
3250 words
People appeal to moderate to high heritability estimates as evidence that a trait is controlled by genes. They then assume that because something has a high heritability then that it must show something about causation. The fact of the matter is, they do not. Heritability estimates assume a false dichotomy of nature vs nurture; it assumes that we can neatly partition genetic from environmental effects. It assumes that the higher a trait’s heritability the more genes control said trait. These are all false. One of the main ways that heritability is estimated is by the CTM (classic twin method). This method, though, has a ton of assumptions poured into it—most importantly, the assumption that DZ and MZ fraternal twins experience roughly equal environments—the equal environments assumption (EEA). Heritability studies are useless for humans; twin studies bias estimates upwards with a whole host of assumptions.
I will show that i) heritability estimates are highly flawed (due to erroneous assumptions); ii) nature vs nurture cannot be separated (like behavior geneticists claim) and so their main tool (the heritability estimate) should be discontinued; iii) genetic reductionism is not a tenable model due to what we now know about how genes work. All three of these reasons are enough to discontinue heritability estimates. If the nature vs nurture debate rests on a fallacy, and this fallacy is used as a vehicle for heritability estimates, then they should be discontinued for humans and only be used for breeding animals where they can control the environment fully (Schonemann, 1997; Moore and Shenk, 2016).
Heritability, twin studies, and equal environments
Back in 2014-2015, there was a debate in the criminological literature that had implications for heritability studies as a whole. Burt and Simons (2014) stated that it was time to get rid of heritability studies. Barnes et al (2015) responded that this was “a de facto form of censorship” (pg 2). Joseph et al (2015) respond to these accusations, writing, “It was good science and not “censorship” when earlier scientists called for ending studies based on craniometry, phrenology, and physiognomy, and any contemporary criminologist calling for the use of astrological charts to predict whether certain people will commit violent crimes would be justifiably ridiculed.” The main thing here, in my opinion, is that heritability estimates are based on an oversimplified (and wrong) model of the gene. Partitioning variance assumes that you can partition how much a trait is influenced by “nature” or “nurture” which is a false dichotomy (Moore, 2002; Schneider, 2007; Moore and Shenk, 2016).
More importantly, no “genes have been found” (I know that’s everyone’s favorite thing to hear) for traits that supposedly have high heritabilities. On page 179 of his book (nook version), Misbehaving Science, Controversy and the Development of Behavior Genetics Panofsky (2014) writes:
Molecular genetics has been a major dissapointment, if not an outright failure, in behavior genetics. Scientists have made many bold claims about genes for behavioral traits or mental disorders only to later retract them or to have them not be replicated by other scientists. Further, the findings that have been confirmed, or not yet falsified, have been few, far between, and small in magnitude.
There seems to be a huge disconnect between heritability estimates gleaned from twin studies and what the actual molecular genetic evidence says. This is because the EEA—that fraternal MZ twins experience roughly similar environments compared to fraternal DZ twins—is false. Fraternal MZ twins end up experiencing more similar environments when compared with fraternal DZ twins. Though most researchers attempt to save face by stating that MZ twins “seek out” and “elicit” their own environments which then makes them more similar compared to DZ twins. However, this is circular logic. The conclusion (that twins experience more similar environments) is in the premise, and therefore it is an invalid argument due to the logical fallacy. (It should also be noted that identical twins’ genes are not identical.)
Heritability studies assume an outdated model of the gene. The flaw regarding heritability estimates is simple: they imply a false dichotomy of nature vs nurture, while also assuming that genes and environment are independent, while the contribution to complex behaviors can be precisely quantified (Charney, 2013). This is one of the most critical parts of the heritability debate. Prenatal environments of DZ twins “can be significantly more stressful than that of DZ twins, and hence a cause of greater stress-related phenotypic concordance, the equal environment assumption will not hold in relation to behavioral phenotypes potentially associated with prenatal stress” (Charney, 2012: 20). This also is cause for concern regarding studies of twins reared apart. While twins are reared apart to eliminate shared environmental confounds, it cannot eliminate perhaps the most important confound of all—the prenatal environment (Moore and Shenk, 2016).
One of the most-cited studies regarding twins reared apart is Bouchard (1990). Though there are a whole slew of problems with this study.
1) You have the huge confound of similar environments before birth.
2) Full details for the MISTRA have never been published, so we don’t know how ‘separated’ the twins were. Though Bouchard et al do say that they were separated between 0 to 48.7 months (table 1) so some pairs spent at least 4 years together. Some of the twins even had reunions and spent a lot of time together.
3) They’re not representative and twins who do sign up for this research are self-selecting. Ken Richardson says in his book (2017, pg 55): “Twins generally tend to be self-selecting in any twin study. They may have responded to advertisements placed by investigators or have been prompted to do so by friends or family, on the grounds that they are alike. Remember, at least some of them knew each other prior to the study. Jay Joseph has suggested that the twins who elected to participate in all twin studies are likely to be more similar to one another than twins who chose not to participate. This makes it difficult to claim that the results would apply to the general population.”
4) And the results aren’t fully reported. Richardson also states that (2017, pg 55) “… of two IQ tests administered in the MISTRA, results have been published for one but not the other. No explanation was given for that omission. Could it be they produced different results?” He even states that attempts to get the data, by researchers like Jay Joseph, have been denied. Why would you refuse to publish, or give to another researcher, your data when asked?
We don’t know the relevant environments, the children’s average age at testing is closer to the biological mother than adopted mother; the biological mother and child will have reduced self-esteem and be more vulnerable to difficult situations, and in this sense they share environments; and conscious or unconscious bias make adopted children different from other family members. Adoption agencies also attempt to put children into similar homes as the biological mother too.
Charney (2012: 25) brings up an important point: “For phenotypes of any degree of complexity, DNA does not contain a determinate genetic program (analogous to the digital code of a computer) from which we can predict phenotype. If DNA were the sole carrier of information relevant to phenotype formation, and contained a genetic program sufficiently determinate that solely by reading it we could predict phenotype, then humans (and all other organisms) would be largely lacking in phenotypic plasticity.” Moore and Shenk (2016) also state that “we inherit developmental resources, not traits.”
1 For twin studies to be valid DZ twins and MZ fraternal twins would have to experience roughly equal environments. 2 Fraternal MZ twins experience much more similar environments than DZ twins. 3 Therefore the EEA is false and no genetic interpretations can be drawn from the data.
Heritability estimates cannot detangle genes and environment, and therefore they should be discontinued or reinterpreted (Joseph et al, 2015). Burt and Simons (2014: 110) also conclude: “Rejecting heritability studies and the false nature–nurture dichotomy and gene-centric model on which they are grounded is a necessary step forward that will pave the way for a reconceptualization of the link between the biological and the social in shaping criminal propensities in ways that are consistent with postgenomic knowledge“. I disagree with Barnes et al (2015) when they say that ending heritability estimates are “a defacto form of censorship“, because if nature vs nurture is a false dichotomy and the gene-centric model that heritability estimates rely on is wrong, then we need to either discontinue or reinterpret the estimates, not saying that ‘this is how much nature contributes to X and this is how much nurture contributes to Y’. (See also Richardson and Norgate, 2005 for more arguments regarding the EEA.)
Sapolsky (2017: 219) writes:
Oh, that’s right, humans. Of all species, heritability scores in humans plummet the most when shifting from a controlled experimental setting to considering the species’ full range of habitats. Just consider how much the heritability score for wearing earrings, with its gender split, has declined since 1958.
Heritability flaws
High heritability estimates have been used as evidence for causation—that genes control a large part of the trait in question. This reasoning, however, is highly flawed. People confuse “heritable” with “inheritable” (Moore and Shenk, 2016). Heritability does not inform us what causes a trait, how much environment contributes to a trait, nor does it tell us the relative influence of genes on a trait. Moore and Shenk (2016) agree with Joseph et al (2015) and Burt and Simons (2014) that heritability studies need to end, but Moore and Shenk’s reasoning slightly differs: they say we should end estimates because people confuse “heritable” with “inheritable”. Likewise, Guo (2000: 299) concurs, writing “it can be argued that the term ‘heritability’, which carries a strong conviction or connotation of something ‘heritable’ in everyday sense, is no longer suitable for use in human genetics and its use should be discontinued.”
Some may say that if a trait turns out to be mildly heritable then we can say that genes have some effect, but we know that genes affect all traits so it seems kind of redundant to have a useless measure that assumes a false dichotomy and relies on an outdated, additive model of the gene.
Rose (2006), too, agrees that heritability estimates imply a false dichotomy of nature vs nurture onto biological systems:
Biological systems are complex, non-linear, and non-additive. Heritability estimates are attempts to impose a simplistic and reified dichotomy (nature/nurture) on non-dichotomous processes.
Likewise, Lewontin (2006) argues we should be analyzing and studying causes, not variance.
There are numerous hereditarian scientific fallacies which include: 1) trait heritability does not predict what would occur when environments/genes change; 2) they’re inaccurate since they don’t account for gene-environment covariation or interaction while also ignoring nonadditive effects on behavior and cognitive ability; 3) molecular genetics does not show evidence that we can partition environment from genetic factors; 4) it wouldn’t tell us which traits are ‘genetic’ or not; and 5) proposed evolutionary models of human divergence are not supported by these studies (since heritability in the present doesn’t speak to what traits were like thousands of years ago) (Bailey, 1997).
Bailey (1997) brings up important arguments against the use of heritability, and even discusses fallacious writing from Rushton on the matter:
Rushton (1995), for example, thinks that if observed differences among the
racial groups that he defines are higher for traits that have high heritability within the groups, the hypothesis of genetically caused differences among the groups is supported.
Bailey (1997) then goes on to discuss three lakes: Otter lake, Welcome lake, and Bark lake. Otter lake has very high primary production, while Bark lake has very little and Welcome lake is somewhere in between (you can see that ‘Otter’, ‘Bark’ and ‘Welcome’ lakes are analogies for ‘Orientals’, ‘Blacks’, and ‘Whites’ as said by Rushton). But there is variation within the lakes, there are high production pockets of water in Bark lake while there are low production pockets of water in Otter lake. All three lakes are visited and measurements are taken. Bailey (1997) states that his conclusion would be that they differ in how much light each receives. Bailey (1997: 131) writes:
If I substitute three groups of people for my lakes, IQ for primary production, and genes for light levels, the fallacy of the slippery scale, as applied to human behaviour genetics, becomes clear. Even if we are sure that there is a difference among groups of people in IQ, and we are sure that IQ has high heritability within
each of the groups (i.e. variation in IQ is largely caused by genetic variation), we can make no inference about the cause of differences in IQ among the groups. The differences might be caused by genetic differences or they might not, but the heritability studies within the groups can’t help us make that judgment.
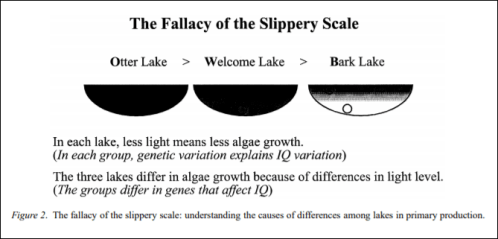
(Genes don’t cause IQ scores—or behavior—but that’s for another day.)
Heritability estimates for, say, IQ, are higher than any other trait in the animal kingdom. Heritability estimates for animal traits are low—lower than the stratospheric heritability of IQ. For example, heritability estimates of the bodyweight of farm animals is about 30 percent, which is the same for egg and milk production. Body fat in pigs and wool on sheep has a heritability of about 50 percent. But these estimates pale in comparison to the heritability estimates of IQ: estimates have been as high as 80 percent (but Schonemann, 1997 states it’s 60 percent but it’s as high as 80-90 percent today); this heritability estimate for IQ “surpasses almost anything found in the animal kingdom” (Schonemann, 1997: 104).
This high heritability estimate for IQ, of course, comes to us from the highly flawed twin studies discussed above. The reason why farmers and botanists use heritability estimates is that they can perfectly control the environment, and therefore get accurate—or close enough to it—estimates that will help them in their breeding efforts. Conversely, for humans, environments cannot be perfectly controlled and it is, of course, unethical to rear twins, MZ and DZ, in a controlled environment. Proponents of the twin method may say “It doesn’t matter if it’s flawed, it still shows there is a genetic component to trait X!”. But as discussed by Moore and Shenk (2016), that’s irrelevant because genetic factors influence all of our characteristics.
Heritability and causation
In the final section, I will shortly discuss how people fallaciously assume that high heritability estimates imply that a trait is strongly influenced by genetic factors.
In his essay in the book Postgenomics: Perspectives on Biology After the Genome, sociologist Aaron Panofsky (2016: 167; nook version) writes:
Heritability estimates do not help identify particular genes or ascertain their functions in development or physiology, and thus, by this way of thinking, they yield no causal information.
This is important to note: to those who truly believe that heritability estimates tell us anything about causation, how could they, logically, give us causal information if genes that lead to trait variation are not identified (Richardson, 2012)?
Panofsky (2014: 102-103) writes:
Experimental evidence from plants and animals suggest that shapes of the curves cannot inferred in advance and rarely follow the smooth, nonintersecting pattern like in figure 3.2. Thus true causal interpretations of heritability are hopeless and must be abandoned. Behavior geneticists did not claim direct experimental evidence, but they thought these various indirect lines of evidence provided a reasonable set of assumptions that would enable them to interpret heritability scores causally—provided they offer apporopriate, reasonable qualifications.
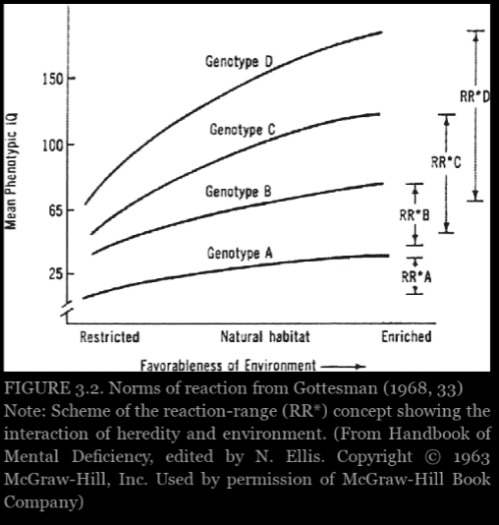
Graph from Panofsky (2014: 103)
Heritability estimates imply nothing about causation. It is about associations with variance, not identity and causes (Richardson, 2017: 69). A heritability of 0 does not mean that genes do not play a role in the development of form and function and phenotypic variation, it just means that, for whatever reason, there is little correlation between the two.
Scheneider (2007) writes (emphasis mine):
Heritability estimates apply only to groups, and are inherently inapplicable to individuals in any sense. And they do not imply causation. As Moore notes, all of these important limitations have been frequently ignored or minimized.
Reductionism
Heritability estimates imply nothing about causation. Behavior geneticists and others assume that heritability estimates will lead to ‘finding the genes’ that ’cause’ or are ‘associated with’ behavior. Their models are also, of course, extremely reductionist. It is then important to note that genes do not determine behavior. To quote Lerner and Overton (2017: 114):
Data presented in a 2016 special section of the journal Child Development indicate
that “some behaviors may be affected by only slight changes in DNA methylation,
while others may require a larger percent change in methylation; of course, the
effects are also likely bidirectional, with behavior impacting changes in methylation” [Lester et al., 2016, p. 31]. This point is key . It underscores the absurdity of genetic reductionist models: Genes do not determine behavior.
Methylation impacts behavior; behavior impacts methylation. It is the relations between methylation and behavior, not the genes acting as the “command center”, the “executive” of human behavior and development, that constitute the basic role of biology across the developmental course. This is the fatal flaw of reductionist models. Lastly, Lerner and Overton (2017: 145) write (emphasis mine):
That is, with the recent advances in understanding the role of epigenetics and recent research findings supporting this role, it should no longer be possible for any scientist to undertake the procedure of splitting of nature and nurture and, through reductionist procedures, come to conclusions that the one or the other plays a more important role in behavior and development.
[Richardson (2017: 129) also writes: “Note that this environmental source of [epigenetic] variation will appear in the behavioral geneticists twin-study as genetic variation: quite probably another way in which heritability estimates are distorted.”]
Reductionism in biology is fatally flawed. Reductionism, of course, has greatly increased our understanding of biology. However, it is time to move past the false dichotomy of nature vs nurture, and with that, move past heritability estimates since they prop up the fallacy of nature vs nurture. There is no way to separate the two since they are intertwined, but behavior geneticists would like you to believe that by studying twins raised apart will tell you anything about how ‘genetic’ or ‘environmental’ variation in a trait is in a population. Since heritability estimates are gleaned from the highly flawed studies of twins reared apart, a whole host of assumptions is poured in and these estimates are highly inflated, showing that genes influence a trait more than they supposedly do.
Twin studies, and along with it, heritability estimates, are useless for figuring out, and describing, trait variation in humans. The developmental system is more complex than the genetic reductionists (behavior geneticists) would like one to believe. The reductionist model has been heavily attacked in recent years (Regenmortal, 2004; Noble, 2008, 2012, 2015, 2016; Joyner, 2011, b; Joyner and Pederson, 2011).
Nature vs nurture has also shown to be a false dichotomy because the system develops in whichever environment it finds itself in (Oyama, 1985, 1999; 2000; Moore, 2002; Schneider, 2007)
Conclusion
Since the genetic reductionist model is wrong, along with heritability estimates (because of the nature/nurture fallacy), both should be discontinued. One of the main vehicles of these two models—twin studies—should also be discontinued. These fatal flaws of the behavior geneticists’ paradigm should be enough to discontinue these techniques in the study of human development and behavior. Heritability estimates give no causal information and they also use an outdated model of the gene; twin studies assume too many things for it to be a viable model in the discovering how traits manifest (most importantly, twin studies keep the nature/nurture fallacy alive and should be discontinued on that note only, in my opinion); and genetic reductionist models have been shown to be fatally flawed in recent years. We now have a better understanding of what a gene is today (Portin and Wilkins, 2017), and due to this, we should discontinue whatever implies the fallacy of nature vs nurture because it is irrelevant and a false dichotomy. That, alone, should be enough to discontinue twin studies and heritability estimates.

very good.
HBDers think that the only alternative to their theory is skinnerism.
are there any skinnerians left?
HBDers are dumb.
the very statistic “heritability/h^2” ASSUMES that the effects of environment and genes is additive and thus independent.
the idea that a given environment might be good for one genome and bad for another is a thought they can;t think.
sad!
LikeLike
Gene/gene, gene/environment interactions screw up the model. Fisher ignored it; Jensen thought it didn’t matter. They’re wrong. Anyone who thinks that this is still a ‘debate’ needs to read this book:
Beyond Versus: The Struggle to Understand the Interaction of Nature and Nurture
LikeLike
but perhaps unlike rr i do believe that “smart” and “dumb” are real phenomena AND that they can be measured (very imperfectly) by IQ tests AND that rank order in any given population will eventually be predictable from the genome to some extent./
LikeLike
They are ‘real’ phenomena, but we have yet to define ‘smart’ and ‘intelligent’.
I highly doubt that we’ll be able to predict “IQ” from the genome.
LikeLike
why do you deign argue with peepee?
you’re WAY smarter.
she’s a black lesbian. how can you not know this by now?
seriously dude…
were you banned from unz?
i was!
or LotB?
you can get around it with a proxy.
LikeLike
What was your username on UNZ?? id love to read your comments.
LikeLike