
Home » Twin Studies
Category Archives: Twin Studies
Why Heritability Estimates are Flawed: A Conceptual Account
1200 words
Introduction
Heritability estimates have been used as a cornerstone and psychology and genetic research. They are designed to quantify the proportion of phenotypic variance in a population that can be attributed to genetic differences among individuals. We’ve known for a while now that heritability isn’t a measure of genetic strength (Moore and Shenk, 2016), but it’s a population-specific estimate of variance. Here I will provide two a priori arguments (one methodological on twins and the EEA and one theoretical based on Noble’s biological relativity argument). The twin critique shows that the twin researcher’s main assumption (equal environments) does not hold while the biological relativity critique shows that h2 is conceptually invalid. This is why there is a missing heritability problem—it never existed in the first place, and the assumptions twin researchers have are false.
The classical twin method
The CTM compares MZ and DZ twins to attempt to quantify the relative contributions of genes and environment in relation to the origin of trait differences between individuals. Perhaps the biggest assumption of the twin researcher is the equal environments assumption (EEA). The EEA assumes that MZ and DZ twins experience equivalent shared environments.
The EEA seems plausible enough: twins reared together should experience compatible environments, regardless of zygosity. But since DZ twins are more genetically similar than DZ twins so they’ll be more phenotypically similar as well. MZ twins are dressed alike, mistaken for one another, or placed in similar social roles compared to DZ twins which leads to more similar environments. So the shared environmental variance for MZ twins exceeds that for DZ twins, violating the EEA.
Clearly this violation throws a wrench into the logic of the CTM. The formula assumes that the greater similarity in MZ twins stems solely from their genetic identity. But if MZ twins experience more similar environments due to their phenotypic similarity (Fosse, Joseph and Richardson, 2015; Joseph et al, 2015), the difference in correlations between MZs and DZs captures genetic variance and excess environmental similarity. Thus, heritability is overestimated (see eg Bingley, Cappellari, and Tatsiramos, 2023) increasing the effect of genes while masking the effects of the environment—in effect, environment is made to look like genes. Thus, h2 fails to isolate genetic influence as intended. (Note that Grayson 1989 explains this as well, but it seems that it’s just ignored.) Here’s the argument:
(1) The classical twin method assumes that its heritability (h2) estimate (Falconer’s formula) isolates the proportion of phenotypic variance due solely to genetic variance.
(2) For the h2 estimate to isolate genetic variance, the shared environmental variance must be equal for MZ and DZ twins.
(3) MZ twins are more genetically similar than DZ twins.
(4) Genetic similarity between individuals leads to greater similarity in their expressed phenotypic traits, and this phenotypic similarity results in greater similarity in their environmental experiences.
(5) Because MZ twins have greater genetic similarity than DZ twins, and genetic similarity leads to phenotypic similarity, which in turn results in environmental similarity, the shared environmental variance is greater for MZ twins compared to DZ twins.
(6) If the shared environmental variance for MZ twins is greater than that for DZ twins, then the EEA is false because it requires that shared environmental variance be equal for both twin types.
(7) If the EEA is false, then we cannot logically infer genetic conclusions from h2, and thus h2 reflects shared environmental variance (c2), rather than genetic variance.
(8) Any method that relies on an assumption that’s logically inconsistent with the principles governing it’s variables – like the relationship between genetic similarity, environmental similarity and phenotypic similarity – cannot accurately isolate its intended causal component and is therefore conceptually untenable.
(9) Thus, the classical twin method is conceptually and logically untenable since it depends on the EEA which, when false, renders h2 a measure of environmental—not genetic—variance.
The biological relativity critique against h2
This argument is theoretical as opposed to methodological, and it relies on Noble’s (2012) biological relativity argument, where there is no privileged level of causation in biological systems. Genes, cells, tissues, organs, organisms, and the environment form an interdependent network where each level influences and is influenced by the other levels. Phenotypes arise from the interaction between all of these levels, not just due to the independent action of any one of the resources.
Heritability rests on a reductionist assumption—that phenotypic variance can be neatly partitioned into genetic and environmental components with genetic effects isolated as a distinct and quantifiable entity. This framework, clearly, privileges the genetic level treating it as separate from the broader biological and ecological context. But Noble’s argument directly contradicts this view. Genes don’t operate in a vacuum and do nothing on their own.
So by attempting to isolate genetic variance, heritability imposed an artificial simplicity on a complex reality (Rose, 2006). Noble’s principle suggests that separation isn’t just an approximation but a fundamental conceptual flaw. Phenotypic variation emerges from the integrated functioning of all biological levels, which then makes it impossible to assign causation to genes alone.
Thus, h2 is conceptually flawed, since it seeks to measure a genetic contribution that cannot be meaningfully disentangled from the holistic system in which it operates. Obviously the conceptual foundation of h2 contradicts the principle of biological relativity. Since h2 attempts to assign a specific portion of trait variance to genes alone, h2 implicitly privileges the genetic level, suggesting that it can be disentangled from the broader biological system. Noble’s argument denies that this is possible while emphasizing holism and rejecting reductionism. Thus, a priori, h2 estimates are fundamentally flawed because they rest on a reductionist framework which assumes a separability of causes which is incompatible with the holistic, relativistic nature of biological causation. Here’s the argument:
(1) Biological relativity holds that there is no privileged level of causation in biological systems: all levels (genes, cells, tissues, organs, organisms, environments) are interdependent in producing phenotypes.
(2) h2 assumes that genetic variance can be isolated and quantified as a distinct contributor to phenotypic variance.
(C) Since biological relativity rejects the isolation of genetic effects, h2 is conceptually invalid as a measure of genetic influence.
Conclusion
Both of these arguments show the same thing—h2 is a deeply flawed concept. The EEA critique exposes a methodological weakness: since MZ twins experience more similar environments than DZ twins, the excess environmental similarity experienced by MZs masquerades as genetic influence, leaving h2 incapable of isolating genetic variance.
But Noble’s biological relativity argument strikes at a deeper conceptual flaw in this practice, since it challenges the theoretical aspects of h2 itself. Since it highlights the interdependence of biological systems, it dismantles the reductionist notion that genetic effects can be separated from other levels of causation. The gene-centric assumption is at ends with the reality of phenotypes being emergent properties of multi-level interactions, which then renders the concept of h2 conceptually incoherent. Therefore, h2 isn’t only empirically questionable but it is theoretically untenable. The conceptual model is just not sound due to how genes really work (Burt and Simon, 2015)
Thus, again, hereditarianism fails conceptually. Even their main “tool” fails for a modicum of reasons not least the main theoretical killshot for heritability estimates—the principle of biological relativity. The reductionist hereditarian paradigm is conceptually and logically untenable, it’s time to throw it away, it’s time to throw it to the dustbin of history.
Evaluating Heritability in an Imagined Metabolic Ward Study of Twins
2000 words
Introduction
For the better part of 100 years, twin studies have been used to prove that there is a genetic influence to all traits. The claim is that “all human traits are heritable”, which is the “first law of behavioral genetics” as stated by Turkheimer. Using twin, family, and adoption studies, it is claimed that some traits are “more genetic than others”, especially IQ. But a meta-analysis 10 years ago found that heritability was 49 percent for all traits (Polderman et al, 2014; see here for critique). However due to the falsity of the EEA—which holds that environments are similar for both MZ and DZ twins—the so-called heritability is shared environment (h2=c2). (See Joseph, 2014 for arguments against twin interpretations and Joseph 2022, 2023 for arguments against Thomas Bouchard’s twin studies. See Bouchard 2023 for response to Joseph 2022, and Joseph 2023 is Joseph’s response.)
But imagine we could bypass such devestating issues for twin studies that invalidates them for their stated aims. What would we find? Here, I will create a thought experiment in which 100 MZs and 100 DZs were placed in a metabolic ward immediately after birth. In this scenario, every single aspect of the environment is meticulously controlled—as is the case in animal breeding studies—which would then eliminate shared environmental influences and therefore would give us a “real look” into the (obviously context-dependent) heritability of traits.
However unethical this study is, though, even if it were possible there are a slew of conceptual and empirical issues that would still invalidate the estimates. The issues include the limitations of assuming additive genetic effects, the presence of GxE, the non-representative nature of twins, and the missing heritability problem. The argument I will mount here will show that even with perfectly controllable environments, we still wouldn’t be able to “estimate heritability” and furthermore due to the interaction problem that this study still wouldn’t overcome, that would further make any such results meaningless.
The unethical twin metabolic ward study
A metabolic ward is a controlled environment which is usually found in a hospital, research facility or clinic in which the participant’s can be closely monitored and their metabolic processes studied under tightly regulated conditions. Researchers can control diet, diet quality, sleep, physical activity, and environmental influences to investigate nutrient absorption, energy expenditure, hormone regulation and metabolic disorders. Controlled settings like this allow researchers to get precise measurements, ensure that a participant is following a diet correctly as to study their metabolic processes on that diet, and understand the physiological mechanisms better. (See Hodges and Bean, 1960.)
Now imagine all of that but imagine that 100 MZ twins and 100 DZ twins were—after birth—immediately whisked away to a metabolic ward and from that moment on, they were in a tightly regulated environment as to attempt to minimize or completely eliminate shared environmental influences. What would happen?
We could learn about the genetic influence of genetics on metabolic processes, like the influence of genes on nutrient absorption, energy expenditure, hormone regulation and metabolic disorders. So by comparing MZ and DZ twins on the similarity and differences in these traits, we could then ascertain the genetic influence that underlies the variability between them. This would also allow researchers to completely control sleep, eating, physical activity levels and other environmental exposures which would then allow researchers to tease a part the relative influenced of genes and environment. This would also allow us to see how GxE affects a trait. We could learn about how genetic predispositions interact with dietary factors or how lifestyle choices modulate metabolic health outcomes and disease risk. It could also give us insights into individual variability so we can give individualized metabolic approaches.
However, even if IRB can be passed, there are of course potential psychological and social implications for those studied. The issue is, we have minds and we therefore react differently to things independently of our genetic makeup. So although we could potentially learn some interesting things about human metabolic processes, when it comes to psychological ones, due to the unrepresentative environment, these will be less valid than metabolic processes.
We technically do “measure” heritability of traits now, but they’re heavily confounded, and even though—intuitively—it may seem like such an experiment would be valid and show “true heritability” (whatever that means), we know that h2 is context-dependent and not useful for individuals. Such a study would be about perfectly controlling every aspect of the environment as best we could to decrease whatever environmental effect would bias the h2 upwards. But I think a slew of conceptual issues would still even then invalidate such interpretations.
Cattle and humans are quite obviously different in their propensities, and even then, if we could pass IRB ethics and lock 100 MZs and 100 DZs in a metabolic ward and perfectly control every aspect of the environment, the issue there would be our experiences in society and our interactions with others are extremely meaningful to how we develop as humans. So if that kind of experiment were possible, then the h2s would be way lower than twin, family and adoption studies give us (we see this in animal breeding studies—way lower h2 in those traits than for IQ, see Schonemann, 1997).
There are also further conceptual issues like the falsity of the addivitiy assumption, GxE, GxG (Zuk et al, 2011), psychological distress, and the fact that we become human and gain our minds through our interactions with people in broader society would also further confound such interpretations. Furthermore, since the EEA is false, h2 equals c2, meaning the heritability is shared environment. That’s the best interpretation we have of twin studies. And the missing heritability issue that plagues GWAS and other molecular approaches to trait heritability further throws a wrench in this, since it’s completely possible that GxE (and other interactions) contribute to trait variance, and this—along with epigenetic and non-genetic factors—aren’t accounted for by h2 estimates (since they assume additive independent genetic effects). So even if such an experiment were possible and the conceptual issues I raised weren’t an issue, that alone would invalidate the study.
One big issue is the fact that twins aren’t representative of the population at large. For example we know that twins have a higher chance of having low birth weights, are more likely to be born premature than non-twins (Isakkson, Ruchkin, and Ljungstrom, 2023). Low birth weight is also predictive of health outcomes in adulthood (Hassan et al, 2021). Twins are also more likely to experience complications during pregnancy and childbirth like preterm birth, intrauterine birth restriction and childbirth trauma. The intrauterine birth restrictions could also influence their metabolic development and responses within the ward, which This then would influence their outcomes in adulthood compared to non-twins. We also know that there are a slew of environmental effects that increase the chance for DZ twinning between groups (MZ twinning is relatively the same between them). Moreover, due to the complications that twins face before, during, and after childbirth, this could then affect their metabolic health and responses within the metabolic ward. Therefore, while this thought experiment proposes ways in which researchers can use a metabolic ward to study the relative influence of genes and environment on traits (assuming that there are no other conceptual issues with such an endeavor), researchers would still need to take into account the ways in which twins differ from the general population which would then influence the results that would be observed in the metabolic ward study.
Joseph (2009) proposed an interesting thought experiment:
Finally, we could conduct a thought experiment on political behavior and social attitude correlations among reared-apart MZ twin pairs who, while genetically identical, grow up in truly uncorrelated environments in different eras. Suppose one male MZA twin is placed at birth in an aristocratic Japanese family in 1802. The other male MZA twin is placed at birth in a poor peasant family living in the highlands of El Salvador in 1965. Unlike previous TRA studies, inwhich the investigators calculate correlations among partially reared-apart twins sharing many cultural influences, in our thought experiment we eliminate cultural influences such as family (which most MZAs share to some extent), mutual association and influence (which most MZAs also share to some extent), nation, region, ethnic, religion, economic class, and birth cohort (of course, they would still be the same sex).75 I conclude this thought experiment by posing the following question: Would we expect a study of genetically identical pairs of this type to find sizable correlations for political behavior and social attitudes?
The correct conclusion to draw here is that although the twins share genetic makeup (to whatever degree they do), but they are raised in different environments and different time periods, the divergence in their experiences shows that heritability can’t fully capture the role of environment in capturing in shaping not only these traits but all traits studied by twin researchers.
Conclusion
In the end, if such a study were logistically possible and ethical, it wouldn’t show h2 anywhere near twin, family and adoption studies. We’ve basically reached an upper bound on molecular h2 estimates. And of course twins aren’t representative of the general population at large (since they have lower birth weights/injuries and a higher mortality rate). And even then, twins in these studies aren’t even representative of all twins. This is, yet again, a conceptual failure of hereditarianism—the assumption that twin studies will tell us anything about “the genetic architecture” of traits merely due to genetic similarity and being so-called “reared apart.”
Even if such a study were possible, there would be a lack of generalizability due to the fact that the observations occurred during rightly controlled conditions. Such a study is fraught with conceptual, empirical and ethical issues. Biases in sample selection, the health differences between twins and non-twins, prenatal and peri-natal factors, psychological and social dynamics and the lack of generalizability highlight why twins aren’t a representative of the population at large. Moreover, the complexity of metabolic traits and the assumption of genetic independence further complicate interpretations in the ward setting—especially since DNA interacts with all other developmental resources (Noble, 2012).
But even in a possible world where such limitations can be addressed and made meaningless, there are still inherent limitations of heritability estimates as a measure of the genetic determination of traits (in the popular culture, see Moore et al, 2016 for a critique if the heritability concept). Heritability estimates are context-dependent and subject to numerous confounding factors.
But disregarding this thought experiment and looking to the real world, we know that there is no way to estimate interactions in population studies. The so-called solution is to assume that there are NO gene-environment and gene-gene interactions, and this therefore biases such estimates upwards (Ho, 2013). This shows that the assumption of additive effects and no GxG or GxE interactions would still pose a problem—an insurmountable one. Thus, even estimates from this study would still be subject to upward bias due to the nature of the assumptions of heritability. This shows that even in a perfectly controlled environment that measuring h2 is impossible due to the disregarded biological facts inherent in the heritability formula. Joseph and Richardson (2024) also show that Herrnstein and Murray (1994) didn’t give any valid evidence that genes influence IQ scores within groups.
So the conclusion here is very simple—since heritability studies can’t tell us anything about the relative contributions of genes and environments due to the inherent nature of the heritability formula which disregards interactions, and due to the whole slew of other conceptual and empirical issues raised here, even in a perfectly controlled environment we still wouldn’t be able to get “true h2” estimates of any kind of trait in the metabolic ward. So the goal of behavioral genetics is an impossible one.
On the So-Called “Laws of Behavioral Genetics”
2400 words
In the year 2000, psychologist Erik Turkheimer proposed three “laws of behavioral genetics” (LoBG hereafter):
● First Law. All human behavioral traits are heritable.
● Second Law. The effect of being raised in the same family is smaller than the effect of genes.
● Third Law. A substantial portion of the variation in complex human behavioral traits is not accounted for by the effects of genes or families. (Turkheimer, 2000: 160)
In March of 2021, I asked Turkheimer how he defined “law.” He responded: “With tongue in cheek. In fact, it’s a null hypothesis: an expected result when nothing in particular is going on.“
In 2015, Chabris et al (2015) proposed a 4th “law”, that a typical behavioral trait is associated with many variants which each explain a small amount of behavioral variability. They state that the “4th law” explains the failure of candidate gene studies and also the need for higher sample sizes in GWA studies. (It seems they are not aware that larger sample sizes increase the probability of spurious correlations—which is all GWA studies are; Claude and Longo, 2016; Richardson, 2017; Richardson and Jones, 2019) Nice ad hoc hypothesis to save their thinking.
One huge proponent of the LoBG is JayMan, who has been on a crusade for years pushing this nonsense. He added a “5th law” proposed by Emil Kirkegaard, which states that “All phenotypic relationships are to some degree genetically mediated or confounded.”
But what is a “law” and are these “laws of behavioral genetics” laws in the actual sense? First I will describe what a “law” is and if there even are biological laws. Then I will address each “law” in turn. I will then conclude that the LoBG aren’t real “laws”, they are derived from faulty thinking about the relationship between genes, traits, environment and the system and how the “laws” were derived rest on false assumptions.
What is a law? Are there biological laws?
Laws are “true generalizations that are “purely quantitative” … They have counterfactual force” (Sober, 1993: 458). Philosopher of mind Donald Davidson argued that laws are strict and exceptionless (Davidson, 1970; David-Hillel, 2003). That is, there must be no exceptions for that law. Sober (1993) discusses Rosenberg’s and Beatty’s arguments against laws of biology—where Rosenberg states that the only law in biology is “natural selection.” (See Fodor, 2008 and Fodor and Piattelli-Palmarini, 2009, 2010 for the argument against that claim and for arguments against the existence of laws of selection that can distinguish between causes and correlates of causes.) It has even been remarked that there are “so few” laws in biology (Dhar and Giuliani, 2010; also see Ruse, 1970).
Biology isn’t reducible to chemistry or physics (Marshal, 2021), since there are certain things about biology that neither chemistry or physics have. If there are laws of biology, then they will be found at the level of the organism or its ecology (Rull, 2022). In fact, it seems that although three laws of biology have been proposed (Trevors and Sailer Jr., 2008), they appear to be mere regularities, including McShea and Brandon’s (2010) first law of biology; all “laws of biology” seem to be mere laws of physics (Wayne, 2020). The “special sciences”, it seems, “are not fit for laws” (Kim, 2010). There seem to be, though, no uncontroversial laws or regularities in biology (Hamilton, 2007).
Now that I have described what laws are and have argued that there probably aren’t any biological laws, what does that mean for the LoBG? I will take each “law” in turn.
“Laws” of behavioral genetics
(1) All human behavioral traits are heritable.
JayMan gives derivations for the “laws”, and (1) and (2) have their bases in twin studies. We know that the equal environments assumption is false (Charney, 2012; Joseph, 2014; Joseph et al, 2015), and so if the EEA is false then we must reject genetic claims from twin study proponents. Nevertheless, the claim that these “laws” have any meaning gets pushed around a lot.
When it comes to the first law, the claim is that “ALL human behavioral traits are heritable”—note the emphasis on “ALL.” So this means that if we find only ONE behavioral trait that isn’t heritable, then the first law is false.
Reimann, Schilke, and cook (2017) used a sample of MZ and DZ twins and asked questions related to trust and distrust. They, of course, claim that “MZ and DZ twins share comparable environments in their upbringing“—which is false since MZ twins have more comparable environments. Nevertheless, they conclude that while trust has a heritability or 30%, “ACE analyses revealed that the estimated heritability [for] distrust is 0%.” This,therefore, means, that the “1st law” is false.
This “first law”, the basis of which is twin, family, and adoption studies, is why we have poured countless dollars into this research, and of course people have their careers (in what is clear pseudoscience) to worry about, so they won’t stop these clearly futile attempts in their search for “genes for” behavior.
(2) The effect of being raised in the same family is smaller than genes.
This claim is clearly nonsense, and one reason why is that the first “law” is false. In any case, there is one huge effect on, children’s outcomes due to birth order and how, then, parental attitudes–particularly mothers—affect child outcomes (Lehmann, Nuevo-Chiquero, and Vidal-Fernandez, 2018).
Why would birth order have an effect? Quite simply, the first-born child will get more care and attention than children who are born after, and so variations in parental behavior due to birth order can explain differences in education and life outcomes. They conclude that “broad shifts in parental behavior appear to set later-born children on a lower path for cognitive development and academic achievement, with lasting impact on adult outcomes.” Thus, Murray’s (2002) claim that birth order doesn’t matter and JayMan’s claim that “that the family/rearing environment has no effect on eventual outcomes” is clearly false. Thus, along with this and the falsity of the “1st law”, the “2nd law” is false, too.
(3) A substantial portion of the variation in complex human behavioral traits is not accounted for by the effects of genes or families.
This “law” covers the rest of the variance not covered in the first two “laws.” It was coined due to the fact that the first two “laws” had variance left that wasn’t “explained” by them. So this is basically unique experience. This is what behavioral genetics call “non-shared environment.” Of course, unique experiences (that is, subjective experiences) would definitely “shape who we are”, and part of our unique experiences are cultural. We know that cultural differences can have an impact on psychological traits (Prinz, 2014: 67). So the overall culture would explain why these differences aren’t “accounted for” in the first two “laws.”
Yet, we didn’t need the LoBG for us to know that individual differences are difference-makers for differences in behavior and psychology. So this means that what we choose to do can affect our propensities and then, of course, our behavior. Non-shared environmental effects are specific to the individual, and can include differing life events. That is, they are random. Non-shared environment, then, is parts of the environment that aren’t shared. Going back to Lehmann, Nuevo-Chiquero, and Vidal-Fernandez (2018) above, although children to grow up in the same family under the same household, they are different ages and so they also experience different life events. They also experience the same things differently, due to the subjectivity of experience.
In any case, the dichotomy between shared and non-shared environment is a dichotomy that upholds the behavioral geneticists main tool—the heritability estimate—from which these “laws” derive (from studies of twins, adoptees, and families). So, due to how the law was formulated (since there were still portions “unaccounted for” by the first two “laws”), it doesn’t really matter and since it rests on the first two false “laws”, therefore the third “law” is also false.
(4) Human behavioral traits are associated with many genes of small effect which contribute to a small amount of behavioral variability.
This “law” was formulated by Chabris et al (2015) due to the failure of molecular genetic studies which hoped to find genes with large effects to explain behavior. This “law” also “explains why the results of “candidate-gene” studies, which focus on a handful of genetic variants, usually fail to replicate in independent samples.” What this means to me is simple—it’s an ad-hoc account, meaning it was formulated to save the gene-searching by behavioral geneticists since the candidate gene era was a clear failure, as Jay Joseph noted in his discussion of the” 4th law.”
So here is the time line:
(1) Twin studies show above-0 heritabilities for behavioral traits.
(2) Since twin studies show high heritabilities for behavioral traits, then there must be genes that will be found upon analyzing the genome using more sophisticated methods.
(3) Once we started to peer into the genome after the completion of the human genome project, we then came to find candidate genes associated with behavior. Candidate gene studies “look at the genetic variation associated with disease within a limited number of pre-specified genes“, they refer to genes “believed to be” associated with a trait in question. Kwon and Goat (2000) wrote that “The candidate gene approach is useful for quickly determining the association of a genetic variant with a disorder and for identifying genes of modest effect.” But Sullivan (2020) noted that “Historical candidate gene studies didn’t work, and can’t work.” Charney (2022) noted that the candidate gene era was a “failure” and is now a “cautionary tale.”
Quite clearly, they were wrong then, and the failure of the candidate gene era led to the ad-hoc “4th law.” This has then followed us to the GWAS and PGS era, where it is claimed that we aren’t finding all of the heritability that twin studies say we should find with GWAS, since the traits under review are due to many genes of small effect. It’s literally just a shell game—when one claim is shown to be false, just make a reason why what you thought would be found wasn’t found, and then you can continue to search for genes “for” behavior. But genetic interactions create a “phantom heritability” (Zuk et al, 2011), while behavioral geneticists assume that the interactions are additive. They simply outright ignore interactions, although they pay it lip service.
So why, then, should we believe behavioral geneticists today in 2023 that we need larger and larger samples to find these mythical genes “for” behavior using GWAS? We shouldn’t. They will abandon GWAS and PGS in a few years when the new kid on the block shows up that they can they champion and claim that the mythical genes “for” behavior will finally be found.
(5) All phenotypic relationships are to some degree genetically mediated or confounded.
This claim is something that comes up a lot—the claim of genetic confounding (and mediation). A confound is a third variable that influences both the dependent and independent variable. The concept of genetic confounding was introduced during the era where it was debated whether or not smoking caused lung cancer (Pingault et al, 2021). (Do note that Ronald Fisher (1957), who was a part of this debate, claimed that smoking and lung cancer were both “influenced by a common cause, in this case individual genotype.“
However, in order for the genetic confounding claim to work, they need to articulate a mechanism that explains the so-called genetic confounding. They need to articulate a genetic mechanism which causally explains X and Y, explains X independent of Y and explains Y independent of X. So for the cop-out genetic confounding claim to hold any water: G confounds X and Y iff there is a genetic mechanism which causally explains X and Y, causally explains X independent of Y and Y independent of X.
Conclusion
The “laws of behavioral genetics” uphold the false dichotomy of genes and environment, nature and nurture. Though, developmental systems theorists have rightly argued that it is a false dichotomy (Homans, 1979; Moore, 2002; Oyama, 2002; Moczek, 2012) and that it is just not biologically plausible (Lewkowicz, 2012). In fact, the h2 statistics assumes that G and E are independent, non-interacting factors, so if the claim is false then—for one of many reasons—we shouldn’t accept their conclusions. The fact that G and E interact means that, of course, we should reject h2 estimates, and along with it, the entire field of behavioral genetics.
Since the EEA is false, h2 equals c2. Furthermore, h2 equals 0. So Polderman’s (2015) meta analysis doesn’t show that for all traits in the analysis that h2 equals 49%. (See Jay Joseph’s critique.) Turkheimer (2000: 160) claimed that the nature-nurture debate is over, since everything is heritable. However, the debate is over because developmental systems approach has upended the false dichotomy of nature vs nurture, since all developmental resources interact and are therefore irreducible to development.
However, for the field to continue to exist, they need to promulgate the false dichotomy, since their heritability estimates depend on it. They also need to hold onto the claim that twin, family and adoption studies can show the “genetic influence” on traits to justify the continued search for genes “for” behavior. Zuk and Spencer (2020) called the nature-nurture “debate” “a zombie idea, one that, no matter how many times we think we have disposed of it, springs back to life.” This is just like Oyama (2000) who compared arguing against gene determinism like battling the undead (Griffiths, 2006).
Jay Joseph proposed a 5th “law” in 2015 where he stated:
Behavior genetic Laws 1-4 should be ignored because they are based on many false assumptions, concepts, and models, on negative gene finding attempts, and on decades of unsubstantiated gene discovery claims.
The “laws” should quite obviously be ignored. Since the whole field of behavioral genetics is based on them, why not abandon the search for “genes for behavior”? At the end of the day, it seems like there are no “laws” of behavioral genetics, since laws are strict and exceptionless. So why do they keep up with their claims that their “laws” tell us anything about human behavior? Clearly, it’s due to the ideology of those who hold that the all-important gene causes traits and behavior, so they will do whatever it takes to “find” them. But in 2023, we know that this claim is straight up false.
Twin Studies, Adoption Studies, and Fallacious Reasoning
2350 words
Twin and adoption studies have been used for decades on the basis that genetic and environmental causes of traits and their variation in the population could be easily partitioned by two ways: one way is to adopt twins into separate environments, the other to study reared-together or reared-apart twins. Both methods rest on a large number of (invalid) assumptions. These assumptions are highly flawed and there is no evidential basis to believe these assumptions, since the assumptions have been violated which invalidates said assumptions.
Plomin et al write (2013) write: For nearly a century, twin and adoption studies have yielded substantial estimates of heritability for cognitive abilities.
But the validity of the “substantial estimates of heritability for cognitive abilities” is strongly questioned due to unverified (and false) assumptions that these researchers make.
Adoption studies
The problem with adoption studies are numerous, not least: restricted range of adoptive families; selective placement; late separation; parent-child attachment disturbance; problems with the tests (on personality, ‘IQ’); the non-representativeness of adoptees compared to non-adoptees; and the reliability of the characteristic in question.
In selective placement, the authorities attempt to place children in homes close to their biological parents. They gage how “intelligent” they believe they are (on the basis of parental SES and the child’s parent’s perceived ‘intelligence’), thusly this is a pretty huge confound for adoption studies.
According to adoption researcher Harry Munsinger, a “possible source of bias in adoption studies is selective placement of adopted children in adopting homes that are similar to their biological parents’ social and educational backgrounds.” He recognized that “‘fitting the home to the child’ has been the standard practice in most adoption agencies, and this selective placement can confound genetic endowment with environmental influence to invalidate the basic logic of an adoptive study (Munsinger, 1975, p. 627). Clearly, agency policies of “fitting the home to the child” are a far cry from random placement of adoptees into a wide range of adoptive homes. (Joseph, 2015: 30-1)
Richardson and Norgate (2005) argue that simple additive effects for both genetic and environmental effects are false; that IQ is not a quantitative trait; while other interactive effects could explain the IQ correlation.
1) Assignment is nonrandom. 2) They look for adoptive homes that reflect the social class of the biological mother. 3) This range restriction reduces the correlation estimates between adopted children and adopted parents. 4) Adoptive mothers come from a narrow social class. 5) Their average age at testing will be closet to their biological parents than adopted parents. 6) They experience the womb of their mothers. 7) Stress in the womb can alter gene expression. 8) Adoptive parents are given information about the birth family which may bias their treatment. 9) Biological mothers and adopted children show reduced self-esteem and are more vulnerable to changing environments which means they basically share environment. 10) Conscious or unconscious aspects of family treatment may make adopted children different from other adopted family members. 11) Adopted children also look more like their biological parents than their adoptive parents which means they’ll be treated accordingly.
Twin studies
Personally, my favorite thing to discuss. Twin studies rest on the erroneous assumption that DZ and MZ environments are equal; that they get treated equally the same. This is false, MZ twins get treated more similarly than DZ twins, which twin researchers have conceded decades ago. But in order to save their field, they attempt to use circular argumentation, known as Argument A. Argument A states that MZTs (monozygotic twins reared together) are more genetically similar than DZTs (dizygotic twins reared together) and thusly this causes greater behavioral similarity. But this is based on circular reasoning: the researchers already implicitly assumed that genes played a role in their premise and, not surprisingly, in their conclusion genes are the cause for the similarities of the MZTs. So Argument A is used, twin researchers circularly assume that MZTs greater behavioral similarity is due to genetic similarity, while their argument that genetic factors explain the greater behavioral similarity of MZTs is a premise and conclusion of their argument. “X is true because Y is true; Y is true because X is true.” (Also see Joseph et al, 2015.)
We have seen that circular reasoning is “empty reasoning in which the conclusion rests on an assumption whose validity is dependent on the conclusion” (Reber, 1985, p. 123). … A circular argument consists of “using as evidence a fact which is authenticated by the very conclusion it supports,” which “gives us two unknowns so busy chasing each other’s tails that neither has time to attach itself to reality” (Pirie, 2006, p. 27) (Joseph, 2016: 164).
Even if Argument A is accepted, the causes of behavioral similarities between MZ/DZ twins could still come down to environment. Think of any type of condition that is environmentally caused but is due to people liking what causes the condition. There are no “genes for” that condition, but their liking the thing that caused the condition caused an environmental difference.
Argument B also exists. Those that use Argument B also concede that MZs experience more similar environments, but then argue that in order to show that twin studies, and the EEA, are false, critics must show that MZT and DZT environments differ in the aspects that are relevant to the behavior in question (IQ, schizophrenia, etc).
An example of an Argument B environmental factor relevant to a characteristic or disorder is the relationship between exposure to trauma and post-traumatic stress disorder (PTSD). Because trauma exposure is (by definition) an environmental factor known to contribute to the development of PTSD, a finding that MZT pairs are more similarly exposed to trauma than DZT pairs means that MZT pairs experience more similar “trait-relevant” environments than DZTs. Many twin researchers using Argument B would conclude that the EEA is violated in this case. (Joseph, 2016: 165)
So twin researchers need to rule out and identify “trait-relevant factors” which contribute to the cause of said trait, along with experiencing more similar environments, invalidates genetic interpretations made using Argument B. But Argument A renders Argument B irrelevant because even if critics can show that MZTs experience more similar “trait-relevant environments”, they could still argue that the twin method is valid by stating that (in Argument A fashion) MZTs create and elicit more similar trait-relevant environments.
One more problem with Argument A is that it shows that twins behave accordingly to “inherited environment-creating blueprint” (Joseph, 2016: 164) but at the same time shows that parents and other adults are easily able to change their behaviors to match that of the behaviors that the twins show, which in effect, allows them to “create” or “elicit” their own environments. But the adults’ “environment-creating behavior and personality” should be way more unchangeable than the twins’ since along with the presumed genetic similarity, adults have “experienced decades of behavior-molding peer, family, religious, and other socialization influences” (Joseph, 2016: 165).
Whether or not circular arguments are “useful” or not has been debated in the philosophical literature for some time (Hahn, Oaksford, and Corner, 2005). However, assuming, in your premise, that your conclusion is valid is circular and therefore While circular arguments are deductively valid, “it falls short of the ultimate goal of argumentative discourse: Whatever evaluation is attached to the premise is transmitted to the conclusion, where it remains the same; no increase in degree of belief takes place” (Hahn, 2011: 173).
However, Hahn (2011: 180) concludes that “the existence of benign circularities makes clear that merely labeling something as circular is not enough to dismiss it; an argument for why the thing in question is bad still needs to be made.” This can be simply shown: The premise that twin researchers use (that genes cause similar environments to be constructed) is in their conclusion. They state in their premise that MZT behavioral similarity is due to greater MZT genetic similarity in comparison to DZTs (100 vs. 50 percent). Then, in the conclusion, they re-state that the behavioral similarities of MZTs is due to their genetic similarity compared to DZTs (100 vs. 50 percent). Thus, a convincing argument for conclusion C (that genetic similarity explains MZT behavioral similarity) cannot rest on the assumption that conclusion C is correct. Thus, Argument A is fallacious due to its circularity.
What causes MZT behavioral similarities is their more similar environment: they get treated the same by peers and parents, and have higher rates of identity confusion and had a closer emotional bond compared to DZTs. The twin method is based on the (erroneous) assumption that MZT and DZT pairs experience roughly equal environments, which twin researchers conceded was false decades ago.
Richardson and Norgate (2005: 347) conclude (emphasis mine):
We have shown, first, that the EEA may not hold, and that well-demonstrated treatment effects can, therefore, explain part of the classic MZ–DZ differences. Using published correlations, we have also shown how sociocognitive interactions, in which DZ twins strive for a relative ‘apartness’, could further depress DZ correlations, thereby possibly explaining another part of the differences. We conclude that further conclusions about genetic or environmental sources of variance from MZ–DZ twin data should include thorough attempts to validate the EEA with the hope that these interactions and their implications will be more thoroughly understood.
Of course, even if twin studies were valid and the EEA was true/ the auxiliary arguments used were true, this would still not mean that heritability estimates would be of any use to humans, since we cannot control environments as we do in animal breeding studies (Schonemann, 1997; Moore and Shenk, 2016). I have chronicled how 1) the EEA is false and how flawed twin studies are; 2) how flawed heritability estimates are; 3) how heritability does not (and cannot) show causation; and 4) the genetic reductionist model that behavioral geneticists rely on is flawed (Lerner and Overton, 2017).
So we can (1) accept the EEA, that the greater behavioral resemblance indicates the importance of genetic factors underlying most human behavioral differences and behavioral disorders or we can (2) reject the EEA and state that the greater behavioral resemblance is due to nongenetic (environmental) factors, which means that all genetic interpretations of MZT/DZT studies must be rejected. Thus, using (2), we can infer that all twin studies measure is similarity of the environment of DZTs, and it is, in fact, not measuring genetic factors. Accepting explanation 2 does not mean that “twin studies overestimate heritability, or that researchers should assess the EEA on a study-by-study basis, but instead indicates that the twin method is no more able than a family study to disentangle the potential influences of genes and environment” (Joseph, 2016: 181).
What it does mean, however, is that we can, logically, discard all past, future, and present MZT and DZT comparisons and these genetic interpretations must be outright rejected, due to the falsity of the EEA and the fallaciousness of the auxiliary arguments made in order to save the EEA and the twin method overall.
There are further problems with twin studies and heritability estimates. Epigenetic supersimilarity (ESS) also confounds the relationship. Due to the existence of ESS “human MZ twins clearly cannot be viewed as the epigenetic equivalent of isogenic inbred mice, which originate from separate zygotes. To the extent that epigenetic variation at ESS loci influences human phenotype, as our data indicate, the existence of ESS establishes a link between early embryonic epigenetic development and adult disease and may call into question heritability estimates based on twin studies” (Van Baak et al, 2018). In other words, ESS is an unrecognized phenomenon that contributes to the phenotypic similarity of MZs, which calls into question the usefulness of heritability studies using twins. The uterine environment has been noted to be a confound by numerous authors (Devlin, Daniels, and Roeder, 1997; Charney, 2012; Ho, 2013; Moore and Shenk, 2016).
Conclusion
Adoption studies fall prey to numerous pitfalls, most importantly, that children are adopted into similar homes compared to their birth parents, which restricts the range of environments for adoptees. Adoption placement is also non-random, the children are placed into homes that are similar to their biological parents. Due to these confounds (and a whole slew of other invalidating problems), adoption studies cannot be said to show genetic causation, nor can they separate genetic from environmental factors.
Twin studies suffer from the biggest flaw of all: the falsity of the EEA. Since the EEA is false—which has been recognized by both critics and supporters of the assumption—the supporters of the assumption have attempted to redefine the EEA in two ways: (1) that MZTs experience more similar environments due to genetic similarity (Argument A) and (2) that it is not whether MZTs experience more similar environments, but whether or not they share more similar trait-relevant environments. Thus, unless these twin researchers are able to identify trait-relevant factors that contribute to the trait in question, we must conclude that (along with the admission from twin researchers that the EEA is false; that MZTs experience more similar environments than DZTs) genetic interpretations made using Argument B are thusly invalidated. Fallacious reasoning (“X causes Y; Y causes X) does not help any twin argument. Because their conclusion is already implicitly assumed in their premise.
The existence of ESS (epigenetic supersimilarity) further shows how invalid the twin method truly is, because the confounding starts in the womb. Attempts can be made (however bad) to control for shared environment by adopting different twins into different homes, but they still shared a uterine environment which means they shared an environment, which means it is a confound and it cannot be controlled for (Charney, 2012).
Adoption and twin studies are highly flawed. Like family studies, twin studies are no more able to disentangle genetic from environmental effects than a family study, and thus twin studies cannot separate genes from environment. Last, and surely not least, it is fallacious to assume that genes can be separated so neatly into “heritability estimates” as I have noted in the past. Heritability estimates cannot show genetic causation, nor can it show how malleable a trait is. They’re just (due to how we measure) flawed measures that we cannot fully control so we must make a number of (false) assumptions that then invalidate the whole paradigm. The EEA is false, all auxiliary arguments made to save the EEA are fallacious; adoption studies are hugely confounded; twin studies are confounded due to numerous reasons, most importantly the uterine environment (Van Baak et al, 2018).
Behavior Genetics and the Fallacy of Nature vs Nurture
3250 words
People appeal to moderate to high heritability estimates as evidence that a trait is controlled by genes. They then assume that because something has a high heritability then that it must show something about causation. The fact of the matter is, they do not. Heritability estimates assume a false dichotomy of nature vs nurture; it assumes that we can neatly partition genetic from environmental effects. It assumes that the higher a trait’s heritability the more genes control said trait. These are all false. One of the main ways that heritability is estimated is by the CTM (classic twin method). This method, though, has a ton of assumptions poured into it—most importantly, the assumption that DZ and MZ fraternal twins experience roughly equal environments—the equal environments assumption (EEA). Heritability studies are useless for humans; twin studies bias estimates upwards with a whole host of assumptions.
I will show that i) heritability estimates are highly flawed (due to erroneous assumptions); ii) nature vs nurture cannot be separated (like behavior geneticists claim) and so their main tool (the heritability estimate) should be discontinued; iii) genetic reductionism is not a tenable model due to what we now know about how genes work. All three of these reasons are enough to discontinue heritability estimates. If the nature vs nurture debate rests on a fallacy, and this fallacy is used as a vehicle for heritability estimates, then they should be discontinued for humans and only be used for breeding animals where they can control the environment fully (Schonemann, 1997; Moore and Shenk, 2016).
Heritability, twin studies, and equal environments
Back in 2014-2015, there was a debate in the criminological literature that had implications for heritability studies as a whole. Burt and Simons (2014) stated that it was time to get rid of heritability studies. Barnes et al (2015) responded that this was “a de facto form of censorship” (pg 2). Joseph et al (2015) respond to these accusations, writing, “It was good science and not “censorship” when earlier scientists called for ending studies based on craniometry, phrenology, and physiognomy, and any contemporary criminologist calling for the use of astrological charts to predict whether certain people will commit violent crimes would be justifiably ridiculed.” The main thing here, in my opinion, is that heritability estimates are based on an oversimplified (and wrong) model of the gene. Partitioning variance assumes that you can partition how much a trait is influenced by “nature” or “nurture” which is a false dichotomy (Moore, 2002; Schneider, 2007; Moore and Shenk, 2016).
More importantly, no “genes have been found” (I know that’s everyone’s favorite thing to hear) for traits that supposedly have high heritabilities. On page 179 of his book (nook version), Misbehaving Science, Controversy and the Development of Behavior Genetics Panofsky (2014) writes:
Molecular genetics has been a major dissapointment, if not an outright failure, in behavior genetics. Scientists have made many bold claims about genes for behavioral traits or mental disorders only to later retract them or to have them not be replicated by other scientists. Further, the findings that have been confirmed, or not yet falsified, have been few, far between, and small in magnitude.
There seems to be a huge disconnect between heritability estimates gleaned from twin studies and what the actual molecular genetic evidence says. This is because the EEA—that fraternal MZ twins experience roughly similar environments compared to fraternal DZ twins—is false. Fraternal MZ twins end up experiencing more similar environments when compared with fraternal DZ twins. Though most researchers attempt to save face by stating that MZ twins “seek out” and “elicit” their own environments which then makes them more similar compared to DZ twins. However, this is circular logic. The conclusion (that twins experience more similar environments) is in the premise, and therefore it is an invalid argument due to the logical fallacy. (It should also be noted that identical twins’ genes are not identical.)
Heritability studies assume an outdated model of the gene. The flaw regarding heritability estimates is simple: they imply a false dichotomy of nature vs nurture, while also assuming that genes and environment are independent, while the contribution to complex behaviors can be precisely quantified (Charney, 2013). This is one of the most critical parts of the heritability debate. Prenatal environments of DZ twins “can be significantly more stressful than that of DZ twins, and hence a cause of greater stress-related phenotypic concordance, the equal environment assumption will not hold in relation to behavioral phenotypes potentially associated with prenatal stress” (Charney, 2012: 20). This also is cause for concern regarding studies of twins reared apart. While twins are reared apart to eliminate shared environmental confounds, it cannot eliminate perhaps the most important confound of all—the prenatal environment (Moore and Shenk, 2016).
One of the most-cited studies regarding twins reared apart is Bouchard (1990). Though there are a whole slew of problems with this study.
1) You have the huge confound of similar environments before birth.
2) Full details for the MISTRA have never been published, so we don’t know how ‘separated’ the twins were. Though Bouchard et al do say that they were separated between 0 to 48.7 months (table 1) so some pairs spent at least 4 years together. Some of the twins even had reunions and spent a lot of time together.
3) They’re not representative and twins who do sign up for this research are self-selecting. Ken Richardson says in his book (2017, pg 55): “Twins generally tend to be self-selecting in any twin study. They may have responded to advertisements placed by investigators or have been prompted to do so by friends or family, on the grounds that they are alike. Remember, at least some of them knew each other prior to the study. Jay Joseph has suggested that the twins who elected to participate in all twin studies are likely to be more similar to one another than twins who chose not to participate. This makes it difficult to claim that the results would apply to the general population.”
4) And the results aren’t fully reported. Richardson also states that (2017, pg 55) “… of two IQ tests administered in the MISTRA, results have been published for one but not the other. No explanation was given for that omission. Could it be they produced different results?” He even states that attempts to get the data, by researchers like Jay Joseph, have been denied. Why would you refuse to publish, or give to another researcher, your data when asked?
We don’t know the relevant environments, the children’s average age at testing is closer to the biological mother than adopted mother; the biological mother and child will have reduced self-esteem and be more vulnerable to difficult situations, and in this sense they share environments; and conscious or unconscious bias make adopted children different from other family members. Adoption agencies also attempt to put children into similar homes as the biological mother too.
Charney (2012: 25) brings up an important point: “For phenotypes of any degree of complexity, DNA does not contain a determinate genetic program (analogous to the digital code of a computer) from which we can predict phenotype. If DNA were the sole carrier of information relevant to phenotype formation, and contained a genetic program sufficiently determinate that solely by reading it we could predict phenotype, then humans (and all other organisms) would be largely lacking in phenotypic plasticity.” Moore and Shenk (2016) also state that “we inherit developmental resources, not traits.”
1 For twin studies to be valid DZ twins and MZ fraternal twins would have to experience roughly equal environments. 2 Fraternal MZ twins experience much more similar environments than DZ twins. 3 Therefore the EEA is false and no genetic interpretations can be drawn from the data.
Heritability estimates cannot detangle genes and environment, and therefore they should be discontinued or reinterpreted (Joseph et al, 2015). Burt and Simons (2014: 110) also conclude: “Rejecting heritability studies and the false nature–nurture dichotomy and gene-centric model on which they are grounded is a necessary step forward that will pave the way for a reconceptualization of the link between the biological and the social in shaping criminal propensities in ways that are consistent with postgenomic knowledge“. I disagree with Barnes et al (2015) when they say that ending heritability estimates are “a defacto form of censorship“, because if nature vs nurture is a false dichotomy and the gene-centric model that heritability estimates rely on is wrong, then we need to either discontinue or reinterpret the estimates, not saying that ‘this is how much nature contributes to X and this is how much nurture contributes to Y’. (See also Richardson and Norgate, 2005 for more arguments regarding the EEA.)
Sapolsky (2017: 219) writes:
Oh, that’s right, humans. Of all species, heritability scores in humans plummet the most when shifting from a controlled experimental setting to considering the species’ full range of habitats. Just consider how much the heritability score for wearing earrings, with its gender split, has declined since 1958.
Heritability flaws
High heritability estimates have been used as evidence for causation—that genes control a large part of the trait in question. This reasoning, however, is highly flawed. People confuse “heritable” with “inheritable” (Moore and Shenk, 2016). Heritability does not inform us what causes a trait, how much environment contributes to a trait, nor does it tell us the relative influence of genes on a trait. Moore and Shenk (2016) agree with Joseph et al (2015) and Burt and Simons (2014) that heritability studies need to end, but Moore and Shenk’s reasoning slightly differs: they say we should end estimates because people confuse “heritable” with “inheritable”. Likewise, Guo (2000: 299) concurs, writing “it can be argued that the term ‘heritability’, which carries a strong conviction or connotation of something ‘heritable’ in everyday sense, is no longer suitable for use in human genetics and its use should be discontinued.”
Some may say that if a trait turns out to be mildly heritable then we can say that genes have some effect, but we know that genes affect all traits so it seems kind of redundant to have a useless measure that assumes a false dichotomy and relies on an outdated, additive model of the gene.
Rose (2006), too, agrees that heritability estimates imply a false dichotomy of nature vs nurture onto biological systems:
Biological systems are complex, non-linear, and non-additive. Heritability estimates are attempts to impose a simplistic and reified dichotomy (nature/nurture) on non-dichotomous processes.
Likewise, Lewontin (2006) argues we should be analyzing and studying causes, not variance.
There are numerous hereditarian scientific fallacies which include: 1) trait heritability does not predict what would occur when environments/genes change; 2) they’re inaccurate since they don’t account for gene-environment covariation or interaction while also ignoring nonadditive effects on behavior and cognitive ability; 3) molecular genetics does not show evidence that we can partition environment from genetic factors; 4) it wouldn’t tell us which traits are ‘genetic’ or not; and 5) proposed evolutionary models of human divergence are not supported by these studies (since heritability in the present doesn’t speak to what traits were like thousands of years ago) (Bailey, 1997).
Bailey (1997) brings up important arguments against the use of heritability, and even discusses fallacious writing from Rushton on the matter:
Rushton (1995), for example, thinks that if observed differences among the
racial groups that he defines are higher for traits that have high heritability within the groups, the hypothesis of genetically caused differences among the groups is supported.
Bailey (1997) then goes on to discuss three lakes: Otter lake, Welcome lake, and Bark lake. Otter lake has very high primary production, while Bark lake has very little and Welcome lake is somewhere in between (you can see that ‘Otter’, ‘Bark’ and ‘Welcome’ lakes are analogies for ‘Orientals’, ‘Blacks’, and ‘Whites’ as said by Rushton). But there is variation within the lakes, there are high production pockets of water in Bark lake while there are low production pockets of water in Otter lake. All three lakes are visited and measurements are taken. Bailey (1997) states that his conclusion would be that they differ in how much light each receives. Bailey (1997: 131) writes:
If I substitute three groups of people for my lakes, IQ for primary production, and genes for light levels, the fallacy of the slippery scale, as applied to human behaviour genetics, becomes clear. Even if we are sure that there is a difference among groups of people in IQ, and we are sure that IQ has high heritability within
each of the groups (i.e. variation in IQ is largely caused by genetic variation), we can make no inference about the cause of differences in IQ among the groups. The differences might be caused by genetic differences or they might not, but the heritability studies within the groups can’t help us make that judgment.

(Genes don’t cause IQ scores—or behavior—but that’s for another day.)
Heritability estimates for, say, IQ, are higher than any other trait in the animal kingdom. Heritability estimates for animal traits are low—lower than the stratospheric heritability of IQ. For example, heritability estimates of the bodyweight of farm animals is about 30 percent, which is the same for egg and milk production. Body fat in pigs and wool on sheep has a heritability of about 50 percent. But these estimates pale in comparison to the heritability estimates of IQ: estimates have been as high as 80 percent (but Schonemann, 1997 states it’s 60 percent but it’s as high as 80-90 percent today); this heritability estimate for IQ “surpasses almost anything found in the animal kingdom” (Schonemann, 1997: 104).
This high heritability estimate for IQ, of course, comes to us from the highly flawed twin studies discussed above. The reason why farmers and botanists use heritability estimates is that they can perfectly control the environment, and therefore get accurate—or close enough to it—estimates that will help them in their breeding efforts. Conversely, for humans, environments cannot be perfectly controlled and it is, of course, unethical to rear twins, MZ and DZ, in a controlled environment. Proponents of the twin method may say “It doesn’t matter if it’s flawed, it still shows there is a genetic component to trait X!”. But as discussed by Moore and Shenk (2016), that’s irrelevant because genetic factors influence all of our characteristics.
Heritability and causation
In the final section, I will shortly discuss how people fallaciously assume that high heritability estimates imply that a trait is strongly influenced by genetic factors.
In his essay in the book Postgenomics: Perspectives on Biology After the Genome, sociologist Aaron Panofsky (2016: 167; nook version) writes:
Heritability estimates do not help identify particular genes or ascertain their functions in development or physiology, and thus, by this way of thinking, they yield no causal information.
This is important to note: to those who truly believe that heritability estimates tell us anything about causation, how could they, logically, give us causal information if genes that lead to trait variation are not identified (Richardson, 2012)?
Panofsky (2014: 102-103) writes:
Experimental evidence from plants and animals suggest that shapes of the curves cannot inferred in advance and rarely follow the smooth, nonintersecting pattern like in figure 3.2. Thus true causal interpretations of heritability are hopeless and must be abandoned. Behavior geneticists did not claim direct experimental evidence, but they thought these various indirect lines of evidence provided a reasonable set of assumptions that would enable them to interpret heritability scores causally—provided they offer apporopriate, reasonable qualifications.
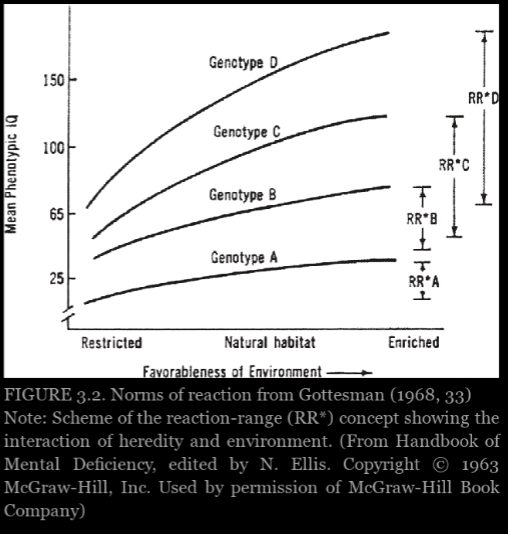
Graph from Panofsky (2014: 103)
Heritability estimates imply nothing about causation. It is about associations with variance, not identity and causes (Richardson, 2017: 69). A heritability of 0 does not mean that genes do not play a role in the development of form and function and phenotypic variation, it just means that, for whatever reason, there is little correlation between the two.
Scheneider (2007) writes (emphasis mine):
Heritability estimates apply only to groups, and are inherently inapplicable to individuals in any sense. And they do not imply causation. As Moore notes, all of these important limitations have been frequently ignored or minimized.
Reductionism
Heritability estimates imply nothing about causation. Behavior geneticists and others assume that heritability estimates will lead to ‘finding the genes’ that ’cause’ or are ‘associated with’ behavior. Their models are also, of course, extremely reductionist. It is then important to note that genes do not determine behavior. To quote Lerner and Overton (2017: 114):
Data presented in a 2016 special section of the journal Child Development indicate
that “some behaviors may be affected by only slight changes in DNA methylation,
while others may require a larger percent change in methylation; of course, the
effects are also likely bidirectional, with behavior impacting changes in methylation” [Lester et al., 2016, p. 31]. This point is key . It underscores the absurdity of genetic reductionist models: Genes do not determine behavior.
Methylation impacts behavior; behavior impacts methylation. It is the relations between methylation and behavior, not the genes acting as the “command center”, the “executive” of human behavior and development, that constitute the basic role of biology across the developmental course. This is the fatal flaw of reductionist models. Lastly, Lerner and Overton (2017: 145) write (emphasis mine):
That is, with the recent advances in understanding the role of epigenetics and recent research findings supporting this role, it should no longer be possible for any scientist to undertake the procedure of splitting of nature and nurture and, through reductionist procedures, come to conclusions that the one or the other plays a more important role in behavior and development.
[Richardson (2017: 129) also writes: “Note that this environmental source of [epigenetic] variation will appear in the behavioral geneticists twin-study as genetic variation: quite probably another way in which heritability estimates are distorted.”]
Reductionism in biology is fatally flawed. Reductionism, of course, has greatly increased our understanding of biology. However, it is time to move past the false dichotomy of nature vs nurture, and with that, move past heritability estimates since they prop up the fallacy of nature vs nurture. There is no way to separate the two since they are intertwined, but behavior geneticists would like you to believe that by studying twins raised apart will tell you anything about how ‘genetic’ or ‘environmental’ variation in a trait is in a population. Since heritability estimates are gleaned from the highly flawed studies of twins reared apart, a whole host of assumptions is poured in and these estimates are highly inflated, showing that genes influence a trait more than they supposedly do.
Twin studies, and along with it, heritability estimates, are useless for figuring out, and describing, trait variation in humans. The developmental system is more complex than the genetic reductionists (behavior geneticists) would like one to believe. The reductionist model has been heavily attacked in recent years (Regenmortal, 2004; Noble, 2008, 2012, 2015, 2016; Joyner, 2011, b; Joyner and Pederson, 2011).
Nature vs nurture has also shown to be a false dichotomy because the system develops in whichever environment it finds itself in (Oyama, 1985, 1999; 2000; Moore, 2002; Schneider, 2007)
Conclusion
Since the genetic reductionist model is wrong, along with heritability estimates (because of the nature/nurture fallacy), both should be discontinued. One of the main vehicles of these two models—twin studies—should also be discontinued. These fatal flaws of the behavior geneticists’ paradigm should be enough to discontinue these techniques in the study of human development and behavior. Heritability estimates give no causal information and they also use an outdated model of the gene; twin studies assume too many things for it to be a viable model in the discovering how traits manifest (most importantly, twin studies keep the nature/nurture fallacy alive and should be discontinued on that note only, in my opinion); and genetic reductionist models have been shown to be fatally flawed in recent years. We now have a better understanding of what a gene is today (Portin and Wilkins, 2017), and due to this, we should discontinue whatever implies the fallacy of nature vs nurture because it is irrelevant and a false dichotomy. That, alone, should be enough to discontinue twin studies and heritability estimates.
