, 2014). Another meta-analysis found that 697 genetic variants explain about 20 percent of the genetic variation (Wood et al, 2014). Furthermore, according to geneticist David Goldstein, “most genes are height genes” (Goldstein, 2009).
Author of the book DNA is not Destiny and cultural and social psychologist Steven J. Heine writes:
“This means if you wanted to genetically engineer a designer baby who you would like to grow up to be tall, you would have to make almost 300,000 genetic alterations to the genome and you still would only be half way there. When the genetic evidence suggests that almost all genes are related to height, then in a way, we learn close to nothing about the genetic basis of height.” (Heine, 2017: 30)
Hirschhorn, Deloukas, and Lettre, (2017) found 83 rare and low-frequency genes that explain 1.7 percent of the adult heritability of height, along with newly identified and novel variants that explained 2.4 percent, “and all independent variants, known and novel together explained 27.4% of heritability. By comparison, the 697 known height SNPs explain 23.3% of height heritability in the same dataset (vs. 4.1% by the new height variants identified in this ExomeChip study)” (pg 7). So 27.4 percent of the variance is explained by known common variants and these new variants discovered.
Americans who drink more milk are, on average, half an inch taller than Americans who don’t recall drinking as much milk, even after controlling for race, income, and education (Wiley, 2005). This shows the importance milk has on skeletal muscle growth. This increase has even been noticed in Japan, where they increased their milk intake using school lunch programs (Takahasi, 1984), which increased their height by 4 inches (Funatogawa et al, 2009).
We also grow more in the spring and summer than in the fall and winter. This is due to ultraviolet radiation from the sun’s rays that synthesize some of the vitamin D we drink that is in the cow’s milk. Clearly, environmental factors (UV rays, milk consumption, overall nutrition, etc) all have a part to play in human height variation (Heine, 2017: 30). However, if all genes may be height genes, may all genes be IQ genes?
In regards to IQ, 3 genetic variants explain .3 IQ points (Rietvald et al, 2014):
After adjusting the estimated effect sizes of the SNPs (each R2 ∼ 0.0006) for the winner’s curse, we estimate each as R2 ∼ 0.0002 (SI Appendix), or in terms of coefficient magnitude, each additional reference allele for each SNP is associated with an ∼0.02 SD increase in cognitive performance [or 0.3 points on the typical intelligence quotient (IQ) scale].
This is the gene with the highest known effect that we currently know of. No “but undiscovered X means Y!!”, because science isn’t based on ‘what ifs’.
To predict one’s intelligence, you would need all genes on an SNP chip—which contains about 500,000 SNPs—to be able to predict half of the individual variation in IQ (Davies et al, 2011; Chabris et al, 2012; Heine, 2017: 175). Just as is the case with height, it seems that it’s possible that most—if not all—genes are IQ genes.
So, clearly, intelligence is highly polygenic, and, contrary to what Plomin says, it’s doubtful that we’ll be able to genotype one to guesstimate their intelligence level.
This is because you need more than 500,000 SNPs on a gene chip and even still, that would only explain half of the variance. So it’s reasonable to assume—as is the case with height—that all genes are IQ genes.
Chabris et al (2012) write:
One SNP, rs2760118 in SSADH (also known as ALDH5A1), exhibited a nominally significant association with g (t = 2.01, p = .04), but this association did not survive a Bonferroni correction. The mean g values (transformed to the IQ scale) by genotype for this SNP were 98.3, 99.7, and 100.6 for genotypes TT, TC, and CC respectively.
So it seems that all genes are height genes and all genes could possibly be IQ genes (that is, having a small effect). If most genes are height genes, and height is linked to IQ, then most genes should be IQ genes as well. Therefore, it is plausible that all genes are IQ genes.
Finally, I need to talk about the study that everyone is talking about, the study that found 52 new genes for intelligence (Sniekers et al, 2017). However, Razib Khan cautions: “My plain words are this: do not trust, and always verify“. A Google search for “gene found for” brings up 26,300,000 hits. As can be seen with the study that was published the other day on the supposed ‘new hominin’ found in Europe, science journalists use fancy and catchy headlines. “Genes for ___ and ___” is a bad way to put it—few traits are caused by a single gene, and most traits are highly polygenic, height and IQ included.
Do I think we’ll disentangle the intricacies involved with height and IQ? One day. But since at the moment, 500,000 SNPs need to be loaded on a gene chip to explain half of the variation in individual IQ.
Since most—or all—genes are related to height and the same may be so for IQ, we don’t really learn anything knowing the genes that control for these two traits. In regards to Heine’s (2017) example of genetic engineering 300,000 SNPs for height and you’d only be halfway there, I’d assume the same would be true for IQ. Both traits are highly polygenic, with thousands of genes controlling these traits. Genetic engineering a human for high intelligence or height looks to be a long shot—at least until far into the future.
Rebutting Heartiste on the Twinkie Diet
1300 words
Nutritional myths run amok everywhere. One of the most persistent is that ‘a calorie is a calorie’, that is, every macronutrient will be processed the same by the body. Another assumption is that the body doesn’t ‘care’ about where the calories come from—they can come from fat, protein, or carbs and the response will be the same: bodyweight will be reduced until one reaches their goal. However, it’s not as simple as that. He also has the assumption that “diets work”, when the best meta-analysis I know of on the matter shows the opposite (Mann et al, 2007, see especially table 1). They control for studies where weight was self-reported. They conclude that dieting does not work. This is what, as Heartiste says, “iScience!” says on the matter, so he should believe everything I state in this article, which is backed by “iScience!”.
Chateau Heartiste published an article back in 2010 titled The Twinkies Diet Proves Fatty Fats Are Fat Because They Eat Too Much. He is referring to professor of human nutrition Mark Haub and his success on ‘the twinkies diet’, where 2/3rds of his caloric intake came from junk food such as Twinkies. He lost 27 pounds in a two month period while his LDL cholesterol decreased by 20 percent and his HDL cholesterol increased by 20 percent. His level of triglycerides also decreased by 37 percent, with his body fat decreasing from 33.4 to 24.9 percent. So he ate 1800 kcal per day—2/3rds of it being junk food—for two months and lost 27 pounds. Case closed, right? Eat junk food at a deficit and lose weight? A calorie is a calorie? There are a few problems with this contention which will be addressed below.
Heartiste writes:
Big bottom line: Being fat itself is bad for your health. “Fat and fit” is a myth. The change that counts the most is losing the weight, which can only be done by PUSHING AWAY FROM THE TABLE.
Except fit and overweight and obese individuals have similar mortality rates than their normal weight counterparts (Barry et al, 2014). However, more recently a study was published purporting that overweight and obese individuals being healthy despite excess weight is a myth. The researchers state that in a sample of millions of Britons that overweight and obese individuals had a higher risk of heart disease than their normal-weight counterparts. Unfortunately, I cannot locate the study since it wasn’t published in a journal (and thusly not peer reviewed). I wonder if variables such as diet, smoking and other lifestyle factors were taken into account. Nevertheless, the debate on fitness and fatness continues.
Another large meta-analysis shows that grade 1 obesity (BMI 30->35) had the same mortality risk as normal-weight individuals with grade 2 obese (BMI +35) having a significantly higher risk of death (Flegal, Kit, and Orpana, 2013).
Heartiste claims that ‘a calorie is a calorie’. This is a common fallacy. This suggests that the body will process all foods the same way—that is, processing them the same metabolically. This, however, is not the case. Haub himself is a sample size of 1. If Heartiste can use a sample size of 1 to make a claim, then I can too.
Sam Feltham ate +5,000 kcal per day for 21 days and only gained 1.3 kg when he should have gained 7.3 kg based on the amount of kcal he ate. A calorie is a calorie, right? This is a fallacious statement. The statement “a calorie is a calorie” violates the second law of thermodynamics (Feinman and Fine, 2004). Heartiste writes:
That first law of thermodynamics looms large over everything.
The first law of thermodynamics is irrelevant to human physiology. It only states that an organism gets bigger if it consumes more energy; it doesn’t state why this occurs, which is due to the hormone insulin which causes weight gain.
He does rightly state that an omega 3/6 imbalance is part of the reason but then handwaves it away. Western-like high-fat diets (i.e., diets with an imbalance of linoleic acids (LA; and n-6 fatty acid) with n-3) are sufficient enough to induce gradual enhancement in fat mass across the generations (Massiera et al, 2010). This obviously includes the average 55 percent carbohydrate diet that the AHA recommends (Eckel et al, 2014). The Standard American Diet (aptly named the “SAD diet”) has the n-3/n-6 imbalance along with being high in carbohydrates which spike insulin which impedes fat being unlocked from the adipocyte.
Heartiste doesn’t understand that if you reduce the ‘in’, the ‘out’ also decreases. This was noted in the famous starvation experiment headed by Ancel Keys. They took 36 healthy men who ate normally for three months while being their behavior and personality was monitored. In the next six months, they were reduced to eating half of their initial intake (they started at 2000 kcal and dropped to 1000 kcal; some individuals going lower than that) and their metabolic rate decreased by 40 percent (Keys et al, 1945). This is proof for the contention that the body decreases its metabolic rate due to what is ingested. A similar study was done on Vermont prisoners, except they were told to gorge on food. Since they were in a controlled setting, the prisoners could be monitored to ensure they ate all of the food.
At the end of the study, their metabolic rates had increased by 50 percent. This is evidence that the body was trying to get back to its original weight. In six months, the prisoners went back to their normal weight as they ate normally (Salas, Horton, and Sims, 1971) One man only gained ten pounds eating all of those calories. Clearly, the body was resisting weight gain and when they were allowed to eat normally, they effortlessly regained their normal weights.
Finally, on the topic of Haub, Big Food shill, I will address a few things about him and his ‘research’ that recently came to light.
Intermittent fasting and obesity expert Dr. Jason Fung showed that in 2016 after Coca-Cola released their funding reports after criticisms of transparency, Mark Haub was found to be one of the many researchers that were backed by Coca-Cola. This is an attempt to show that ‘a calorie is a calorie’ and that ‘all calories are created equal’. This has been rebutted above.
In 2016—six years after his ‘experiment—it was revealed that he was funded by Coca-Cola. No doubt in order to ‘prove’ that ‘a calorie is a calorie’ and have people continue to gorge on high carbohydrate/insulinogenic foods. However, the human body is a lot more complex than to just reduce it to simply calories in and calories out—which I have written about in depth.
People like Heartiste need to get an actual understanding of the literature and what Coca-Cola has been trying to do for years, which is to make eating junk food ‘OK’ because ‘it doesn’t cause obesity’. Children consume 45 percent more food when exposed to advertisements (Harris, Bargh, and Brownell, 2009). So to begin to curb obesity rates we don’t need to ‘eat junk food’, we need to not eat junk food and eat a diet more ancestral to us—that is, one lower in processed carbs and higher in animal fat and protein. Big Food shills like Haub need to be exposed for what they are—people who do ‘research’ for a quick buck, i.e., not furthering our understanding of a complex issue as he would like you to believe. Exercise also doesn’t induce weight loss. So the claims of ‘eat less and move more’ (eat less according to the 55 percent carbohydrate recommendations) is bound to fail.
If Heartiste can make a claim using one man as an example then so can I. Read the above article by Sam Feltham in which he writes about hs experience eating 5,000 kcal per day for 21 days while only gaining 1.3 kg. I can use this example to say that eating low carb and high fat at 5,000 kcal per day will lead to negligible weight gain, however, I don’t use n=1 sample sizes to make claims and no one else should either.
Modern Man Did Not Begin in Europe
1300 words
A lot of buzz is going around about a recent study that purports that the human-chimp split occurred in the Mediterranean—not Africa as is commonly thought (Fuss et al, 2017). This claim, however, is based off of a few teeth and jawbone with one tooth in it of a supposed hominin named Graecopithecus freybergi. A lot of wild conclusions are being jumped to about this study and these claims need to be put to rest.
On altright.com, an article was written titled Recent Discovery Shows Humans Came From Europe. The article claims that the OoA hypothesis has been “debunked by hard evidence”. Though to disprove the OoA hypothesis, a lot more will be needed than a few teeth and a jawbone. This is similar to another article on the Dailystormer, New Discovery Shows Pre-Human Hominids in Europe Before Africa, which, again, makes more wild claims.
white people are just Negroes who have lighter skin because during the Ice Age, they were wearing more clothing and thus needed lighter skin to absorb more Vitamin D. needed lighter skin to absorb more Vitamin D.
So, the reasoning behind the theory is that we are all very close together, genetically, so there isn’t really any problem with mixing us all together, and pretty much, race is a social construct.
Race is a social construct of a biological reality. OoA is based on solid evidence. Just because ‘we are close together, genetically’, as egalitarians would say, doesn’t mean we should destroy the human diversity we currently have.
Much damage to evolutionary research was done by the Jew Stephen Jay Gould3, who argued in favor of the idea of rapid evolution
This is bullshit. Stephen J. Gould and Niles Eldredge proposed Puncuated Equilibria (PE). PE occurs when a species becomes as ‘adapted’ to its environment as possible and then remains in stasis. Species speciate when the environment changes (climate), or, say an earthquake occurs and splits a population of 100 peacocks in half. Fifty of the peacocks will change due to drift, natural and sexual selection. But if the environmental conditions stay the same then species cannot change.
Punctuated Equlibria is an alternative to phyletic gradualism.
What is punctuated equilibrium? What is macroevolution? A response to Pennell et al
The proposal was that after a long time in stasis that quick speciation would occur—that would be in response to the environmental change that drives the evolution of species.
He also made much damage to the field of sociobiology, literally arguing that evolution has no role in human behavior
He argued that many functions of the ‘higher’ functions of the human brain evolved for other reasons and were coopted for other reasons, which is why he coined the term ‘exaptation’.
And the debate about spandrels—which is a phenotypic trait that is a byproduct of the evolution of another trait and not due to adaptive selection. There is a tendency to assume that all—or most—traits are due to adaptive selection. This is not true.
PZ Myers – Bad Biology: How Adaptationist Thinking Corrupts Science
With this new discovery of prehumans in Europe, they are dating the European fossil as older, but we would basically end up with the same conclusions with regards to rapid evolution and thus “race not existing.” So I don’t see anything for racists to get all excited about, with the way it is currently being presented.
Punctuated Equilibria is a lot more nuanced than you’re making it out to be. It’s looking at the whole entire fossil record and noticing that for most of a species’ evolutionary history that it remains in stasis and that evolution then occurs in quick bursts.
This theory postulates that Africans, Asians, Europeans and Aboriginal Australians all evolved completely separately from different hominidae
This is not tenable. This isn’t even how it works. Neanderthals and Homo sapiens are derived from Heidelbergensis.
Erectus is in our family tree beginning 2 mya. He is an origin of AMH and us as well. However, what you’re talking about needs to be proven with genetic testing.
Africans of course are more violent (and larger) not so much because of IQ, but because of higher levels of testosterone, but no one has explained what caused this.
Claims about substantially higher levels of testosterone in blacks are not true.
Then, of course, the Asians who moved north developed higher IQs and lighter skin because of climate-related reasons.
East Asians needed bigger brains for expertise capcity; not IQ. Light skin did evolve for climatic reasons; not sexual selection as some claim.
People need to 1) learn the basics of evolutionary theory; 2) learn the basics of the OoA hypothesis; 3) stop jumping to conclusions based on little evidence and large conjecture; 4) never trust anything at face value; always do more research into something and put all ideas under intense scrutiny, even ones you strongly believe. That way, articles like the ones above don’t get writtent with complete disregard for modern-day evolutionary theory.
John Hawks, paleoanthropologist writes:
Here’s what I think: Paleoanthropology must move past the point where a mandibular fragment is accepted as sufficient evidence.
He also states that this may be a case of apes evolving “supposed hominin characters” in the Miocene, citing a study by him and his colleagues showing that features that supposedly link Ardipethicus and Sahelanthropus are also found in other Miocene fossils (Wolpoff et al, 2006). Graecopithecus shares few features with Australopithecus, so Hawks says that we should begin to think about the possibility of Graecopithecus being “part of a diversity of apes that are continuous across parts of Africa and Europe.”
Finally, there is not enough evidence to back the claim that humans originated in Europe. Vertebrate paleontologist and paleobiologist Dr. Julian Benoit states that the author’s claim of the fourth molar root in Graecopithecus being similar to hominins is unfounded because “This is not a character that is conventionally used in palaeoanthropology, especially because not all hominins have similar tooth roots. This character is rather variable – and the authors go on to acknowledge this – so it’s unreliable for classification.” Further, humans aren’t the only apes with small canines and the jawbone and teeth aren’t too well preserved.
We have found thousands of hominin fossils in Africa. We know that the LCA between apes and humans existed between 6-12 mya in Africa. Graecopithecus was probably an ape species not related to humans. Even if the claim were true, it wouldn’t completely disprove the hypothesis that Man originated in Africa. Extraordinary claims require extraordinary evidence; this is not it.
People need to stop letting their biases and political beliefs get in the way of rational thought. Never take claims at face value; always look at things objectively. There needs to be a lot more evidence for the claim that Man originated in Europe; and even then, there is a mountain of evidence that anatomically modern humans arose in Africa.
In order to prove that Graecopithecus was a hominin and not another species of non-human ape, more fossils need to be found and a phylogenetic analysis needs to be done on the jawbone, comparing it with other species to see the closest relationship on the phylogeny. I assume when this is done it will show that it is related to non-human apes; not humans. Nevertheless, extraordinary claims require extraordinary evidence and people need to stop believing and agreeing with everything that ‘agrees’ with their worldviews as a fact without taking an objective look at the data. Never trust claims and always attempt to verify that what someone claims has a basis in reality. Only ask yourself what the facts are and what they show—without bias.
Dinosaurs, Brains, and ‘Progressive Evolution’
1800 words
Would dinosaurs have reached human-like intellect had the K-T extinction (an asteroid impact near the Yucatan peninsula) not occurred? One researcher believes so, and he believes that a dinosaur called the troodon would have evolved into a bipedal, human-like being. This is, of course, the old progressive evolution shtick. This assumes that a man-like being is an inevitability, and that sentience is a forgone conclusion.
This belief largely comes from Rushton’s citation of one Dale Russel, the discoverer of the dinosaur the troodon:
Paleontologist Dale Russell (1983,1989) quantified increasing neurological complexity through 700 million years of Earth history in invertebrates and vertebrates alike. The trend was increasing encephalization among the dinosaurs that existed for 140 million years and vanished 65 million years ago. Russell (1989) proposed that if they had not gone extinct, dinosaurs would have progressed to a large-brained, bipedal descendent. For living mammals he set the mean encephalization, the ratio of brain size to body size, at 1.00, and calculated that 65 million years ago it was only about 0.30. Encephalization quotients for living molluscs vary between 0.043 and 0.31, and for living insects between 0.008 and 0.045 but in these groups the less encephalized living species resemble forms that appeared relatively early in the geologic record, and the more encephalized species resemble those that appeared later. (Rushton, 1997: 294)
This argument is simple to rebut. What is being described is complexity. The simplest possible organism are bacteria, which reside at the left wall of complexity. The left wall “induces right-skewed distributions”, whereas the right wall induces “left-skewed distributions” (Gould, 1996: 55). Knowing this, biological complexity is a forgone conclusion, which exists at the extreme end of the right tail curve. I’ve covered this in my article Complexity, Walls, 0.400 Hitting and Evolutionary “Progress”
Talking about what Troodons may have looked like (highly, highly, doubtful. The anthropometric bias was pretty strong) is a waste of time. I’ve stated this a few times and I’ll state it yet again: without our primate body plan, our brains are pretty much useless. Our body needs our brain; our brain needs our body. Troodons would have stayed quadrupedal; they wouldn’t have gone bipedal.
He claims that some dinosaurs would have eventually reached an EQ of humans—specifically the troodon. They had EQs about 6 times higher than the average dinosaur, had fingers to grasp, had small teeth, ate meat, and appeared to be social. Dale Russel claims that had the K-T extinction not occurred, the troodon would look similar to us with a brain size around 1100 cc (the size of erectus before he went extinct). This is what he believes the dinosauroid troodon would look like had they not died out 65 mya:

When interviewed about the dinosauroid he imagined, he stated:
The “dinosauroid” was a thought experiment, based on an observable, general trend toward larger relative brain size in terrestrial vertebrates through geologic time, and the energetic efficiency of an upright posture in slow-moving, bipedal animals. It seems to me that such speculation remains acceptable, particularly if directed toward non-anthropoid anatomical configurations. However, I very nearly decided not to publish the exercise because of the damaging effects it might have had on the credibility of my work in general. Most people remained polite, although there were hostile reactions from those with “ultra-quantitative” and “ultra-intuitive” world views.
Why does it look so human? Why does he assume that the ‘ideal body plan’ is what we have? It seems to be extremely biased towards a humanoid morphology, just as other reconstructions are biased towards what we think about certain areas today and how the people may have looked in our evolutionary past. Anthropocentric bias permeates deep in evolutionary thinking, this is one such example.
Thinking of this thought experiment of a possible ‘bipedal dinosauroid’ we need to be realistic in terms of thinking of its anatomy and morphology.
Let’s accept Russel’s contention as true; that troodontids or other ‘highly encephalized species’ reached a human EQ, as he notes, of 9.4, with troodontids at .34 (the highest), archaeopteryx at .32, triconodonts (early extinct mammal of the cretaceous) with a .29 EQ, and the diademodon with an EQ of .20 (Russel, 1983). Russel found that the troodontids had EQs 6 times higher than the average dinosaur, so from here, he extrapolated that the troodon would have had a brain our size. However, Stephen Jay Gould argued the opposite in Wonderful Life writing:
If mammals had arisen late and helped to drive dinosaurs to their doom, then we could legitimately propose a scenario of expected progress. But dinosaurs remained dominant and probably became extinct only as a quirky result of the most unpredictable of all events—a mass dying triggered by extraterrestrial impact. If dinosaurs had not died in this event, they would probably still dominate the large-bodied vertebrates, as they had for so long with such conspicuous success, and mammals would still be small creatures in the interstices of their world. This situation prevailed for one hundred million years, why not sixty million more? Since dinosaurs were not moving towards markedly larger brains, and since such a prospect may lay outside the capability of reptilian design (Jerison, 1973; Hopson, 1977), we must assume that consciousness would not have evolved on our planet if a cosmic catastrophe had not claimed the dinosaurs as victims. In an entirely literal sense, we owe our existence, as large reasoning mammals, to our lucky stars. (Gould, 1989: 318)
If a large brain was probably outside of reptilian design, then a dinosaur—or a descendant (troodon included)—would have never reached human-like intelligence. However, some people may say that dinosaur descendants may have evolved brains our size since birds have brains that lie outside of reptilian design (supposedly).
However, one of the most famous fossils ever found, archaeopteryx, was within reptilian design, having feathers and along with wings which would have been used for gliding (whether or not they flew is debated). Birds descend from therapods. Anchiornis, and other older species are thought to be the first birds. Most of birds’ traits, such as bipedal posture, hinged ankles, hollow bones and S-shaped neck in birds are derived features from their ancestors.
If we didn’t exist, then if any organism were to come close to our intelligence, I would bet that some corvids would, seeing as they have a higher packing density and interconnections compared to the “layered mammalian brain” (Olkowicz et al, 2016). Nick Lane, biochemist and author of the book The Vital Question: Evolution and the Origins of Complex Life believes a type of intelligent ocotopi may have evolved, writing:
Wind back the clock to Cambrian times, half a billion years ago, when mammals first exploded into the fossil record, and let it play forwards again. Would that parallel be similar to our own? Perhaps the hills would be crawling with giant terrestrial octopuses. (Lane, 2015: 21)
We exist because we are primates. Our brains are scaled-up primate brains (Herculano-Houzel, 2009). Our primate morphology—along with our diet, sociality, and culture—is also why we came to take over the world. Our body plan—which, as far as we know, only evolved once—is why we have the ability to manipulate our environment and use our superior intelligence—which is due to the number of neurons in our cerebral cortex, the highest in the animal kingdom, 16 billion in all (Herculano-Houzel, 2009). Why postulate that a dinosaur could have looked even anywhere close to us?
This is also ignoring the fact that decimation and diversification also ‘decide the fates’ so to speak, of the species on earth. Survival during an extinction event is strongly predicated by chance (and size). The smaller an organism is, the more likely it will survive an extinction event. Who’s to say that the troodon doesn’t go extinct due to an act of contingency, say, 50 mya if the K-T extinction never occurred?
In conclusion, the supposed ‘trend’ in brain size evolution is just random fluctuations—inevitabilities since life began at the left wall of complexity. Gould wrote about a drunkard’s walk in his book Full House (Gould, 1996) in which he illustrates an example of a drunkard walking away from a bar with the bar wall being the left wall of complexity and the gutter being the right wall. The gutter will always be reached; and if he hits the wall, he will lean against the wall “until a subsequent stagger propels him in the other direction. In other words, only one direction of movement remains open for continuous advance—toward the gutter” (Gould, 1996: 150).
I bring up this old example to illustrate but one salient point: In a system of linear motion structurally constrained by a wall at one end, random movement, with no preferred directionality whatever, will inevitably propel the average position away from a starting point at the wall. The drunkard falls into the gutter every time, but his motion includes no trend whatever toward this form of perdition. Similarly, some average or extreme measure of life might move in a particular direction even if no evolutionary advantage, and no inherent trend, favor that pathway (Gould, 1996: 151).
We humans are lucky we are here. Contingencies of ‘just history’ are why we are here, and if we were not here—if the K-T extinction never occurred—and the troodon or another dinosaur species survived to the present day, they would not have reached our ‘level’ of intelligence. To believe so is to believe in teleological evolution—which certainly is not true. Anthropometric bias runs deep in evolutionary biology and paleontology. People assume that since we are—according to some—the ‘pinnacle’ of evolution, that us, or something like us, would eventually have evolved.
Any ‘trends’ can be explained as life moving away from the left wall of complexity, with the left wall—the mode of life, the modal bacter-–being unchanged. We are at the extreme tail of the distribution of complexity while bacteria are at the left wall. Complex life was inevitable since bacteria, the most simple life, began at the left wall. And so, these ‘trends’ in brain size are just that, increasing complexity, not any type of ‘progressive evolution’. Evolution just happens, natural selection occurs based on the local environment, not any inherent or intrinsic ‘progress’.
References
Gould, S. J. (1989). Wonderful life: the burgess Shale and the nature of history. New York: Norton.
Gould, S. J. (1996). Full house: The Spread of Excellence from Plato to Darwin. New York: Harmony Books.
Herculano-Houzel, S. (2009). The human brain in numbers: a linearly scaled-up primate brain. Frontiers in Human Neuroscience,3. doi:10.3389/neuro.09.031.2009
Lane, N. (2015). The vital question: energy, evolution, and the origins of complex life. New York: W.W. Norton & Company.
Olkowicz, S., Kocourek, M., Lučan, R. K., Porteš, M., Fitch, W. T., Herculano-Houzel, S., & Němec, P. (2016). Birds have primate-like numbers of neurons in the forebrain. Proceedings of the National Academy of Sciences,113(26), 7255-7260. doi:10.1073/pnas.1517131113
Rushton J P (1997). Race, Evolution, and Behavior. A Life History Perspective (Transaction, New Brunswick, London).
Russell, D. A. (1983). Exponential evolution: Implications for intelligent extraterrestrial life. Advances in Space Research,3(9), 95-103. doi:10.1016/0273-1177(83)90045-5
Traumatic Brain Injury and IQ
1900 words
What is the relationship between traumatic brain injury (TBI) and IQ? Does IQ decrease? Stay the same? Increase? A few studies have looked at the relationship between TBI and IQ, and the results may be quite surprising to some. Tonight I will look through a few studies and see what the relationship is between TBI and IQ—does IQ decrease substantially or is there only a small decrease? Does it decrease for all subtests or only some?
TBI and IQ
In a sample of 72 people with TBI who had significant brain injuries had an average IQ of 90 (study 1; Bigler, 1995). Bigler also says that whatever correlation exists between brain size and IQ “does not persist post injury” (pg 387). This finding has large implications: can there be a minimal hit to IQ depending on age/severity of injury/brain size/education level?
As will be seen when I review another study on IQ and brain injury, every individual in the cohort in Bigler (1995) was tested after 42 days of brain injury. This does matter, as I will get into below.
Table 1 in study 1 shows that whatever positive relationship between IQ and brain size that is there before injury does not persist after injury (Bigler, 1995: 387). Study 1 showed that, even with mild-to-severe brain damage, there was little change in measured IQ—largely because the correlation between brain size and IQ is .51 at the high end (which I will use—the true correlation is between .24 [Pietschnig et al, 2015] to .4 [Rushton and Ankney, 2009]), this means that if the correlation were to be that high, brain size would only explain 25 percent of the variation in IQ (Skoyles, 1999). That leaves a lot of room for other reasons for differences in brain size and IQ in individuals and groups.
In study 2 (Bigler, 1995: 389-391), he looked into whether or not there were differences in IQ between high and low brain volume people (95 men). Results summed in table 3 (pg 390). Those with low brain volume (1185), aged 28, had an IQ of 82.61 while those with high brain volume (1584), aged 34 had an IQ of 92 (both cohorts had similar education). Bigler showed in study 1 IQ was maintained post injury, so we can say that this was their IQ preinjury.
In table 2, Bigler (1995) compares IQs and brain volumes of mild-to-moderate and moderate-to-severe individuals with TBI. Brain volume in the moderate-to-severe group was 1289.2 whereas for the mild-to-moderate TBI-suffering individuals had a mean brain volume of 1332.9. Amazingly, both groups had IQ scores in the normal range (90.0 for moderate-to-severe TBI and 90.7 for individuals suffering from mild-to-moderate TBI. In study 3, Bigler (1995) shows that trauma-induced atrophic changes in the brain aren’t related to IQ postinjury, nor to the amount of focal lesion volume.
Nevertheless, Bigler (1995) shows that those with bigger brains had less of a cognitive hit after TBI than those with smaller brains. PumpkinPerson pointed me to a study that shows that TBI stretches far back into our evolutionary history, with TBI seen in australopithecine fossils along with erectus fossils found throughout the world. This implies that TBI was a driver for brain size (Shivley et al, 2012); if the brain is bigger, then if/when TBI is acquired, the cognitive hit will be lessened (Stern, 2002). This is a great theory for explaining why we have large brains despite the negatives that come with them—if we were to acquire TBI in our evolutionary past, then the hit to our cognition would not be too great, and so we could still pass our genes to the next generation.
The fact that changes in IQ are minimal when brain damage is acquired shows that brain size isn’t as important as some brain-size-fetishists would like you to believe. Though, preinjury (PI) IQ was not tested, I have one study where it was.
Wood and Rutterford (2006) showed results similar to Bigler (1995)—minimal change to IQ occurs after TBI. The whole cohort pre-injury (PI) had a 99.79 IQ. T1 (early measure) IQ for the cohort was 90.96 while T2 (late measure) IQ for the cohort was 92.37. For people with greater than 11th-grade education (n=30), IQ decreased from 106.57 PI to 95.19 in T1 to 100.17 in T2. For people with less than an 11th-grade education (n=44), IQ PI was 95.16 and decreased to 86.99 in T1 and increased to 87.96 in T2. Male (n=51) and female (n=23) were similar, with male PI IQ being 99.04 to women’s 101.44 with a 90.13 IQ in T1 for men with a 90.72 IQ in T1 for women. In T2 for men it was 92.94 and for women, it was 92.83. So this cohort shows the same trends as Bigler (1995).
The most marked difference in subtests post-injury was in vocabulary (see table 3) with similarities staying the same, and digit symbol, and block design increasing between T1 and T2. Neither group differed between T1 and T2. The only significant association in performance change over time was years of education. Less educated people were at greater risk for cognitive decline (see table 2).
The difference for PI IQ after T2 for less educated people was 7.2 whereas for more educated people it was 6.4. Though more educated people gained back more IQ points between T1 and T2 (4.98 points) compared to less educated people (.97 IQ points). And: “The participants in our study represent a subgroup of patients with severe head injury reported in a larger study assessing long‐term psychosocial outcome.”
Bigler (1995) didn’t have PI IQ, but Wood and Rutterford (2006) did, and from T1 to T2 (Bigler 1995 tested what would be equivalent to T1 in the Wood and Rutterford 2006 study), IQ hardly increased for those with lower education (.97 points) but substantially increased for those with higher education (4.98 points) with there being a similar difference between PI IQ and T2 IQ for both groups.
Brain-derived neurotrophic protective factor (BDNF) also promotes survival and synaptic plasticity in the human brain (Barbey et al, 2014). They genotyped 156 Vietnam War soldiers with frontal lobe lesion and “focal penetrating head injuries” for the BDNF polymorphism. Though they did find differences in the groups with and without the BDNF polymorphism, writing that there were “substantial average differences between these groups in general intelligence (≈ half a standard deviation or 8 IQ points), verbal comprehension (6 IQ points), perceptual organization (6 IQ points), working memory (8 IQ points), and processing speed (8 IQ points) after TBI” (Barbey et al, 2014). This supports the hypothesis that BDNF is protective against TBI; and since BDNF was important in our evolutionary history which is secreted by the brain while endurance running (Raichlen and Polk, 2012), this could have also been another protective factor against hits to cognition that were acquired, say, during hunts or fights.
Nevertheless, one study found in a sample of 181 children Crowe et al (2012) found that children with mild-to-moderate TBI had IQ scores in the average range, whereas children with severe TBI had IQ scores in the low average range (80 to 90; table 3).
Infants with mild TBI had IQ scores of 99.9 (n=20) whereas infants with moderate TBI has IQs of 98.0 (n=23) and infants with severe TBI had IQs of 90.7 (n=7); preschoolers with mild TBI had IQ scores of 103.8 (n=11), whereas preschoolers with moderate TBI had IQ scores of 100.1 (n=19) and preschoolers with severe TBI had IQ scores of 85.8 (n=13); middle schoolers with mild TBI had IQ scores of 93.9 (n=10), whereas middle schoolers with moderate TBI had IQ scores of 93.5 (n=21), and middle schoolers with severe TBI had IQ scores of 86.1 (n=14); finally, children with mild TBI in late childhood had a mean FSIQ of 107.3 (n=17), while children with moderate TBI had IQs of 99.5 in late childhood (n=15), and children with severe TBI in late childhood had FSIQs of 94.7 (Crowe et al, 2012; table 3). This shows that age of acquisition and severity influence IQ scores (along with their subtests), and that brain maturity matters for maintaining average intelligence post-TBI. Königs et al (2016) also show the same trend; the outlook is better for children with mild TBI, while children faired far worse with severe TBI compared to mild when compared to adults (also seen in Crowe et al, 2012).
People who got into motor vehicle accidents suffered a loss of 14 IQ points (n=33) after being tested 20 months postinjury (Parker and Rosenblum, 1996). The WAIS-IV Technical and Interpretive Manual also shows a similar loss of 16 points (pg 111-112), however, the 22 subjects were tested within 6 to 18 months within acquiring their TBI, with no indication of whether or not a follow-up was done. IQ will recover postinjury, but education, brain size, age, and severity all are factors that contribute to how many IQ points will be gained. However, adults who suffer mild, moderate, and severe TBIs have IQs in the normal range. TBI severity also had a stronger effect on children aged 2 to 7 years of age at injury, with white matter volume and results on the Glasgow Coma Scale (which is used to assess consciousness after a TBI) were related to the severity of the injury (Levin, 2012).
Conclusion
TBI can occur with a minimal hit to IQ (Bigler, 1995; Wood and Rutterford, 2006; Crowe et al, 2012). IQs can still be in the average range at a wide range of ages/severities, however the older one is when they suffer a TBI, the more likely it is that they will incur little to no loss in IQ (depending on the severity, and even then they are still in the average range). It is interesting to note that TBI may have been a selective factor in our brain evolution over the past 3 million years from australopithecines to erectus to Neanderthals to us. However, the fact that people with severe TBI can have IQ scores in the normal range shows that the brain size/IQ correlation isn’t all it’s cracked up to be.
References
Barbey AK, Colom R, Paul E, Forbes C, Krueger F, Goldman D, et al. (2014) Preservation of General Intelligence following Traumatic Brain Injury: Contributions of the Met66 Brain-Derived Neurotrophic Factor. PLoS ONE 9(2): e88733. https://doi.org/10.1371/journal.pone.0088733
Bigler, E. D. (1995). Brain morphology and intelligence. Developmental Neuropsychology,11(4), 377-403. doi:10.1080/87565649509540628
Crowe, L. M., Catroppa, C., Babl, F. E., Rosenfeld, J. V., & Anderson, V. (2012). Timing of Traumatic Brain Injury in Childhood and Intellectual Outcome. Journal of Pediatric Psychology,37(7), 745-754. doi:10.1093/jpepsy/jss070
Green, R. E., Melo, B., Christensen, B., Ngo, L., Monette, G., & Bradbury, C. (2008). Measuring premorbid IQ in traumatic brain injury: An examination of the validity of the Wechsler Test of Adult Reading (WTAR). Journal of Clinical and Experimental Neuropsychology,30(2), 163-172. doi:10.1080/13803390701300524
Königs, M., Engenhorst, P. J., & Oosterlaan, J. (2016). Intelligence after traumatic brain injury: meta-analysis of outcomes and prognosis. European Journal of Neurology,23(1), 21-29. doi:10.1111/ene.12719
Levin, H. S. (2012). Long-term Intellectual Outcome of Traumatic Brain Injury in Children: Limits to Neuroplasticity of the Young Brain? Pediatrics, 129(2), e494–e495. http://doi.org/10.1542/peds.2011-3403
Parker, R. S., & Rosenblum, A. (1996). IQ loss and emotional dysfunctions after mild head injury incurred in a motor vehicle accident. Journal of Clinical Psychology,52(1), 32-43. doi:10.1002/(sici)1097-4679(199601)52:1<32::aid-jclp5>3.3.co;2-1
Pietschnig, J., Penke, L., Wicherts, J. M., Zeiler, M., & Voracek, M. (n.d.). Meta-Analysis of Associations Between Human Brain Volume And Intelligence Differences: How Strong Are They and What Do They Mean? SSRN Electronic Journal. doi:10.2139/ssrn.2512128
Raichlen, D. A., & Polk, J. D. (2012). Linking brains and brawn: exercise and the evolution of human neurobiology. Proceedings of the Royal Society B: Biological Sciences,280(1750), 20122250-20122250. doi:10.1098/rspb.2012.2250
Rushton, J. P., & Ankney, C. D. (2009). Whole Brain Size and General Mental Ability: A Review. The International Journal of Neuroscience, 119(5), 692–732. http://doi.org/10.1080/00207450802325843
Shively, S., Scher, A. I., Perl, D. P., & Diaz-Arrastia, R. (2012). Dementia Resulting From Traumatic Brain Injury: What Is the Pathology? Archives of Neurology, 69(10), 1245–1251. http://doi.org/10.1001/archneurol.2011.3747
Skoyles R. J. (1999) HUMAN EVOLUTION EXPANDED BRAINS TO INCREASE EXPERTISE CAPACITY, NOT IQ. Psycoloquy: 10(002) brain expertise
Stern, Y. (2002). What is cognitive reserve? Theory and research application of the reserve concept. Journal of the International Neuropsychological Society,8(03), 448-460. doi:10.1017/s1355617702813248
Wood, R. L., & Rutterford, N. A. (2006). Long‐term effect of head trauma on intellectual abilities: a 16‐year outcome study. Journal of Neurology, Neurosurgery, and Psychiatry, 77(10), 1180–1184. http://doi.org/10.1136/jnnp.2006.091553
Homo Erectus in America?
1800 words
Note: This article is high speculation based on the finding that occurred last week of the modification of mastodon bones in Ice Age California. If it is an actual archaeological site, along with being the age it’s purported to be, there are, in my opinion, only two possibilities for who could be responsible: erectus or the Denisova. Though I will cover evidence that Erectus did make it to America between 40-130,000ya, and rule out that Neanderthals are the hominid responsible.
It was discovered last week that there was human activity at an archeological site in San Diego, California, dated to about 130,000 years ago. Researchers discovered pieces of bone and teeth from a mastodon—that looked to have been modified by early humans. This discovery—if it shows that there was a hominid in the Americas 130,000ya—would have us rethink hominin migrations in the ancient past.
The bones and teeth show signs of having been modified by humans with “manual dexterity and experiential knowledge.” The same pattern was discovered in Nebraska and Kansas, where it was ruled out that carnivorous animals were responsible (Holen et al, 2017).
Now, we only have a few pieces of broken bone and some teeth from a mastodon. It is possible that ‘Natives’ dug up the mastodon skull and modified it, but I like to think outside of the box sometimes. When I first read the ScienceDaily article on the matter, the first hominin that popped into my head that could be responsible for this is erectus. But what is the evidence that he could have made it to the Americas that long ago?
Erectus in America
Evidence for erectus in America is scant. We have discovered no erectus skeletons in the Americas, and we only have a few pieces of bone to go off of to guess which hominid did this (and I doubt it was Homo sapiens or Neanderthals, I will explain my reasoning below).
I’ve been documenting on my blog for the past six months that, contrary to popular belief, erectus was not a ‘dumb ape’ and that, in fact, erectus had a lot of modern behaviors. If it turns out to be true that erectus made it to America, that wouldn’t really surprise me.

—Erectus territory
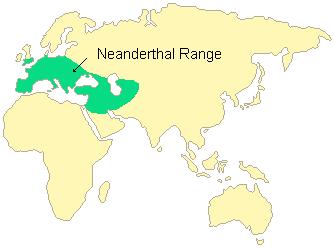
–Neanderthal territory
Erectus had a wider territory than the other hominid candidates (Neanderthals, Homo sapiens) and the other candidate—the Denisova—were situated more to the middle of the Asian continent. So this, really, leaves us only with erectus as the only possible candidate for the mysterious hominin in Caliofornia—and there is evidence that (albeit, extremely flimsy), erectus may have possibly made it to America, from a paper published back in 1986. Dreier (1986) writes that there is evidence of Man in America before 30kya, and if this is true, then it must be erectus since the estimated dates are between 50-70 kya—right around the time that AMH began migrating out of Africa. Dreier (1986) goes through a few different discoveries that could have been erectus in America, yet they were only modern skeletons. However, absence of evidence is not evidence of absence. (Though I will return to this specific point near the end of the article.)
How could erectus have possibly made it to America?
This is one of the most interesting things about this whole scenario. There is evidence that erectus made rafts. If erectus did make it to Flores (Stringer, 2004; Hardaker, 2007: 263-268; Lieberman, 2013)—eventually evolving into floresiensis (or from habilis or a shared common ancestor with habilis)—then he must have had the ability to make rafts. Since we have found erectus skulls at Java, and since certain bodily proportions of floresiensis are ‘scaled-down’ from erectus, along with tools that erectus used, it’s not out of the realm of possibility that erectus had the ability to navigate the seas.
One way that hominins can get to America is through the Bering strait. However, Dreier (1986) assumes that erectus was not cold-adapted, and insists that erectus could have only gone into higher latitudes for only a few months out of the year when it was warmer. As you can see from the above map of erectus’ territory, he lived along the coast of China and into some of the islands around SE Asia. While we don’t have any skeletal evidence, we can infer that it was late Asian erectus who, could have possibly, made it to the Americas. So since it was late in erectus’ evolution, we would expect him to have a large brain size in order to 1) survive in Africa and 2) since brain size predicts the success of a species in novel environments (Sol et al, 2008), erectus would have had a larger brain in these locations. So it seems that erectus did have the same adaptability that we do—especially if he actually did make it to the Americas.
Dreier (1986) posits that erectus could have traveled along the Aluetian island chain in Alaska, eating marine life (shells, mollusks, clams, etc), and so he would not have had to “deviate from the 53 north latitude vitamin D barrier drastically since almost the entire Aleutian Island chain falls between the 50 and 55 north latitude lines, and access via this route may have been possible during glaciation when sea levels in the area dropped as much as 100 meters” (Dreier, 1986: 31). Erectus could have gotten vitamin D from shells, mollusks and other marine life, as they are extremely high in vitamin D (Nair and Maseeh, 2012). I will contend that erectus rafted to America, but the Aluetian island route is also plausible.
Dreier (1986) ends up concluding that our best bet for finding erectus skeletons in America is along with Pacific coast, and there may be some submerged underwater. However, with the new discovery last week, I await more work into the site for some more answers (and of course questions).
However, contra Dreier’s (1986) claim that we should stop looking for sites with human activity earlier than 30,000 years, this new finding is promising.
Why not Neanderthals?
Neanderthals were seafarers, just like erectus, and later, us. However, there is evidence for Neanderthals sailing the seas 100kya, however, earlier dates of seafaring activity “as far back as 200 ka BP can not be excluded.” (Ferentinos et al, 2012). Further—and perhaps most importantly—the range of the Neanderthals was nowhere near the Pacific Ocean—whereas erectus was. So since there is little evidence of seafaring 200kya (which cannot be excluded), then we’re still left with the only possibility being erectus go to the Americas either by walking the Aleutian islands or rafting across the Pacific.
Could erectus have killed animals as large as a mastodon?
Erectus was killing elephants (Elephas antiquus) around 400kya in the Levant (Ben-Dor et al, 2011). Then, when the elephants went extinct, erectus had to hunt smaller, quicker game and thus evolved a smaller body to deal with the new environmental pressure—chasing a new food source. So erectus did have the ability to kill an animal that big, another positive sign that this is erectus we are dealing with in California 130,000 years ago.
An erectus skeleton in America?
An osteologist discovered a brow bone in the Americas, and in an unpublished report in 1990, he says the brow’s thickness and structure is comparable to African erectus, with a reanalysis showing it was closer to Asian erectus—just what we would expect since Asian erectus may have been a seafarer (Hardaker, 2007). However, the author of the book reiterates the Texas A&M osteologists’ findings writing: “these comparisons do not imply that preHomo sapiens were in the Americas” (Steen-McIntyre, 2008).
Humanlike cognition in erectus?
Humanlike thinking evolved 1.8 mya, right around the time erectus came into the picture (Putt et al, 2017). Volunteers created Auchulean tools while wearing a wearing a cap that measured brain activity. Visual attention and motor control were needed to create the “simpler Oldowan tools”, whereas for the “more complex Auchelian tools” a “larger portion of the brain was engaged in the creation of the more complex Acheulian tools, including regions of the brain associated with the integration of visual, auditory and sensorimotor information; the guidance of visual working memory; and higher-order action planning.” This discovery pushes back the advent of humanlike congition, since the earliest tools of this nature are found around 1.8 mya. There is a possibility that some erectus may have had IQs near ours, as studies of microcephalics show that a large amount have higher than average IQs (Skoyles, 1999).
Conclusion
Evidence is mounting that erectus was more than the ‘dumb ape’ that some people say he is. If erectus did make it to America—and the possibility is there—then human migratory patterns need to be rewritten. I hope there is more evidence pointing to what hominid was in the area at that time—and if there is evidence of humanlike activity there, it most likely is erectus. It is extremely possible that erectus could have gotten to America, as there is evidence that he was at least in northern China. So he could have sailed to the Americas or walked along the Aluetian islands.
The evidence for erectus in America is compelling, and I hope more is discovered about what went on at this site and who was there. Even if it wasn’t erectus, there is still some compelling evidence that he did make it to America.
References
Ben-Dor, M., Gopher, A., Hershkovitz, I., & Barkai, R. (2011). Man the Fat Hunter: The Demise of Homo erectus and the Emergence of a New Hominin Lineage in the Middle Pleistocene (ca. 400 kyr) Levant. PLoS ONE,6(12). doi:10.1371/journal.pone.0028689
Dreier, Frederick G., (1986). Homo Erectus in America: Possibilities and problems. Lambda Alpha Journal of Man, v.17, no.1-2, 1985-1986. Citing: Gifford, E.W., (1926). California Anthropometry. University of California Publications in Archaeology and Ethnology.22:217-390
Ferentinos, G., Gkioni, M., Geraga, M., & Papatheodorou, G. (2012). Early seafaring activity in the southern Ionian Islands, Mediterranean Sea. Journal of Archaeological Science,39(7), 2167-2176. doi:10.1016/j.jas.2012.01.032
Hardaker, C. (2007). The first American: the suppressed story of the people who discovered the New World. Franklin Lakes, NJ: New Page Books, a division of The Career Press.
Holen, S. R., Deméré, T. A., Fisher, D. C., Fullagar, R., Paces, J. B., Jefferson, G. T., . . . Holen, K. A. (2017). A 130,000-year-old archaeological site in southern California, USA. Nature,544(7651), 479-483. doi:10.1038/nature22065
Lieberman, D. (2013). The Story of the human body – evolution, health and disease. Penguin.
Nair, R., & Maseeh, A. (2012). Vitamin D: The “sunshine” vitamin. Journal of Pharmacology & Pharmacotherapeutics, 3(2), 118–126. http://doi.org/10.4103/0976-500X.95506
Putt, S. S., Wijeakumar, S., Franciscus, G. R., Spencer. P. J. The functional brain networks that underlie Early Stone Age tool manufacture. Nature Human Behaviour, 2017
Skoyles R. J. (1999) HUMAN EVOLUTION EXPANDED BRAINS TO INCREASE EXPERTISE CAPACITY, NOT IQ. Psycoloquy: 10(002) brain expertise
Sol, D., Bacher, S., Reader, S., & Lefebvre, L. (2008). Brain Size Predicts the Success of Mammal Species Introduced into Novel Environments. The American Naturalist,172(S1). doi:10.1086/588304
Steen-McIntyre, V. (2008) A Review of the Valsequillo, Mexico Early-Man Archaeological Sites (1962-2004) with Emphasis on the Geological Investigations of Harold E. Malde. Presentation at 2008 Geological Society of America Joint Annual Meeting Oct. 5-9, Houston, Texas
Stringer, C. (2004, October 27). A stranger from Flores. Retrieved May 09, 2017, from http://www.nature.com/news/2004/041027/full/news041025-3.html
The Evolution of Racial Pelvis Variation
1850 words
It’s well-known that blacks have narrower hips than whites (Rushton, 1997; Handa et al, 2008). These pelvic differences then account for part of the variation in elite sporting events such as sprinting and jumping (Entine, 2000). These pelvic differences are the result of climatic variation and sexual selection.
The evolution of the pelvis is due to bipedalism. We are bipeds because of our S-shaped spine, which helps us to cope with differing loads. The human pelvis had to evolve in two ways—to make birthing babies easier and to become more efficient for bipedal walking. Termed the ‘obstetric dilemma’, it has implications for osteoarthritis in both men and women (Hogervorst, Heinse, and de Vos, 2009). Having a more efficient bipedal gait meant the body could allocate energy to other parts of the body—mainly our growing brains/neuronal count. Over time, the brain grew while the pelvis had to shrink for more efficient bipedalism. The pelvis also got narrower in our evolution, being wider in Australopithicenes, while becoming more narrow when erectus appeared—which is the first instance of a humanlike pelvis in the fossil record—which increased how far we could travel as well as reduce our energy expenditure (Lieberman, et al, 2006). Further discussion can be found in my article Man the Athlete.
So we began evolving a narrower pelvis in comparison to our ancestors because it was more efficient for heat dissipation. Smaller trunks are more efficient for heat dissipation (Lieberman, 2015), whereas wider trunks are more efficient for thermoregulation in colder climes (Weaver and Hublin, 2008; Weaver, 2009; Gruss and Schmidt, 2015). Now, simply applying this logic to Eurasians and Africans (I am grouping East Asians and Europeans together since they were a single breeding population up until about 23,000-6,500ya), we can see one reason why that population has wider pelves than Africans.
When anatomically modern humans (AMH) left Africa between 50-100kya, human skeletal morphology was just like modern-day Africans’ today. When Man migrated into northerly climes, however, a wider pelvis was needed to retain heat in colder climes (Gruss and Schmidt, 2015). So, along with a wider pelvis evolving due to climatic demands on the body, as we migrated north the human brain expanded due to the climate of the area, along with expanding the pelvis to better thermoregulate (which a bigger brain also does in northerly climes). I did argue two months back (and added to Skoyles’ (1999) theory) that brain size increased for expertise capacity and not IQ since Arctic people needed more tools, as well as tools that were more complex, in comparison to peoples who evolved in a hotter climate. So selection then occurred for larger brains and pelvis due to the demand for thermoregulation and bigger brains—which then led to earlier births and more helpless babes, which higher levels of intelligence were then needed to care for them (Piantadosi and Kidd, 2016). The helplessness of infants predicts the intelligence of adults in the primate genera (Piantadosi and Kidd, 2016), so I will assume that this holds within primate species as well (I am not able to locate a citation that this doesn’t hold within the primate genera; if I am in error, please provide a citation). Since African children are born earlier and more mature than Eurasian children who are born slightly later and more helpless/less developed, this is one reason why Eurasians have higher levels of intelligence than Africans (which is independent of any direct effects of climate I may add!).
So since Eurasians needed a larger brains to make more tools in the Arctic/colder climes, their brains needed to expand in size for increased expertise capacity, which would then have further selected for wider pelves in Eurasian women. Climatic variation caused the wider hips/bigger brains in Eurasians, which then allowed the evolution of larger brains in comparison to those who remained in Africa.
Finally, the obstetric dilemma has been recently called into question; there is evidence that a wider pelvis does not increase locomotor costs in humans (Warrener et al, 2015), a treadmill tracked their gait, as well as the motion of their pelvis. This study is used as evidence that the obstetric dilemma is wrong—they argue that there is no trade-off between narrower hips in men and wider hips in women. However, as the authors point out, all subjects in the study walked/ran at the same speed. Let’s say that the speed was heightened; do you think the women/men with wider pelves would have had the same locomotor costs as the men/women with narrower pelves? The answer is, obviously, no.
The pelvis of all of the races of Man has evolved the way they are due to environmental/climatic demands. A wider pelvis is better for thermoregulation in colder climates, while a narrower pelvis/body is more efficient for heat loss (Gruss and Schmidt, 2015).
Thus, we can look at the evolution of brain size/pelvic size in a few ways: 1) The amount of tools/complexity of the tools in the area that led to a need for an increase in brain size for more ‘chunks’ (Gobet and Simon, 1998), which then—along with colder climates—selected for larger brains and a wider body/pelvis which made birthing babes with large heads/brains easier along with helping to conserve heat due to the wider body (Gruss and Schmidt, 2015); 2) Since people in higher altitudes needed a high amount of expertise to survive, further selection for bigger brains, wider pelves occurred because of this; 3) Africans have smaller pelves in comparison to Eurasians because they evolved in hotter climes and didn’t have the amount of tools that peoples in more northerly climes did—which also increased brain size; 4) putting this all together, we can say that because Africans live in hotter climates, they need narrow pelves in order to lose body heat; Eurasians, after they migrated into more northerly climes, needed a wider body/pelvis in order to retain heat. When Man migrated north, he needed the ability to become an expert in, say, tool-making and thus needed a bigger brain for more informational chunks (Simon and Gobet, 1998; Skoyles, 1999). Due to this, Eurasians have wider pelves since they needed larger brains for a higher expertise capacity (Skoyles, 1999).
When Man migrated north, he needed the ability to become an expert in, say, tool-making and thus needed a bigger brain for more informational chunks (Simon and Gobet, 1998; Skoyles, 1999). Due to this, Eurasians have wider pelves than Africans; so they can birth larger-brained children. The width of the female pelvis, too, was shaped by sexual selection (Lassek and Gaulin, 2009). Therefore, the evolution of the modern pelvis in human populations comes down to climatic variation, which, in turn, affects how large of a brain the babe is able to have. Climate constrains brain size in either ‘direction’, big or small. We don’t even need to look at the variation within modern Homo sapiens to see the pattern in pelvic size we do today; because the pelvic differences noted among Man definitely were in effect millions of years ago, with hominids in colder climates having wider pelves while hominids in warmer climates had narrower pelves.
Along with everything above, the evolution of the human pelvis has a few implications for the human races today. Some recent studies have shown that there is no obstetric dilemma at all, with birth complications being caused by babies with higher weights than in our ancestral past, due to environmental mismatches causing higher-weight babies (Warrener et al, 2015; Betti, 2017), which was also beneficial for the evolution of our large brains (Cunnane and Crawford, 2003) with the largest amount of cortical neurons in the animal kingdom. However, marked differences in locomotion would be seen in people who had wide pelves compared to narrow pelves; which is what we see in elite running competitions: the elite runners have narrower pelves. So wider pelves don’t impede normal bipedal walking, but it does impede being able to efficiently run, as evidenced in participants of elite sprinting and marathon competitions. Looking at champion athletes and studying their locomotion (along with other traits as I’ve covered here) you can see that those with narrower pelves win more competitions than those with wider pelves (and happen to have different muscle fiber competition, fat distribution/percent, and morphology).
Racial differences in the pelvis explain the reasons behind why a certain race dominates in certain elite competitions; it largely comes down to skeletal morphology. These skeletal differences have evolutionary underpinnings, with the same pelvic differences seen in hominins that evolved in colder/warmer climates in the past. These pelvic differences (along with body fat percentage/distribution, musculoskeletal morphology, muscle fiber type, lean mass percentage, lower Vo2 max, poorer running economy, a larger Q-angle [4.6 degrees greater than men], etc) are why women are less efficient runners. People with wider hips are more likely to have be endomorphic while people with narrower hips are more likely to be ecto and meso. Not surprisingly, people from northerly climes consistently win WSM competitions whereas East and West Africans dominate bodybuilding and sprinting/marathons due to having a narrower pelvis and other advantageous morphological traits that lead to success in the sport. Nevertheless, pelvic differences between the races largely come down to differences in climate, which was also seen in ancient hominins. These pelvic differences further lead to racial differences in elite sporting competition.
References
Betti, L. (2017). Human Variation in Pelvic Shape and the Effects of Climate and Past Population History. The Anatomical Record,300(4), 687-697. doi:10.1002/ar.23542
Cunnane, S. C., & Crawford, M. A. (2003). Survival of the fattest: fat babies were the key to evolution of the large human brain. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology,136(1), 17-26. doi:10.1016/s1095-6433(03)00048-5
Dr. John R. Skoyles (1999) HUMAN EVOLUTION EXPANDED BRAINS TO INCREASE EXPERTISE CAPACITY, NOT IQ. Psycoloquy: 10(002) brain expertise
Entine, J. (2000). Taboo: why Black athletes dominate sports and why we are afraid to talk about it. New York: PublicAffairs.
Gobet, F., & Simon, H. A. (1998). Expert Chess Memory: Revisiting the Chunking Hypothesis. Memory,6(3), 225-255. doi:10.1080/741942359
Gruss, L. T., & Schmitt, D. (2015). The evolution of the human pelvis: changing adaptations to bipedalism, obstetrics and thermoregulation. Philosophical Transactions of the Royal Society B: Biological Sciences,370(1663), 20140063-20140063. doi:10.1098/rstb.2014.0063
Hogervorst, T., Heinse W.B., & de Vos J., (2009) Evolution of the hip and pelvis. Acta Orthopaedica, 80:sup336, 1-39, DOI: 10.1080/17453690610046620
Lieberman, D. E., Raichlen, D. A., Pontzer, H., Bramble, D. M., & Cutright-Smith, E. (2006). The human gluteus maximus and its role in running. Journal of Experimental Biology,209(11), 2143-2155. doi:10.1242/jeb.02255
Lieberman, D. E. (2015). Human Locomotion and Heat Loss: An Evolutionary Perspective. Comprehensive Physiology, 99-117. doi:10.1002/cphy.c140011
Piantadosi, S. T., & Kidd, C. (2016). Extraordinary intelligence and the care of infants. Proceedings of the National Academy of Sciences,113(25), 6874-6879. doi:10.1073/pnas.1506752113
Rushton J P (1997). Race, Evolution, and Behavior. A Life History Perspective (Transaction, New Brunswick, London).
Handa, V. L., Lockhart, M. E., Fielding, J. R., Bradley, C. S., Brubakery, L., Cundiffy, G. W., … Richter, H. E. (2008). Racial Differences in Pelvic Anatomy by Magnetic Resonance Imaging. Obstetrics and Gynecology, 111(4), 914–920.
Warrener, A. G., Lewton, K. L., Pontzer, H., & Lieberman, D. E. (2015). A Wider Pelvis Does Not Increase Locomotor Cost in Humans, with Implications for the Evolution of Childbirth. PLoS ONE, 10(3), e0118903.
Weaver, T. D., & Hublin, J. (2009). Neandertal birth canal shape and the evolution of human childbirth. Proceedings of the National Academy of Sciences,106(20), 8151-8156. doi:10.1073/pnas.0812554106
Weaver, T. D. (2009). The meaning of Neandertal skeletal morphology. Proceedings of the National Academy of Sciences,106(38), 16028-16033. doi:10.1073/pnas.0903864106
