
Vegans/Vegetarians vs. Carnivores and the Neanderthal Diet
2050 words
The vegan/vegetarian-carnivore debate is one that is a false dichotomy. Of course, the middle ground is eating both plants and animals. I, personally, eat more meat (as I eat a high protein diet) than plants, but the plants are good for a palate-switch-up and getting other nutrients in my diet. In any case, on Twitter, I see that there is a debate between “carnivores” and “vegans/vegetarians” on which diet is healthier. I think the “carnivore” diet is healthier, though there is no evolutionary basis for the claims that they espouse. (Because we did evolve from plant-eaters.) In this article, I will discuss the best argument for ethical vegetarianism and the evolutionary basis for meat-eating.
Veganism/Vegetarianism
The ethical vegetarian argument is simple: Humans and non-human animals deserve the same moral consideration. Since they deserve the same moral consideration and we would not house humans for food, it then follows that we should not house non-human animals for food. The best argument for ethical vegetarianism comes from Peter Singer from Unsanctifying Animal Life. Singer’s argument also can be extended to using non-human animals for entertainment, research, and companionship.
Any being that can suffer has an interest in avoiding suffering. So the equal consideration of interests principle (Guidi, 2008) asserts that the ability to suffer applies to both human and non-human animals.
Here is Singer’s argument, from Just the Arguments: 100 of the Most Important Arguments in Western Philosophy (pg. 277-278):
P1. If a being can suffer, then that being’s interests merit moral consideration.
P2. If a being cannot suffer, then that beings interests do not merit moral consideration.
C1. If a being’s interests merit moral consideration, then that being can suffer (transposition, P2).
C2. A being’s interests merit moral consideration if and only if that being can suffer (material equivalence, P1, C1).
P3. The same interests merit the same moral consideration, regardless of what kind of being is the interest-bearer (equal consideration of interests principle).
P4. If one causes a being to suffer without adequate justification, then one violates that being’s interests.
P5. If one violates a being’s interests, then one does what is morally wrong.
C3. If one causes a being to suffer without adequate justification, then one does what is morally wrong (hypothetical syllogism, P4, P5).
P6. If P3, then if one kills, confines, or causes nonhuman animals to experience pain in order to use them as food, then one causes them to suffer without adequate justification.
P7. If one eats meat, then one participates in killin, confining, and causing nonhuman animals to experience pain in order to use them as food.
C4. If one eats mea, then one causes nonhuman animals to suffer without adequate justification (hypothetical syllogism, P6, P7).
C5. If one eats meat, the one does what is morally wrong (hypothetical syllogism, C3, C4).
This argument is pretty strong, indeed it is sound. However, I personally will never eat a vegetarian/vegan diet because I love eating meat too much. (Steak, turkey, chicken.) I will do what is morally wrong because I love the taste of meat.
In an evolutionary context, the animals we evolved from were plant-eaters. The amount of meat in our diets grew as we diverged from our non-human ancestors; we added meat through the ages as our tool-kit became more complex. Since the animals we evolved from were plant-eaters and we added meat as time went on, then, clearly, we were not “one or the other” in regard to diet—our diet constantly changed as we migrated into new biomes.
So although Singer’s argument is sound, I will never become a vegan/vegetarian. Fatty meat tastes too good.
Nathan Cofnas (2018) argues that “we cannot say decisively that vegetarianism or veganism is safe for children.” This is because even if the vitamins and minerals not gotten through the diet are supplemented, the bioavailability of the consumed nutrients are lower (Pressman, Clement, and Hayes, 2017). Furthermore, pregnant women should not eat a vegan/vegetarian diet since vegetarian diets can lead to B12 and iron deficiency along with low birth weight and vegan diets can lead to DHZ, zinc, and iron deficiencies along with a higher risk of pre-eclampsia and inadequate fetal brain development (Danielewicz et al, 2017). (See also Tan, Zhao, and Wang, 2019.)
Carnivory
Meat was important to our evolution, this cannot be denied. However, prominent “carnivores” take this fact and push it further than it goes. Yes, there is data that meat-eating allowed our brains to grow bigger, trading-off with body size. Fonseca-Azevedo and Herculano-Houzel (2012) showed that metabolic limitations resulting from hours of feeding and low caloric yield explain the body/brain size in great apes. Plant foods are low in kcal; great apes have large bodies and so, need to eat a lot of plants. They spend about 10 to 11 hours per day feeding. On the other hand, our brains started increasing in size with the appearance of erectus.
If erectus ate nothing but raw foods, he would have had to eat more than 8 hours per day while hominids with neurons around our level (about 86 billion; Herculano-Houzel, 2009). Thus, due to the extreme difficulty of attaining the amount of kcal needed to power the brains with more neurons, it is very unlikely that erectus would have been able to survive on only plant foods while eating 8+ hours per day. Indeed, with the archaeological evidence we have about erectus, it is patently ridiculous to claim that erectus did eat for that long. Great apes mostly graze all day. Since they graze all day—indeed, they need to as the caloric availability of raw foods is lower than in cooked foods (even cooked plant foods would have a higher bioavailability of nutrients)—then to afford their large bodies they need to basically do nothing but eat all day.
It makes no sense for erectus—and our immediate Homo sapiens ancestors—to eat nothing but raw plant foods for what amounts to more than a work day in the modern world. If this were the case, where would they have found the time to do everything else that we have learned about them in the archaeological record?
There is genetic evidence for human adaptation to a cooked diet (Carmody et al, 2016). Cooking food denatures the protein in it, making it easier to digest. Denaturation is the alteration of the protein shape of whatever is being cooked. Take the same kind of food. That food will have different nutrient bioavailability depending on whether or not it is cooked. This difference, Herculano-Houzel (2016) and Wrangham (2009) argue is what drove the evolution of our genus and our big brains.
Just because meat-eating and cooking was what drove the evolution of our big brains—or even only allowed our brains to grow bigger past a certain point—does not mean that we are “carnivores”; though it does throw a wrench into the idea that we—as in our species Homo sapiens—were strictly plant-eaters. Our ancestors ate a wide-range of foods depending on the biome they migrated to.
The fact that our brain takes up around 20 percent of our TDEE while representing only 2 percent of our overall body mass, the reason being our 86 billion neurons (Herculano-Houzel, 2011). So, clearly, as our brains grew bigger and acquired more neurons, there had to have been a way for our ancestors to acquire the energy need to power their brains and neurons and, as Fonseca-Azevedo and Herculano-Houzel (2012) show, it was not possible on only a plant diet. Eating and cooking meat was the impetus for brain growth and keeping the size of our brains.
Take this thought experiment. An asteroid smashes into the earth. A huge dust cloud blocks out the sun. So the asteroid would have been a cause of lowering food production. This halting of food production—high-quality foods—persisted for hundreds of years. What would happen to our bodies and brains? They would, of course, shrink depending on how much and what we eat. Food scarcity and availability, of course, do influence the brain and body size of primates (Montgomery et al, 2010), and humans would be no different. So, in this scenario I have concocted, in such an event, we would shrink, in both brain and body size. I would imagine in such a scenario that high-quality foods would disappear or become extremely hard to come by. This would further buttress the hypothesis that a shift to higher-quality energy is how and why our large brains evolved.
Neanderthal Diet
A new analysis of the tooth of a Neanderthal apparently establishes that they were mostly carnivorous, living mostly on horse and reindeer meat (Jaouen et al, 2019). Neanderthals did indeed have a high-meat diet in northerly latitudes during the cold season. Neanderthals in Southern Europe—especially during the warmer seasons—however, ate a mixture of plants and animals (Fiorenza et al, 2008). Further, there was a considerable plant component to the diet of Neanderthals (Perez-Perez et al, 2003) (with the existence of plant-rich diets for Neanderthals being seen mostly in the Near East; Henry, Brooks, and Piperno, 2011) while the diet of both Neanderthals and Homo sapiens varied due to climatic fluctuations (El Zataari et al, 2016). From what we know about modern human biochemistry and digestion, we can further make the claim that Neanderthals ate a good amount of plants.
Ulijaszek, Mann, and Elton (2013: 96) write:
‘Absence of evidence’ does not equate to ‘evidence of absence,’ and the meat-eating signals from numerous types of data probably swamp the plant-eating signlas for Neanderthals. Their dietary variability across space and time is consistent with the pattern observed in the hominin clade as a whole, and illustrates hominin dietary adaptatbility. It also mirrors trends observed in modern foragers, whereby those populations that live in less productive environments have a greater (albeit generally not exclusive) dependance on meat. Differences in Neanderthal and modern human diet may have resulted from exploitation of different environments: within Europe and Asia, it has been argued that modern humans exploited marginal areas, such as steppe environments, whereas Neanderthals may have preferred more mosaic, Mediterranean-type habitats.
Quite clearly, one cannot point to any one study to support an (ideologically driven) belief that our genus or Neanderthals were “strictly carnivore”, as there was great variability in the Neanderthal diet, as I have shown.
Conclusion
Singer’s argument for ethical vegetarianism is sound; I personally can find no fault in it (if anyone can, leave a comment and we can discuss it, I will take Singer’s side). Although I can find no fault in the argument, I would never become a vegan/vegetarian as I love meat too much. There is evidence that vegan/vegetarian diets are not good for growing children and pregnant mothers, and although the same can be said for any type of diet that leads to nutrient deficiencies, the risk is much higher in these types of plant-based diets.
The evidence that we were meat-eaters in our evolutionary history is there, but we evolved as eclectic feeders. There was great variability in the Neanderthal diet depending on where they lived, and so the claim that they were “full-on carnivore” is false. The literature attests to great dietary flexibility and variability in both Homo sapiens and Neanderthals, so the claim that they ate meat and only meat is false.
My conclusion in my look into our diet over evolutionary time was:
It is clear that both claims from vegans/vegetarians and carnivores are false: there is no one “human diet” that we “should” be eating. Individual variation in different physiologic processes implies that there is no one “human diet”, no matter what type of food is being pushed as “what we should be” eating. Humans are eclectic feeders; we will eat anything since “Humans show remarkable dietary flexibility and adaptability“. Furthermore, we also “have a relatively unspecialized gut, with a colon that is shorter relative to overall size than in other apes; this is often attributed to the greater reliance on faunivory in humans (Chivers and Langer 1994)” (Ulijaszek, Mann, and Elton, 2013: 58). Our dietary eclectism can be traced back to our Australopithecine ancestors. The claim that we are either “vegetarian/vegan or carnivore” throughout our evolution is false.
There is no evidence for both of these claims from both of these extreme camps; humans are eclectic feeders. We are omnivorous, not vegan/vegetarian or carnivores. Although we did evolve from plant-eating primates and then added meat into our diets over time, there is no evidence for the claim that we ate only meat. Our dietary flexibility attests to that.
Athleticism is Irreducible to Biology: A Systems View of Athleticism
1550 words
Reductionists would claim that athletic success comes down to the molecular level. I disagree. Though, of course, understanding the molecular pathways and how and why certain athletes excel in certain sports can and will increase our understanding of elite athleticism, reductionist accounts do not tell the full story. A reductionist (which I used to be, especially in regard to sports; see my article Racial Differences in Muscle Fiber Typing Cause Differences in Elite Sporting Competition) would claim that, as can be seen in my article, the cause for elite athletic success comes down to the molecular level. Now, that I no longer hold such reductionist views in this area does not mean that I deny that there are certain things that make an elite athlete. However, I was wrong to attempt to reduce a complex bio-system and attempt to pinpoint one variable as “the cause” of elite athletic success.
In the book The Genius of All of Us: New Insights into Genetics, Talent, and IQ, David Shenk dispatches with reductionist accounts of athletic success in the 5th chapter of the book. He writes:
2. GENES DON’T DIRECTLY CAUSE TRAITS; THEY ONLY INFLUENCE THE SYSTEM.
Consistent with other lessons of GxE [Genes x Environment], the surprising finding of the $3 billion Human Genome Project is that only in rare instances do specific gene variants directly cause specific traits or diseases. …
As the search for athletic genes continues, therefore, the overwhelming evidence suggests that researchers will instead locate genes prone to certain types of interactions: gene variant A in combination with gene variant B, provoked into expression by X amount of training + Y altitude + Z will to win + a hundred other life variables (coaching, injuries, etc.), will produce some specific result R. What this means, of course, What this means, of course, is that we need to dispense rhetorically with thick firewall between biology (nature) and training (nurture). The reality of GxE assures that each persons genes interacts with his climate, altitude, culture, meals, language, customs and spirituality—everything—to produce unique lifestyle trajectories. Genes play a critical role, but as dynamic instruments, not a fixed blueprint. A seven- or fourteen- or twenty-eight-year-old is not that way merely because of genetic instruction. (Shenk, 2010: 107) [Also read my article Explaining African Running Success Through a Systems View.]
This is looking at the whole entire system: genes, to training, to altitude, to will to win, to numerous other variables that are conducive to athletic success. You can’t pinpoint one variable in the entire system and say that that is the cause: each variable works together in concert to produce the athletic phenotype. One can invoke Noble’s (2012) argument that there is no privileged level of causation in the production of an athletic phenotype. There are just too many factors that go into the production of an elite athlete, and attempting to reduce it to one or a few factors and attempt to look for those factors in regard to elite athleticism is a fool’s errand. So we can say that there is no privileged level of causation in regard to the athletic phenotype.
In his paper Sport and common-sense racial science, Louis (2004: 41) writes:
The analysis and explanation of racial athleticism is therefore irreducible to
biological or socio-cultural determinants and requires a ‘biocultural approach’
(Malina, 1988; Burfoot, 1999; Entine, 2000) or must account for environmental
factors (Himes, 1988; Samson and Yerl`es, 1988).
Reducing anything, sports included, to environmental/socio-cultural determinants and biology doesn’t make sense; I agree with Louis that we need a ‘biocultural approach’, since biology and socio-cultural determinants are linked. This, of course, upends the nature vs. nurture debate; neither “nature” nor “nurture” has won, they causally depend on one another to produce the elite athletic phenotype.
Louis (2004) further writes:
In support of this biocultural approach, Entine (2001) argues that athleticism is
irreducible to biology because it results from the interaction between population-based genetic differences and culture that, in turn, critiques the Cartesian dualism
‘which sees environment and genes as polar-opposite forces’ (p. 305). This
critique draws on the centrality of complexity, plurality and fluidity to social
description and analysis that is significant within multicultural common sense. By
pointing to the biocultural interactivity of racial formation, Entine suggests that
race is irreducible to a single core determinant. This asserts its fundamental
complexity that must be understood as produced through the process of
articulation across social, cultural and biological categories.
Of course, race is irreducible to a single core determinant; but it is a genuine kind in biology, and so, we must understand the social, cultural, and biological causes and how they interact with each other to produce the athletic phenotype. We can look at athlete A and see that he’s black and then look at his somatotype and ascertain that the reason why athlete A is a good athlete is conducive to his biology. Indeed, it is. One needs a requisite morphology in order to succeed in a certain sport, though it is quite clearly not the only variable needed to produce the athletic phenotype.
One prevalent example here is the Kalenjin (see my article Why Do Jamaicans, Kenyans, and Ethiopians Dominate Running Competitions?). There is no core determinant of Kalenjin running success; even one study I cited in my article shows that Germans had a higher level of a physiological variable conducive to long-distance running success compared to the Kalenjin. This is irrelevant due to the systems view of athleticism. Low Kenyan BMI (the lowest in the world), combined with altitude training (they live in higher altitudes and presumably compete in lower altitudes), a meso-ecto somatotype, the will to train, and even running to and from where they have to go all combine to show how and why this small tribe of Kenyans excel so much in these types of long-distance running competitions.
Sure, we can say that what we know about anatomy and physiology that a certain parameter may be “better” or “worse” in the context of the sport in question, no one denies that. What is denied is the claim that athleticism reduces to biology, and it does not reduce to biology because biology, society, and culture all interact and the interaction itself is irreducible; it does not make sense to attempt to partition biology, society, and culture into percentage points in order to say that one variable has primacy over another. This is because each level of the system interacts with every other level. Genes, anatomy and physiology, the individual, the overarching society, cultural norms, peers, and a whole slew of other factors explain athletic success not only in the Kalenjin but in all athletes.
Broos et al (2016) showed that in those with the RR genotype, coupled with the right morphology and fast twitch muscle fibers, this would lead to more explosive contractions. Broos et al (2016) write:
In conclusion, this study shows that a-actinin-3 deficiency decreases the contraction velocity of isolated type IIa muscle fibers. The decreased cross-sectional area of type IIa and IIx fibers may explain the increased muscle volume in RR genotypes. Thus, our results suggest that, rather than fiber force, combined effects of morphological and contractile properties of individual fast muscle fibers attribute to the enhanced performance observed in RR genotypes during explosive contractions.
This shows the interaction between the genotype, morphology, fast twitch fibers (which blacks have more of; Caeser and Henry, 2015), and, of course, the grueling training these elite athletes go through. All of these factors interact. This further buttresses the argument that I am making that different levels of the system causally interact with each other to produce the athletic phenotype.
Pro-athletes also have “extraordinary skills for rapidly learning complex and neutral dynamic visual scenes” (Faubert, 2013). This is yet another part of the system, along with other physical variables, that an elite athlete needs to have. Indeed, as Lippi, Favalaro, and Guidi (2008) write:
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.
So now we have: (1) strong mental background; (2) genes; (3) morphology; (4) Vo2 max; (5) altitude; (6) will to win; (7) training; (8) coaching; (9) injuries; (10) peer/familial support; (11) fiber typing; (12) heart strength etc. There are of course myriad other variables that are conducive to athletic success but are irreducible since we need to look at it in the whole context of the system we are observing.
In conclusion, athleticism is irreducible to biology. Since athleticism is irreducible to biology, then to explain athleticism, we need to look at the whole entire system, from the individual all the way to the society that individual is in (and everything in between) to explain how and why athletic phenotypes develop. There is no logical reason to attempt to reduce athleticism to biology since all of these factors interact. Therefore, the systems view of athleticism is the way we should view the development of athletic phenotypes.
(i) Nature and Nurture interact.
(ii) Since nature and nurture interact, it makes no sense to attempt to reduce anything to one or the other.
(iii) Since it makes no sense to attempt to reduce anything to nature or nurture since nature and nurture interact, then we must dispense with the idea that reductionism can causally explain differences in athleticism between individuals.
Race and Menarche
1100 words
Back in 2016 I wrote about racial differences in menarche and how there is good evidence that leptin is a strong candidate for the cause in my article Leptin and its Role in the Sexual Maturity of Black Girls (disregard the just-so stories). Black girls are more likely to hit puberty at a younger age than white girls. Why? One reason may be that leptin may play a role in the accelerated growth and maturation of black girls, since there was a positive relationship between leptin concentration and obesity in black girls (Wagner and Heyward, 2000). When girls start to develop at younger and younger ages, a phrase you hear often is “It’s the chemicals in the food” in regard to, for example, early breast development on a young, pre-teen girl.
Black girls are more likely to be obese than white girls (Freedman et al, 2000) and it is thought that body fat permits the effects of earlier menarche due to leptin being released from the adipocyte (fat cell) (Salsberry, Reagen, and Pajer, 2010). Freedman et al (2000) showed that black girls experienced menarche 3 months earlier than white girls on average, while over a 20 year period the median age decreased by 9.5 months. There is also evidence of earlier thelarche (breast development) in black girls, which was mediated by gonadotropin (Cabrera et al, 2014). Wong et al (1998) found that circulating serum leptin levels were correlated with earlier menarche in black girls which was related to body fatness and age but lessened after fat mass, maturation and physical fitness. There is a ton of evidence that exists that body fatness is related to obesity and, as I said above, the mechanism is probably fat cells releasing leptin, permitting earlier menarche (see Kaplowitz, 2008). Higher levels of body fat cause earlier menarche; earlier menarche does not cause higher levels of body fat. The evidence is there that leptin indeed plays the permissive role to allow a girl to enter into puberty earlier, and that this is how and why black girls enter menarche earlier than white girls.
So when fat mass increases, so does leptin; when leptin increases, girls have puberty at an earlier age (Apter, 2009). Black girls have higher levels of circulating leptin than white girls (Ambrosious et al, 1998). So knowing the relationship between leptin and obesity and how fat cells release leptin into the body permissing earlier puberty, we can confidently say that leptin is a major cause of earlier pubertal development in black girls. Total body fat correlates with fasted leptin (Ebenibo et al, 2018).
Siervogel et al (2003) write:
A negative relationship between age at menarche, BMI and body fatness in girls has been shown [63, 64, 65]. It is still unclear, however, whether increased early childhood adiposity induces an earlier onset of puberty, if rapid maturation and early puberty induce an increase in body fat later in life, or whether both of these phenomena occur.
The average age of menarche in black girls was 12 years of age whereas for white girls it was 12.5 (Regan et al, 2013). Since we now know the causes of earlier menarche, we can talk about ultimate causation.
Since menarche is correlated with obesity and circulating leptin levels, then food quality would be an easy culprit to look at.
Deardorff et al (2014) write:
Three sets of findings stood out. One, grandmother’s lower education was related to later menarche for Black girls. Two, mother’s unmarried status (at birth and age 7) was associated with earlier menarche for Hispanics and Whites, but not for Blacks. Three, family income at child’s birth was related to earlier menarche for Blacks and Hispanics, but not Whites.
Lower family income at birth was associated with earlier menarche for Blacks (and to some extent for Hispanics), but not for white girls.
Deardorff et al’s (2014) study suggests that social determinants of health can be intervened upon and differences in pubertal timing can be ameliorated since they are driven by social factors.
Black children have higher levels of insulino-glucose ratios than white children, even after adjusting for confounds (Wong et al, 1999). There were similar findings when comparing normal-weight black and white girls matched for age, bone age, weight, and BMI. Black girls also had higher levels of insulin than white girls. Black girls grow faster than white girls beginning at 2 years of age, with the cause hypothesized to be “higher serum insulin concentrations in healthy African American girls suppress the hepatic production of IGFBP-1, which results in higher circulating concentrations of free IGF-I, contributing to the accelerated growth observed in African American girls compared with their white counterparts” (Wong et al, 1999: 297).
Wong et al (1999) found that black girls were older and more sexually mature than white girls according to the Tanner stages of development (using pubic hair as a measure). Even after adjusting for age, black girls were still more sexually mature, heavier, and had higher levels of fat-free mass. Both serum insulin levels and serum concentrations of IGF-1 were significantly higher in black girls compared to white girls. Even after they controlled for differences in pubic hair development and fat mass, the differences in IGF-1 and IGFBP-1 remained significant. So black girls in this study had higher levels of IGF-1 and insulin than white girls.
This is significant because higher levels of IGF-1 at age 8 are associated with earlier menarche in girls (Thankamoney et al, 2012). Earlier menarche is related to higher levels of adiposity, as noted above, and the mechanism for earlier menarche is fat cells releasing insulin which has a permissive effect on menarche. Further, there is evidence that lower levels of circulating growth hormones are related to delayed menarche (Circo, 2014).
IGF-1 is a protein “coded for” by the IGF-1 gene (Hoppener et al, 1985). IGF-1 is molecularly similar to insulin (Laron, 2001). This makes sense. Body fat cannot be stored unless insulin is high; since they are molecularly similar, they both cause growth, albeit in different ways.
In conclusion, there is strong evidence that body fatness is related to earlier menarche, with the mechanism being fat cells releasing leptin which has a permissive effect on pubertal development. Further evidence indicates that circulating hormones like insulin and IGF-1 also have permissive effects on pubertal development. Black girls are more likely to be obese than white girls. Black girls are more likely to have higher levels of circulating hormones like IGF-1 and insulin than white girls. Thus, a cause—a major cause in my opinion—for earlier menarche in black girls compared to white girls is higher levels of body fat and higher levels of circulating hormones like IGF-1 and insulin which have a permissive effect on menarche.
Racial Differences in the Palmaris Longus
1050 words
There are many physical differences between racial/ethnic groups. Some of these differences are obvious to the naked eye, others much less so. One racial difference that exists is variation in having a tendon called the palmaris longus (PL). This muscle rests between the flexor carpi radialis and the flexor carpi urinalis. The radius and ulna are bones in the forearm. The radius supports the lateral (thumb) side of the hand whereas the ulna supports the medial (pinky) side; they enable wrist rotation. To see if you have the tendon, take your pinky and touch it to your thumb. If you have the tendon you should see it poking out of your wrist. Fourteen percent of the population lacks the PL, but there is considerable variation by race. These differences, of course, have should be taken into account when doing a tendon graph operation.
The action that the PL performs is flexing the wrist; the origin is the medial epicondyle of the humerus; it inserts in the palmar aponeurosis and flexor rentinaculum of the hand. The antagonist muscles are extensor carpi radialis brevis, extensor carpi radialis longus, and extensor carpi urinalis.
There are unilateral (affecting one side of the body) differences in the variation of this tendon along with bilateral (affecting both sides of the body) differences. There is differential absence of the tendon depending on which hand is dominant (Eric et al, 2011). The tendon also has been found to contribute to the strength of thumb abuction (Gangata, Ndou, and Louw, 2013). However, it has been shown that whether or not one has the PL or not does not contribute to grip/pinch strength (Sebastin et al, 2005).
Soltani et al (2012) followed patients at hand surgery clinics LA county +University of Southern California Medical Center and Keck Medical Center of the University of Southern California. Their objective was to observe the variation in the PL in regard to race, sex and ethnicity (indeed, they placed people into races based on the US Census designations; see Spencer, 2014, they excluded mixed-race people from the ethnicity part of the analysis). They wanted to see the extent that the PL was missing and whether or not it was bilateral or unilateral. They evaluated a group of 516 multi-ethnic individuals while age, race, ethnicity and sex was accounted for. They then administered the Schaeffer test:
![]()
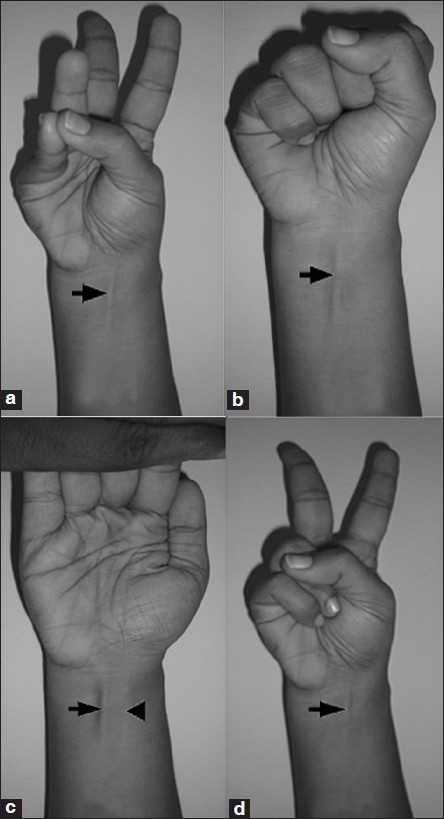
Obviously, this only works with lean individuals. So obese patients needed to get an ultrasound to ascertain whether or not they had the tendon.
In their sample of 516, 415 were Caucasian, 55 were African American, 35 were Asian and 11 were mixed race. The age range was 12-94, while an even number of men and women were tested (288, 288). Soltani et al (2012) write:
There were no differences in the absence of the PL based on laterality. The right side was absent in 11.8% and left 12.0% of the time (see Table 1). Further, there were no differences in the absence of the PL based on gender, value 0.369 (see Table 2). Ethnically, there was no difference in the absence of the PL between White (non-Hispanic) and White (Hispanic) patients, with prevalence of 14.9% and 13.1%, respectively. However, African American (4.5%) and Asian (2.9%) patients had significantly fewer absences of the PL than the Hispanic reference group.
They then write in the discussion that this has implications for plastic surgery—this anatomic variation between the races has implications for surgery:
This is information that should be taken into account preoperatively when planning surgical algorithms in treating tendon injuries or palsy. The PL is one such option as a tendon transfer for opponensplasty in restoring intrinsic function in cases of recurrent median nerve injury. If the PL is absent on the affected side, it is important to know preoperatively to plan using another donor muscle such as the extensor indicis proprius. In our study, the African American population had a statistically significantly lower rate of absent PL (4.5%), which is radically different than previously published reports from Nigeria, where the absence rates were much higher (31%). This could be due to the ethnic heterogeneity of the African American population of the United States compared to the Nigerian population. Nevertheless, the PL is present in high likelihood in this particular ethnic group which bodes well for using the PL in a surgical scenario. The PL is used quite frequently in cases of secondary tendon reconstruction, and it is useful for the surgeon to be aware of that issue preoperatively for surgical safety and efficiency in harvesting the tendon graft. The patient needs to be aware of the location of possible surgical incisions for tendon harvesting. Further, the surgeon should examine all possible tendon donors preoperatively, and one’s suspicion might be heightened by knowing the patient’s ethnicity. This is particularly important for the White population which in our study had the highest rates of absence, in both the Hispanic and non-Hispanic subsets. The surgeon must be aware in these patients that it is more likely that the PL might be absent. Thus, in Caucasian patients, it is particularly important to have a thorough examination of possible tendon donor sites.
Touching on the point of there being no difference between having or not having a PL and grip/pinch strength: we should not expect that, since whites dominate strength competitions (and the Chinese powerlifters are no slouches either, look at some videos of them). So, just because whites have a higher prevalence of not having the tendon does not mean that they do not have a strong hand grip. Indeed, senior blacks did have a stronger hand grip than whites (Araujo et al, 2010), but the PL is not the cause of the slightly higher hand grip strength (this was seen in a Turkish study; Setin et al, 2013). However, tennis players need a strong grip and the PL is more likely to be found in elite tennis players over the recreational athletes (Vercruyssen, Scafoglieri, and Cattrysse, 2016). But Nekkanti et al (2018) conclude that “There was no statistically significant correlation between agenesis of PL and the mean hand grip. Right-handed dominant individuals had a higher incidence of PL.”
In sum, this is tendon varies by race/ethnicity/sex/handedness like many other traits. understanding these differences will lead to a better understanding of plastic surgery in regard to the tendon. There is a racial difference, but it is of no functional relevance in regard to hand grip/power/strength sports.
Is Diet and Exercise Eugenic?
1550 words
Eugenics can be defined as “the science of improving a human population by controlled breeding to increase the occurrence of desirable heritable characteristics.” Though, in his book Genetic Ethics: An Introduction, Farrelly (2018: 30) cites Bertrand Russel’s definition of eugenics. Eugenics is “the attempt to improve the biological character of a breed by deliberate methods adopted to that end.” So, eugenics does not have to be ‘bad’, if it is morally justifiable and defensible, if they treat all people as free and equal (which most eugenic movements in the past have not done. See my article Eugenics and the State for a history of eugenics and the policies that arose from it.)
Dieting (defined as “what one eats”, not the other commonly-used definition “when one is in caloric restriction under TDEE”) and exercise change the expression of genes in the genome. For example, mature skeletal muscle can adapt to numerous stressors—indeed, if it could not, then we would not be able to choose to gain (or lose) muscle mass. Exercise induces the activity of certain genes (Vissing, Anderson, and Schjerling, 2005). One study on endurance athletes showed that there are pronounced effects of gene expression on exercised and non-exercised muscles (Catoire et al, 2012). One study showed that the ingestion of glucose during exercise decreases the gene expression of genes associated with fatty acid metabolism (Civitarese et al, 2005). Numerous epigenetic changes are also induced by exercise (Ntanasis-Stathopoulos, Tzanninis, and Koutsilieris, 2013). Miyamoto-Mikami et al (2018) showed that young men who participated in high-intensity intermittent exercise training showed that 79 genes had an elevated expression whereas 73 genes were significantly reduced.
Williams and Neufer (1996) show that long-term responses in regard to adaptations in regard to a specific exercise “require changes in gene expression, mediated by changes in the rate of transcription of specific genes and in the rate of synthesis of
specific proteins.” Further, diet and exercise can change the transcriptional properties of skeletal muscle, which induce further physiological changes (Hargreaves and Cameron-Smith, 2002). There is even preliminary evidence that diet and exercise affects the epigenome over several generations (Barres and Zierath, 2016). Since changes occur to the epigenome due to environmental stressors, and exercise is an environmental stressor, it follows that exercise, too, can change the epigenome.
People visit the gym to change their biology. Since people visit the gym to change their biology, are they involved in “self-eugenics”? I would say yes, going with the definition from Russell quoted above. That one wants to change their biology means that they are most likely not currently happy with the way their biological phenotype currently is. So they visit the gym, begin a diet (defined here as “caloric restriction”) in order to change their biological phenotype since they are not happy with it. Methods like diet and exercise seek to improve the biological character of a breed through deliberate methods, and so, fall under the umbrella of “eugenics.”
But there is a difference between this type of “eugenics” and methods commonly thought of when “eugenics” are discussed. “When eugenic measures exemplify moral and epistemic virtue rather than vice, they are morally obligatory rather than simply morally permissible” (Farrelly, 2018: 42). Prescribing (a sensible) diet and exercise to a populace can and will improve their health; further, educating people on the right and wrong things to eat (“right and wrong things to eat” in regard to their current goals) and how these things we eat affect our physiology is not morally objectionable nor is it coercive.
Think of the eugenic policies I discussed in my article on eugenics and the State. The policies discussed (such as forced sterilization, infanticide, forced contraceptive measures, and selective breeding) are immoral: the State is attempting to force its ideals on the populace, and so it can be argued that it is immoral since individual autonomy is taken away (or attempted to be taken away). On the other hand, prescribing diet and exercise is not eugenic in this manner: it’s just a prescription, what one should do if they would like to live their life to the highest quality. This includes staying away from highly processed and refined foods (carbs) and other, “non-natural” foodstuffs. This is only a suggestion based on the current state of nutritional knowledge; if one wants to live the best-possible life then they should diet and exercise.
On the other hand, we can take a State-measure and, using the definition in this article from Russell, can say that this measure is eugenic, but its similarities to what is being argued here is irrelevant, since I am arguing for education, not forcing people to do something (though I will state my views on this matter at the end, which I still honestly think about since it conflicts with some of my views.)
Back in the beginning of this decade, then-mayor of NYC Michael Bloomberg “proposed [a] regulation that would bar food service establishments from selling certain sugary drinks in containers larger than sixteen ounces” in an effort to “reduce the city’s obesity rate.” When this law was proposed, I was all for it. People cannot make decisions for themselves, because when they do, they make the “wrong” (in regard to, what I would assume to be what people try to achieve—a healthy lifestyle) decisions, and so, I thought that a policy like this was a good idea, because who the hell needs a 64 oz. Big Gulp soda (which could have up to 700 kcal in the cup) from 7-11? Why would someone need to down almost 2 liters of soda in one go? Note that, I would assume, the individual would not be caught dead drinking out of a 2-liter bottle of soda (though I have seen quite a lot of people do so, even early in the morning). But there is no problem using the cup since its size is kind of deceptive—deceptive in the manner that it does not look like the 2-liter bottle of (family-sized) soda.
I loved this proposal when it was announced. It would, I thought, attempt to address one aspect of our obesity problem (since nearly 40 percent of all of the added sugars we consume come from sugary drinks). Back then, I was more libertarian in my politics, but I thought that the policy was a good idea, even though it conflicted with my views on politics. I now do not believe we should take these types of measures—I believe that education is sufficient, along with getting rid of food deserts which hamper the ability of those in those deserts to get access to good, high-quality food (which affects certain races over others; National Research Council, 2009).
The view held by Bloomberg, and now current NYC mayor Bill Deblasio, is an example of a policy that would take away one’s choice to drink what they would like. Bloomberg’s rationale was that, if people wanted more of the drink, then they can go over and refill their cup so that they can see what and how much they are actually drinking. This, on its face, is sensible. If one wants to drink the same amount they would have drank in, say, a 64 oz. Big Gulp, they can keep refilling their cup in order to get the same amount of liquid they would have gotten in the bigger cup. But, what if someone wants a 64 oz. Big Gulp? What about a 128 oz. Ultra Big Gulp? A 256 oz. Super Ultra Big Gulp? Is there anywhere we should draw this line? Should we?
In any case, I have shown that exercise and diet is “eugenic” in the sense of Russel’s definition. But it is not “eugenic” in the sense of, in my opinion, what most people mean when they discuss “eugenics”: taking away one’s individual autonomy to do what they want, forcing them to do something. (Though, they are not being told they cannot drink sugary drinks, they are being told that there is a size limit on how big their cup is; they would be forced to drink a sugary drink in a small cup.)
Bloomberg’s proposed measure is quite obviously eugenic since it “attempt[s] to improve the biological character of a breed by deliberate methods adopted to that end.” Is the health of the populace more important than individual autonomy to be able to buy their 256 oz. Super Ultra Big Gulp? Or is one’s ability to freely drink their 256 oz. Ultra Big Gulp more important? If it can be shown that this policy would reduce the number of obese people in the City, should it be attempted?
These are important (moral) questions to answer. I am honestly undecided here; this issue is incredibly complex. Though, we do know one thing: exercise and diet is “eugenic” in Russel’s sense, and the measures exemplify moral and epistemic virtue so it is not like “State-forced” eugenics of old. Whether or not there is a negative connotation to “eugenics” depends on whether or not it pursues sound and morally justifiable aims. Therefore, though prescribing diet and exercise is eugenic since they both influence the epigenome and gene expression (along with it falling under Russel’s definition of eugenics), it is not eugenic in a negative sense, since there are sound and morally justifiable aims in prescribing diet and exercise to the population at large.
Race as a Genuine Kind
1600 words
Race realism can be defined as the belief that our racial categories pick out real kinds in nature. Picking out real kinds in nature is a necessary condition for race realism to be true. I’ve extensively covered Hardimon’s (2017) and Spencer’s (2014) arguments on the existence of race. They definitively prove that, all though there is a social/cultural dimension to race, biological racial realism is true, since our racial categories truly do pick out real kinds in nature.
In the philosophy of race, questions such as “What is race?” and “Is race biological?” are asked. Philosophers of race use many tools at their disposal to attempt to answer these and other questions regarding the metaphysics of race. Four main views on race exist: racial anti-realism, the belief that race does not exist; race realism, the belief that race is real; social constructivism, the belief that race is a social construct; and biological racial realism, the belief that race is biologically real (Spencer, 2011).
Note that social constructivists about race are race realists; they believe that race is real but that it is strictly a social reality. Racial anti-realists take the belief that race does not exist at all; so those who make the claim that social constructivists about race say that race is not real, they really mean to say that racial anti-realists claim that race is not real. People hear the phrase “social construct” and automatically assume that that individual is attempting to argue that race is not real. But social constructivists about race do believe that race is real—that is, they are race realists, but only in a social, not biological, manner.
Spencer (2011: 9) argues that genuine kinds are kinds that contribute to long-term scientific progress, meaning that genuine kinds are “a valid kind in a well-ordered SRP [scientific research program].” Spencer (2011: 9) states that valid kinds in an SRP are “useful for playing an epistemic role of scientific kinds in that SRP, as well as a kind that is adequately epistemically justified in its SRP” while also stating that “ a valid kind in an SRP must be useful for playing an epistemic role of scientific kinds in its SRP.” Kinds are valid in SRPs when they can help scientists generate observations for understanding the natural world. Genuine kinds can be epistemically useful without being inductively useful.
Genuine kinds can be epistemically real or just a human (social) construct. Genuine kinds are real enough to use in scientific research—because they tell us just enough about the world to be epistemically useful in a SRP. That a kind is genuine means that it is “real enough” to be used in scientific research.
Now let’s look at Spencer’s argument that biological racial realism should mean “race is a genuine kind in biology”:
(1) The meaning of ‘biological racial realism’ in the race debate should be a metaphysically minimal interpretation of important scientific kindhood that also does the most justice to what counts as an important scientific kind.
(2) A “metaphysically minimal” interpretation of important scientific kindhood is one that does not adopt unnecessary and contentious metaphysical assumptions.
(3) The interpretation of important scientific kindhood that does the most justice to what counts as an important scientific kind is the one that best captures epistemically important scientific kinds—or ‘EIS kinds’ for short.
(4) The candidates for important scientific kindhood in the race debate are naturalo kinds, naturali kinds, naturalu kinds, naturalp kinds, realp biological kinds, reali biological kinds, and genuine kinds.
(5) No kind of kind in the race debate is both metaphysically minimal and does a better job of capturing EIS kindhood than genuine kinds.
(6) Therefore, the meaning of ‘biological racial realism’ in the race debate should be ‘race is a genuine kind in biology’.
This argument provided by Spencer establishes the fact that biological racial realism in the race debate should be ‘race is a genuine kind in biology.’ Now that I have laid out that an entity being biologically real in the race debate is not whether or not it is objective but whether it is epistemically justified in a biological research program, I will now turn to other author’s views on biological racial realism.
The topic on “Race as a Biological Kind” on PhilPapers states that:
One option is to say that one’s ancestor is a member of race X in virtue of sharing similar phenotypic, or observable, properties specific to other members of one’ s reproductively isolated breeding population. A second option is to say that one’s ancestor is a member of race X in virtue of sharing similar genotypic, or genetic, properties specific to other members of one’s reproductively isolated breeding population.
While “… ancestral relations among reproductively isolated breeding populations and either genotypic or phenotypic properties is one way to develop a naturalistic account of race.” Note how this is almost, to the tee, Hardimon’s (2017) populationist race concept.
Andreason (2000) writes that “Most constructivists assume that biological realism and social constructivism are incompatible views about race; I argue that the two conceptions can be compatible.” Indeed, the claim that race is a social construct of a biological reality is a tempting view to take (and one I take to myself). That we socially construct groups does not mean that there is no biological reality to them, as Spencer (2014) shows with his Blumenbachian partitions. The claim that race is a social construct of a biological reality is completely at ends with the claim that race is purely social/political (as Dorothy Roberts argues) and the claim that race is purely biological. Though, we cannot separate ‘social/political’ and ‘biological’ terminology from our ontology. We must use both in conjunction in order to tease out what ‘race’ truly means. Stating that race is a social construct and only a social construct betrays the biology behind race. Stating that race is only biological betrays the social aspects of race. (Though, Hardimon (2017) has one race concept—the minimalist race concept—in which there is nothing ‘social’ about his proposed concept of race.)
Hardimon has four concepts of race: (1) the racialist concept of race—the claim that significant intellectual and moral differences exist between races (which he dispatched in his book); (2) the minimalist concept of race—the claim that groups that exhibit patterns of visible physical features which correspond to geographic ancestry satisfy the conditions of minimalist race; (3) the populationist race concept of race (the scientization of the minimalist race concept)—the claim that “race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population” (Hardimon, 2017: 99); and (4) the socialrace concept of race—the claim that race is a social, not biological, reality, and that society constructs what ‘race’ is.
Our racial categories pick out real kinds in nature, therefore race realism is true. This is established by both Hardimon (2017) and Spencer (2014), who discuss Rosenberg et al’s (2002) paper on the existence of population clusters deemed continental-level minimalist races by Hardimon (2017) and Blumenbachian partitions by Spencer (2014). Both arguments provided by Hardimon and Spencer are sound—they answer numerous objections that critics bring up to argue against Rosenberg et al’s (2002) study and other’s interpretation of what they say.
This article discusses Rosenberg et al (2002), and has this to say about the Kalash (the sixth cluster in that Rosenberg et al (2002) found):
The sixth population which is composed entirely of members of the Kalash, an isolated population in central Pakistan, is omitted by Wade on the grounds it “makes no genetic or geographic sense”. But the Kalash have a significant degree of genetic isolation from others, comparable in magnitude to that of other groups such as Native Americans. Moreover, they all live in the same place.
This is nonsense. That Structure labels a population as genetically distinct does not entail that that population is a continental-level minimalist race. That Structure picks out the Kalash as a genetically distinct group does not undercut Hardimon’s (2017) arguments on the existence of continental-level minimalist races. So, both K=6 and K=5 show that continental-level minimalist races are genetically structured (Hardimon, 2017: 88). (Further responses to critiques of Rosenberg et al can be found here and in Hardimon, 2017, chapter 5 and Spencer, 2014.)
Lastly, I’ll take some time to respond to commenter Oliver D. Smith’s objections to cranial measures and geographic ancestry.
Smith cited Sierp and Henneberg’s (2015) paper Can ancestry be consistently determined from the skeleton? in which the authors show that “no one individual was identified as belonging to only one ‘racial class’” (Sierp and Henneberg, 2015: 23). I don’t find this to be a problem. If we take Hardimon’s minimalist and populationist race concepts, the three conditions that need to be satisfied to delineate races are differences in phenotype which correspond to geographic ancestry, geographic location, and geographic ancestry. The critique from Sierp and Henneberg (2015) only counts against one parameter—if that—which are skulls. When delineating races, we don’t only look at skulls (one visible physical feature), we look at the whole suite of traits that make a “race” “racial.” Therefore, Sierp and Henneberg’s (2015) critique has no bearing on Hardimon’s (2017) or Spencer’s (2014) arguments.
Is race a genuine kind in biology? Do our racial categories pick out real kinds in nature? The answer to both questions is “yes.” Race is a genuine kind in biology since it captures EIS kindhood; our racial categories do pick out real kinds in nature, as shown by Spencer (2014) and his Blumenbachian partitions. Hardimon’s and Spencer’s arguments are definitive: biological racial realism is true.
(i) If biological racial realism is true, then our racial categories would have to pick out real kinds in nature.
(ii) Our racial categories pick out real kinds in nature.
(iii) Therefore, biological racial realism is true.
