
Home » Posts tagged 'Philosophy'
Tag Archives: Philosophy
Evolutionary “Progress”: Gould’s Full House Argument
1600 words
Wind back the tape of life to the origin of modern multicellular animals in the Cambrian explosion, let the tape play again from this identical starting point, and the replay will populate the earth (and generate a right tail of life) with a radically different set of creatures. The chance that this alternative set will contain anything remotely like a human being must be effectively nil, while the probability of any kind of creature endowed with self‐consciousness must also be extremely small. (Gould, 1996. Full House)
Wind back the tape of life to the early days of the Burgess Shale; let it play again from an identical starting point, and the chance becomes vanishingly small that anything like human intelligence would grace the replay. (Gould, 1987. Wonderful Life)
Wind back the clock to Cambrian times, half a billion years ago, when mammals first exploded into the fossil record, and let it play forwards again. Would that parallel be similar to our own? Perhaps the hills would be crawling with giant terrestrial octopuses. (Lane, 2015: 21. The Vital Question)
I first read Full House (Gould, 1996) about two years ago. I never was one to believe in evolutionary “progress”, though. As I read through the book, seeing how Gould weaved his love for baseball into an argument against evolutionary “progress” enthralled me. I love baseball, I love evolution, so this was the perfect book for me (indeed, one of my favorite books I have read in my life—and I have read a lot of them). The basic argument goes like this: There are more bacteria on earth than other animals deemed more “advanced”; if evolutionary “progress”—as popularly believed— were true, then there would be more “advanced” mammals than bacteria; there are more bacteria (“simpler: animals) than mammals (more “advanced” animals); therefore evolutionary “progress” is an illusion.
Evolutionary “progress” is entrenched in our society, as can be seen from popular accounts of human evolution (see picture below):

This is the type of “progress” that permeates the minds of the public at large.
Some may look at the diversity of life and conclude that there is a type of “progress” to evolution. However, Gould dispatches with this type of assertion with his drunkard argument. Imagine a drunkard leaving the bar. There is the bar wall (the left wall of complexity) and the gutter (the right wall of complexity). As the drunkard walks, he may stumble in between the left wall and the gutter, but he will always end up in the gutter every time.
Gould explains then explains his reasoning for using this type of argument:
I bring up this old example to illustrate but one salient point: In a system of linear motion structurally constrained by a wall at one end, random movement, with no preferred directionality whatever, will inevitably propel the average position away from a starting point at the wall. The drunkard falls into the gutter every time, but his motion includes no trend whatever toward this form of perdition. Similarly, some average or extreme measure of life might move in a particular direction even if no evolutionary advantage, and no inherent trend, favor that pathway (Gould, 1996: 151).
The claim that there is a type of “progress” to evolution is only due to the fact—in my opinion—that humans exist and are the most “advanced” species on earth.
It seems that JP Rushton did not read this critique of evolutionary “progress”, since not even a year after Gould published Full House, Rushton published anew edition of Race, Evolution, and Behavior (Rushton, 1997) where Rushton argues (on pages 292-294) that there is, indeed, “progress” to evolution. He cites Aristotle, Darwin (1859), Wilson (1975) Russell (1983, 1989; read my critique of Russel’s theory), and Bonner.
To be brief:
The Great Chain of Being (which Rushton’s r/K selection theory attempts to revive) is not valid; Wilson’s idea of “biological progression” is taken care of by Gould’s drunkard argument; Bonner asks why there has been evolution from simple to advanced, and this, too, is taken care of by Gould’s drunkard argument, and finally Dale Russel’s argument about the troodon (I will expand on this below).
Rushton claims that Russell, in his 1989 book Odysseys in Time: Dinosaurs of North America (which I bought specifically to get more info on Russel’s thoughts on the matter and to get more information for an article on it) that “if [dinosaurs] had not gone extinct, dinosaurs would have progressed to a large-brained, bipedal descendent” (Rushton, 1997: 294). Either Rushton only glanced at Russel’s writings or he’s being inherently dishonest: Russel claimed that had the dinosaurs not gone extinct, one dinosaur—the troodon—would have evolved into a bipedal, human-like being. Russel made these claims since the troodon had EQs about 6 times the size of the average dinosaur and they ran on two legs and had use of their ‘hands.’ So, due to this, Russel argues that had the dinosaurs not gone extinct, the troodons could possibly have been human-like. However, there are two huge problems for this hypothesis.
In the book Up From Dragons, Skoyles and Sagan (2002: 12) write:
But cold-bloodedness is a dead-end for the great story of this book—the evolution of intelligence. Certainly reptiles could evolve huge sizes, as they did over vast sweeps of Earth as dinosaurs. But they never could have evolved our quick-witted and smart brains. Being tied to the sun restricts their behavior: Instead of being free and active, searching and understanding the world, they spend too much time avoiding getting too hot or too cold.
So, since dinosaurs are cold-blooded and being tied to the sun restricts their behavior, if they would have survived the K-T extinction event, then it is highly implausible that they would have grown brains our size.
Furthermore, Hopson (1977: 444) writes:
I would argue, as does Feduccia (44), that the mammalian/avian levels of activity claimed by Bakker for dinosaurs should be correlated with a great increase in motor and sensory control and this should be reflected in increased brain size. Such an increase is not indicated by most dinosaur endocasts.
Gould even writes in Wonderful Life:
If mammals had arisen late and helped to drive dinosaurs to their doom, then we could legitimately propose a scenario of expected progress. But dinosaurs remained dominant and probably became extinct only as a quirky result of the most unpredictable of all events—a mass dying triggered by extraterrestrial impact. If dinosaurs had not died in this event, they would probably still dominate the large-bodied vertebrates, as they had for so long with such conspicuous success, and mammals would still be small creatures in the interstices of their world. This situation prevailed for one hundred million years, why not sixty million more? Since dinosaurs were not moving towards markedly larger brains, and since such a prospect may lay outside the capability of reptilian design (Jerison, 1973; Hopson, 1977), we must assume that consciousness would not have evolved on our planet if a cosmic catastrophe had not claimed the dinosaurs as victims. In an entirely literal sense, we owe our existence, as large reasoning mammals, to our lucky stars. (Gould, 1989: 318)
I really don’t think it’s possible that brains our size would have evolved had the dinosaurs not gone extinct, and the data we have about dinosaurs strongly points to that assertion.
Staying on the topic of progression and brain size, there is one more thing I want to note. Deacon (1990a) argues that fallacies exist in the assertion that brain size progressed throughout evolutionary history. One of Deacon’s fallacies is the “evolutionary progression fallacy.” The concept of “progress” finds refuge “implicit expression in the analysis of brain-size differences and presumed grade shifts in allometric brain/body size trends, in theories of comparative intelligence, in claims about the relative proportions of presumed advanced vs. primitive brain areas, in estimates of neural complexity, including the multiplication and differentiation of brain areas, and in the assessment of other species with respect to humans, as the presumed most advanced exemplar” (Deacon, 1990a: 195).
This, in my opinion, is the last refuge for progressionists: looking at the apparent rise of brain size in evolutionary history and saying “Aha! There it is—progress!” So, the so-called progress in brain size evolution is only due to allometric processes, there is no true “progress” in brain size, no unbiased allometric baseline exists, therefore these types of claims from progressionists fail. Lastly, Deacon (1990b) argues that so-called brain size progress vanishes when functional specialization is taken into account.
Therefore it is unlikely that dinosaurs would have evolved brains our size.
In sum, there are many ways that progressionists attempt to show that there is “progress” in evolution. However, they all fail since Gould’s argument is always waiting to rear its head. Yes, some organisms have evolved greater complexity—i.e., moved toward the right wall—though this is not evidence for “progress.” Many—if not all—accounts of “progress” fail. There is no “progress” in brain size evolution; there would not be human-like dinosaurs had the dinosaurs not gone extinct in the K-T extinction event. We live on a planet of bacteria, and since we live on a planet of bacteria—that is, since bacteria are the most numerous type of organism on earth, evolutionary progress cannot be true.
Complexity—getting to the right wall—is an inevitability, just as it is an inevitability that the drunkard would eventually stumble to the gutter. But this does not mean that there is “progress” to evolution.
The argument in Gould’s Full House can be simply stated like this:
P1 The claim that evolutionary “progress” is real and not illusory can only be justified iff organisms deemed more “advanced” outnumber “lesser” organisms.
P2 There are more “lesser” organisms (bacteria/insects) on earth than “advanced” organisms (mammals/species of mammals).
C Therefore evolutionary “progress” is illusory.
The Concepts of Racialist Race and Socialrace
2500 words
I have chronicled Hardimon’s minimalist and populationist race concepts in a few different articles. They show that race is a biological reality. The arguments that show that race exists are sound. Hardimon, unlike Spencer, distinguishes social from biological concepts of race. For Spencer, race is both a social and biological concept. For Hardimon, race is either a biological or social concept which is why he delineates the social concept from the biological concepts. The two concepts—racialist race and socialrace—are similar, and because they are similar they will both be discussed in the same article.
The racialist concept of race
The racialist concept of race is the first concept of race that Hardimon (2017) defines. He sets out six things that need to be true of human genetics, in his eyes, for the racialist concept of race to be true:
(a) The fraction of human genetic diversity between populations must exceed the fraction of diversity between them.
(b) The fraction of human genetic diversity within populations must be small.
(c) The fraction of diversity between populations must be large.
(d) Most genes must be highly differentiated by race.
(e) The variation in genes that underlie obvious physical differences must be typical of the genome in general.
(f) There must be several important genetic differences between races apart from the genetic differences that underlie obvious physical differences.
Note: (b) says that racialist races are genetically racially homogeneous groups; (c)-(f) say that racialist races are distinguised by major biological differences.
Call (a)-(f) the racialist concept of race’s genetic profile. (Hardimon, 2017: 21)
The racialist race concept, as opposed to the populationist and minimalist race concepts, propose to rank races on traits such as intelligence, morals, and cultural characters to different races. Though, he does strawman the racialist concept (which would be the HBD concept of race, I’d say) because he says things like “To be a member of a particular race would be to be a particular person who is disposed to behave in certain ways. Because of this, if racialist races existed, race would constitute a very significant kind.” This statement aside, though, race does not constitute a very significant kind, it constitutes a modest biological kind.
If one condition for the racialist concept of race is said to be that of an essence of different races, then the racialist concept of race is surely false. This is due to population thinking. Population thinking is a type of thinking that maintains that there is no single way in which genotypes are expressed by genotypes. Thus, there is no phenotypic or genotypic property that could play the role of racial essence. “Since there are no biological essences, there are no racialist race essences, and since there are no racialist race essences, there are no racialist races. The existence of racialist races is incompatible with a broad structural principle of biology” (Hardimon, 2017: 20).
Hardimon has many arguments against the existence of racialist races, including:
The corresponding argument against the truth of the racialist concept of race is that science has not found it to be the case that members of the groups thought to be racialist races share a very large number of important properties by virtue of which they count as members of such groups. Nor does it seem likely that science will find that members of groups thought to be racialist races share a very large number of such properties. The results of Lewontin’s 1972 study and Rosenberh and colleague’s 2002 study strongly suggest that it is extremely unlikely that there are many important genetic differences between races apart from the genetic differences that underlie the obvious physical differences. (pg 24)
Another argument he puts for is that if racialist races exist, then the races would be sharply distinguished between phenotypic and genotypic characteristics. Though, since most of the variation between human races are clinal, he argues, “human populations are not sharply distinguished from one another along a broad range of phenotypic and genotypic dimensions. It follows from this that there are no racialist races” (pg 25).
Hardimon also says that, if racialist races did indeed exist, human populations would be sharply divided by skin color, yet they aren’t. These differences between races are continuous and vary between populations, thus racialist races do not exist. He also says since genetic variation in Homo sapiens is nonconcordant “there are no racialist races” (pg 25).
The likelihood that racialist races exist is especially low relative to the available alternative hypotheses, which indlude the hypothesis that there are no races, period, and the hypothesis that, whereas racialist races do not exist, minimalist races do exist. It is safe to conclude that there are no racialist races, period. (pg 25)
The racialist race concept lacks scientific respectability, it does not represent any “facts of the matter“, and it “supports and legalizes domination” (Hardimon, 2017: 62). It is therefore, socially constructed in a pernicious sense. Racialist races are both ideological and social constructions which then purport to pick out biological kinds.
The concept of socialrace
Socialrace is simply defined as the nonracialist concept of social groups that are taken to be racialist races. Socialraces refer to:
(1) a social group that is taken to be a racialist race,
(2) the social position is occupied by a particular social group that is a socialrace, or
(3) the system of social positions that are socialraces. (Hardimon, 2017: 131)
Socialraces are social groups that are taken or thought to be racialist races. Thus, they are wrongly taken to be racialist races. The two concepts socialrace and racialist race are similar—they both are hierarchical. Since socialrace is a social reality, then it must play a role in our everyday social lives.
Hardimon discusses many things regarding socialrace that many readers—myself included, on some of the things—would disagree with. For instance, he states that “institutional racism”. Hardimon claims that institutional racism “obtains when and where socialrace obtains” (pg 133).
He argues that socialrace is “inter alia a relation of power“, and so the institution is also characterized “by the unequal distribution of social goods such as liberty and opportunity, income and wealth, and the bases of self-respect. Socialrace is a system of advantages (purportedly) based on racialist race” (Hardimon, 2017: 133).
He then argues that differences in socialrace are associated with differences in life outcomes of the socialraces. Thus, “the belief that people are members of a biological race . . . is essential to the social construction of races” (Bernard Boxill, quoted in Hardimon, 2017: 133). Of course, the belief that people are members of a biological race is essential to the social construction of races, because the biological correlate of these socially constructed racial groups is the minimalist concept of race.
Socialrace, of course, has a biological correlate. That biological correlate is minimalist races. Minimalist races can be understood through the populationist race concept. Of course, saying that socialrace has a biological correlate in minimalist race does not necessarily mean that there are corresponding minimalist races for every socialrace. For example, “Hispanics/Latinos” can be said to be a socialrace, but they do not have a corresponding minimalist race because they do not genetically transmit distinct phenotypic characters which correspond to geographic ancestry. Using this terminology, the Irish were, at one point, a socialrace, whereas the Jews are treated as a socialrace today, when they are a subrace of the Caucasian race (they are not their own separate race, that’s like saying the British or Germans are a separate race). Minimalist race then appears to be a necessary condition for socialraces because they pick out real patterns of distinct physical features which correspond to geographic ancestry.
Socialraces, though, do not need to exhibit patterns of visible physical features; even if no such differences existed, groups could still be socialraces, for instance, social classes and other groups can be constructed to be socialraces. The concept of socialrace is a distinctive “race” concept (Hardimon, 2017: 139). Thus, the concept of socialrace is a distinctive concept and it is needed (because it discusses social realities). The idea of a social system based on the racialist concept of race can, therefore, said to be the idea of socialrace (Hardimon, 2017: 140).
Socialrace is clearly separate from the term ‘race’, and so, it is not a ‘race’ concept like the other three concepts, it is distinct with the “socialrace” moniker. This is how we show that, when talking about races in a social sense, there are differences between this concept and the scientific minimalist and populationist race concepts. One can think of it this way: the socialrace concept of race can be of use for sociologists and others whereas the minimalist and populationist concepts of race can be useful for biologists and population geneticists.
Whether or not a group is counted as a socialrace is contingent on whether or not the group in question is treated as a socialrace by the larger society. So, in this sense, “Hispanics/Latinos” can be said to be a socialrace, but, remember, they do not have a corresponding minimalist (populationist) race.
One is “properly” counted a member of a socialrace SR if one in fact satisfies the socially accepted criteria in the correlative putative racialist race. Thus, for example, a person belongs to the socialrace black (in the United States) if he or she has any identifiable sub-Saharan African ancestry because he or she satisfies accepted US criteria for bring a member of the racialist race black.
To “pass” (for example, for white) in a system of socialrace is to be taken to be white (to satisfy the socially accepted criteria for whiteness) despite the dact that one does not satisfy those criteria (for example, by virtue of possesion of “one drop” of “black blood”). The possibility of “passing” (in a given society) points to an important variable in the practical significance of socialrace membership. An individual counts as a member of a socialrace cimply by virtue of satisfying the socially accepted criteria for membership in the corresponding racialist race. Socialrace membership is itself a real social status with real social consequences. In the case of the socialrace black, subjection to antiblack racism is a standing possibility. But the actial practical signifigance of membership in a socialrace will vary with the degree to which the individual is subject to the norms associated with the racialist race to which the individual is taken to belong. (Hardimon, 2017: 144)
Socialraces are a real, social reality. They have a biological correlate in minimalist races and are taken to be racialist races, but racialist races do not exist therefore socialraces are social, not biological, in nature, even though the minimalist race concept can be said to be its biological correlate.
My contention is that the concept I have reflectively uncovered is aleeady in general circulation without being fully recognized as the concept that it is. I have endeavered to make it possible to get a proper hold on the concept and to secure an adequate reflective understanding of its content. If the reader thinks that my account of the concept SOCIALRACE captures a notion she or he has already been using, so much the better. I hope to have clarified that concept. As for the word ‘socialrace’, my hope is that it catches on, that its dissemenation promotes understanding of the phenomenon of socialrace, and that this in turn contribubtes to the dismantling of the latter’s existence.
Hardimon says that the concept of SOCIALRACE can help us better grasp the phenomenon of socialrace. Socialrace is real and illusory, it has “real causal powers (for example, causing people’s death) and that is illusory insofar that it appears to be racialist race” (Hardimon, 2017: 172). Socialraces exist and are a significant social reality, especially since socially determined categories can make biological realities (Kaplan, 2010).
When discussing socialraces, we can say that Jews are sometimes taken to be a socialrace (although they are a part of the Caucasian race) as are Arabs (who are also Caucasian, and thusly not a separate race from Europeans or Jews). However, the US Census Bureau categories take these groups to be “white”; that is the socialgroup “white” in the United States.
Regarding the black socialrace, they comprise numerous “Hispanic/Latino” populations (such as many Dominicans, some Puerto Ricans, Nicaraguans and other Latin American countries with high African admixture). Australian Aborigines and Pacific Islanders can be said to be part of the black socialrace in America, too. The fact of the matter is, socialrace in America just pretty much chooses features that people “think” go with race A, and if they see similar-looking people from two different continents, they will assume that they are part of the same race (as is the case with Aborigines and Pacific Islanders being black, even though they are a distinct group; McEvoy et al, 2010; Spencer, 2014).
Lastly, are Asians. In America, “Asians” are taken to be just East Asians (though in the UK when they talk about “Asians” in their police statistics or in the news, they most probably mean Pakistanis. They are not wrong, they are on the Asian continent. However, that is to mislead the people into thinking that what people term Asian (East Asians) are committing the crimes or whatnot, when it’s Pakistanis. The socialrace of Asian in America comprises East and South Asians (some would include Indians, too but they are Caucasian). The socialraces closely mirror Rushton’s three main races, but they are arbitrary, putting populations into groups where they do not belong.
Conclusion
As can be seen, the concepts of socialrace and racalist race are similar. These two concepts are needed to understand each other. Most people, when talking about race, discuss the socialrace concept so it would do them some good to read up more on the concept itself. The racialist concept of race purports to pick out biological kinds and then rank them on a hierarchy in a slew of different traits (which are not physical). Racialist races also purport that racial essences exist, but since essences as a whole do not exist, then racial essences do not exist either. Further, since genetic variation in Homo sapiens is nonconcordant, racialist races do not exist.
Socialrace is simple. Most likely, however you’ve thought about race throughout your life is the socialrace concept. It has a biological correlate in minimalist races. For socialraces to exist, there must be distinctive visible patterns of visible physical features which correspond to geographic ancestry. Thus, minimalist race is a necessary condition for socialrace. Socialrace is a social reality. And if socialrace is a social reality, then it must play a role in our everyday lives.
When these two concepts are looked at together, we can see how and why both of the concepts have been around for as long as they have: they purport to pick out a biological kind, a distinct biological kind. Racialist races do not exist, but just because racialist races do not exist does not mean that socialraces do not exist, because even if, say, minimalist or populationist races did not exist, the concept of socialrace would still be important because socially determined categories can make biological realities (Kaplan, 2010).
How Much Admixture?: On Social Isolation, the One-Drop Rule, and the Maintenance of Races
3000 words
How much admixture does it take for one race to no longer exist? The answer to the question is intuitive, and using Hardimon’s (2017) minimalist race concept, it is also easily answerable on logical grounds. For example, the answer to the question will show that the “one-drop rule” (that “one drop” of “black blood” makes one black) doesn’t make logical sense. These kinds of holdovers are from the racialist concept. Racialist races do not exist, therefore the concept of the “one-drop rule” does not either, since there are no facts of the matter the two concepts explain.
Social Isolation
The maintenance of the races that current exist depend on, at the moment, social barriers to reproduction, such as racism, segregation, differences in culture and class, role segregation and racial discrimination. Thus, social isolation is important for the maintenance of the current races. Social isolation, like geographic isolation (i.e., oceans, mountains, deserts, etc.) impedes racial interbreeding and thus ensures the continuation of the genetic transmission of distinct patterns of visible physical features which correspond to geographic ancestry.
Social isolation mechanisms have been in effect for hundreds of years, which began with the advent of African slavery to the New World. Laws against miscegenation existed in some states (Phillips, Odunlami, and Bonham, 2007), which is part of the reason why it’s (an unspoken) taboo to racially intermarry and bear children with someone not of their own race. Due to this, the few interracial unions that did produce children were specifically barred—in the eyes of society—to only be able to have children with others of their same socialrace at the lower ends of the social hierarchy.
Social isolation mechanisms have ensured the continuation of human races after the discovery of the New World when the geographic isolation mechanisms began breaking down due to exploring new lands. These isolating mechanisms on the populace ensured little admixture in the European population, but compared to European Americans, African Americans have a higher percentage of the opposite admixture. Understanding racial admixture and the genetic transmission of distinct visible physical features which correspond to geographic ancestry is extremely important to understanding when races “disappear” due to inbreeding.
Therefore, social isolation—ever since 1492—and the laws/rules that came after the breakdowns of geographic isolation between races still ensured the existence of the races as we know them today. Social factors acted as de facto physical barriers that impeded the races from breeding, thusly keeping their visible physical features intact, which means keeping their racial phenotype intact since races are defined—most importantly—on the basis of visible physical features. Social isolation can, clearly, be just about as “strong” as geographic isolation, since the social repercussions of interracial unions may exile them from the groups they were in. Thus, people would be wary of interracial unions, even if—as it seems—our culture in America seems to be swaying towards inclusivity in regard to interracial relationships, people still generally associate with and date people who look like themselves and their parents (see below).
How Much Admixture?
How much admixture can one race take before said race ceases to exist? Since C 1 (a group is distinguished from another group on the basis of distinct visible physical features) doesn’t require sharp lines between said visible physical features, C 2 (members linked by peculiar ancestry) also doesn’t require that all of the ancestors of Rs (races) be Rs.
The best possible example for an answer to the question of “How much admixture?” is simple. Think of Europeans (a subrace of the Caucasian race). When Europeans interbreed with non-Europeans, they begin to lose their distinct pattern of visible physical features which correspond to their geographic ancestry. Thus, in the case of Europeans, the answer to the question of “How much admixture?”, meaning “How much interbreeding can the European subrace take before it is “bred out” of existence?” is, of course, not too much.
Think of a union between a black woman and white man (using the social race designation; their populationist race is African and Caucasian, respectively). The child the woman bears will share some of her physical features, but barely. The baby will look more like the non-European parent, but of course, a baby who is the product of the union between an African and European will share features with both parents, and thus, the baby can “roughly fit the pattern” of a minimalist race. We can easily explain this: mixed-race individuals can err, physically, to one minimalist race over another because they are the products of individuals who do fit the patterns (of visible physical features which correspond to geographic ancestry).
Contrary to the alarmist claims heard in the media and from the altright, trends in interracial marriages do not indicate that minimalist (populationist) races are coming to an end (in this case, the white (social) race).
It is true that in the modoern (post-1492) world there is vastlty more racial interbreeding than there was before 1492. And if one is referring to the very long run, then races are almost certainly on their way out. But it is one thing to say that the human races will cease to exist at some point in the distant future and quite another to say that they are likely to disappear anytime soon. It is by no means clear that we are in an epistemic position to make the latter claim.
Contrary to what some writers suggest, recent trends in racial intermarriage in the United States do not indivate the imminent end of populationist (or minimalist) races. 5 The skyrocketing rates of intermarriage in this country notwithstanding, it remains true that the vast majority of Americans continue to marry within their own conventionally designated racial group. Despite the remarkable fact that the multiracial, multi-ethnic Americans have apparently become the fastest-growing demographic group in the United States, their numbers are still swamped by individuals who are members of a single continental-level minimalist races. 6 I don’t think that the significant fraction of DNA traceable to “Europeans” in most black Americans, and the small but real fraction of DNA traceable to “Africans” in white Americans, makes the end of the populationist (or minimalist) race significantly more imminent.
There is no evidence of which I am aware indicating that the rate at which racial interbreeding in the United States (or anywhere else) is occurring is one that would lead to the elimination of all racial differences—a situation in which no two groups could be distinguished on the basis of patterns of visible physical corresponding to differences in geographic ancestry—in the near future. To sum up: the increase frequency of encountering individuals of mixed racial ancestry does not mean that the concept of race is going to go out of business anytime soon. (Hardimon, 2017: 122)
Yaeger et al (2009) show that, in their sample, self-identification as African American is a reliable indicator of ancestry. Their findings also “suggest that self-reported race and ancestry can predict ancestral clusters, but do not reveal the extent of admixture.” Thus, self-identified race—even in the presence of admixture as is the case with African Americans—can show the racial category that an individual belongs to (based on their ancestry).
Hardimon (2017: 49) articulates a simple rule that employs the minimalist concept of race:
If both parents of an individual belong to one particular racial group R, that individual will belong to R.
What happens, however, if one parent belongs to R1 and the other parent belongs to R2. The minimalist concept of race does not say. Still less does it tell us what one’s race is if one’s grandparents belongs to an R1, another to R2, another to R3, and another to R4. This is a further respect in which the minimalist race concept is vague.
Particular conceptions of race (for example, the infamous “one-drop rule”) may specify the race of the individuals of “mixed” parentage, but the minimalist concept of race does not. The idea that a genune concept of race must specify the race of each individual is a hangover from the racialist race concept. Recall here that the minimalist racehood is not defined in terms of the characteristics of the individuals who belong to races. It is defined in terms of characteristics of groups.
So, the minimalist concept of race is vague, just like the populationist concept. But we can make one claim on the answer to the question “How much admixture?”: “Once a race loses its specific phenotype due to racial interbreeding, then the race ceases to exist.”
One drop?
The one drop rule (also known as the law of hypodescent), is a form of racial essentialism (Perez and Hirschman, 2009), which states that “one drop” of another, inferior (on the basis of racialist races) race’s blood denotes him to the inferior race in the social hierarchy. The one drop rule was created back during the slave days and signified who could breed with who, on the basis of how “pure” their blood was. It was, and still is today, a way for race deniers to deny the existence of race.
The one-drop rule stated that anyone with one black ancestor was classified as black (Pauker et al, 2009). That is, his position on the socialrace hierarchy (a hierarchy since it’s based on the false racialist race concept) is based on the fact that he has one black ancestor. Due to this, and other differing amounts of admixture in certain ethnic groups and other social groups taken to be races, people have—fallaciously—stated that races do not exist since the unions of two separate races “erases” one, or both, of the races in question.
This rule helped to ensure the maintenance of populationist races, since society frowned upon interracial marriage. This, obviously, was a social custom. The Jim Crow laws helped to ensure the maintenance of the physical characteristics of the races in question, though the laws were enacted to ensure the “racial purity” (whatever that is) of the European race, it helped to ensure lower amounts of admixture in black Americans. Thus, black Americans would be expected to self-identify as black (Liebler and Zacher, 2017).
Liebler and Zacher (2017)‘s data “supports the notion that this “rule” has some power even today, as there are almost 30 times as many people reporting that they are racially black with American Indian ancestry (weighted N=522,607) as there are people reporting American Indian race with black ancestry (weighted N=16,226).” Bryc et al (2015) show that, despite the expectations of the one drop rule “individuals identify roughly with the majority of their genetic ancestry.”
Most people in one sample that had less than 20 percent African ancestry identified as white. In the US, “Latinos” (a social-race) were estimated to have 65.1 percent European, 6.2 percent African, and 18.6 percent Native American DNA. Overall, 3.5 percent of European Americans had 1 percent or more African ancestry, while 1.4 percent of self-reported European Americans had were estimated to carry at least 2 percent African ancestry (Bryc et al, 2015).
Importantlty, Guo et al (2014) write:
The one-drop rule represents an important case in which social context trumps bio-ancestry. When asked to classify into a single race, most individuals with 30 % to 60 % African ancestry self-report as black; virtually all respondents with >60 % African ancestry self-classify as black. In contrast, a substantially higher proportion of European ancestry is “required” to self-classify or to be classified by an interviewer as white than the proportion of African ancestry necessary to self-classify or be classified as black. However, when given the option of identifying as multiracial, the majority of individuals with 40 % to 60 % African ancestry in both ROOM and Add Health and substantial proportions of individuals with >60 % African ancestry in ROOM stopped self-classifying as only black and primarily chose a multiracial classification.
“The infamous one-drop rule is peculiar to this country [America] but it is a feature of the American conception of race, not the minimalist concept of race.” (Hardimon, 2017: 56) The one-drop rule is a clear tell to how the socialrace concept acts. It is an essentialist concept, which means that it is necessarily racialist—since “one drop” of black blood makes one black—according to the rule.
The Maintenance of Races
It is possible that one society could take social measures to ensure the existence of their specific racial phenotype (that is, the existence of their minimalist race or subrace). Such a society would have to grapple with the moral and ethical underpinnings of such measures to ensure the maintenance of their phenotype (see Glannon, 2001’s book Genes and Future People for an extensive review of the moral, political, social, and ethical implications of human genetic engineering). This could also include genetic modification, though sound arguments exist that show that the way most people view genetic modification depends on a “strong view” of genetic determinism, which is false (Resnick and Vorhaus, 2006). However, it is possible that, through the will of the people in the society, that social isolation can lead to a de facto “physical” isolation through the social norms of the society in question.
However, since the races as they currently are are in no danger of non-existence, such measures, while they would (presumably) work, do not need to be taken. Such measures, though, do not need to be taken, since most people want to court with others who look like themselves, and those who are more likely to look like themselves are people of their own ethny, which is to say, people of their own populationist race. Thus, social measures to ensure the maintenance of races do not need to be taken.
As noted above, certain concepts from the days of the one drop rule are still in effect today, as a holdover from the days of Jim Crow and before. Some of these holdover concepts, though, help to maintain the races we know today. However, there is a possibility that our populationist races, too, have benefits socially constructed. Hardimon (2017: 126) writes (emphases his):
If populationist races exist, the role human action plays in their maintenance is rather more pronounced then the role it played in their genesis. Insofar as social norms and practices prohibiting or discouraging intermarriage have been the primary mechanisms preventing racial interbreeding since 1492, the maintenance of the separation has been intentional: this outcome is the very point of the discriminatory activity and practices in question. There is thus an especially strong sense in which, if populationist races exist, populationist race has been socially constructed since 1492.
Hardimon (2017: 126) goes on to say that the maintenance of populationist races “is not a natural process outside of human control”, nor is it “immutable or inalterable“, while “its existence is not an invariant, unchangeable,”natural” fact” and “The continued existence of populationist races, if it is a fact, is a fact within our power to change.” Thus, if populationist races exist (and they do), they exist by virtue of existing in nature.
So the races are not in danger of non-existence anytime soon, since the percentage of interracial unions are not too high compared to those who marry within their populationist races. The maintenance of populationist races comes down to—and will come down to, as long as humans are around—to social policies, whether enacted by state/country governments or the people themselves, sans any laws on miscegenation.
It has been said that we are attracted to people “who look like us“, “who look like our parents“, and “‘who are more similar to ourselves“. This means—NECESSARILY—that people are more likely to be attracted to people of their own race/ethnic group. People “who look like us” are co-ethnics and people of the same racial background; people who “look like our parents”, are, again, people who would share the same geographic ancestry. Since the physical features that delineate races are genetically transmitted from parent to offspring, then, people are more likely to be attracted to people of their same race. Finally, “people more similar to ourselves” doesn’t necessarily mean “people more racially/ethnically similar to ourselves”, since, of course, there are many other things that individuals have in common other than their race/ethnic group. However, it has been established that we are attracted more to people who share more similar genes than ourselves (Rushton,1997, 1998; Sebro et al, 2017). Thus, logically, since we are attracted to people who look like ourselves and our parents, we are attracted to people of our own ethnicity/race, as a matter of fact.
Conclusion
The question “How much admixture does it take for one race to no longer exist” is answered simply once the term “RACE” is defined: the amount of admixture it takes for one race to be “bred out” of existence is proportional to the amount of admixture it takes for one race’s physical features which correspond to geographic ancestry which are exhibited by the real group in question (this case being a subrace of a minimalist/populationist race). Europeans can’t take “much”, if any, other admixture, otherwise the traits that make Europeans European (which are, of course, not mutually exclusive to them, but the traits they—and their ethnies—exhibit are distinct) will disappear and so one of the Caucasian subraces will disappear as well. Social isolation, at the moment, is maintaining the races as we know them—and will far into the foreseeable future (there is no evidence that they will disappear anytime soon). “Violations” of the one drop rule abound, but they mean little to the minimalist/populationist concepts of race since the visible physical features which distinguish the races remain intact.
The fact that people are more attracted to people who look like themselves and their parents is an implicit way of saying that people are more attracted to people who are physically similar to themselves—that is, racially/ethnically similar to themselves—and shows that the races will not be going anywhere for the foreseeable future.
Human races will continue to exist as long as the social barriers that impede racial interbreeding remain. (Of course, if these social barriers did not exist, a majority of people still would court people who look like themselves and their families.) This is evidence that, contra social laws that impede or frown upon interracial marriages, we do not need such laws/rules because people stick to their own anyway. Therefore, the races are not in danger of disappearing anytime soon.
Races and Populations: Existence and Reality
3000 words
Over at the blog Anthropology 365 the author—Adam Johnson, biocultural anthropologist—wrote an article titled Populations, Race, and The Sorites Paradox, in which he argues that, since there are no “clear lines” and they are “wuzzy”, we cannot say where one race ends and another begins, therefore race does not exist. His whole argument is largely just the continuum fallacy—that since we cannot show where one race, in this instance, ends and another begins, therefore, race does not exist. This reasoning, however, is very flawed.
The beginning of his article is concerned with laying out the sorites paradox. Imagine zero grains of sand, then continuously add grains of sand, 1, 5, 10, 100, 1000, etc. When does the heap become a pile of sand? Johnson attempts to use this logic regarding races and populations: where does one population end and another begin? (You already know where this is headed; it seems that this is the ‘argument’ that gets the most play nowadays when it comes to race-denialism and racial eliminativism when there are better, non-fallacious, arguments out there to attack the concept of race in our ontology. Using the old and tired “continuum fallacy” no longer makes sense because the objection that “Race does not exist because we cannot tell where one race ends and another begins” has been responded to numerous times, most recently (and forcefully) by philosophers of race Michael Hardimon and Quayshawn Spencer.)
He defines “population”, stating that—in biocultural anthropology—that a population is simply a group of like kinds that interbreed with each other which are separated by geographic barriers. Nothing wrong with that—it’s true. He then makes the huge leap in logic to a within-country comparison (America), showing two arbitrarily circled “populations” on the east and west coasts of America. He admits the circles are “arbitrary”, then adds another purple circle in the middle, and finally a green and purple circle in between the original circles, signifying five populations (the image can be seen below).

He says that “It is often impossible to draw neat boundaries around a group”, but I am aware of no author making any claim that it IS possible (and easy) to draw neat boundaries around groups. To do so, you only need simple conditions; and if there is any deviation out of those conditions, then the population in question do not fit the definition of what you were constructing and they can thus be removed. Johnson says “where does yellow end and purple begin?” since there is so much overlap between all five colors in this image. He says that this reasoning shows how “crude” the concept of population is regarding the accepted definition: a group of like kinds that can interbreed but are geographically separated.
One who denies Hardimon’s (2017) 3 conditions for to establish that populations are minimalist races (C1. visible patterns of distinct physical features which correspond to geographic ancestry; C2. that the members in this group are linked by a common ancestry; and C3. they must originate from a distinct geographic location) may then take to this idea that these arbitrarily drawn circles which are supposed to be “populations” (to Johnson) are then races; but Johnson never left any conditions, only a vague definition. One could argue that two of those clusters satisfy C1-C3 (that the cluster in question shares visible patterns of distinct physical features which correspond to geographic ancestry [the people who, say, make up one town in one of the arbitrarily drawn circles may have different visible patterns of distinct physical features which correspond with their ‘geographic ancestry’], that the members are linked by a common ancestry [the town they now live in, say], and they derive from a distinct geographic location [the arbitrarily drawn circle is a distinct geographic location].
However, for one to say that C1 holds for these arbitrarily drawn circles, they have to stretch the definition in order to accept random populations within a country. They then need to say that C2 refers to any type of “common ancestry” of a certain town; and that C3 then shows that they derive from a distinct geographic location. However, in regard to C2 and C3, one who would attempt such an argument would be equivocating on “geographic ancestry” and “distinct geographic location”, thusly claiming that an infinitude of races exist because the conditions are vague. While I do admit that minimalist concept is vague, in my view, it does not allow for one to equivocate on certain words used in the argument to show that any and all arbitrary populations can be called “races”; it does not work like that because there are distinctive conditions that must be met before further thinking on whether or not a population in question is a “race” or not.
Johnson then quotes Scientific American writer John Terrel who writes in his article “Plug and Play” Genetics, Racial Migrations and Human History:
“Distinguishing between races and populations is effectively making a distinction without a difference. If this comes across as sounding crazy to you, then tell me this. What is a population? How can you tell whether you are “inside” a population or “outside” it? How many of them are there “out there” in the real world? How many did there used to be? More than today, or fewer? (Now substitute in these simple questions the word “race.” Doesn’t make much difference, right?)”
What is a population? Good question. The definition left by Johnson above is alright, but we can refine it. I can simply cite Michael Hardimon’s definition of “populationist race” (Hardimon, 2017: 99; my emphasis):
“A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.”
Using this definition of race, a race is a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the groups’ geographic ancestry. Thus, with “population” having a much more non-vague definition, we can then begin to look for populations that exist in reality (not arbitrarily demarcated “populations” like Johnson did—using arbitrary circles as population groups in America).
Now that population is defined, what about the next question: “How can you tell whether you are “inside” a population or “outside” it?” Since we now have a better grasp of what “population” means in this context, then this question is simple to answer. You can tell whether you are “inside”‘ or “outside” a population by looking in a mirror and then thinking about any “population” as defined above. It really is that simple. However, it is hard when “population” is defined so vaguely, and so you get flaws in reasoning like the one from Johnson.
Now that we know that we can tell whether or not we are “inside” or “outside” a population, his next question is: “How many of them are there “out there” in the real world?” According to the definition presented by Hardimon above, there are 5 current races in the human subspecies. That’s the number of races that are ““out there” in the real world” (as opposed to a possible world we can imagine—which is not the topic of contention).
Now that we know how many of “them” [races] exist, the next questions are: “How many did there used to be? More than today, or fewer?” I won’t pretend to know the answer to this question, but I will say one thing: the number of races that used to exist in the past comes down to the number of populations that exhibit a distinctive pattern of visible physical features which are genetically transmitted by geographically and reproductively isolated founding populations. Though, the number of races that “used to” exist is irrelevant to the fact that races exist today and the number of races that do exist today.
Johnson then claims that we, in the West, have a “long history” of constructing different races. And while this is true, this does not go against the claim that biological racial realism is true. Johnson says that “We homogenized entire continents of people into essential “types” and used the assumptions intrinsic to those types to make grand statements about the “natural” divisions in the human species and the value and meaning associated.” Well, these “entire homogenized continents of people” DO fit into “types”—though they are not “essential”; there are “natural” divisions within the human species BUT one does not have to put value and meaning onto the existence of these populations that we call ‘races’, since they are based solely on distinct pattern of genetically transmitted characters which then correspond with the group’s geographic ancestry.
“Anthropology has since moved on from it’s [sic] assumption that the human species is divided up into natural kinds“, Johnson writes. It seems that Johnson is ignorant to the work of Hardimon (2017) and his racial typology using the minimalist concept of race along with its “scientific equivalent” the populationist race concept. Minimalist races are a biological kind “if only a modest one” (Hardimon, 2017: 91), and so, just because “Anthropology has since moved on from it’s [sic] assumption that the human species is divided up into natural kinds” DOES NOT MEAN THAT there are no “kinds” within the human species. The argument for the existence of minimalist races establishes the claim that the human species is, in fact, divided up into kinds:
P1) There are differences in patterns of visible physical features which correspond to geographic ancestry
P2) These patterns are exhibited between real groups, existing groups (i.e., individuals who share common ancestry)
P3) These real, existing groups that exhibit these physical patterns by geographic ancestry satisfy conditions of minimalist race
C) Therefore race exists and is a biological reality
Minimalist races exist and are biologically real; if minimalist races exist, then populationist races exist; populationist race is the “scientization” of minimalist race; minimalist races entail kinds, and so since minimalist races entail kinds then so do populationist races; therefore both concepts speak to kinds within the human species and their biological reality.
Either way, we can also accept that anthropology has moved away from the assumption that the human race is divided into kinds and not have to give up the argument for the existence of race. Instead of arguing that human races are “kinds” as Hardimon (2017) does, Spencer (2014) argues that since Americans defer to the US Census Bureau regarding race, the must be referring to biologically real groups. The US Census Bureau defers to the Office of Management and Budget. The OMB discusses “sets of” populations. K= 5 delineates populations that Americans refer to when referring to race. So since Americans defer to the Census Bureau and the Census Bureau defers to the OMB, when we Americans talk about race, we talk about proper names for population groups as denoted by the OMB—even though ‘race’ looks like a ‘kind’ term, according to Spencer (2014: 1028) “its current use in US racial discourse is that of a proper name. It is a term that rigidly designates a particular set of “population groups.” This means that race is a particular, not a kind.”
So, there are two sound arguments for the existence of race (the argument for the existence of populationist races from Hardimon and the argument for the existence of Blumenbachian partitions—which both use the same population genetics paper (Rosenberg et al, 2002) to buttress their claims that their “kinds” (Hardimon, 2017) and “partitions” (Spencer, 2014) exist in reality.
Lastly, Johnson cites Galanter et al (2012) who genotyped “populations” throughout South America:
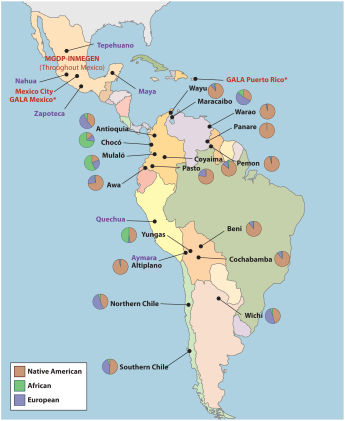
He then states that we have a bunch of South American populations here, all with differing amounts of admixture (which, of course, coincide with three of the five populationist races). He pretty much says, “How can we draw neat circles around these populations to call them “populations”, and what about those other populations not sampled in the analysis?” It makes no sense; when you’re just drawing circles anywhere on any map and then claiming that they are “populations” that satisfy a vague criteria/definition, then you don’t understand any of the newer arguments put forth by philosophers on the existence and reality of racial population groups.
He concludes the article simply:
To conclude, it’s always important to parse in our assumptions and take into account that our levels of analysis (the unit we are studying) may not represent reality. When we equivocate levels of analysis with levels of reality when examining human diversity, as Terrell says, we end up making a distinction between race and populations with no real difference. However, if we understand that the “population(s)” of interest are not reflections of reality, but merely constructed entities that represents an amalgamated web of kinship, political, biological, economic, and random histories at a particular time and place, we can avoid the trap of racial thinking (without using ‘race’) that some scholars fall in to.
He seems to be conflating two concepts here: how we view these visible physical features which correspond to geographic ancestry (our socialview of these populations) and their actual existence completely removed from our social conventions. Yes, socialraces are groups that are taken to be racialist races (that is to say, they are taken to have a specific essence particular to that race and only that race); but the concept of socialrace—the types of social values we give to these populations (think that the minimalist concept of race denotes certain social groups on the basis of distinct visible patterns which correspond to geographic ancestry; the socialrace concept is a good concept since it presents a way of thinking about (1) social groups that are taken to be races (such as ‘Latinos’/’Hispanics’); (2) the social positions that the social groups occupy; and (3) the systems of social structure of which those positions are parts (Hardimon, 2017: 139).
The “populations of interest”, are, indeed, of interest because they pick out what ‘we already know to be’ races.
Races, then, are both socially and biologically constructed. The minimalist concept of race shows the phenotypes that the socialrace concept chooses out when denoting a population its socialrace status in a given society. It shows that there are both biological and social underpinnings to racial categories—that is, there is both a “biological” and “social” realm to race in our ontology, and if we want to understand both ontologies, then we must first think of the consequences of thinking of “race” as only a biological concept and only a social concept and then—after we have thought of “race” as a biological and social concept on its own—we can think of “race” as both a social and biological phenomenon because that’s the best way to describe race in out ontology.
I find it funny how Johnson brings up “population thinking”; but I am probably thinking of it in a different way then he was in his article. When he brings up “population thinking” he wants you to think in terms of his definition of “population”, which pretty much means any group he circles is deemed a population, and thus, since there is no easy way to delineate populations from each other, therefore race does not exist (we must be eliminativist about race). Though when I think of the term “population thinking”, I think of Ernst May’s use of the phrase populationist thinking is more apt: “populationist thinking” is directly opposed to “typological thinking”: “populationist thinking” holds that there are no intrinsic “biological essences”, nor any property—or set of properties—that all, and only all, members of a population share.
For the populationist “all organisms and organic phenomena are composed of unique features and can be described in collectively only in statistical terms. Individuals, or any kind of biological entities, form populations of which we can determine the artihmetic mean and the statistics of variation. Averages are merely statistical abstractions. . . . For the typologist the type (eidos) is real and the variation is an illusion, while for the populationist the type (average is an abstraction and only the variation is real (Mayr, 1976; quoted in Hardimon, 2017: 20).
For example, “Caucasian” is a valid taxonomic category when discussing populationist races. One classified as “Caucasian” might have absolutely none of the genotypic or phenotypic markers associated with “Caucasian-ness”; that is, population thinking does not assume that any one genotype or phenotype is essential to any one population. Thus, there are no intrinsic properties that all members of a race—and only members of that race—share.
To conclude, contrary to the claims of Johnson and Terrel, race does exist and there are reasons why we should accept the existence of these population groups we call races. Johnson largely uses the old and tired continuum fallacy—the fallacy of the beard, whichever name you like—to attempt to argue that “race” does not exist. But he did not even state any conditions on what “population” entails; he just drew random, overlapping circles proclaiming “Ha! Where does X color end and Y color begin!!??” This type of thinking, though, is fallacious, as can be seen. It is completely possible to delinate races on the basis of visible physical features which correspond to geographic ancestry.
Articles like Johnson’s and Terrel’s are easy to come by: they just adopt a racial eliminativist stance on race (that it should be removed from our ontology entirely). They use fallacies like the continuum fallacy to show that since there is no clear ‘genetic line’ (see my article You Don’t Need Genes to Delineate Race) separating so-called races, therefore races do not exist (we must then take an eliminativist approach to race). I’m of the belief that the answer to the question “Does race exist?” will be—and only can be—answered by philosophers of race. We know that geographic variation exists—however small it may be. We know that we can distinguish continental populations on the basis of visible physical features. From there, it’s only a short bit of reasoning to reason, correctly, that race exists and is a biological reality (as the arguments in Spencer, 2014 and Hardimon, 2017 attest to).
The Existence and Reality of Populationist Races
2300 words
Michael Hardimon has some of the best defenses of the reality of race that I am aware of. His 4 concepts are: the racialist concept (he says racialist races do not exist, which I will cover in the future), the minimalist race concept, the socialrace concept (which also will be covered more in depth in the future) and the populationist race concept. Racialist races do not exist, according to Hardimon. However, that does not mean that race does not exist nor does it mean that race isn’t real. On the contrary, race exists and is a biological reality. Simple arguments for the existence of race do indeed exist and see where mixed-race individuals, ‘Latinos’, and Brazilians fall. (Author of the book A Theory of Race Joshua Glasgow also reviewed Hardimon’s book (Glasgow, 2018), and I also left my thoughts on his review.)
Now, minimalist races exist and are biologically real. The concept, though, is vague. It doesn’t state which populations are races, but the populationist race concept, however, does. Hardimon (2017: 99) defines populationist races:
“A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.”
Are there groups that exhibit patterns of a distinctive pattern of visible physical features which are genetically transmitted and correspond to the group’s geographic ancestry? Are there groups that belong to a biological line of descent which was initiated by geographically and reproductively isolated founding populations? The answer is, obviously, yes. Which groups satisfy the definition of populationist races? I will discuss this below.
An important question to answer is: are races subspecies? The two terms are similar. Merriam Webster defines subspecies as: “a category in biological classification that ranks immediately below a species and designates a population of a particular geographic region genetically distinguishable from other such populations of the same species and capable of interbreeding successfully with them where its range overlaps theirs.” While “race” is similarly defined. So, are races subspecies?
The fixation index (Fst) is a measure of population differentiation due to genetic structure which is estimated from SNPs or microsattelites. Generally, the accepted criterion for subspeciation is between .25 and .30. Human groups have an Fst between .05 and .15, so human groups fall way short of subspeciation. Fst estimates for humans fall between .05 and .15, which is far and away what the consensus is on the delineation of subspecies within a group of like kinds. Further, Fst does not support the existence of distinct clusters in humans (Maglo, Mersha, and Martin, 2016; it should be noted that they believe that for human races to exist, human races must be subspecies—similar views are held by philosopher of science Adam Hochman—but their contentions were addressed by Spencer, 2015). Human populations are not subspecies, and the fact that they are not subspecies does not rail against the existence of populationist races.
Hochman (2013) makes the case that in order to claim that clusters represent subspecies, four conditions have to be met: “(i) the range of allele frequency differences between genetic Fstclusters corresponding to race must be relatively uniform, (ii) there must be a determinate number of such clusters, (iii) the allelic frequencies within such clusters must be relatively homogeneous, and (iv) there must be a large jump in genetic differences between such clusters” (Hardimon, 2017: 108).
Thus, the human species does not contain subspecies in the technical sense of the word, as humans Fst estimates range between .05 to .15. This further attests to the fact that the clusters—identified by Rosenberg et al (2002)—are not subspecies. “There is no need for US racial groups to be subspecies or clades, have high genetic variation among them, or be fundamental categories in human population genetics just in order to be biologically real races. Rather, in order for US racial groups to be biologically real races, they just need to be races and biologically real (Spencer, 2015: 6).
The populationist race concept, however, does not require that a division in a species be represented by a particular Fst estimated. It further doesn’t say that Hochman’s (2013) conditions must be met in order for the clusters to be races. Therefore the populationist race concept is not a subspecies concept; there are no subspecies in our genus. Though, if we were forced to accept Hochman’s (2013) conditions (which we do not have to), human races do not exist.
Next is the concept of phylogeny. If phylogenetic is taken to in the normal biological terminology, then the question is whether or not racial lines of descent capture evolutionary significant relationships. And if “evolutionary significant relationships” are taken in the normal biological context then the answer to the question is “no.” This is because the term “evolutionary significance”, taken in the general biological terminology, is understood in a way that for a relationship between populations to be “evolutionarily significant”, then the differences between these populations must be blocked by extensive gene flow.
However, regarding the populations that we take to be populationist races, if the features of these races have adaptive significance, such as skin color for differing climates, then the populationist race concept is of interest to evolutionary biologists since biological raciation makes it possible for divisions of Homo sapiens to survive in different climates. Thus, when discussing how and why divisions of our species adapted to different climates—physically speaking—then this concept is of use to evolutionary biologists since it can explain the adaptive physical features of divisions of Homo sapiens. We then have two choices. We can then further take the idea that to be “phylogenetic”, populations must block extensive gene flow, though we can grant that populationist races may well be of interest to evolutionary biologists (due to their adaptive features that arose due to climatic adaption), despite the fact that populationist races are nonphylogenetic (Hardimon, 2017: 111).
The populationist race concept is a candidate scientific concept. This is because the concept uses biological terminology such as “reproductive isolation”, “transmitted phenotypic characteristics”, “founding population”, and “geographic ancestry.” Hardimon then discusses how and why the concept can form a scientific concept:
“… concept C has the “form” of a scientific concept in biology if
(i) it is formulated in a “biological vocabulary”,
(ii) it is framed in terms of an accepted biological outlook,
(iii) it is suitable for deployment in an accepted branch of biological inquiry, and
(iv) it presents the scientific ground of the phenomenon it represents” (Hardimon, 2017: 112).
This concept satisfies all four conditions. It satisfies (i) since it uses biological vocabulary (e.g., phenotype, reproductive isolation). It satisfies (ii) since it’s framed in what Mayr terms “population thinking” (which is the rejection of essentialism—“the view that some properties of objects are essential to them.”. It satisfies (iii) since it is suitable for deployment in ecology, ethology and evolutionary biology. Areas of study, for example, can focus on how and why differing populationist races have differing patterns of visible physical features (i.e., how and why phenotypes changed as migration occurred out of Africa into Eurasia, the Pacific Islands and the Americas). And it satisfies (iv) in that representing populationist races as having arisen from reproducively isolated founding populations.
Now which groups are candidates for populationist races? There are two conditions: (1) they exhibit distinctive patterns of phenotypic characters which correspond to that population’s geographic ancestry and (2) belong to biological lines of descent which then trace back to geographically separated and reproductively isolated founding populations.
There are populations which exhibit distinctive patterns of visible physical features which correspond to geographic ancestry, and they are Sub-Saharan Africans, Caucasians, East Asians, Native Americans and Pacific Islanders. The distinctive patterns of visible physical features are genetically transmitted, and they correspond to geographic ancestry. These populations belong to biological lines of descent which can then be traced back to geographically separated and reproductively isolated founding populations. Thus, conditions (1) and (2) are satisfied, therefore populationist races exist.
Further support for (iii) (that the populationist race concept can be deployed in the biological sciences) can be found in my article You Don’t Need Genes to Delineate Race. I discussed differences in gross morphology between the races; I discussed differences in physiognomy between the races; and, of course, the differences in geographic ancestry that caused the differences in morphology and physiognomy (see here for discussions on skin color). Differences in climate that Homo sapiens encountered after trekking out of Africa then caused the distinctive differences in visible physical features which correspond with geographic ancestry which then make up populationist races. Thus, the study of populationist races will elucidate the caused of phenotypic differences between populationist races since they exist and are a biological reality.
There is a relationship between populationist and minimalist races, though they’re defined by different concepts. However if minimalist races are populationist races, then the kind minimalist race=populationist race. “The claim that minimalist race=populationist race is analogous to the claim that water=H2O. The latter claim, since true, provides scientific insight into the nature of minimalist race” (Hardimon, 2017: 120).
Furthermore, we can assume that the populations identified by Lewontin (1972) as races can be interpreted as lending support to the biological reality of populationist races exist. We can also assume that African, Caucasians, East Asians, Oceanians, and Native Americans constitute populationist races, then Rosenberg et al (2002) show support for the biological reality of populationist races, even though the fraction of diversity separating the clusters is between 3-5 percent, this still shows that populationist races capture a portion of biological human variation, no matter how small it is.
“If it is assumed that Africans, Eurasians, East Asians, Oceanians, and Americans constitute continental-level populationist races, Rosenberg and colleagues’ 2002 study can be interpreted as providing support for the biological reality of populationist race inasmuch as it shows that a very small fraction (3-5 percent) of human genetic variation is due to difference among continental-level populationist races. Modulo our assumption, the study results indicate that populationist race is a minor principle of human genetic structure and that populationist race is a minor principle of human variation.” (Hardimon, 2017: 124)
The same points made that minimalist races are human population partitions, that races can be distinguished at the level of the gene, and that the continental-level minimalist races differ in a small number of coding genes, also carry over to the populationist race concept since minimalist race=populationist race, so the biological reality of minimalist race carry over to populationist race. So if the five populations are populationist races, then populationist race correspond to a partition of genetic variation found between the races in the human species, which is then evidence for the existence of populationist races.
The five populations that make up populationist races are Native Americans, Caucasians, East Asians, Pacific Islanders, and Sub-Saharan Africans. These populations are biologically real, and they exist. They generically transmit phenotypic characteristics across the generations; these phenotypic characteristics differ due to geographic ancestry. These populations are identified in numerous K = 5 runs. So if we assume that the five populations are populationist races then K = 5 shows the real, but small, human genetic variation found within continental-level populationist races which is how the visible patterns of visible physical features which correspond to geographic ancestry are genetically transmitted.
The populationist race concept is a candidate scientific concept. This is a way to study the small genetic variation between the continental-level clusters. Human phenotypic (and physiologic) differences arose due to adaption to different climates. Thus, since populationist race is a biological reality then studying populationist races will better elucidate how and why differences in phenotype arose.
Both the populationist and minimalist race concepts are vague, I admit. However, they’re not so vague that one could argue that villages, countrys, social classes etc are populationist races. It should be noted, though, that it is implicitly stated in the definition for populationist race, that a morphological component exists. Therefore, groups like the Amish, social classes etc. Thus, the populationist race concept gaurentees that races will be races in the ordinary sense of the word (see Hardimon, 2003). So we can take two groups—G1 and G2—and if G1 does not have any pattern of visible physical features which distinguish it from another group, G2, then G1 is not a race. These visible physical differences that distinguish races from one another are biological in nature—hair color/type, skin color, eye type, morphology etc. This gaurentees that different villages, countries, economic classes and ethnies within a race are not counted as “races”, so defined.
The thing about the populationist race concept is that it directly relates to the minimalist race concept. Once we acknowledge that races exist and are real (since minimalist races exist and are real), then we start thinking “Which populations sastisfy the conditions of populationist races?” The populationist race concept—in tandem with the minimalist race concept—shows us that the patterns in visible physical features are genetically transmitted characters which which correspond to the population’s geoprahic ancestry who belong to biological lines of descent which were initiated by geographically separated and genetically isolated founding populations. The populationist race concept supports the claim that the minimalist race concept is a biological concept and secures the existence of minimalist races since minimalist race=populationist race.
P1) The five populations demarcated by Rosenberg et al (2002) are populationist races; K = 5 demarcates populationist races.
P2) Populationist race=minimalist race.
P3) If populationist race=minimalist race, then everything from showing that minimalist races are a biological reality carrys over to populationist races.
P4) Populationist races capture differences in genetic variation between continents and this genetic variation is responsible for the distinctive patterns of visible physical features which correspond to geographic ancestry who belong to biological lines of descent which were initiated by geographically isolated founding populations.
C) Therefore, since populationist races=minmalist races, and visible physical features which correspond to geographic ancestry are genetically transmitted by populations who belong to biological lines of descent, initiated by reproductively isolated founding populations, then populationist races exist and are biologically real.
Blumenbachian Partitions and Mimimalist Races
2100 words
Race in the US is tricky. On one hand, we socially construct races. On the other, these socially constructed races have biological underpinnings. Racial constructivists, though, argue that even though biological races are false, races have come into existence—and continue to exist—due to human culture and human decisions (see the SEP). Sound arguments exist for the existence of biological races. Biological races exist, and they are real. One extremely strong view is from philosopher of science Quayshawn Spencer. In his paper A Radical Solution to the Race Problem, Spencer (2014) argues that biological races are real; that the term “race” directly refers; that race denotes proper names, not kinds; and these sets of human populations denoted by Americans can be denoted as a partition of human populations which Spencer (2014) calls “the Blumenbach partition”.
To begin, Spencer (2014) defines “referent”: “If, by using appropriate evidential methods (e.g., controlled experiments), one finds that a term t has a logically inconsistent set of identifying conditions but a robust extension, then it is appropriate to identify the meaning
of t as just its referent.” What he means is that the word “race” is just a referent, which means that the term “race” lies in what points out in the world. So, what “race” points out in the world becomes clear if we look at how Americans define “race”.
Spencer (2014) assumes that “race” in America is the “national meaning” of race. That is, the US meaning of race is just the referent to the Census definitions of race, since race-talk in America is tied to the US Census. But the US Census Bureau defers to the Office of Management and Budget (OMB). Therefore, since the US Census Bureau defers to the OMB on matters of race, and since Americans defer to the US Census Bureau, then Americans use the OMB definitions of race.
The OMB describes a “comprehensive set” of categories (according to the OMB) which lead Spencer (2014) to believe that the OMB statements on race are pinpointing Caucasians, Africans, Pacific Islanders, East Asians, and Amerindians. Spencer (2014: 1028-29) thusly claims that race in America “is a term that rigidly designates a particular set of “population groups.” Now, of course, the question is this: are these population groups socially constructed? Do they really exist? Are the populations identified arbitrary? Of course, the answer is that they identify a biologically real set of population groups.
To prove the existence of his Blumenbachian populations, Spencer (2014) invokes populational genetic analyses. Population geneticists first must make the assumption at how many local populations exist in the target species. According to Spencer, “The current estimate for humans is 7,105 ethnic groups, half of which are in Africa and New Guinea.” After the assumptions are made, the next step is to sample the species’ estimated local populations. Then they must test noncoding DNA sequences. Finally, they must attempt to partition the sample so that each partition at each level is unique which then minimizes genetic differences in parts and maximizes genetic differences among parts. There are two ways of doing this: using structure and PCA. For the purposes of this argument, Spencer (2014) chooses structure, invoking a 5-population racial model, (see e.g., Rosenberg et al, 2002).
K = 5 corresponds to 5 populational clusters which denote Africans, Oceanians, East Asians, Amerindians, and Caucasians (Spencer, 2014; Hardimon, 2017b). K = 5 shows that the populations in question are genetically structured—that is, meaningfully demarcated on the basis of genetic markers and only genetic markers. Thus, that the populations in question are meaningfully demarcated on the basis of genetic markers, this is evidence that Hardimon’s (2017b) minimalist races are a biological reality. Furthermore, since Rosenberg et al (2002) used microsatellite markers in their analysis, this is a nonarbitrary way of constructing genetic clusters which then demarcate the continental-level minimalist races (Hardimon, 2017b: 90).
Thus, Spencer (2014) argues to call the partition identified in K = 5 “the Blumenbachian partition” in honor of Johann Blumenbach, anthropologist, physician, physiologist, and naturalist. (Though it should be noted that one of his races “Malays” was not a race, but Oceaninans are, so he “roughly discovered” the population partition.) So we can say that “the Blumenbach partition” is just the US meaning of “race”, the partitions identified by K = 5 (Rosenberg et al, 2002).
Furthermore, like Lewontin (1972), Rosenberg et al (2002) found that a majority of human genetic variation is between individuals, not races. That is, Rosenberg et al (2002) found that only 4.3 percent of human genetic variation was found to lie between the continental-level minimalist races. Thus, minimalist races are a biological kind, “if only a modest one” (Hardimon, 2017b: 91). Thus, Rosenberg et al (2002) support the contention that minimalist races exist and are a biological reality since a fraction of human population variation is due to differences among continental-level minimalist races (Africans, Caucasians, East Asians, Oceanians, and Amerindians). The old canard is true, there really is more genetic variation within races than between them, but, as can be seen, that does not rail against the reality of race, since that small amount of genetic variation shows that humanity is meaningfully clustered in a genetic sense.
Spencer (2014: 1032) then argues why Blumenbachian populations are “race” in the American sense:
It is not hard to generate accessible possible worlds that support the claim that US race terms are just aliases for Blumenbachian populations. For example, imagine a possible world τ where human history unfolded exactly how it did in our world except that every Caucasian in τ was killed by an infectious disease in the year 2013. Presumably, we have access to τ, since it violates no logical, metaphysical, or scientific principles. Then, given that we use ‘white’ in its national American meaning in our world, and given that we use ‘Caucasian’ in its Blumenbachian meaning in our world, it is fair to say that both ‘Caucasian’ and ‘white’ are empty terms in τ in 2014—which makes perfect sense if ‘white’ is just an alias for Caucasians. It is counterfactual evidence like this that strongly suggests that the US meaning of ‘race’ is just the Blumenbach partition.
Contrary to critics, this partition is biologically real and demarcates the five genetically structured populations of the human race. Rosenberg et al (2005) found that if sufficient data are used, “the geographic distribution of the sampled individuals has little effect on the analysis“, while their results verify that genetic clusters “arise from genuine features of the underlying pattern of human genetic variation, rather than as artifacts of uneven sampling along continuous gradients of allele frequencies.”
Some may claim that K = 5 is “arbitrary”, however, constructing genetic clusters using microsatellites is nonarbitrary (Hardimon, 2017b: 90):
Constructing genetic clusters using microsatellites constitutes a nonarbitrary way of demarcating the boundaries of continental-level minimalist races. And the fact that it is possible to construct genetic clusters corresponding to continental-level minimalist races in a nonarbitrary way is itself a reason for thinking that minimalist race is biologically real 62.
It should also be noted that Hardimon writes in note 62 (2017b: 197):
Just to be perfectly clear, I don’t think that the results of the 2002 Rosenberg article bear on the question: Do minimalist races exist? That’s a question that has to be answered separately. In my view, the fundamental question in the philosophy of race on which the results of this study bear is whether minimalist race is biologically real. My contention is that they indicate that minimalist race (or more precisely, continental-level minimalist race) is biologically real if sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races.
Sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races, therefore race is a biological reality. We can pinpoint them on the basis of patterns of visible physical features; these visible physical features correspond to geographic ancestry; this satisfies the criteria for minimalist races; therefore race exists. Race exists as a biological kind.
Furthermore, if these five populations that Rosenberg et al (2002) identified (the Blumenbachian populations) are minimalist races, then minimalist race is “a minor principle of human genetic structure” (Hardimon, 2017b: 92). Since minimalist races constitute a dimension within the small amount of human genetic variation that is captured between the continental-level minimalist races (4.3 percent), then it is completely possible to talk meaningfully about the racial structure of human genetic variation which consists of the human genetic variation which corresponds to continental-level minimalist races.
Thus, the US meaning of race is just a referent; the US meaning of race refers to a particular set of human populations; races in the US are classically-defined races (Amerindian, Caucasian, African, East Asian, and Oceanians; the Blumenbach partition); and race is both a biological reality as well as socially constructed. These populations are biologically real; if these populations are biologically real, then it stands to reason that biological racial realism is true (Hardimon, 2012 2013, 2017a; 2017b; Spencer, 2014, 2015).
Human races exist, in a minimalist biological sense, and there are 5 human races. Defenders of Rushton’s work—who believed there are only 3 primary races: Caucasoids, Mongoloids, and Negroids (while Amerindians and others were thrown into the “Mongoloid race” and Pacific Islanders being grouped with the “Negroid race” (Rushton, 1988, 1997; see also Liberman, 2001 for a critique of Rushton’s tri-racial views)—are forced into a tri-racial theory, since he used this tri-racial theory as the basis for his, now defunct, r/K selection theory. The tri-racial theory, that there are three primary races of man—Caucasoid, Mongoloid, and Negroid—has fallen out of favor with anthropologists for decades. But what we can see from new findings in population genetics since the sequencing of the human genome, however, is that human populations cluster into five populations and these five populations are races, therefore biological racial realism is true.
Biological racial realism (the fact that race exists as a biological reality) is true, however, just like with Hardimon’s minimalist races, they do not denote “superiority”, “inferiority” for one race over another. Most importantly, Blumenbachian populations do not denote those terms because the genetic evidence that is used to support the Blumenbachian partition use noncoding DNA. (It should also be noted that the terms “superior” and “inferior” are nonsensical, when used outside of their anatomic contexts. The head is the most superior part of the human body, the feet are the most inferior part of the human body. This is the only time these terms make sense, thus, using the terms outside of this context makes no sense.)
It is worth noting that, while Hardimon’s and Spencer’s views on race are similar, there are some differences between their views. Spencer sees “race” as a referent, while Hardimon argues that race has a set descriptive meaning on the basis of C (1)-(3); (C1) that, as a group, is distinguished from other groups of human beings by patterns of visible physical features, (C2) whose members are linked be a common ancestry peculiar to members of that group, and (C3) that originates from a distinctive geographic location” (Hardimon, 2017b: 31). Whether or not one prefers Blumenbachian partitions or minimalist races depends on whether or not one prefers race in a descriptive sense (i.e., Hardimon’s minimalist races) or if the term race in America is a referent to the US Census discourse, which means that “race” refers to the OMB definitions which then denote Blumenbachian partitions.
Hardimon also takes minimalist races to be a biological kind, while Spencer takes them to be a proper name for a set of population groups. Both of these differing viewpoints regarding race, while similar, are different in that one is describing a kind, while the other describes a proper name for a population group; these two views regarding population genetics from these two philosophers are similar, they are talking about the same things and hold the same deflationary views regarding race. They are talking about how race is seen in everyday life and where people get their definitions of “race” from and how they then integrate it into their everyday lives.
“Race” in America is a proper name for a set of human population groups, the five population groups identified by K = 5. Americans defer to the US Census Bureau on race, who defers to the Office of Management and Budget to define race. They hold that races are a “set”, and these “sets” are Oceanians, Caucasians, East Asians, Amerindians, and Africans. Race, thusly, refers to a set of population groups; “race” is not a “kind”, but a proper name for known populational groups. K = 5 then shows us that the demarcated clusters correspond to continental-level minimalist races, what is termed “the Blumenbach partition.” This partition is “race” in the US sense of the term, and it is a biological reality, therefore, like Hardimon’s minimalist races, the Blumenbach partition identifies what we in America know to be race. (It’s worth noting that, obviously, the Blumenbach partition/minimalist races are one in the same, Spencer is a deflationary realist regarding race, just like Hardimon.)
Defending Minimalist Races: A Response to Joshua Glasgow
2000 words
Michael Hardimon published Rethinking Race: The Case for Deflationary Realism last year (Hardimon, 2017). I was awaiting some critical assessment of the book, and it seems that at the end of March, some criticism finally came. The criticism came from another philosopher, Joshua Glasgow, in the journal Mind (Glasgow, 2018). The article is pretty much just arguing against his minimalist race concept and one thing he brings up in his book, the case of a twin earth and what we would call out-and-out clones of ourselves on this twin earth. Glasgow makes some good points, but I think he is largely misguided on Hardimon’s view of race.
Hardimon (2017) is the latest defense for the existence of race—all the while denying the existence of “racialist races”—that there are differences in mores, “intelligence” etc—and taking the racialist view and “stripping it down to its barebones” and shows that race exists, in a minimal way. This is what Hardimon calls “social constructivism” in the pernicious sense—racialist races, in Hardimon’s eyes, are socially constructed in a pernicious sense, arguing that racialist races do not represent any “facts of the matter” and “supports and legalizes domination” (pg 62). The minimalist concept, on the other hand, does not “support and legalize domination”, nor does it assume that there are differences in “intelligence”, mores and other mental characters; it’s only on the basis of superficial physical features. These superficial physical features are distributed across the globe geographically and these groups are real and exist who show these superficial physical features across the globe. Thus, race, in a minimal sense, exists. However, people like Glasgow have a few things to say about that.
Glasgow (2018) begins by praising Hardimon (2017) for “dispatching racialism” in his first chapter, also claiming that “academic writings have decisively shown why racialism is a bad theory” (pg 2). Hardimon argues that to believe in race, on not need believe what the racialist concept pushes; one must only acknowledge and accept that there are:
1) differences in visible physical features which correspond to geographic ancestry; 2) these differences in visible features which correspond to geographic ancestry are exhibited between real groups; 3) these real groups that exhibit these differences in physical features which correspond to geographic ancestry satisfy the conditions of minimalist race; C) therefore race exists.
This is a simple enough argument, but Glasgow disagrees. As a counter, Glasgow brings up the “twin earth” argument. Imagine a twin earth was created. On Twin Earth, everything is exactly the same; there are copies of you, me, copies of companies, animals, history mirrored down to exact minutiae, etc. The main contention here is that Hardimon claims that ancestry is important for our conception of race. But with the twin earth argument, since everything, down to everything, is the same, then the people who live on twin earth look just like us but! do not share ancestry with us, they look like us (share patterns of visible physical features), so what race would we call them? Glasgow thusly states that “sharing ancestry is not necessary for a group to count as a race” (pg 3). But, clearly, sharing ancestry is important for our conception of race. While the thought experiment is a good one it fails since ancestry is very clearly necessary for a group to count as a race, as Hardimon has argued.
Hardimon (2017: 52) addresses this, writing:
Racial Twin Americans might share our concept of race and deny that races have different geographical origins. This is because they might fail to understand that this is a component of their race concept. If, however, their belief that races do not have different geographical origins did not reflect a misunderstanding of their “race concept,” then their “race concept” would not be the same concept as the concept that is the ordinary race concept in our world. Their use of ‘race’ would pick out a different subject matter entirely from ours.
and on page 45 writes:
Glasgow envisages Racial Twin Earth in such a way that, from an empirical (that is, human) point of view, these groups would have distinctive ancestries, even if they did not have distinctive ancestries an sich. But if this is so, the groups [Racial Twin Earthings] do not provide a good example of races that lack distinctive ancestries and so do not constitute a clear counterexample to C(2) [that members of a race are “linked by a common ancestry peculiar to members of that group”].
C(2) (P2 in the simple argument for the existence of race) is fine, and the objections from Glasgow do not show that P(C)2 is false at all. The Racial Twin Earth argument is a good one, it is sound. However, as Hardimon had already noted in his book, Glasgow’s objection to C(2) does not rebut the fact that races share peculiar ancestry unique to them.
Next, Glasgow criticizes Hardimon’s viewpoints on “Hispanics” and Brazilians. These two groups, says Glasgow, shows that two siblings with the same ancestry, though they have different skin colors, would be different races in Brazil. He uses this example to state that “This suggests that race and ancestry can be disconnected” (pg 4). He criticizes Hardimon’s solution to the problem of race and Brazilians, stating that our term “race” and the term in Brazil do not track the same things. “This is jarring. All that anthropological and sociological work done to compare Brazil with the rest of the world (including the USA) would be premised on a translation error” (pg 4). Since Americans and Brazilians, in Glasgow’s eyes, can have a serious conversation about race, this suggests to Glasgow that “our concept of race must not require that races have distinct ancestral groups” (pg 5).
I did cover Brazilians and “Hispanics” as regards the minimalist race concept. Some argue that the “color system” in Brazil is actually a “racial system” (Guimaraes 2012: 1160). While they do denote race as ‘COR’ (Brazilian for ‘color), one can argue that the term used for ‘color’ is ‘race’ and that we would have no problem discussing ‘race’ with Brazilians, since Brazilians and Americans have similar views on what ‘race’ really is. Hardimon (2017: 49) writes:
On the other hand, it is not clear that the Brazilian concept of COR is altogether independent of the phenomenon we Americans designate using ‘race.’ The color that ‘COR’ picks out is racial skin color. The well-known, widespread preference for lighter (whiter) skin in Brazil is at least arguably a racial preference. It seems likely that white skin color is preferred because of its association with the white race. This provides a reason for thinking that the minimalist concept of race may be lurking in the background of Brazilian thinking about race.
Since ‘COR’ picks out racial skin color, it can be safely argued that Brazilians and Americans at least are generally speaking about the same things. Since the color system in Brazil pretty much mirrors what we know as racial systems, demarcating races on the basis of physical features, we are, it can be argued, talking about the same (or similar) things.
Further, the fact that “Latinos” do not fit into Hardimon’s minimalist race concepts is not a problem with Hardimon’s arguments about race, but is a problem with how “Latinos” see themselves and racialize themselves as a group. “Latinos” can count as a socialrace, but they do not—can not—count as a minimalist race (such as the Caucasian minimalist race; the African minimalist race; the Asian minimalist race etc), since they do not share visible physical patterns which correspond to differences in geographic ancestry. Since they do not exhibit characters that demarcate minimalist races, they are not minimalist races. Looking at Cubans compared to, say, Mexicans (on average) is enough to buttress this point.
Glasgow then argues that there are similar problems when you make the claim “that having a distinct geographical origin is required for a group to be a race” (pg 5). He says that we can create “Twin Trump” and “Twin Clinton” might be created from “whole cloth” on two different continents, but we would still call them both “white.” Glasgow then claims that “I worry that visible trait groups are not biological objects because the lines between them are biologically arbitrary” (pg 5). He argues that we need a “dividing line”, for example, to show that skin color is an arbitrary trait to divide races. But if we look at skin color as an adaptation to the climate of the people in question (Jones et al, 2018), then this trait is not “arbitrary”, and the trait is then linked to geographic ancestry.
Glasgow then goes down the old and tired route that “There is no biological reason to mark out one line as dividing the races rather than another, simply based on visible traits” (pg 5). He then goes on to discuss the fact that Hardimon invokes Rosenberg et al (2002) who show that our genes cluster in specific geographic ancestries and that this is biological evidence for the existence of race. Glasgow brings up two objections to the demarcation of races on both physical appearance and genetic analyses: picture the color spectrum, “Now thicken the orange part, and thin out the light red and yellow parts on either side of orange. You’ve just created an orange ‘cluster’” (pg 6), while asking the question:
Does the fact that there are more bits in the orange part mean that drawing a line somewhere to create the categories orange and yellow now marks a scientifically principled line, whereas it didn’t when all three zones on the spectrum were equally sized?
I admit this is a good question, and that this objection would indeed go with the visible trait of skin color in regard to race; but as I said above, since skin color can be conceptualized as a physical adaptation to climate, then that is a good proxy for geographic ancestry, whether or not there is a “smooth variation” of skin colors as you move away from the equator or not, it is evidence that “races” have biological differences and these differences start on the biggest organ in the human body. This is just the classic continuum fallacy in action: that X and Y are two different parts of an extreme; there is no definable point where X becomes Y, therefore there is no difference between X and Y.
As for Glasgow’s other objection, he writes (pg 6):
if we find a large number of individuals in the band below 62.3 inches, and another large grouping in the band above 68.7 inches, with a thinner population in between, does that mean that we have a biological reason for adopting the categories ‘short’ and ‘tall’?
It really depends on what the average height is in regard to “adopting the categories ‘short’ and ‘tall’” (pg 6). The first question was better than the second, alas, they do not do a good job of objecting to Hardimon’s race concept.
In sum, Glasgow’s (2018) review of Hardimon’s (2017) book Rethinking Race: The Case for Deflationary Realism is an alright review; though Glasgow leaves a lot to be desired and I do think that his critique could have been more strongly argued. Minimalist races do exist and are biologically real.
I am of the opinion that what matters regarding the existence of race is not biological science, i.e., testing to see which populations have which differing allele frequencies etc; what matters is the philosophical aspects to race. The debates in the philosophical literature regarding race are extremely interesting (which I will cover in the future), and are based on racial naturalism and racial eliminativism.
(Racial naturalism “signifies the old, biological conception of race“; racial eliminativism “recommends discarding the concept of race entirely“; racial constructivism “races have come into existence and continue to exist through “human culture and human decisions” (Mallon 2007, 94)“; thin constructivism “depicts race as a grouping of humans according to ancestry and genetically insignificant, “superficial properties that are prototypically linked with race,” such as skin tone, hair color and hair texture (Mallon 2006, 534); and racial skepticism “holds that because racial naturalism is false, races of any type do not exist“.) (Also note that Spencer (2018) critiques Hardimon’s viewpoints in his book as well, which will also be covered in the future, along with the back-and-forth debate in the philosophical literature between Quayshawn Spencer (e.g., 2015) and Adam Hochman (e.g., 2014).)
“Latinos”, Brazilians, Mixed-Race Individuals and Race Concepts
2050 words
How do “Latinos”, Brazilians, and mixed-race individuals fit into Hardimon’s (2017) differing race concepts (racialist, minimalist, populationist, and socialrace)? It’s easier explaining how “Latinos” fit into this, but mixed-race individuals are a bit trickier (for instance, the minimalist concept of race does not say anything about it and is therefore vague in that respect). This article will discuss these two populations and see where they fit into these categories.
Mixed-race individuals
Mixed-race individuals are tricky to place in these conceptions of race that Hardimon (2017) lays out and defends. For example, minimalist race itself is vague; it does not say which populations/individuals with populational characteristics would be placed, the argument just establishes the biological reality and significance of race. The concept of the “one drop rule” (was a legal) is a social standard in that anyone with “one drop” of “black blood” was deemed black (which, it seems, did well for so-called conceptions of “racial purity” since most white Americans have low amounts of black ancestry; Bryc et al, 2015).
The one drop rule was an attempt to limit racial miscegenation (racial mixing), and it seems to have, for the most part, worked since many white Americans have low to no African ancestry (since 95 percent of white Americans have no African ancestry; Bryc et al, 2015).
Though, as Hardimon (2017: 49) writes, the fact that individuals must have a race is a holdover from the racialist concept; minimalist race, as I’ve covered, is not defined by the features of an individual but is defined by the features of the group said individual belongs to. It is defined in terms of group—not individual—characteristics. So just because individual I doesn’t look like their R but instead looks like an R2, for example, doesn’t make individual I an R2; individual I is still an R even though they look like an R2 since the concept is based on shared group characteristics.
Hardimon doesn’t really discuss mixed-race individuals in his book; there’s only really one note on the subject (and it’s about social race, pg. 209, note 54):
People who are members of more than one socialrace in a socialrace regime that does not recognize mixed race as a racial position will be in the anomalous situation of not having an established socialrace position in society. Having such a position is one way of being a “normal” member of society organized around the institution of socialrace. Not to have such a position is to have no place in the social world along the dimension of socialrace. Hence, perhaps, the pathos of mixed-race individuals seeking social recognition for their distinctive mixed race identity.
Though, in certain (racialist/socialrace) societies, a mixed-race individual would become what the “lower” race is in that society. For instance, if an individual were half white and half black—like the former President of the United States Barack Obama—he would be designated as “black”, as we all know (since he’s the first black President of the United States). This is known as the concept of “hypodescent” and has its basis in Hardimon’s socialrace concept since racial status of the offspring is designated to the parent who has lower “standing” than the majority in said country. So, therefore, the concept of hypodescent is the concept of socialrace in action.
Mixed-race individuals are seen as members of their “lower-status parent group“, which shows how racial constructivism is alive today. One is designated the lower-status of their parent in a hierarchical manner—one way in which the socialrace concept borrows from the racialist race concept in that it is hierarchical.
The researchers found, for example, that one-quarter-Asian individuals are consistently considered more white than one-quarter-black individuals, despite the fact that African Americans and European Americans share a substantial degree of genetic heritage. One Drop Rule Persists—Harvard
Brazil
How does this work in Brazil? Surely this makes problems for racial concepts, right?
Brazilians don’t use the term “race” (raca), but the term “color” (cor). “The reason the word Color (capitalized to call attention to this particular meaning) is preferred to race in Brazil is probably because it captures the continuous aspects of phenotypes” (Parra et al, 2003). Clearly, the conception of “race” (raca) in Brazil comes down to what “color” (cor) one is; and so we should state that Brazilian society is stratified into “colors” and not true “races”. The 1872 Brazilian census created four “color” groups “white, caboclo, black and brown (branco, caboclo (mixed indigenous-European), negro and pardo). These groups were always defined by the same formula: Colour group = members of a pure race + phenotypes of this race in the process of reversion (Guimaraes 1999)” (Guimaraes, 2012: 1157).
Though, distinct from Hardimon (2017), Guimaraes (2012: 1160) argues that the Brazilian color system is, in fact, a racial system:
What makes colour in Brazil a racial term is precisely the fact that the physiognomic traits used by racialists to distinguish different human races became convoluted with the original European system of classification based on shades of skin colour.
Hardimon (2017: 49) writes:
On the other hand, it is not clear that the Brazilian concept of COR is altogether independent of the phenomenon we Americans designate using ‘race.’ The color that ‘COR’ picks out is racial skin color. The well-known, widespread preference for lighter (whiter) skin in Brazil is at least arguably a racial preference. It seems likely that white skin color is preferred because of its association with the white race. This provides a reason for thinking that the minimalist concept of race may be lurking in the background of Brazilian thinking about race.
Someone who is “white” (identifies as white) in Brazil will (most likely) not be white in America; this is due to their differing classification system based on color, which is loosely tied with the minimalist concept, but is distinct from it in that it’s just based largely on the color of one’s skin (one of many requisites for minimalist racehood).
The role of race and color in regard to Brazilian society is complex, biologically, sociologically, and psychologically; but it’s clear that some concept of what we could call a “classical” race concept—however crude—could be said to be in use in Brazil today.
Latinos/Hispanics
The US Census Bureau states that “People who identify their origin as Hispanic, Latino, or Spanish may be of any race.” This is true; just because one derives from a Latin American country does not mean that they are some kind of “Latino” or “Hispanic” race.
Take, as an example, the case of Alberto Fujimori. Alberto Fujimori is the son of Japanese immigrants to Peru, and he eventually ended up becoming the President of Peru. His parents emigrated to Peru and he was born in Lima. Now here’s where things get tricky: is he all of a sudden some new type of “race” called “Hispanic” or “Latino”, all because he was born across the ocean? Is Pope Francis all of a sudden not Italian by ancestry since he was born in Argentina? Are people who are born in Argentina, Chile, and Paraguay with direct ancestry to Germany and Italy all of a sudden no longer German or Italian but some new “Hispanic” or “Latino” race? No! Just because you’re born not in your ancestral home does not mean you “become” whatever society designates that part of the world (in this case “Latin America”). Their race does not change on the basis of where they are born; their individual ancestries can still be traced back to their countries of origin, therefore attempting to “racialize” the terms “Hispanic” or “Latino” do not make any sense.
“Hispanics” do not—and cannot—count as a minimalist race on the basis of one condition: they do not share a single pattern of visible physical features. No one pattern of hair, skin color, lip shape, eye shape etc. They do not share a single geographic origin; they have a mixture of Ancestry from Africa, Europe, and Asia. “Hispanics” can be seen, obviously, as a mixture of minimalist races. “Hispanics” are denied minimalist racehood since they do not exhibit characteristics of minimalist races, which is even echoed, as shown above, by the US Census Bureau.
Hardimon (2017: 39) writes:
To deny that Latinos constitute a race is not to deny that individual Latinos or Latinos as a group can be the targets of racism (for example, owing to skin color). Nor is it to deny that Latinos are often regarded as “racially other” (as differing in some essential humanly important way corresponding to skin color) by members of other racialized groups (for example, Anglos). … Nor is it to deny that they constitute a socialrace in my sense of the term. Still less does it imply that Latinos ought not to aspire to a degree of solidarity connoted by the Spanish word raza.
So “Latinos” can be designated as a socialrace (though socialraces do not always have a mirrored minimalist race), but not as a minimalist race since they do not fit the criteria for minimalist racehood. Many “Latinos” can be said to be mestizos, which are half European (normally of Spanish descent) and half Indian. Still, further, they can be castizos, about three-quarters European and one-quarter Indian. Then you have the “Latino” Carribean countries (Dominican Republic, Cuba, Puerto Rico) with differing amounts of admixture from all over, from different minimalist races. Though, in America, most Dominicans would be counted as “black” under the concept of “hypodescent”—the one drop rule. Many Cubans can be seen to have majority Spanish ancestry, and its the same for Puerto Ricans.
“Latinos/Hispanics” do not constitute a major race (in the minimalist and populationist sense) because they are a mixture of different minimalist races. This does not mean, however, that one designated as “Latino” in America does not have full ancestry to a European country; this is how the concept of “Latino” in America uses the concept of socialrace—it’s only based on the perceived race of the individual (that they derive from a “Latino” country) and that therefore makes them “Latino”, all the while ignoring their actual racial ancestry.
There is even a phrase in Latin America “Mejorar la raza” or “improve the race” by having children with lighter-skinned people since light skin is seen as beautiful in Latin America. They want to better “their race” (even though they—as “Latinos”—don’t have a race), and so they will attempt to have children with lighter and lighter people (i.e., people who have more and more European ancestry) to “improve their race” (i.e., their socialrace) since European features are seen as more beautiful in Latin America.
Most of the ruling class in Latin America is of European descent, while the lower classes are of admixed/unadmixed Indians (coming from differing tribes). This, one can say, is one way that socialrace is used in Latin America.
Conclusion
Brazilians and “Latinos/Hispanics” clearly could have been grouped in the mixed-race category, but each of these subjects has distinct concepts it needed to discuss. The Brazilian concept of “cor” and “raca” are loosely intertwined; they can be said to use aspects of mimimalist races. On the other hand, “Latinos/Hispanics” are not designated minimalist racehood on the basis that they do not share a single pattern of physical features, nor do they share a geographic origin, since the groups that make up “Latinos” (which are minimalist races) are the geographic locations in question. “Latinos/Hispanics” are not minimalist races because they do not exhibit the features of minimalist races.
Mixed-race individuals, regarding the socialrace concept, can be seen to be the “lower” of the races they are admixed with on the social ladder—which is how it is in America (the concept of hypodescent). The existence of mixed-race individuals does not invalidate the concept of race; minimalist racehood is not defined on the basis of individual characters, but on the basis of the characters of the group. Therefore this does not go against the concept of minimalist race.
The concepts of race can definitely survive these anomalies when describing the biological realities of race; some of them can be said to be socialraces for one respect, whereas in reality, they are mixtures of minimalist races. Races exist and the existence of Brazilians (even with their own categorization of races/colors) and “Latinos/Hispanics” and other mixed-race groups/individuals do not rail against any concepts that purport to argue for the existence and biological reality of race.
A Simple Argument for the Existence of Race
1550 words
Race deniers say that there is too small of a genetic distance between races to call the so-called races “races”. They latch on to Lewontin’s 1974 analysis, trumpeting that genetic distance is too small for there to be true “races”. There is, however, a simple way to bypass the useless discussions that would ensue if one cites genetic evidence for the existence of race: just use this simple argument:
P1) There are differences in patterns of visible physical features which correspond to geographic ancestry
P2) These patterns are exhibited between real groups, existing groups (i.e., individuals who share common ancestry)
P3) These real, existing groups that exhibit these physical patterns by geographic ancestry satisfy conditions of minimalist race
C) Therefore race exists and is a biological reality
This argument is simple; anyone who denies this needs to provide a good enough counter-argument, and I’m not aware of any that exist to counter the argument.
P1 shows that there are patterns of visible physical features which correspond to geographic ancestry. This is due to the climates said race evolved in over evolutionary history. Since these phenotypes are not randomly distributed across the globe, but show distinct patterning based on geographic ancestry, we can say that P1 is true; different populations show patterns of different physical features which are not randomly distributed across the globe. Further, since P1 establishes that races are populations that look different from each other, it guarantees that groups like the Amish, social classes etc are not counted as races. P1 further allows a member of a given race to not show the normative physical characters that are characteristic of that race. It further allows for the possibility that individuals from two different races may not differ in their physical characters. These visible physical characters that differ by populations we then call races also need to be heritable to be biological. “Because the visible physical features of race are heritable, the skin color, hair type, and eye shape of children of Rs tend to resemble the skin color, hair type, and eye shape of their parents” (Hardimon, 2017: 35). P1 is true.
P2 shows that these patterns of visible physical features are exhibited between real, existing groups. That is, the groups that exhibit these patterns exist in reality. No one denies this either. Differences in physical features that these real, existing groups exhibit can then be used as proxies for factors in P1. Though, like with which populations figure into this concept, the minimalist race concept doesn’t say—it only establishes the biological existence of races. “In recent years the concept of the continent has come under fire for not being well defined. 59 It is of interest that the formation of the concepts CONTINENT and RACE are roughly coeval. One wonders if the geneses of the two ideas are mutually entwined. Could it be that our idea of continent derives in part from the idea of the habitat of a racial group? Could it be that the idea of a racial group gets part of its content from the idea of a group whose aboriginal home is a distinctive continent? Perhaps the concepts should be thought of as having formed in tandem, each helping to fix the other’s reference” (Hardimon, 2017: 51). Since these real, existing populations that were geographically separated for thousands of years show these visible physical patterns, P2 is true.
P3 follows from the specification of the concept of minimalist race. If these populations that exhibit these distinct visible characters and if they are non-randomly distributed across the globe then this satisfied the argument for the concept of minimalist race. The specification of the minimalist concept of race states that groups satisfy the requisites for the concept by being distinguishable by patterns of visible physical features (P1) and that individuals who share a common ancestry peculiar to them which derive from a distinct geographic location (P2) exist as real groups. Since P1 and P2 are true, P3 follows logically from P1 and P2, which then leads us to the conclusion which is true and establishes the argument for the minimalist concept of race as a sound and valid argument.
C is then the logical conclusion of the three premises: race exists and is a biological reality since the patterns of visible physical features are non-randomly distributed across the globe and are exhibited by real, existing groups. Since all three premises are true and the conclusion is true, it is a valid argument; since the premises are true the argument is sound. No one can—logically—deny the existence of race when presented with this logical proof.
Though notice the argument doesn’t identify which populations are designated as “races” (that’s for another article), the argument just establishes that race exists and it exists as a biological reality. Notice also how this conception of race is sort of like the “racialist concept”, but it takes it down to its barest bones—only taking the normatively important, superficial biological physical features (these features establish minimalist races as biologically existing).
Notice, too, that I did not appeal to any genetic differences between the races, indeed, in my opinion, they are not needed when discussing race. All that is needed when discussing race and whether or not it is a biological reality is asking three simple questions:
1) Are there differences in patterns of visible physical features that correspond to geographic ancestry? Yes.
2) Are these patterns exhibited by real, existing groups? Yes.
3) Do these real, existing groups satisfy conditions of minimalist race? Yes
Therefore race exists.
These three simple questions (just take the premises and ask them as questions) will have one—knowingly or not—admit to the biological reality and existence of race.
Do note, though, nothing in this argument brings up anything about what we “can’t see”, meaning things like “intelligence” or mores of these races. This concept is distinct from the racialist concept in that it does not mention normatively important characters; it does not posit a relationship between visible physical characters and normatively important characters; and it does not “rank” populations on some type of scale. “Also, the conjoint fact that a group is characterized by a distinctive pattern of visible physical characteristics and consists of members who are linked by a common ancestry and originates from a distinctive geographic location is of no intrinsic normative significance. The status of being a minimalist race has no intrinsic normative significance“ (Hardimon, 2017: 32).
Clearly, one does not need to invoke genetic differences to show that race exists as a biological reality. That races differ in patterns of visible physical features which are inherited from the parents and are heritable establishes the biological reality of minimalist race. I really see no way that one could, logically, deny the existence of race given the argument provided in this article. Race exists and is a biological reality and the argument for the existence of minimalist race establishes this fact. Races differ in physiognomy and morphology; these physical differences are non-randomly distributed by geographic ancestry/at the continental level. These populations that show these physical differences share a peculiar ancestry. Knowing these facts, we can safely infer the existence of race. It is the only logical conclusion to come to. Note that the minimalist concept is deflationist—meaning that racialist races do not exist and that this concept enjoys what the racialist concept was supposed to, it is deflationary in the aspect that it takes the normative physical differences from the racialist concept. It is realist since it acknowledges the existence of minimalist race as genetically grounded and relatively superficial but still very significant biological reality of race.
Races can exist as minimalist races and socialraces—no contradiction exists. minimalist race, and its “scientific” companion populationist race (which I will cover in the future) show that there is a well-formulated argument for the existence of race (minimalist race concept) whereas the other concept shows how it is grounded in science and partitions populations to races (populationist concept; both are deflationary). (Read the descriptions of racialist race, minimalist race, populationist race, and socialrace.) You don’t need genes to delineate race; you only need a sound, valid argument based on biological principles. Minimalist races exist.
Race exists and is a biological reality, even if it is ‘socially constructed’ (what isn’t?), our social constructs still correspond to differing breeding populations who share peculiar ancestry and show patterns of visible physical features establish the existence of race.

From Hardimon (2017: 177)
(I also came across a book review from philosopher Joshua Glasgow (Book Review Rethinking Race: The Case for Deflationary Realism, by Michael O. Hardimon. Harvard University Press, 2017. Pp. 240.), author of A Theory of Race (2009) who has some pretty good critiques against Hardimon’s theses in his book, but not good enough. I am going to cover a bit more about these concepts then discuss his article. I will also cover “Latinos” and mixed race people as regards these concepts as well.)
4/19/2018 edit: Two more simple arguments:
(Where P is population, C is continent and T is trait(s)
Population P that evolved in continent C have physical traits T which correspond to C.
Minimalist Races Exist and are Biologically Real
3050 words
People look different depending on where their ancestors derived from; this is not a controversial statement, and any reasonable person would agree with that assertion. Though what most don’t realize, is that even if you assert that biological races do not exist, but allow for patterns of distinct visible physical features between human populations that then correspond with geographic ancestry, then race—as a biological reality—exists because what denotes the physical characters are biological in nature, and the geographic ancestry corresponds to physical differences between continental groups. These populations, then, can be shown to be real in genetic analyses, and that they correspond to traditional racial groups. So we can then say that Eurasian, East Asian, Oceanian, black African, and East Asians are continental-level minimalist races since they hold all of the criteria needed to be called minimalist races: (1) distinct facial characters; (2) distinct morphologic differences; and (3) they come from a unique geographic location. Therefore minimalist races exist and are a biological reality. (Note: There is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016), but this does not mean that the minimalist biological concept of race has no grounding in biology.)
Minimalist race exists
The concept of minimalist race is simple: people share a peculiar geographic ancestry unique to them, they have peculiar physiognomy (facial features like lips, facial structure, eyes, nose etc), other physical traits (hair/hair color), and a peculiar morphology. Minimalist races exist, and are biologically real since minimalist races can survive findings from population genetics. Hardimon (2017) asks, “Is the minimalist concept of race a social concept?” on page 62. He writes that social concepts are socially constructed in a pernicious sense if and only if it “(i) fails to represent any fact of the matter and (ii) supports and legitimizes domination.” Of course, populations who derive from Africa, Europe, and East Asia have peculiar facial morphology/morphology unique to that isolated population. Therefore we can say that minimalist race does not conform to criteria (i). Hardimon (2017: 63) then writes:
Because it lacks the nasty features that make the racialist concept of race well suited to support and legalize domination, the minimalist race concept fails to satisfy condition (ii). The racialist concept, on the other hand, is socially constructed in the pernicious sense. Since there are no racialist races, there are no facts of the matter it represents. So it satisfies (i). To elaborate, the racialist race concept legtizamizes racial domination by representing the social hierarchy of race as “natural” (in a value-conferring sense): as the “natural” (socially unmediated and inevitable) expression of the talent and efforts of the inidividuals who stand on its rungs. It supports racial domination by conveying the idea that no alternative arrangment of social institutions could possibly result in racial equality and hence that attempts to engage in collective action in the hopes of ending the social hierarchy of race are futile. For these reasons the racialist race concept is also idealogical in the prejorative sense.
Knowing what we know about minimalist races (they have distinct physiognomy, distinct morphology and geographic ancestry unique to that population), we can say that this is a biological phenomenon, since what makes minimalist races distinct from one another (skin color, hair color etc) are based on biological factors. We can say that brown skin, kinky hair and full lips, with sub-Saharan African ancestry, is African, while pale/light skin, straight/wavy/curly hair with thin lips, a narrow nose, and European ancestry makes the individual European.
These physical features between the races correspond to differences in geographic ancestry, and since they differ between the races on average, they are biological in nature and therefore it can be said that race is a biological phenomenon. Skin color, nose shape, hair type, morphology etc are all biological. So knowing that there is a biological basis to these physical differences between populations, we can say that minimalist races are biological, therefore we can use the term minimalist biological phenomenon of race, and it exists because there are differences in the patterns of visible physical features between human populations that correspond to geographic ancestry.
Hardimon then talks about how eliminativist philosophers and others don’t deny that above premises above the minimalist biological phenomenon of race, but they allow these to exist. Hardimon (2017: 68-69) then quotes a few prominent people who profess that there are, of course, differences in physical features between human populations:
… Lewontin … who denies that biological races exist, freely grants that “peoples who have occupied major geographic areas for much of the recent past look different from one another. Sub-Saharan Africans have dark skin and people who have lived in East Asia tend to have a light tan skin and an eye color and eye shape that is difference from Europeans.” Similarly, population geneticist Marcus W. Feldman (final author of Rosenberg et al., “Genetic Stucture of Human Populations” [2002]), who also denies the existence of biological races, acknowledges that “it has been known for centuries that certain physical features of humans are concentrated within families: hair, eye, and skin color, height, inability to digest milk, curliness of hair, and so on. These phenotypes also show obvious variation among people from different continents. Indeed, skin color, facial shape, and hair are examples of phenotypes whose variation among populations from different regions is noticeable.” In the same vein, eliminative anthropologist C. Loring Brace concedes, “It is perfectly true that long term residents of various parts of the world have patterns of features that we can identify as characteristic of they area from which they come.”
So even these people who claim to not believe in “biological races”, do indeed believe in biological races because what they are describing is biological in nature and they, of course, do not deny that people look different while their ancestors came from different places so therefore they believe in biological races. We can then use the minimalist biological phenomenon of race to get to the existence of minimalist races.
Hardimon (2017: 69) writes:
Step 1. Recognize that there are differences in patterns of visible physical features of human beings that correspond to their differences in geographic ancestry.
Step 2. Observe that these patterns are exhibited by groups (that is, real existing groups).
Step 3. Note that the groups that exhibit these patterns of visible physical features correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race.
Step 4. Infer that minimalist race exists.
Those individuals mentioned previously who deny biological races but allow that people with ancestors from differing geographic locales look differently do not disagree with step 1, nor does anyone really disagree with step 2. Step 4’s inference immediately flows from the premise in step 3. “Groups that exhibit patterns or visible physical features that correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race. Call (1)-(4) the argument from the minimalist biological phenomenon of race” (Hardimon, 2017: 70). Of course, the argument does not identify which populations may be called races (see further below), it just shows that race is a biological reality. Because if minimalist races exist, then races exist because minimalist races are races. Minimalist races exist, therefore biological races exist. Of course, no one doubts that people come from Europe, sub-Saharan Africa, East Asia, the Americas, and the Pacific Islands, even though the boundaries between them are ‘blurry’. They exhibit patterns of visible physical characters that correspond to their differing geographic ancestry, they are minimalist races therefore minimalist races exist.
Pretty much, the minimalist concept of race is just laying out what everyone knows and arguing for its existence. Minimalist races exist, but are they biologically real?
Minimalist races are biologically real
Of course, some who would assert that minimalist races do not exist would say that there are no ‘genes’ that are exclusive to one certain population—call them ‘race genes’. Of course, these types of genes do not exist. Whether or not one individual is a part of one race or not does not rest on the basis of his physical characters, but is determined by who his parents are, because one of the three premises for the minimalist race argument is ‘must have a peculiar geographic ancestry’. So it’s not that members of races share sets of genes that other races do not, it’s based on the fact that they share a distinctive set of visible physical features that then correspond with geographic ancestry. So of course if the minimalist concept of race is a biological concept then it entails more than ‘genes for’ races.
Of course, there is a biological significance to the existence of minimalist biological races. Consider that one of the physical characters that differ between populations is skin color. Skin color is controlled by genes (about half a dozen within and a dozen between populations). Lack of UV rays for individuals with dark skin will lead to diseases like prostate cancer, while darker skin is a protectant against UV damage to human skin (Brenner and Hearing, 2008; Jablonksi and Chaplin, 2010). Since minimalist race is biologically significant and minimalist races are partly defined by differences in skin color between populations then skin color has both medical and ecological significance.
(1) Consider light skin. People with light skin are more susceptible to skin cancer since they evolved in locations with poor UVR radiation (D’Orazio et al, 2013). The body needs vitamin D to absorb and use calcium for maintaining proper cell functioning. People who evolved near the equator don’t have to worry about this because the doses of UVB they absorb are sufficient for the production of enough previtamin D. While East Asians and Europeans on the other hand, became adapted to low-sunlight locations and therefore over time evolved lighter skin. This loss of pigmentation allowed for better UVB absorption in these new environments. (Also read my article on the evolution of human skin variation and also how skin color is not a ‘tell’ of aggression in humans.)
(2) While darker-skinned people have a lower rate of skin cancer “primarily a result of photo-protection provided by increased epidermal melanin, which filters twice as much ultraviolet (UV) radiation as does that in the epidermis of Caucasians” (Bradford, 2009). Dark skin is thought to have evolved to protect against skin cancer (Greaves, 2014a) but this has been contested (Jablonski and Chaplin, 2014) and defended (Greaves, 2014b). So therefore, using (1) and (2), skin color has evolutionary signifigance.
So as humans began becoming physically adapted to their new niches they found themselves in, they developed new features distinct from the location they previously came from to better cope with the new lifestyle due to their new environments. For instance “Northern Europeans tend to have light skin because they belong to a morphologically marked ancestral group—a minimalist race—that was subject to one set of environmental conditions (low UVR) in Europe” (Hardimon, 2017: 81). Of course explaining how human beings survived in new locations falls into the realm of biology, while minimalist races can explain why this happened.
Minimalist races clearly exist since minimalist races constitute complex biological patterns between populations. Hardimon (2017: 83) writes:
It [minimalist race] also enjoys intrinsic scientific interest because it represents distinctive salient systematic dimension of human biological diversity. To clarify: Minimalist race counts as (i) salient because human differences of color and shape are striking. Racial differences in color and shape are (ii) systematic in that they correspond to differences in geographic ancestry. They are not random. Racial differences are (iii) distinctive in that they are different from the sort of biological differences associated with the other two salient systematic dimensions of human diversity: sex and age.
[…]
An additional consideration: Like sex and age, minimalist race constitutes one member of what might be called “the triumverate of human biodiversity.” An account of human biodiversity that failed to include any one of these three elements would be obviously incomplete. Minimalist race’s claim to be biologically real is as good as the claim of the other members of the triumverate. Sex is biologically real. Age is biologically real. Minimalist race is biologically real.
Real does not mean deep. Compared to the biological associated with sex (sex as contrasted with gender), the biological differences associated with minimalist race are superficial.
Of course, the five ‘clusters’ and ‘populations’ identified by Rosenberg et al’s (2002) K=5 graph, which told structure to produce 5 genetic clusters, corresponds to Eurasia, Africa, East Asia, Oceania, and the Americas, are great candidates for minimalist biological races since they correspond to geographic locations, and even corroborates what Fredrich Blumenbach said about human races back in the 17th century. Hardimon further writes (pg 85-86):
If the five populations corresponding to the major areas are continental-level minimalist races, the clusters represent continental-level minimalist races: The cluster in the mostly orange segment represents the sub-Saharan African continental-level minimalist race. The cluster in the mostly blue segment represents the Eurasian continental-level minimal race. The cluster in the mostly pink segment represents the East Asian continental-level minimalist race. The cluster in the mostly green segment represents the Pacific Islander continental-level minimalist race. And the cluster in the mostly purple segment represents the American continental-level minimalist race.
[…]
The assumption that the five populations are continental-level minimalist races entitles us to interpret structure as having the capacity to assign individuals to continental-level minimalist races on the basis of markers that track ancestry. In constructing clusters corresponding to the five continental-level minimalist races on the basis of objective, race-neutral genetic markers, structure essentially “reconstructs” those races on the basis of a race-blind procedure. Modulo our assumption, the article shows that it is possible to assign individuals to continental-level races without knowing anything about the race or ancestry of the individuals from whose genotypes the microsattelites are drawn. The populations studied were “defined by geography, language, and culture,” not skin color or “race.”
Of course, as critics note, the researchers predetermine how many populations that structure demarcates, for instance, K=5 indicates that the researchers told the program to delineate 5 clusters. Though, these objections do not matter. For the 5 populations that come out in K=5 “are genetically structured … which is to say, meaningfully demarcated solely on the basis of genetic markers” (Hardimon, 2017: 88). K=6 brings one more population, the Kalash, a group from northern Pakistan who speak an Indo-European language. Though “The fact that structure represents a population as genetically distinct does not entail that the population is a race. Nor is the idea that populations corresponding to the five major geographic areas are minimalist races undercut by the fact that structure picks out the Kalash as a genetically distinct group. Like the K=5 graph, the K=6 graph shows that modulo our assumption, continental-level races are genetically structured” (Hardimon, 2017: 88).
Though of course there are naysayers. Svante Paabo and David Serre, Hardimon writes, state that when individuals are sampled from homogeneous populations from around the world, the gradients of the allele frequencies that are found are distributed randomly across the world rather than clustering discretely. Though Rosenberg et al responded by verifying that the clusters they found are not artifacts of sampling as Paabo and Serre imply, but reflect features of underlying human variation. Though Rosenberg et al agree with Paabo and Serre in that that human genetic diversity consists of clines in variation in allele frequencies (Hardimon, 2017: 89). Other naysayers also state that all Rosenberg et al show is what we can “see with our eyes”. Though a computer does not partition individuals into different populations based on something that can be done with eyes, it’s based on an algorithm.
Hardimon also accepts that black Africans, Caucasians, East Asians, American Indians and Oceanians can be said to be races in the basic sense because “they constitute a partition of the human species“, and that they are distinguishable “at the level of the gene” (Hardimon, 2017: 93). And of course, K=5 shows that the 5 races are genetically distinguishable.
Hardimon finally discusses some medical significance for minimalist races. He states that if you are Caucasian that it is more likely that you have a polymorphism that protects against HIV compared to a member of another race. Meanwhile, East Asians are more likely to carry alleles that make them more susceptible to Steven-Johnson syndrome or another syndrome where their skin falls off. Though of course, the instances where this would matter in a biomedical context are rare, but still should be at the back of everyone’s mind (as I have argued), even though instances where medical differences between minimalist races are rare, there are times where one’s race can be medically significant.
Hardimon finally states that this type of “metaphysics of biological race” can be called “deflationary realism.” Deflationary because it “consists in the repudiation of the ideas that racialist races exist and that race enjoys the kind of biological reality that racialist race was supposed to have” and realism which “consists in its acknowledgement of the existence of minimalist races and the genetically grounded, relatively superficial, but still significant biological reality of minimalist race” (Hardimon, 2017: 95-96).
Conclusion
Minimalist races exist. Minimalist races are a biological reality because distinct visible patterns show differences between geographically isolated populations. This is enough for the classification of the five classic races we know of to be called race, be biologically real, and have a medical significance—however small—because certain biological/physical traits are tied to different geographic populations—minimalist races.
Hardimon (2017: 97) shows an alternative to racialism:
Deflationary realism provides a worked-out alternative to racialism—it it a theory that represents race as a genetically grounded, relatively superficial biological reality that is not normatively important in itself. Deflationary realism makes it possible to rethink race. It offers the promise of freeing ourselves, if only imperfectly, from the racialist background conception of race.
It is clear that minimalist races exist and are biologically real. You do not need to speak about supposed mental traits between these minimalist races, they are irrelevant to the existence of these minimalist biological races. As Hardimon (2017: 67) writes: “No reference is made to normatively important features such as intelligence, sexuality, or morality. No reference is made to essences. The idea of sharp boundaries between patterns of visible physical features or corresponding geographical regions is not invoked. Nor again is reference made to the idea of significant genetic differences. No reference is made to groups that exhibit patterns of visible physical features that correspond to geographic ancestry.”
The minimalist biological concept of race stands up to numerous lines of argumentation, therefore we can say without a shadow of a doubt that minimalist biological race exists and is real.
