
r/K Selection Theory: A Response to Anonymous Conservative
2800 words
I knew the article about r/K selection would stir a bit of debate. Anonymous Conservative has replied to both articles that were published the other day. However, he seems confused. He doesn’t talk about r/K selection theory in terms of density-dependence/independence. That’s what r/K theory was based on before it was discredited for age-specific mortality (Reznick et al, 2002). The theory was discredited decades ago. This article will be a response to him. How can you use age-specific mortality for your theory?
Combining all African and all European populations probably dulls the degree to which certain populations are r and K.
Combining the ethnies of all three populations makes no sense if you’re attempting to infer how behavior X evolved in ecosystem Y using r/K selection theory. To conduct such a study, you would need to study the races in the ecosystem that the selection was hypothesized to have occurred. r/K selection is—as I’ve already brought up—proven false. I will get to that below.
If r/K selection did apply to humans, then since Africans have been in their habitat—according to Rushton—for 140ky and Mongoloids have been in their habitat for 40ky, then Africans would have had more opportunity to approach the environmental carrying capacity while Mongoloids who migrated into novel environments (cold weather, as mentioned above) would experience r-selected traits since they are in a novel environment (r pressure) and facing cold weather (another r pressure). Per Rushton’s own arguments—along with how r/K theory was really used—Africans are K and Mongoloids are r.
Take the most r populations in Africa and you would also see highly obvious differences deviating from normal human behavior.
Which populations in Africa are ‘the most r’? What is ‘normal human behavior’?
Goal number one should be to get people forced to acknowledge that some humans are exhibiting the r-strategy compared to others.
If this were the case, then Mongoloids would be r while Africans would be K—if r/K selection theory weren’t discredited and if human races qualified as local populations. This, of course, comes from Rushton own words, who asserts that Mongoloids have cold-weather adaptations. So if Mongoloids have cold-weather adaptations and cold weather is an agent of r-selection as described previously, then Mongoloids are r-selected. This argument comes straight from Rushton’s own theory. Furthermore, Africans would be K-selected since endemic disease is an agent of K-selection. This is simple enough to understand, especially if you read a few papers on r/K selection.
I get the impression the author is a pot-stirrer ginning up debate, which I can respect. But I would counter that I think this argument requires a slightly more complex view on a few points, and it seeks to cite the established literature on r/K a little too much.
Citing papers is what’s needed when discussing scientific matters. If your arguments are not backed by scientific papers then your argument is pretty much moot.
Most of the literature on r/K is incredibly shallow in its analyses. I suspect nobody really cared about the theory on an emotional level, so nobody really bothered to look too closely at it, or tried to understand why some arguments would seemingly violate simple common sense. One person would assert things that would make no sense in certain contexts, and nobody would ever try to highlight the complexity required for a fuller understanding of the issue. It is either that, or the more powerful minds gravitated somewhere else in the sciences with more practical application.
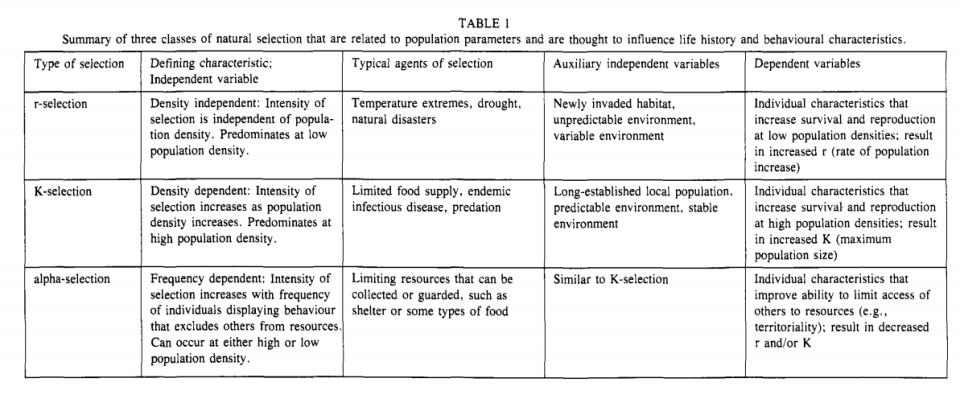
This looks pretty clear-cut to me. r/K selection theory has been extensively tested and falsified. Of course people cared about it, it dominated biology and ecology literature for about twenty years after Pianka’s (1970) paper where he proposed his now debunked ‘r/K continuum’. As I have said, Pianka gave no experimental rationale on why he chose the traits he did for the continuum (Graves, 2002: 135). This is simple enough to understand on its own.
As an example, the author cites papers that say drought is an r-selective pressure. Drought can be r or K, depending on the abilities of the organisms confronted with it. Mice will die in a drought, and have short enough life cycles to reproduce in the wet periods following it. So with mice, after the drought, there will be free resources and that makes drought a huge r-selection pressure.
But suppose you have an organism with the intelligence to envision how to survive the drought, and which thinks in terms of long time frames. Now that drought will cull the relatively r-selected individuals who are designed to exploit a glut with no thought of the future, while favoring those who planned for the drought and stockpiled water, or organized a way to acquire it. Is the drought still an r-selective pressure? Being human, with a high IQ and an ability to plan for the future changes a lot of these rules.
Drought is an agent of r-selection. How about earthquakes and volcanic eruptions? Are those agents of K-selection as well if you can ‘plan for the future changes’? Provide references for your assertion or your claim is unfounded.
On the issue of colder climates being K, the author cites research which makes the case that cold climates kill back the population in the winter, and then allow explosive growth in the summer, and thus are r-selecting.
This will be true in things like insects with short lifespans and no ability to plan for the winter. But in humans, this will favor those who can defer pleasures in the summer, looking forward to the winter and sacrificing by setting aside resources to get themselves through the colder period. It will also favor groups which can work together in pursuit of common goals.
You don’t get it. Mongoloids being r-selected is straight from Rushton. He asserts that they have cold-adaptations. Cold adaptations are due to cold weather. Cold weather is an agent of r-selection (temperature extreme). If cold weather is an agent of r-selection and Mongoloids further migrated into a novel environment (another agent of r-selection), then, per Rushton’s own words, Mongoloids are r-selected. Conversely, Rushton describes endemic disease and drought in Africa (without references), but let’s assume it’s true. As described above, drought is an agent of r (see the table from Anderson above) while endemic disease is an agent of K-selection.
Endemic (native) disease is an agent of K-selection. Since the disease is constant, then the population under that agent of K-selection can prepare ahead for disease. Indeed, in Africa, measures can be taken to reduce the number of those infected with malaria, such as mothers shielding their babies from mosquitoes, to even herbal remedies which have been in use for thousands of years (Wilcox and Bodecker, 2004). If endemic disease is constant (and it is) and Africans are under that constant pressure, then they will be K-selected.
Do groups not work together in Africa to reach common goals? In the Pleistocene as well? Citations? Think before you write (and cite), because hunting bands in our species began with Homo erectus. The capacity for endurance running evolved in erectus which can be seen with the beginnings of our modern pelvis as well as the evolution of the gluteus maximus (Lieberman et al, 2006). So how can you assert that working together to reach common goals only occurred where it was cold—as if tropical environments don’t have their own challenges which require foresight and planning? Think about human evolution and how modern human cognition evolved in Africa.
This will be true of most hardships to some degree. Where they kill back the population massively and randomly, and then allow explosive regrowth, they are r-pressures. But where they are challenges that select for those who can prepare and overcome them, they will tend to favor K, even if they may, strictly by the numbers, appear to be r.
How can you prepare and overcome a violent winter storm, volcanic eruption, earthquake, and drought (which vary wildly)? At a certain point, you can be the smartest one around but one would still succumb to the elements.
He also speaks of aggression. There the question is, is aggression borne of a competitive psychology that embraces risk innately because it evolved to embrace risk in a competitive environment where resources are scarce, or is aggression an opportunistic seizure of free resources from the weak and helpless.
A criminal who sees an old lady and pushes her to the ground to steal her purse is not the same as a Marine who proceeds to selflessly storm enemy lines and kill fifteen men with his bare hands simply to try and save his fellow Marines in battle. The criminal will seek out the weak and vulnerable to victimize safely for personal gain, while the Marine would find that in conflict with his nature. The Marine will sacrifice himself for his group and nothing more, while the criminal would view that as pointless and stupid. Those are two vastly different forms of aggression.
Aggression and violence can be principled and daring, or opportunistic and cowardly. Each is driven by a different psychology, and you can see this difference extend to sexual drive, promiscuity, and even rearing investments. I think there needs to be a difference cited there. One aggressive psychology is r and one is K. One is designed to take free resources in a world with no consequences, while the other is programmed to fight with anyone to try and get a share of scarce resources, because if they didn’t they would starve.
I speak of aggression in regards to testosterone and Richard Lynn’s claims that gonadotropin levels and testosterone lend further support for Rushton’s theory. However, I’ve falsified Ross et al (1986) numerous times. Further, the correlation between testosterone and physical aggression is a pitiful .08 (Archer, Graham-Kevan, and Lowe 2005). The point is that testosterone is not related to aggression, nor crime. Furthermore, the time of day that crime is committed at the highest rates for teens (3 pm) and adults (10 pm) discredit the testosterone-causing-crime theory since testosterone levels are highest at 8 am and lower at 8 pm. You did not address my arguments on testosterone—try again.
Then there is disease. Disease can be r or K, depending on epidemiology. If a disease is sexually transmitted, it is going to take out those with a high sex drive, promiscuity, and reduced disgust. That doesn’t means the disease is K-selecting, so much as it preferentially kills those with an r-selected psychology, and fosters the rise of K.
What about if a disease is endemic? Endemic disease (Rushton’s assertion) is an agent of K, this is not up for discussion. Endemic disease reduces carrying capacity and thusly is an agent of K-selection.
This is simple enough to understand, especially if you understand r/K selection theory.
On the other hand, if a disease infects and kills randomly, such as one transmitted by mosquito, then it will open up free resources by killing the population back below the carrying capacity. That will favor the rise of the r-selected psychologies.
Nope.
I have found the vast majority are written by individuals looking to create quick rules of thumb for much more complex variables that can only be looked at in the context of the mechanisms they are a part of. In many cases, I see authors claiming something is always r or K, when the truth is they are more often the opposite for reasons which the authors seem strangely blind to.
The vast majority of what was written about r/K in its heyday was written by biologists and ecologists. Why reduce a complex biological system interacting with its almost equally complex environment down to a discredited theory? It doesn’t make sense to reduce what organisms do to some ‘simple model’ when the real world—and by proxy ecological theories—are much more complex than a ‘simple model’.
r and K are simple adaptation to either free or limited resource availabilities. To understand how the environment affects the evolution of r and K psychologies, you have to understand that those adaptations to free or limited resources imbue certain psychological predispositions. Once imbued, all other selective pressures have to be examined with an eye to how they either confer advantage or disadvantage on those who express those psychological traits.
r/K selection theory is based on density-dependence and density-independence. As a matter of fact, searching for ‘density-dependent‘ brings up no hits and for ‘density-independent‘, the only hit is for your response to my article. Which makes me believe that you don’t understand r/K selection theory since it’s based on density-dependence and density-independence. It’s also impossible to predict which life history traits will be favored by selection unless you know which particular ecological factors influence life history traits as well as needing a model as to how they function (Anderson, 1991). Rushton did neither, and so he was wrong with his application of r/K to human races.
A sexually transmitted disease that savages a population will open up resource availability and reduce the population well below the carrying capacity, and thus could be mistaken for an r-selecting pressure. But if it wipes out every promiscuous r-strategist, and leaves behind only the monogamous K-strategists, then it is not an r-selective pressure at all. It is favoring the K-psychology, even as from a raw numerical standpoint it would appear an r-pressure.
Which STD? Which population(s)? Source? Even then, STDs such as chancroid (in the US and Europe) were endemic in the early 20th century (Aral, Fenton, and Holmes, 2007). Which populations are you describing? An event like that would be part of the density-dependence aspect of what r/K described. The population would dip and then go right back to environmental carrying capacity (K).
It is necessary—for a K-selected history—to have some sort of density-dependent pressure. Density-dependent pressures are things such as endemic disease in Africa—which is necessary for a K-selected history since density-dependent natural selection occurs at or close to the environmental carrying capacity (Anderson, 1991: 58). If you truly understood r/K selection theory, you’d understand how it’s based on density dependence. You’d understand that ‘r’ and ‘K’ are not adjectives.
(Indeed, I suspect a golden age in the context of human history will be found to often be such an unusual circumstance, where a population is K-ified, even as it is placed in an r-selected environment of free resource availability. The opposite, an r-ified population placed in a grossly overpopulated environment of shortage will be found to reliably be Hell on earth. Guess which one we have coming.)
You should learn about what r/K selection really is (it is density-dependent selection).
The complete absence of that type of detailed understanding of the effects of selective pressures in the literature about r/K Selection Theory is why I don’t waste extensive time here quoting the source texts on the subject. Most seem strangely shallow in their analyses.
It is detailed, see the table above. Where does alpha-selection fit into your theory? Are conservatives alpha-selected? Not speaking about alpha-selection throws a wrench into the theory. The r/K continuum doesn’t even exist!
I am amused to see the author mention r/K Selection Theory has been linked to ideology, without any mention of where. My greatest hope has always been that r/K Theory would become so ever present in the dialog that nobody would remember where it first arose. When that happens, r/K will be everywhere, and nobody will have any idea who to blame.
Well, the ‘one’s to blame’ would be the originators of the theory, MacArthur and Wilson. But r/K selection is a dead concept in biology and population ecology. Don’t worry, r/K selection is dead and isn’t coming back. I’ve shown how it’s a discredited model.
In regards to r/K being falsified, when the theory was tested, key life history variables did not conform to the predictions of the theory (Graves, 2002: 137). People should stop pushing discredited theories.
By the way, in regards to the one comment that was left, why breakdown complex biological interactions with the environment into something so simple? Can you explain to me how and why complex biological systems interacting with their environment can be broken down ‘simply’? You, as well, have no idea what r/K selection is either.
Anonymous Conservative should try to be aware of his political biases. That much is clear. Although, now I know what will happen. We will see a case of the backfire effect where these corrections will increase his misconceptions of r/K selection theory (Nyhan and Reifler, 2012). Everyone should try keep this quote in mind at all times:
When you are studying any matter, or considering any philosophy, ask yourself only what are the facts and what is the truth that the facts bear out. Never let yourself be diverted either by what you wish to believe, or by what you think would have beneficent social effects if it were believed. But look only, and solely, at what are the facts. That is the intellectual thing that I should wish to say. —Bertrand Russel, 1959
E.O. Wilson on Rushton’s r/K Theory and More on Endemic Disease
1300 words
I thought I’d address what E.O. Wilson’s thoughts on Rushton’s theory and clarify some things on endemic disease and cold winter and how they relate to this r/K paradigm. Proponents of Rushton may look to it and say ‘Well, E.O. Wilson said X, so therefore the reason why it’s not accepted is Y!” However, this comes from a faulty misunderstanding of what Wilson said.
I think Phil is an honest and capable researcher. The basic reasoning by Rushton is solid evolutionary reasoning; that is, it is logically sound. If he had seen some apparent geographic variation for a non-human species – a species of sparrow or sparrow hawk, for example – no one would have batted an eye. … when it comes to [human] racial differences, especially in the inflamed situation in this country, special safeguards and conventions need to be developed.
This little blurb does not address anything, really. Yes, it does address the fact that people attacked Rushton for his research on human racial differences. What it does not address is Rushton’s incorrect application of the theory, as covered yesterday. So, bringing up Wilson’s thoughts on Rushton and the controversy surrounding his theory is a moot point.
I don’t understand why people cannot just accept that Rushton was wrong with his misuse of the theory. Notice how I never said anything about his data—I only talked about his misuse of the theory. People act as if both his data and theory need to be correct, well, why can’t one be right and the other wrong (the data and the theory)? Because that’s how it is in reality.
Rushton’s data was largely correct, however, his misapplication of r/K theory shows that he just saw, for instance, current TFRs (total fertility rates) and just arbitrarily placed Africans as r and Eurasians as K, when looking at what Rushton said about both environments—tropical and cold—would lead to K selection for the tropics, since Rushton asserts that endemic and infectious disease is a selective agent (with no references) while Asia was ‘unbearably cold (also with no reference). This characterization of Pleiositicine environments as ‘hot and endemic disease’ and ‘unbearably cold’ has literally no basis in reality.
Tropical environments are more challenging than cold/temperate ones (Dobzhansky, 1950: 221). Knowing this, Rushton’s assertion of cold winters selecting for higher levels of intelligence in Eurasians compared to Africans is wrong since life is easy nowhere. This characterization of life being ‘easy’ in tropical environments has no basis in reality. It’s like people assume that in the tropics you can just laze around all day while fruits fall onto your lap and you have to do nothing that’s cognitively demanding. This is not true at all. Just look at how a savanna looks, does that look ‘easy’ to live in?
There are also a few more things I’d like to talk about in regards to Rushton’s theory, mainly on endemic disease and why it is an agent of K-selection; not r. Even then, r characteristics probably wouldn’t be able to evolve in the savanna (Miller, 1991: 670). The thing is, populations that evolve in disease-ridden places are expected to select for high population growth—increasing r. However, populations in other areas would increase K as they would be selected for survival and not disease resistance. So if disease was a main difference in so-called r/K differences between populations, r-selected people would be more disease resistant AND they would live longer lives (Miller, 1991: 672).
Case closed, right? Wrong. Miller (1991) writes: “If differences in disease rates do prove to be part of the explanation, the theory would not be an r vs K selection theory, because resistance to disease and a long life span are considered K characteristics, rather than r characteristics” (pg: 672). It is also doubtful that conditions in Africa are much more variable in comparison to other continents.
Furthermore, if an alien observed us with no prior knowledge of our species and only had Pianka’s (1970) paper to go off of, he would conclude that Mongoloids would be r-selected due to the cold winter temperatures which bring a high mortality rate. This is the direct opposite of what Rushton claimed.
Miller states at the end of the article that these differences between populations clearly need explaining. However, the explanation is not r vs. K selection, as Afrosapiens and I showed yesterday, Rushton reversed r and K for the three races, making Africans r when they really would be K and making Mongoloids K, when in reality they would be r. Miller addresses other possibilities, such as testosterone, citing Ellis and Nyborg (1992) for racial differences in testosterone, however, he notes that the difference is only 3 percent which wouldn’t account for racial differences in behavior. (Also recall my critique of having no measure of central adiposity.) I’ve definitively shown that even if the races did differ largely in testosterone that it would not account for disease acquisition nor higher rates of crime.

from Anderson (1991: 53)
Above are the agents of selection, their defining characteristics, and independent and dependent variables. Notice how for r-selection the typical agents of selection are temperature extremes, drought, and natural disaster. For K-selection, the usual agents of selection are limited food supply, endemic infectious disease, and predation. Alpha-selection selects for competitive ability and is thus closer to K than r. Limited resources that can be collected or guarded such as shelter or food are agents of selection.
Clearly, as you can see, if this theory did apply to the human races, Mongoloids would be r and Africans would be K. Endemic disease is an agent of K-selection, not r. This is because endemic disease usually imposes density-dependent selection while cold winters impose density-independent selection. Furthermore, and perhaps most importantly, intelligence can be selected for due to agents of r- or K-selection! Rushton had no reason to add intelligence as a ‘K trait’, as Pianka did not even add it to his continuum. Further, Pianka gave no experimental rationale as to why he placed those traits on r or K (Graves, 2002: 135)! So due to this, Rushton’s claims are wrong and people should stop pushing his r/K theory.
Clearly, Rushton reversed r and K selection and wrongly applied them to the races of man. The three races he describes are NOT local populations, so any inferences made off of any so-called evolutionary environment are not warranted because he did not use the right variable (r or K) for Africans or Eurasians. However, some people may not want to admit that Rushton—and by extension, them—were wrong so they will attempt whatever kind of mental gymnastics possible to attempt to prove that Rushton was ‘right’. As I’ve already said, I don’t have a problem with Rushton’s data; I have a problem with his misapplication of r/K to humans—which I’ve made a strong case that he was wrong and didn’t know what he was talking about in terms of ecology and evolution.
Rushton’s theory was no longer viable 3 years after it was proposed when Judith Anderson got her hands on it, writing the paper Rushton’s Racial Comparisons: An Ecological Critique of Theory and Method. There is literally no saving his application of r and K to humans because he used it wrong! I don’t care what E.O. Wilson said, because he didn’t address Rushton’s application of r/K to human races. He only said if he noticed this variation between another species that no one would have batted an eye. That says absolutely nothing about Rushton’s erroneous application of r/K selection to the races of man.
I hope any HBDers reading this will stop and think for a moment before stating that Eurasians are K and Africans are r. This canard needs to stop in this sphere and I hope I set the wheels in motion to end it.
r/K Selection Theory: A Response to Rushton
3300 words
by RaceRealist and Afrosapiens
Introduction
Jean Phillipe Rushton (1943-2012) was a British-born Canadian psychologist known for his theories on genetically determined racial differences in cognition and behavior between Africans, Europeans, and East Asians. While marginal among experts, Rushton’s theories are still widely accepted amongst the proponents of eugenics and racialism. This article will focus on Rushton’s Differential K-theory which tries to apply the r/K selection model to racial differences in behavioral traits. To be fair, Rushton wasn’t the only one to use r/K selection as an explanation for psychological differences within humanity. For instance, some have associated the continuum with left-wing vs. right-wing ideologies. And although ecologists (the specialists of ecosystems) find applying r/K selection to humans inappropriate, the behavioral sciences have identified life-history patterns that roughly correspond to the colloquial fast vs. slow life differences in life history. For that reason, Rushton may have accidentally discussed variables and trends that are largely acknowledged by experts but his theory lies on a misunderstanding of core principles of the r/K model as well as using flawed (or non-existent) data.
Agents of selection
To begin, confusion about the modes of selection in an ecological context needs to be cleared up. There are classes of natural selection in ecological theory to be discussed: r-selection where the agent of selection acts in a density-independent way; K-selection where the agent of selection acts in a density-dependent way; and alpha selection which is selection for competitive ability (territoriality, aggression). Typical agents of K-selection include food shortage, endemic and infectious disease, and predation. Typical agents of r-selection temperature extremes, droughts, and natural disasters. Typical agents of alpha-selection are limited resources that can be collected or guarded, examples being shelter and food (Anderson, 1991).
As you can see, the third mode of selection in ecological theory is alpha-selection—which Rushton failed to bring up as a mode of selection to explain racial differences in behavior. He didn’t explain his reasoning as to why he did not include it—especially since alpha-selection is selection for competitive ability. One may wonder why Rushton never integrated alpha-selection into his theory—either he was ignorant to the reality of alpha-selection or it could occur in numerous ecosystems—whether temperate/cold or tropical. The non-application of alpha-selection throws his theory into disarray and should have one questioning Rushton’s use of ecological theory in application to human races.
The Misuse of r/K Theory
Ecoregions
Rushton’s model starts with the erroneous assumption that the populations he describes as humanities three main races qualify as ecological populations. When studying the adaptive strategies of organisms, ecologists only consider species within their evolutionary niche—that is, the location that the adaptation was hypothesized to have occurred. When it comes to humans, this can only be done by studying populations in their ancestral environments. For this reason, Africans, Europeans, Amerindians—any population that is not currently in their ancestral environments—are not suitable populations to study in an evolutionary ecological context. The three populations no longer inhabit the environment that the selection was hypothesized to have occurred, so any conclusions based on observing modern-day populations must be viewed with extreme caution (Anderson, 1991). Even in the Old World, constant gene flow between ecoregions, as well as alterations of the environment due to agriculture and then industrialization, make such a study virtually impossible as it would require ecologists to study only hunter-gatherers that have received no admixture from other areas.
Rushton’s next misuse of the theory is not discussing density-dependence and density-independence and how they relate to agents of selection and the r/K model. K-selection works in a density-dependent way while r-selection works in a density-independent way. Thusly, K-selection is expected to favor genotypes that persist at high densities (increasing K) whereas r-selection favors genotypes that increase more quickly at low densities (increasing r) (Anderson, 1991). Rushton also failed to speak about alpha-selection. Alpha-selection selection for competitive abilities and, like with K-selection, occurs at high population densities, but could also occur with low population densities. Alpha-selection, instead of favoring genotypes that increase at high densities “it favours genotypes that, owing to their negative effects on others, often reduce the growth rate and the maximum population size” (Anderson, 1991: 52).
The r/K continuum
The r/K continuum—proposed by Pianka (1970)—has been misused over the decades (Boyce, 1984) and that is where Rushton got the continuum and applied it to human racial differences. Different agents of r-selection produce different selection pressures, as does K-selection. However, where Rushton—and most who cite him—go wrong is completely disregarding the agents of selection, along with perhaps the most critical part, reversing r and K in application to human races (if it were applicable to human races, that is), which will be covered below.
Dobzhansky (1950: 221) notes that “Tropical environments provide more evolutionary challenges than do the environments of temperate and cold lands.” It is erroneously assumed that living in colder temperatures is somehow ‘harder’ than it is in Africa. People believe that since food is ‘readily available’, that it must be ‘harder’ to find food in the temperate/Arctic environments so, therefore, selection for high intelligence occurred in Eurasians while Africans have lower intelligence since it’s so ‘easy’ to live in Africa, as well as other tropical environments.
Africans, furthermore, have been in roughly the same environment since the OoA migration occurred (the Ice Age ‘ended’ about 11,700 ya, although we are still in an Ice Age since the planets caps still have ice), and so any assumptions about it being ‘harder’ for the ancestors of Eurasians to survive and pass on their genes is a baseless assumption. Tropical environments that provide more evolutionary challenges than temperate and cold lands whereas the migration that occurred Out of Africa introduced humans to novel environments. As described above, endemic disease is an agent of K-selection whereas migration to novel environments are agents of r-selection. Thus, cold temperatures would be an agent of r-selection, not K-selection as is commonly believed, whereas endemic disease would be an agent of K-selection.
Even though intelligence nor rule-following were not included on the list of variables that Pianka (1970) noted on his r/K continuum, Rushton chose to include the variables anyway, even though selection for intelligence and rule-following can occur due to agents of r- or K-selection (Anderson, 1991: 55; Graves, 2002: 134-144). Pianka (1970) never gave experimental rationalization as to why he placed the traits he did on his continuum (Graves, 2002: 135). This is one critical point that makes his theory unacceptable in application to racial differences in behavior. By Rushton’s own interpretation of the r/K model, Africans would be selected for intelligence while Eurasians would be selected to breed more since novel environments (i.e., colder temperatures) are agents of r-selection, not K. Using the terms r- and K-selection to describe the traits of an organism is inappropriate; Rushton’s application of r/K theory to the traits of the three races, while ignoring that r/K describes a mode of natural selection “indicates circular reasoning rather than support for Rushton’s hypothesis” (Anderson, 1991: 59).
Reznick et al, (2002: 1518) write: “The distinguishing feature of the r- and K-selection paradigm was the focus on density-dependent selection as the important agent of selection on organisms’ life histories. This paradigm was challenged as it became clear that other factors, such as age-specific mortality, could provide a more mechanistic causative link between an environment and an optimal life history (Wilbur et al. 1974, Stearns 1976, 1977). The r- and K-selection paradigm was replaced by new paradigm that focused on age-specific mortality (Stearns 1976, Charlesworth 1980).” r/K selection theory was dropped for the much stronger life-history approach (Graves, 2002)—which uses some elements of r and K, but otherwise those terms are no longer used since other factors are more important as agents of selection, rather than density dependence and independence as was commonly thought.
Simple models?
One of the main reasons that Rushton’s r/K continuum gets pushed is because it’s a ‘simple model’ that so ‘parsimoniously’ explains racial differences. (e.g., cold winters supposedly take more intelligence to survive in and supposedly are an agent of K-selection.) But ecological systems are never simple; there are numerous interactions between the physical environment and the biological system which interact in complex ways.
Rushton’s use of this ‘simple model’—the r/K continuum—and its application to human races are wrong because 1) the three races described are not local populations; 2) the r/K continuum as described by Pianka (1970) is a poor representation of multidimensional ecological processes; and 3) cold weather is normally an agent of r-selection while endemic disease in Africa—as described by Rushton—is an agent of K-selection. Simple models are not always best—especially for organisms as complex as humans—so attempting to reduce complex biological and environmental interactions into a linear continuum is mistaken (Boyce, 1984). The simpler the ecological model, the more complex ecological sophistication is needed to understand and apply said model. So, although Rushton prefers simple models, in this context it is not apt, as complex biological systems interacting with their environments should not be reduced to a ‘simple model’.
Applying r/K to human races
If the r/K model were applicable to humans, then Caucasoids and Mongoloids would be r-selected while Negroids would be K-selected. Endemic and infectious disease—stated by Rushton to be an r-selected pressure—is actually a K-selected pressure. So Negroids would have been subjected to K-selected pressures (disease) and r-selected pressures (drought). Conversely, for Mongoloids, they migrated into colder temperatures which act in a density-independent way—hence, cold winters (temperature extremes) are an agent of r-selection.
Pianka’s (1970) r/K continuum “confuses the underlying pattern of life history variation with density-dependence, a process potentially involved to explain the pattern” (Gaillard et al, 2016). Furthermore, one cannot make assumptions about an organism’s traits and the selection pressures that caused them without studying said organism in their natural habitat. This seems to be impossible since one would need to study non-admixed hunter-gatherer populations that have received no outside contact.
Gonadotropin levels, testosterone, prostate cancer and r/K theory
Numerous attempts have been made to validate Rushton’s r/K theory. One notable paper by Lynn (1990) attempts to integrate gonadotropin levels and testosterone into Rushton’s r/K continuum. Lynn cites studies showing that blacks have higher testosterone than whites who have higher testosterone than Asians. He then implicates higher levels of both testosterone and gonadotropin levels as the cause for the higher incidence of prostate cancer (PCa) in black Americans.
Lynn (1990) asserts that by having fewer children and showing more care, this is shifting to a K strategy. So, according to Lynn, the best way to achieve this would be a reduction in testosterone. However, there is a fault in his argument.
The study he uses for his assertion is Ross et al (1986). He states that the two groups were both “matched for possible environmental factors which might affect testosterone levels” (Lynn, 1990: 1204). However, this is an erroneous assumption. Ross et al (1986) did control for relevant variables, but made two huge errors. They did not control for waist circumference (WC), and, perhaps most importantly, did not assay the subjects in the morning as close to 8 am as possible.
Testosterone levels are highest at 8 am and lowest at 8 pm. When doing a study like this—especially one to identify a cause of a disease with a high mortality rate—all possible confounds must be identified then controlled for—especially confounds that fluctuate with age. The cohort was assayed between the hours of 10 am and 3 pm. Since testosterone assay time was all over the place for both groups, you cannot draw evolutionary hypotheses from the results. Further, the cohort was a sample of 50 black and white college students—a small sample and a non-representative population. So it’s safe to disregard this hypothesis, on the knowledge that blacks don’t have significantly higher testosterone levels than whites.
Another correlate that is used to show that blacks have higher levels of testosterone is the higher rate of crime they commit. However, physical aggression has a low correlation with testosterone (Archer, 1991; Book et al, 2001) and thusly cannot be the cause of crime. Furthermore, the .14 correlation that Book et al, 2001 found was found to be high. Archer, Graham-Kevan, and Lowe (2005) show that even the .14 correlation between testosterone and aggression is high in a reanalysis of Book et al (2001) since they included 15 studies that should have been omitted. The correlation was then reduced by almost half to .08.
Other theories have been developed to attempt to explain the racial crime gap which centers around testosterone (Ellis, 2017), however, the theory has large flaws which the author rightly notes. Exposure to high levels of testosterone in vitro supposedly causes a low 2d/4d ratio and blacks apparently have the lowest (Manning, 2008). Though, larger analyses show that Asians—mainly the Chinese—have a lower digit ratio compared to other ethnicities (Lippa, 2003; Manning et al, 2007).
Testosterone also does not cause PCa (Stattin et al, 2003; Michaud, Billups, and Partin, 2015). The more likely culprit is diet. Less exposure to sunlight along with low vitamin D intake (Harris, 2006; Rostand, 2010) is a large cause for the prostate cancer discrepancy between the races since low vitamin D is linked to aggressive prostate cancer.
Even then, if there were, say, a 19 percent difference in testosterone between white and black Americans as asserted by Rushton and Lynn, it wouldn’t account for the higher rates of crime, nor higher acquisition and mortality from PCa. If their three claims are false (higher levels testosterone in African-Americans, larger penis size, and high levels of testosterone causing PCa), and they are, then this obliterates Rushton’s and Lynn’s theory.
Differential K Theory has, as noted above, has also been associated with a larger penis for black males in comparison to white males who have larger penises than Asian males (Lynn, 2012), which is not true, there is no reliable data and the data that does exist points to no evidence for the assertion. Lynn, (2012) also used data from a website with unverified and nonexistent sources. In a 2015 presentation, Edward Dutton cites studies showing that, again, Negroids have higher levels of testosterone than Caucasoids who have higher levels of testosterone than Mongoloids. Nevertheless, the claims by Dutton have been rebutted by Scott McGreal who showed that population differences in androgen levels don’t mean anything and that they fail to validate the claims of Lynn and Rushton on racial differences in penis size.
r/K selection theory as an attempt at reviving the scala naturae
Finally, to get to the heart of the matter, Rushton’s erroneous attempt to apply r/K selection theory to the human races is an attempt at reviving the scala naturae concept proposed by Aristotle (Hodos, 2009). The scala naturae organizes living and non-living organisms on a scale from ‘highest’ to ‘lowest’. However, these assumptions are erroneous and have no place in evolutionary biology (Gould, 1996). Rushton (1997: 293) attempted to apply r/K selection theory to human populations to try to revive the concept of the scala naturae, as can be clear by reading the very end of Race, Evolution, and Behavior.
This, of course, goes back to Rushton’s erroneous application of r/K selection theory to human races. He (and others) wrongly assert that Mongoloids are more K-selected than Africans who are more r-selected while Caucasians are in the middle—it also being asserted that K organisms, supposedly Mongoloids, “are the most K evolved” (Lynn, 2012). However, if r/K selection theory were applicable to humans, Mongoloids would be r and Africans would be K. Rushton further attempts to provide evidence for this ‘evolutionary progress’ by citing Dale Russel (1983; 1989) and his thought experiment troodon that he imagines would have eventually have gained human-like bipedalism and a large brain. Nevertheless, Rushton himself doesn’t say that it was only one dinosaur that would have supposedly had human-like intelligence and mobility, Reptile brains, however, lie outside of mammalian design (Hopson, 1977: 443; Gould, 1989: 318), and so, Russel’s theory is falsified.
This use of r/K selection theory as an attempt at bringing back the scala naturae may seem like an intuitive concept; some races/animals may seem more ‘advanced’ or ‘complex’ than others. However, since Rushton’s application of r/K selection theory is not correctly applied (nor does it apply to humans) and any of the claims that Rushton—or anyone else—makes while invoking the theory can be disregarded since he misused r and K selection.
In an attempt to “[restore] the concept of “progress” to its proper place in evolutionary biology,” Rushton (2004) proposed that g—the general factor of intelligence—sits atop a matrix of correlated traits that he proposes to show why evolution is synonymous with ‘progress’, including how and why K evolved organisms are so-called ‘more highly K evolved’—which is a sly attempt to revive the concept of scala naturae. Rushton’s (2004) paper is largely copy and pasted from his 1997 afterword in Race, Evolution, and Behavior—especially the part about ‘progress in evolution’ (which has been addressed in depth).
As can be seen, Ruston attempted to revive the scala naturae by giving it a new name, along with the misuse of ecological theory to make it seem like evolution is synonymous with progress and that K organisms are ‘more evolved’, makes no sense in the context of how ecological theory is (or was) applied to organisms. Rushton’s theory is correct, if and only if he applied r and K correctly to human races. Rushton did not apply r/K selection theory correctly to human races, so Rushton’s claims and any that follow from them are, on their face, immediately wrong. The claims by Rushton et al showing evolution to be ‘progressive’ have been shown to be demonstrably false since evolution is local change, not ‘progress’ (Gould, 1989; 1996).
Conclusion
Rushton’s r/K selection theory has enamored many since he proposed it in 1985. He was relentlessly attacked in the media for his proposals about black penis size, testosterone, brain size, sexual frequency, etc. However, the explanation for said racial differences in behavior—his r/K selection theory—has been summarily rebutted for misapplying ecological theory and not understanding evolution (Anderson, 1991; Graves, 2002). Even ignoring his racial comparisons, his application of the theory would still be unacceptable as he didn’t recognize agents of selection nor alpha selection.
Rushton is wrong because
(i) he misapplied r/K selection in application to human races (Africans would be K, Mongoloids would be r; rule-following and intelligence can be selected for in either environment/with any of the agents of r- or K-selection),
(ii) he arbitrarily designated Africans as r and Mongoloids as K due to current demographic trends (the true application of r and K is described above, which Rushton showed no understanding of),
(iii) the races do not differ in levels of testosterone nor penis size,
(iv) testosterone does not cause prostate cancer nor does it cause crime, so even if there was a large difference between blacks and whites, it would not explain higher rates of PCa in blacks, nor would it explain higher rates of crime,
(v) the scala naturae is a long-dead concept no longer in use by evolutionary biologists, along with its cousin ‘evolutionary progress’, while r/K selection is the attempt at reviving both,
(vi) human races are not local populations; since human races are not local populations then his application of r/K selection to humans is erroneous.
Rushton was informed numerous times he wrongly applied ecological theory to human populations. Yes, E.O. Wilson did say that if Rushton had noticed variation in any other animal that ‘no one would have batted an eye’, however, that does not say a word about Rushton’s incorrect application of r/K selection to human races. No race of humans is more ‘highly evolved’ than another.
Anyone who uses Rushton’s theory as an explanation for observed data is using incorrect/misapplied theory meaning that, therefore, by proxy, their theory is wrong. Rushton’s r/K theory is wrong, and people need to stop invoking it as an explanation for racial differences in behavior, politics, religion, and any other variable they can think of. If Rushton’s application of the theory is wrong, then it logically follows that anything based off of his theory is wrong as well.
The ENA Theory: On Testosterone and Aggressive Behavior by Race/Ethnicity
3250 words
A commenter by the name of bbloggz alerted me to a new paper by Lee Ellis published this year titled Race/ethnicity and criminal behavior: Neurohormonal influences in which Ellis (2017) proposed his theory of ENA (evolutionary neuroandrogenic theory) and applied it to racial/ethnic differences in crime. On the face, his theory is solid and it has great explanatory power for the differences in crime rates between men and women, however, there are numerous holes in the application of the theory in regards to racial/ethnic differences in crime.
In part I, he talks about racial differences in crime. No one denies that, so on to part II.
In part II he talks about environmental causes for the racial discrepancies, that include economic racial disparities, racism and societal discrimination and subordination, a subculture of violence (I’ve been entertaining the honor culture hypothesis for a few months; Mazur (2016) drives a hard argument showing that similarly aged blacks with some college had lower levels of testosterone than blacks with less than high school education which fits the hypothesis of honor culture. Though Ellis’ ENA theory may account for this, I will address this below). However, if the environment that increases testosterone is ameliorated (i.e., honor culture environments), then there should be a subsequent decrease in testosterone and crime, although I do believe that testosterone has an extremely weak association with crime, nowhere near high enough to account for racial differences in crime, the culture of honor could explain a good amount of the crime gap between blacks and whites.
Ellis also speaks about the general stress/strain explanation, stating that blacks have higher rates of self-esteem and Asians the lowest, with that mirroring their crime rates. This could be seen as yet another case for the culture of honor in that blacks with a high self-esteem would feel the need to protect their ‘name’ or whatever the case may be and feel the need for physical altercation based on their culture.
In part III, Ellis then describes his ENA theory, which I don’t disagree with on its face as it’s a great theory with good explanatory power but there are some pretty large holes that he rightly addresses. He states that, as I have argued in the past, females selected men for higher rates of testosterone and that high rates of testosterone masculinize the brain, changing it from its ‘default feminine state’ and that the more androgens the brain is exposed to, the more likely it is for that individual to commit crime.
Strength
Ellis cites a study by Goodpaster et al (2006) in which he measured the races on the isokinetic dynamometry, pretty much a leg extension. However, one huge confound is that participants who did not return for follow-up were more likely to be black, obese and had more chronic disease (something that I have noted before in an article on racial grip strength). I really hate these study designs, but alas, it’s the best we have to go off of and there are a lot of holes in them that must be addressed. Though I applaud the researchers’ use of the DXA scan (regular readers may recall my criticisms on using calipers to assess body fat in the bench press study, which was highly flawed itself; Boyce et al, 2014) to assess body fat as it is the gold standard in the field.
Ellis (2017: 40) writes: “as brain exposure to testosterone surges at puberty, the prenatally-programmed motivation to strive for resources, status, and mating opportunities will begin to fully activate.” This is true on the face, however as I have noted the correlation between physical aggression and testosterone although positive is low at .14 (Archer, 1991; Book et al, 2001). Testosterone, as I have extensively documented, does cause social dominance and confidence which do not lead to aggression. However, when other factors are coupled with high testosterone (as noted by Mazur, 2016), high rates of crime may occur and this may explain why blacks commit crime; a mix of low IQ, high testosterone and low educational achievement making a life of crime ‘the smart way’ to live seeing as, as Ellis points out, and that intelligent individuals find legal ways to get resources while less intelligent individuals use illegal ways.
ENA theory may explain racial differences in crime
In part IV he attempts to show how his ENA theory may explain racial differences in crime—with testosterone sitting at the top of his pyramid. However, there are numerous erroneous assumptions and he does rightly point out that more research needs to be done on most of these variables and does not draw any conclusions that are not warranted based on the data he does cite. He cites one study in which testosterone levels were measured in the amniotic fluid of the fetus. The sample was 59 percent white and due to this, the researchers lumped blacks, ‘Hispanics’ and Native Americans together which showed no significant difference in prenatal testosterone levels (Martel and Roberts, 2014).
Umbilical cord and testosterone exposure
Ellis then talks about testosterone in the umbilical cord, and if the babe is exposed to higher levels of testosterone in vitro, then this should account for racial/ethnic differences in crime. However, the study he cited (Argus-Collins et al, 2012) showed no difference in testosterone in the umbilical cord while Rohrmann et al (2009) found no difference in testosterone between blacks and whites but found higher rates of SHBG (sex hormone-binding globulin) which binds to testosterone and makes it unable to leave the blood which largely makes testosterone unable to affect organ development. Thusly, if the finding of higher levels of SHBG in black babes is true, then they would be exposed to less androgenic hormones such as testosterone which, again, goes against the ENA theory.
He also cites two more studies showing that Asian babes have higher levels of umbilical cord testosterone than whites (Chinese babes were tested) (Lagiou et al, 2011; Troisi et al, 2008). This, again, goes against his theory as he rightly noted.
Circulating testosterone
Next he talks about circulating differences in testosterone between blacks and whites. He rightly notes that testosterone must be assayed in the morning within an hour after waking as that’s when levels will be highest, yet cites Ross et al (1986) where assay times were all over the place and thusly testosterone cannot be said to be higher in blacks and whites based on that study and should be discarded when talking about racial differences in testosterone due to assay time being between 10 am and 3 pm. He also cites his study on testosterone differences (Eliss and Nyborg, 1993), but, however, just as Ross et al (1986) did not have a control for WC (waist circumference) Ellis and Nyborg (1993) did not either, so just like the other study that gets cited to show that there is a racial difference in testosterone, they are pretty hugely flawed and should not be used in discussion when discussing racial differences in testosterone. Why do I not see these types of critiques for Ross et al (1986) in major papers? It troubles me…
He also seems to complain that Lopez et al (2013) controlled for physical activity (which increases testosterone) and percent body fat (which, at high levels, decreases testosterone). These variables, as I have noted, need to be controlled for. Testosterone varies and fluctuated by age; WC and BMI vary and fluctuate by age. So how does it make sense to control for one variable that has hormone levels fluctuate by age and not another? Ellis also cites studies showing that older East Asian men had higher levels of testosterone (Wu et al, 1995). Nevertheless, there is no consensus; some studies show Chinese babes have higher levels of testosterone than whites and some studies show that whites babes have higher levels of testosterone than Chinese babes. Indeed, this meta-analysis by Ethnicmuse shows that Asians have the highest levels, followed by Africans then Europeans, so this needs to be explained to save the theory that testosterone is the cause of black overrepresentation of violence (as well as what I showed that testosterone is important for vital functioning and is not the boogeyman the media makes it out to be).
Bone density and crime
Nevertheless, the next variable Ellis talks about is bone density and its relationship to crime. Some studies find that blacks are taller than whites while other show no difference. Whites are also substantially taller than Asian males. Blacks have greater bone density than the other three races, but according to Ellis, this measure has not been shown to have a relationship to crime as of yet.
Penis size, race and crime
Now on to penis size. In two articles, I have shown that there is no evidence for the assertion that blacks have larger penises than whites. However, states that penis length was associated with higher levels of testosterone in Egyptian babes. He states that self-reported penis size correlates with self-reports of violent delinquency (Ellis and Das, 2012). Ellis’ main citations for the claim that blacks have larger penises than other races comes from Nobile (1982), the Kinsey report, and Rushton and Boagert (1987) (see here for a critique of Rushton and Boagert, 1987), though he does cite a study stating that blacks had a longer penis than whites (blacks averaging 5.77 inches while whites averaged 5.53 inches). An HBDer may go “Ahah! Evidence for Rushton’s theory!”, yet they should note that the difference is not statistically significant; just because there is a small difference in one study also doesn’t mean anything for the totality of evidence on penis size and race—that there is no statistical difference!
He then cites Lynn’s (2013) paper which was based on an Internet survey and thus, self-reports are over-measured. He also cites Templer’s (2002) book Is Size Important?, which, of course, is on my list of books to read. Nevertheless, the ‘evidence’ that blacks average larger penises than whites is extremely dubious, it’s pretty conclusive that the races don’t differ in penis size. For further reading, read The Pseudoscience of Race Differences in Penis Size, and read all of Ethnicmuses’ posts on penis size here. It’s conclusive that there is no statistical difference—if that—and any studies showing a difference are horribly flawed.
2d/4d ratio and race
Then he talks about 2d/4d ratio, which supposedly signifies higher levels of androgen exposure in vitro (Manning et al, 2008) however these results have been challenged and have not been replicated (Koehler, Simmons, and Rhodes, 2004; Yan et al, 2008, Medland et al, 2010). Even then, Ellis states that in a large analysis of 250,000 respondents, Asians had the lowest 2d/4d ratio, which if the hypothesis of in vitro hormones affecting digit length is to be believed, they have higher levels of testosterone than whites (the other samples had small ns, around 100).
Prostate-specific antigens, race, and prostate cancer
He then talks about PSA (prostate-specific antigen) rates between the races. Blacks are two times more likely to get prostate cancer, which has been blamed on testosterone. However, I’ve compiled good evidence that the difference comes down to the environment, i.e., diet. Even then, there is no evidence that testosterone causes prostate cancer as seen in two large meta-analyses (Stattin et al, 2003; Michaud, Billups, and Partin, 2015). Even then, rates of PCa (prostate cancer) are on the rise in East Asia (Kimura, 2012; Chen et al, 2015; Zhu et al, 2015) which is due to the introduction of our Western diet. I will cover the increases in PCa rates in East Asia in a future article.
CAG repeats
He then reviews the evidence of CAG repeats. There is, however, no evidence that the number of CAG repeats influences sensitivity to testosterone. However, intra-racially, lower amounts of CAG repeats are associated with higher spermatozoa counts—but blacks don’t have higher levels of spermatozoa (Mendiola et al, 2011; Redmon et al, 2013). Blacks do have shorter CAG repeats, and this is consistent with the racial crime gap of blacks > whites > Asians. However, looking at the whole of the evidence, there is no good reason to assume that this has an effect on racial crime rates.
Intelligence and education
Next he talks about racial differences in intelligence and education, which have been well-established. Blacks did have higher rates of learning disabilities than whites who had higher levels of learning disabilities then Asians in a few studies, but other studies show whites and South Asians having different rates, for instance. He then talks about brain size and criminality, stating that the head size of males convicted for violent crimes did not differ from males who committed non-violent crimes (Ikaheimo et al, 2007). I won’t bore anyone with talking about what we know already: that the races differ in average brain size. However, a link between brain size and criminality—to the best of my knowledge—has yet to been discovered. IQ is implicated in crime, so I do assume that brain size is as well (no matter if the correlation is .24 or not; Pietschnig et al, 2015).
Prenatal androgen exposure
Now to wrap things up, the races don’t differ in prenatal androgen exposure, which is critical to the ENA theory; there is a small difference in the umbilical cord favoring blacks, and apparently, that predicts a high rate of crime. However, as noted, blacks have higher levels of SHBG at birth which inhibits the production of testosterone on the organs. Differences in post-pubertal testosterone are small/nonexistent and one should not talk about them when talking about differences in crime or disease acquisition such as PCa. DHT only shows a weak positive correlation with aggression—the same as testosterone (Christiansen and Winkler, 1992; however other studies show that DHT is negatively correlated with measures of physical aggression; Christiansen and Krussmann, 1987; further, DHT is not so evil after all).
Summing it all up
Blacks are not stronger than whites, indeed evidence from the races’ differing somatype, grip strength and leverages all have to do with muscular strength. Furthermore, the study that Ellis cites as ‘proof’ that blacks are stronger than whites is on one measure; an isokinetic dynamometry machine which is pretty much a leg extension. In true tests of strength, whites blow blacks away, which is seen in all major professional competitions all around the world. Blacks do have denser bones which is due to androgen production in vitro, but as of yet, there has been no research done into bone density and criminality.
The races don’t differ on penis size—and if they do it’s by tenths of an inch which is not statisitcally significant and I won’t waste my time addressing it. It seems that most HBDers will see a racial difference of .01 and say “SEE! Rushton’s Rule!” even when it’s just that, a small non-significant difference in said variable. That’s something I’ve encountered a lot in the past and it’s, frankly, a waste of time to converse about things that are not statistically significant. I’ve also rebutted the theory on 2d/4d ration as well. Finally, Asians had a similar level of androgen levels compared to blacks, with whites having the least amount. Along with a hole in the theory for racial differences in androgen causing crime, it’s yet another hole in the theory for racial differences in androgens causing racial differences in penis size and prostate cancer.
On intelligence scores, no one denies that blacks have scored about 1 SD lower than whites for 100 years, no one denies that blacks have a lower educational attainment. In regards to learning disabilities, blacks seem to have the highest rates, followed by Native Americans, than non-Hispanic whites, East Asians and the lowest rates found in South Asians. He states only one study links brain size to criminal behavior and it showed a significant inverse relationship with crime but not other types of offenses.
This is a really good article and I like the theory, but it’s full of huge holes. Most of the variables described by Ellis have been shown to not vary at all or much between the races (re: penis size, testosterone, strength [whites are stronger] prostate cancer caused mainly by diet, 2d/4d ratio [no evidence of it showing a digit ratio difference], and bone density not being studied). Nevertheless, a few of his statements do await testing so I await future studies on the matter. He says that androgen exposure ‘differs by race and ethnicity’, yet the totality of evidence shows ‘not really’ so that cannot be the cause of higher amounts of crime. Ellis talks about a lot of correlates with testosterone, but they do not pass the smell test. Most of it has been rebutted. In fact, one of the central tenets of the ENA theory is that the races should differ in 2d/4d ratio due to exposure of differing levels of the hormone in vitro. Alas, the evidence to date has not shown this—it has in fact shown the opposite.
ENA theory is good in thought, but it really leaves a lot to be desired in regards to explaining racial differences in crime. More research needs to be looked into in regards to intelligence and education and its effect on crime. We can say that low IQ people are more likely to drop out of school and that is why education is related to crime. However, in Mazur (2016) shows that blacks matched for age had lower levels of testosterone if they had some college under their belt. This seems to point in the direction of the ENA theory, however then all of the above problems with the theory still need to be explained away—and they can’t! Furthermore, one of the nails in the coffin should be this: East Asian males are found to have higher levels of testosterone than white males, often enough, and East Asian males actually have the lowest rate of crime in the worle!
This seems to point in the direction of the ENA theory, however then all of the above problems with the theory still need to be explained away—and they can’t! Furthermore, one of the nails in the coffin should be this: East Asian males are found to have higher levels of testosterone than white males, often enough, and East Asian males actually have some of the lowest rate of crime in the world (Rushton, 1995)! So this is something that needs to be explained if it is to be shown that testosterone facilitates aggression and therefore, crime.
Conclusion
I’ve shown—extensively—that there is a low positive correlation between testosterone and physical aggression, why testosterone does not cause crime, and have definitively shown that, by showing how flawed the other studies are that purport to show blacks have higher testosterone levels than whites, along with citing large-scale meta-analyses, that whites and blacks either do not differ or the differences is small to explain any so-called differences in disease acquisition or crime. One final statement on the CAG repeats, they are effect by obesity, men who had shorter CAG repeats were more likely to be overweight, which would skew readings (Gustafsen, Wen, and Koppanati, 2003). So depending on the study—and in most of the studies I cite whites have a higher BMI than blacks—BMI and WC should be controlled for due to the depression of testosterone.
It’s pretty conclusive that testosterone itself does not cause crime. Most of the examples cited by Ellis have been definitively refuted, and his other claims lack evidence at the moment. Even then, his theory rests on the 2d/4d ratio and how blacks may have a lower 2d/4d ratio than whites. However, I’ve shown that there is no significant relationship between 2d/4d ratio and traits mediated by testosterone (Kohler, Simmons, and Rhodes, 2004) so that should be enough to put the theory to bed for good.
Why Testosterone Does Not Cause Crime
1900 words
Edit: (The correlation between aggression and testosterone isn’t .14 as Book et al (2001) state; the true correlation is .08 (Archer, Graham-Kevan and Davies, 2005) So it’s even lower than I thought. This is one of the many reasons why testosterone does not cause crime. It’s just feminist bullshit and fear mongering from people who do not understand the hormone and what it does in the body. The misconceptions come from Rushton’s r/K selection bullshit which has been summarily refuted.)
Recently, I’ve written at length on racial differences in testosterone and how the correlation between testosterone and physical aggression is .14. Pitifully low to account for the cause of crime and any overall differences in racial crime (that will be touched on at length in the future). Tonight I will show, yet again, why testosterone does not cause crime by looking at what times most crimes are committed by both adults and children under the age of 18. This will definitively put the ‘testosterone causes crime’ myth to bed for good.
Before I get into the time of day that most crimes are committed, I must talk about the production of testosterone in the body. There are no ‘genes for’ testosterone (although men who had three certain alleles had a 6.5 fold higher risk of having low testosterone; Ohlsson et al, 2011, I am unaware of there being a variation by race; over 10,000 Caucasian men were studied). There is, however, an indirect control of testosterone synthesis by DNA. DNA regulates the production of testosterone by coding for enzymes that convert cholesterol to testosterone (testosterone is a cholesterol-based hormone).
There are five simple steps to the production of testosterone: 1) DNA codes for mRNA; 2) mRNA codes for the synthesis of an enzyme in the cytoplasm; 3) luteinizing hormone stimulates the production of another messenger in the cell when testosterone is needed; 4) this second messenger activates the enzyme; 5) the enzyme then converts cholesterol to testosterone (Leydig cells produce testosterone in the presence of luteinizing hormone). That’s how testosterone is produced in the body. It is indirectly controlled by DNA.

Above is a graph from the Office of Juvenile Justice and Delinquency Prevention which shows the time of day that most crimes are committed. Notice how crime goes up as the time of day goes on and since kids are at school, they’re more likely to fight. This then peaks at 3 pm when kids are getting out of school.
Now look at rates of crime for adults. At its peak of 10 pm, it’s vastly lower than that of people under the age of 18, which is important to keep in mind. You can see how at 8 am that rates of crime are low for adults and high for kids, right when they would be entering school so there would be a lot of other kids around and the chance for violence goes up. Keep the times of 8 am (kids when they enter school), 12 pm (when most kids go on lunch) and 3 pm (when most kids get out of school) along with the hours of 12 pm to 8 pm for adults (when 74 percent of crimes are committed by adults).
- In general, the number of violent crimes committed by adults increases hourly from 6 a.m. through the afternoon and evening hours, peaks at 10 p.m., and then drops to a low point at 6 a.m. In contrast, violent crimes by juveniles peak in the afternoon between 3 p.m. and 4 p.m., the hour at the end of the school day.
- Nearly one-third (29%) of all violent crime committed by juvenile offenders occurs between 3 p.m. and 7 p.m. In comparison, 26% of all violent committed by adult offenders occurs between 8 p.m. and 12 p.m.
So since testosterone varies by day and levels are highest at 8 am and lowest at 8 pm (Brambilla et al, 2009; however testing men aged 45 years of age and older is fine before 2 pm due to a blunted circadian rhythm; Long, Nguyen, and Stevermer, 2015), then how could testosterone account for why men commit most of their crimes at night and why the crime that children commit spikes when they go to school, go to lunch and get out of school? The answer is that it doesn’t because testosterone does not cause crime. What testosterone does cause, however, are feelings of confidence and dominance, which does not—surprisingly—lead to increased aggression and assault on others (Booth et al, 2006).
What testosterone does cause, however, is social dominance and success, not physical aggression and maladjustment (Shcaal et al, 1996). The effects of environment are also more notable on testosterone than are genetics at 5 months of age (Carmaschi et al, 2010). Furthermore, aggressive behavior is first noticed in infancy and reaches its peak before school age (Tremblay et al, 2004; Cote et al, 2006). Though testosterone does seem to have an effect on aggression in preschool boys, however genetic and environmental causality has not been established (Sanchez-Martin et al, 2000).
Nevertheless, the meta-analyses I cited last week show that testosterone has an extremely low correlation of .14, so other factors must be at play. However, Sanchez-Martin et al (2000:778-779) also note that “Tremblay et al (1998) suggested that associations between testosterone titer and physical aggression are likely to be observed in contexts where such attack leads to social dominance. This may be true of the preschool boys in the present study. The data generated in the present study generally support Scerbo and Kolko (1994), who studied older children (7 to 14 years of age). They found a significant relationship between testosterone levels and aggression (as assessed by clinical staff).”
It’s interesting to note that in the case of Scerbo and Kolko (1994) that after controlling for age and size, testosterone correlated with aggression when rated by staff but not parents or teachers. ‘Staff’ refers to clinic staff at a facility where the children were assessed for hyperactivity disorders. Of course, the staff would rate higher levels of aggression compared to parents of teachers—people who are around the children every day—since they would want a higher chance for diagnosis for certain drugs to ‘cure’ the hyperactivity, but I digress. Testosterone does not induce aggression in children, but it does induce social dominance and confidence which does not lead to aggression (Rowe et al, 2004; Booth et al, 2006).
There was also little difference in testosterone between socially dominant prisoners and aggressive prisoners (Ehrenkraz, Bliss, and Sheard, 1974). Furthermore, the testosterone increase leading to pubertal development in boys is not associated with increased aggression (Tremblay et al, 1998; Booth et al, 2006: 171). Indeed, increased body size is a marker for physical aggression in children, and I doubt these children have high muscle mass so, I assume, they have high levels of body fat and thusly lower levels of testosterone than they would have if they were leaner. Yet another strike against the ‘testosterone causes crime/physical aggression’ hypothesis.
Indeed, this has some implications for the honor culture hypothesis of why low-income blacks have higher levels of testosterone than similarly aged blacks with some college (Mazur, 2016). The patterns for crime as shown by the OOJDP shows that crime rises as the day progresses from the morning until its peak at 3 pm for children and then sharply declines while for adults it peaks at 10 pm.
Testosterone does increase when a challenge is issued; when one man feels his reputation is threatened, the propensity for violence is increased, but this was most notably seen in Southern men (Cohen et al, 1996). So the same would be said for this ‘culture of honor’ found in low-income black neighborhoods, the so-called ‘code of the street’ as stated by Anderson (1994: 88): “Moreover, if a person is assaulted, it is important, not only in the eyes of his opponent but in the eyes of his “running buddies,” for him to avenge himself. Otherwise, he risks being “tried” (challenged) or “moved on” by any number of others. To maintain his honor, he must show he is not someone to be “messed with” or “dissed.””
This culture of honor is found all over the world, including Brazil where homicide can be explained by the need to maintain honor and can be understood by taking into account cultural factors; biological, psychological and socioeconomic factors do not explain murder in Northeast Brazil as well as honor and culture (de Souza et al, 2015). People in honor cultures also have a higher chance of self-harm (Osterman and Brown, 2011) as well as a higher chance of committing violence in school (Brown, Osterman, and Barnes, 2009).
Testosterone does not cause crime; it does not cause aggression. Increases in testosterone before, during and after events are a physiologic process to prime the body for competition. As cited above, dominant behavior does not necessarily lead to violence in most cases, which may be surprising for some. Indeed, honor and culture may explain a nice amount of the homicide and violence rate in the South. Since testosterone is highest at 8 am and lowest at 8 pm and the rates of crime committed by adults and children are vastly different than the diurnal variance in the day, then testosterone does not cause crime and its increase is not associated with crime, but social dominance and confidence which does not lead to crime.
Hopefully—if anyone still believes testosterone to be the boogeyman its made out to be—I’ve put those misconceptions to rest. Racial differences in testosterone cannot be the cause of racial differences in crime—because there is either no statistical difference in testosterone between the races or the difference is non-existent. Testosterone is clearly a beneficial hormone—as I have extensively documented. Misunderstandings of the hormone are abound—especially in the HBD sphere—only due to literally a few paragraphs in a book (Rushton, 1997) and one study that showed blacks have higher testosterone than whites which was the cause of their higher rates of prostate cancer (Ross et al, 1986). The study is hard to find so I had to buy access to it. I will cover this in the future, but I discovered that they assayed the subjects when it was convenient for them—between the hours of 10 am and 3 pm—which is unacceptable. You cannot gauge racial differences in testosterone from a small study (n=50) and a non-representative sample (college students). For these reasons, the study should be thrown in the trash—especially when formulating evolutionary hypotheses.
Testosterone is one of the most important hormones for vital functioning. By knowing how it is processed in the body and that there are no ‘genes for’ testosterone (‘low testosterone genes’ notwithstanding) along with how testosterone has a low relationship with physical aggression one should not be scared of having high levels, on the contrary, one should be scared of having low levels. I have once again proven my case that testosterone is not related to violence in showing the diurnal variation in testosterone levels in adults, as well as the time of day that crimes are committed by both adults and children. High testosterone means high confidence and high dominance—and those two traits have a lot to do with masculinity—which do not lead to violence.
I know why testosterone does not cause crime—because I have an understanding of the hormone, how its produced in the body and what its effects on the body are. The most important thing to note here, is that even if blacks had 15 percent higher testosterone than whites, it still wouldn’t explain higher rates of crime or disease such as prostate cancer. So those who try so hard to prove that blacks have higher levels of the hormone do so in vain, because even if they did it wouldn’t mean anything for any theories they may have. The myth of testosterone causing aggression and crime need to be put to bed for good.
Our View of ‘Intelligence’ Needs to Change
900 words
What is intelligence? How would we define it? Would intelligence be reacting to what occurs in the immediate environment; having the ability to have behavioral plasticity or even communicating with others? Amazingly, bacteria have been found to do both things noted above: They have been found to be able to react to their environment, i.e., have the ability for plastic behavior and they have even been shown to communicate with one another. Hell, even something as simple as a slime mold has been found to navigate a maze to find food. Is that not intelligence?
Ken Richardson, author of the book Genes, Brains, and Human Potential: The Science and Ideology Behind Intelligence writes:
Living things, then, need to be good at registering those statistical patterns across everyday experience and then use them to shape the best response, including (in the cell) what genes to recruit for desired products. This is what intelligence is, and it’s origins coincide with the origins of life itself, and life is intelligence. (Richardson, 2017: 115)
In multicelluar systems, of course, the cells are not just responding to one another, but also collectively to the changing environment outside. That requires an intelligent physiology, as described in chapter 5. However, it is still the statistical structure of the changes that matters and that forms the basis of a living intelligence. Even at this level, closest to the genes, then, the environment is emphatically not a loose collection of independent factors to which the cells respond, in stimulus-response fashion, under gene control. This reality makes the additive statistical models of the behavioral geneticist quite unrealistic. (Richardson, 2017: 120)
Currently, our view of intelligence has an anthropometric lean. But, as I’ve been saying for months now, why should we view humans as a sort of ‘apex’ to evolution? Why should we be the measuring stick? If you really think about it to put us—our brains—at the top of a rank order as ‘the best’ and not recognize what other, smaller supposedly ‘archaic’ forms of life can do, then maybe it’s best to take off our human-centric glasses and look at the whole of the animal kingdom as intelligent—including bacteria, as they show the basic things necessary for what we would call intelligence, i.e., behavioral plasticity.
In this paper published just two months ago, the authors write:
Bacteria are far more intelligent than we can think of. They adopt different survival strategies to make their life comfortable. Researches on bacterial communication to date suggest that bacteria can communicate with each other using chemical signaling molecules as well as using ion channel mediated electrical signaling. (Majumdar and Pal, 2017)
Furthermore, looking at definitions of the term ‘behavior’ from ethology, we can see that bacteria exhibit these behaviors that we have deemed ‘human’ or ‘human-like’:
- “Externally visible activity of an animal, in which a coordinated pattern of sensory, motor and associated neural activity responds to changing external or internal conditions” (Beck et al. 1981)
- “A response to external and internal stimuli, following integration of sensory, neural, endocrine, and effector components. Behavior has a genetic basis, hence is subject to natural selection, and it commonly can be modified through experience” (Starr and Taggart 1992)
- “Observable activity of an organism; anything an organism does that involves action and/or response to stimulation” (Wallace et al. 1991)
- “What an animal does” (Raven and Johnson 1989)
Bacteria have been found to fit all of the criteria mentioned above. If organisms can react to how the environment changes, then that organism has—at least a semblance—of intelligence. Bacteria have also been found to be able to learn and they also have memories, so if this is true (and it is), then bacteria are intelligent.
Finally, Westerhoff et al (2014) write that leaving out the terms ‘human’ and our brains as measuring sticks for what is intelligent, that “all forms of life – from microbes to humans – exhibit some or all characteristics consistent with “intelligence.” For people with anthropocentric views of evolution, however, this is a hard pill to swallow. If the data says that bacteria have evidence of ‘cognition’ and an ability to react to outside environmental cues then bacteria have a semblance of intelligence. There is no denying it.
We clearly need to look at intelligence in a different way—one that’s free of any anthropocentric bias—-and if we do, we would recognize numerous species as intelligent that we would never have thought of before since we view ourselves as some sort of ‘apex’ of evolution, that we are supreme on this earth, when the bacteria—the modal bacter—reign supreme and will continue to remain supreme until the Sun explodes. So if bacteria show the ability to communicate with one another and the ability to change their behavior when their environment changes, i.e., that they learn and have ‘memories’ of past events, then maybe it’s time for us to change from our human-centric view of intelligence (which makes a ton of sense; viewing us as an ‘apex’ of evolution makes no sense and doesn’t allow us to appreciate the wide range of variation on earth).
As Gould wrote in Full House, looking at only the right tail we would believe that some sort of ‘progress’ reigns supreme, but looking at the whole sum of variation, we can see that the bacteria are the mode of all life, have been the mode of all life and will remain the mode of all life until the Sun explodes and all life forever perishes from Earth.
Testosterone and Aggressive Behavior
1200 words
Testosterone gets a bad rep. People assume that if one has higher testosterone than average, that they will be a savage, bloodthirsty beast with an insatiable thirst for blood. This, however, is not the case. I’ve documented how testosterone is vital for male functioning, and how higher levels don’t lead to maladies such as prostate cancer. Testosterone is feared for no reason at all. The reason that people are scared of it is that of the anecdotal reports that individual A had higher testosterone when he committed crime B so, therefore, anyone who commits a crime has higher testosterone and that is the ultimate—not proximate—cause of crime. This is erroneous. There is a positive—albeit extremely low—correlation between physical aggression and violence at .14. That’s it. Furthermore, most of these claims of higher levels of testosterone causing violence is extrapolated from animal studies to humans.
Testosterone has been shown to lead to violent and aggressive behavior, largely only in animal studies (Archer, 1991; Book et al, 2001). For years, the relationship between the two variables was thought to be causal, i.e., high levels of testosterone cause violent crimes, which has been called into question over recent years. This is due to how the environment can raise testosterone levels. I have documented how these environmental factors can raise testosterone—and after these events, testosterone stays elevated.
Largely, animal studies are used to infer that high levels of testosterone in and of themselves lead to higher rates of aggression and therefore crime. However, two important meta-analyses show this is not necessarily the case (Archer, 1991; Book et al, 2001). Book et al, 2001 showed that two variables were important in seeing the relationship between aggression and crime—the time of day that the assay was taken and the age of the participant. This effect was seen to be largest in, not unexpectedly, males aged 13-20 (Book et al, 2001: 594). So since age confounds the relationship between aggression and testosterone in males, that is a variable that must also be controlled for (which, in the meta-analyses and other papers I cite on black and white testosterone is controlled for).
More interestingly, Book et al (2001) showed that the nature of the measure of aggression (self-reported or behavioral) did not have any effect on the relationship between testosterone and aggression. Since there is no difference between the two measures, then a pencil-and-paper test is a good enough index of measure of aggression, comparable to observing the behavior of the individual studied.
Archer (1991) also showed the same low—but positive—correlations between aggression and testosterone. Of course, as I’ve extensively documented since there is a positive relationship between the two variables does not necessarily mean that high-testosterone men commit more crime—since the outcome of certain situations can increase and decrease testosterone, no causal factors have been detangled. Book et al (2001) confirmed Archer’s (1991) finding that the correlation between violent and aggressive behavior was positive and low at .14.
Valois et al (2017) showed there was a relationship between emotional self-efficacy (ESE) and aggressive and violent behaviors in a statewide sample of high school children in South Carolina (n=3,386). Their results suggested that there was a relationship between carrying a weapon to school within the past 30 days along with being injured with a club, knife or gun in the past 12 months was significantly associated with ESE for specific race and sex groups.
Black girls who reported a low ESE reported carrying a weapon to school 30 days prior to the survey were 3.22 times more than black girls with a high ESE who did not report carrying a weapon to school within the past 30 days prior to the questionnaire. For black boys with low ESE, they were 3.07 times more likely to carry a weapon to school within the past 30 days in comparison to black boys with high ESE who did not carry a weapon to school in the past 30 days. White girls who reported low ESE had the highest chance of bringing a weapon to school in comparison to white girls with low ESE—they were 5.87 times more likely to carry a weapon to school 30 days prior to the survey. Finally, white boys with low ESE were slightly more than 2 times more likely than white boys with high ESE to carry a weapon to school 30 days prior to the survey.
Low ESE in white and black girls is associated with carrying a weapon to school, whereas low ESE for white and black boys is associated with being threatened. Further, their results suggested that carrying a weapon to school was associated with low ESE in black and white girls suggesting that low ESE is both situation-specific and specific to the female sex. The mediator between these things is low ESE—it is different for both black boys and black girls, and when it occurs different courses of action are taken, whether it’s through bringing a weapon to school or being threatened. What this tells me is that black and white boys with low ESE are more likely to be threatened because they are perceived to be more meek, while black and white girls with low ESE that get provoked at school are more likely to bring weapons. So it seems that girls bring weapons when provoked and boys fight.
The two meta-analyses reviewed above show that there is a low positive (.14) correlation between testosterone and aggression (Archer, 1991; Book et al, 2001). Thusly, high levels of testosterone on their own are not sufficient enough to explain high levels of aggression/violence. Further, there are race- and sex-specific differences when one is threatened at high school with black and white boys being more likely to report being threatened more (which implies a higher rate of physical fighting) while black and white girls when threatened brought weapons to school. These race- and sex-specific differences in the course of action taken when they are physically threatened needs to be looked into more.
I’d like to see the difference in testosterone levels for a matched sample of black and white boys from two neighboring districts with different murder rates as a proxy for the amount of violence in the area. I’d bet that the places with a higher murder rate would have children 1) report more violence and instances of bringing weapons to school and 2) report more harm from these encounters—especially if they have low ESE as seen in Valois (2017) and 3) the children in the high schools along with the residents of the area would have higher testosterone than the place with less violence. I would expect these differences to be magnified in the direction of Valois (2017) in that areas with higher murder rates would have black and white girls report bringing weapons to school when threatened whereas black and white boys would report more physical violence.
High testosterone itself is not sufficient enough to explain violence as the correlation is extremely low at .14. Testosterone levels fluctuate depending on the time of day (Brambilla et al, 2009; Long, Nguyen, and Stevermer, 2015) to the time of year (Stanton, Mullette-Gillman, and Huettel, 2011; Demur, Uslu, and Arslun, 2016). How the genders/races react differently when threatened in adolescence is interesting and deserves further study.
You Are what you Eat, Including Brains
by Scott Jameson
850 words
I’ve been thinking about Omega 3 fatty acids (N3s) recently. We’re clearly adapted to getting more of them than we’re actually getting.
Children whose mothers took fish oil (chock full of N3s EPA and DHA) during pregnancy have higher coordination, and they are smarter, although that difference may not persist later in development. RaceRealist has written about N3s and PISA math scores before.
Here’s a paper summarizing many of the known benefits of fish oil supplementation. Goes over some of the aforementioned results regarding kids, and also shows random improvements such as helping people with Alzheimer’s maintain weight.
A lack of N3s is associated with depression, coronary artery disease, maybe even autism. N3s are known to lessen autism symptoms. This is probably an immune thing. Sulforaphane, an anti-inflammatory agent, is known to reduce autism symptoms, and other research has elucidated the relationship between N3s and the immune system.
This is delicious fodder for a post of its own. There’s a mass of papers about the relationship between autism and the immune system; N3s being neuroprotective and correlating negatively with likelihood of autism/severity of autism symptoms vindicates the idea that autism happens when your immune system cooks your brain (neuroinflammation). You would expect males to be whacked harder by this because they’re not good at producing the important N3 docosahexaenoic acid (DHA). Thus, males ought to have higher autism rates- and they do.
Anyway, here’s what I’m getting at. Either N3s are a counter-intuitive miracle drug, or they’re just an important nutrient of which many of us do not get enough. The latter, I should think! The lemonade that can and will be made from these lemons is that widespread N3 deficiency gives us an opportunity to understand one of the ways that a human brain can get messed up. But we still want to fix the problem- more on that in my next post.
Our ancestors probably got more N3s than we do. They’ve been estimated to, anyway. (Second study mentioning our ancestors’ higher N3 intake.) If they didn’t, they might’ve been selected for better processing of alpha linoleic acid into eicosapentaenoic acid and subsequently docosahexaenoic acid, which real-life humans aren’t great at. As it stands, humans need either a whole bunch of excess ALA to convert to EPA and DHA, or we could have EPA and DHA straight. The excess ALA idea probably isn’t something you can rely on for your N3 needs, but certainly ALAs are better than nothing.
Seeds would’ve been a part of the diet back in the Paleolithic. Flax, an example of a seed rich in ALA, was known to humans quite a darned while ago, so seeds might have been a source of these critical nutrients. It’s worth noting that the first paleolithic diet estimation study linked in the previous paragraph pegs hunter gatherers as getting way more ALA than modern folks do.
Of course, many populations would’ve lived near oceans or at least rivers and lakes, where they could’ve gotten N3s from fish and seafood.
Some of it must have come from insects, themselves actually having a decent amount of N3s.
I’ve got an even weirder guess, though: bioaccumulation in land animal tissue. This possibility must have been important for peoples like the Botai culture with extremely narrow (Tarpan-based) diets, which likely didn’t include enough seeds, insects, and/or seafood. Animals can’t produce N3s of their own, but they eat ALA, convert it into EPA and DHA, and send a lot of it up to the brain, where it’s needed, with the result being that the brain has a lot more and higher quality N3s than any of the plants from which the N3s were derived.
Grog the caveman could’ve gotten a bunch of N3s from his favorite treat: scrambled auroch brains. Many of his progeny carry the practice into the present day, to the disgust of other living humans. Being that humans and our relatives are disgusting in general, Neanderthals occasionally even ate each other’s brains, and according to CNN, they’re still at it!
Nonhuman brains had to be the more popular option, then as now; mentioning the whole cannibalism thing is a bit of a non sequitur I shoved in for sheer entertainment value. But I suspect that there are several reasons we Westerners don’t get enough N3s: we don’t eat brains anymore, we don’t eat bugs anymore, and we don’t get as many ALA-rich plant oils as we used to. Perhaps we’re not getting enough fish, or perhaps the fish we’re getting are less likely to be fatty (e.g. salmonids, tuna, schooling fish) and more likely to be lean (cod, pollock).
Somehow or other we ought to get N3s back in our diet. Could save some kids from autism or, failing that, improve their prognosis. Again, it could increase our math scores, too, particularly in women. Closing part of the gap with other nations and between our sexes- two birds with one stone for our educators- and a boon for our engineering departments as well. Which is a boon for everyone.
The stakes are high. Who can save us from our collective state of starved brains? I’ll post my ideas soon.
Race, Testosterone, and Prostate Cancer
1900 words
I have explicitly shown how the ‘black men have more T’ canard is false. I have provided sufficient evidence for this claim. People claim to be unbiased—like PumpkinPerson—and say they’re only ‘looking for the truth’. However, it’s clear that in all of my conversations with him on the matter, he’s reaching for anything that affirms his worldview when he is shown evidence to the contrary of his beliefs (Nyhan and Reifler, 2012). It’s so very easy to notice this. I will provide yet more robust data on the black/white testosterone ‘gap’ while also critiquing some other studies that didn’t control for some pretty important variables. The gist is: after controlling for the most important variables, (waist circumference, BMI) the ‘testosterone difference’ all but disappears—and I don’t think people will argue for .0068 ng/ml higher testosterone cause things like prostate cancer and higher rates of crime.
Gapstur et al (2002) studied 5,115 individuals from aged 18-30 who completed baseline examinations at one of four locations from 1985-86: Birmingham, Alabama; Chicago, Illinois; Minneapolis, Minnesota; and Oakland, California. Then, 4 follow-ups were completed (Year 2: 1987-88; year 5: 1990-91; year 7: 1992-93; and year 10: 1995-96).
The number of blacks who completed the baseline information was 1157 whereas for whites it was 1171. They measured waist circumference (WC) at the minimal abdominal girth. Whether or not one took medication was self-reported, so Gapstur et al (2002) separated medications into two categories: regulation or interfering with binding likely, or interfering with binding unlikely/impossible.
Now, before I discuss the results of Gapstur et al (2002) I must talk about why they controlled for BMI (body mass index) and WC along with age. Since the obesity rate differs by race/ethnicity, along with obesity frequently changing with age, all three of the variables need to be controlled for to get a clearer picture of what circulating testosterone looks like—on average—between blacks and whites. White men are slightly more likely than black men to be obese/overweight in America (being African-American seems to be a protector against obesity; African American men with more African ancestry are less likely to be obese), however the sample used by Gapstur et al (2002) had blacks who had a higher WC and BMI than whites.
At year 2, there was no significant difference in total serum testosterone between the races with or without adjustment for age, BMI and WC. The only part in this analysis that blacks had higher testosterone levels than whites was at year ten, with black ‘enjoying’ .0063 ng/ml higher levels than whites. Furthermore, there was no significant difference in testosterone between blacks and whites after adjustment for age. Only adjusting for BMI, blacks had substantially higher levels of testosterone (.21 ng/ml) however after including WC and the changes in WC between blacks and whites (blacks had a greater change over the study) this difference all but disappeared. The age-associated changes in testosterone between blacks and whites were similar after adjusting for waist circumference and BMI. Also, after adjusting for the relevant confounds, free testosterone did not differ between blacks and whites.
Measures of body size (i.e., BMI and WC) must be controlled for when comparing race/ethny since levels of obesity vary amongst them as well as obesity constantly changing with age. Furthermore, testosterone increased for both groups between the ages of 20-21 to 22-23 (blacks had a higher T level by .7 ng/ml at age 20-21), with the two groups diverging at age 22-23 with blacks having higher levels of total testosterone by .1 ng/ml.
The average ages for the cohort were 28.4 and 28.8 for blacks and whites respectfully. Thus, blacks (theoretically) had a slight advantage due to the age confound, which had to be controlled for. The unadjusted mean total testosterone, SHGB, and free testosterone were not statistically significant at any point in the study except at year 10 where blacks had slightly higher levels at 5.8 ng/ml compared to whites’ 5.69 ng/ml. The difference in year 2 (when testosterone levels raised for both groups) was a difference of 5.8 and 5.75 for blacks and whites respectfully. Free testosterone did not differ at all from years 2, 7, and 10 (.17 compared to .17 in year 2; .16 to .15 in year 7; and .16 to .15 in year 10; blacks and whites respectfully).
Levels of testosterone were also found to be lower in 12-13-year-old blacks compared to whites (Lopez et al, 2013). In this study, Mexican Americans had higher T levels, while after adjustments for confounds, blacks and whites did not differ. They conclude that testosterone levels were not higher in black compared to white adolescents. Black men have higher levels of estradiol than white men, not testosterone (Roerrmann et al, 2007; Lopez et al, 2013). These studies are done to see the relationship between PCa (prostate cancer) and certain hormones. These and other studies have shown that the black-white difference isn’t large (Richard et al, 1992; Roerrmann et al, 2007; Lopez et al, 2013; Richard et al, 2014) and that prostate cancer is not caused by abnormally high testosterone levels (Stattin et al, 2003; Michaud, Billups, and Partins, 2015).
Ross et al (1986) did not have a measure of central adiposity (WC), thusly the results were confounded. Further, the other study Rushton cited in Race, Evolution, and Behavior is and Nyborg (1992) on discharged army veterans stating a difference of 3 percent, Ellis and Nyborg did not control for WC nor BMI. As seen in the Gapstur et al (2002) study, WC is an extremely important variable to control for as decreases in testosterone are high when central adiposity is high (Wang et al, 2011) so if we are trying to compare two ethnies on one variable such as testosterone, all of the above variables MUST be controlled for, and if they are not then they can be safely disregarded.
The non-inclusion of a measure of WC is the most likely cause of the different results found in Gapstur et al (2002). If you don’t control for WC, then you cannot get an actual and reliable testosterone reading since the results will be confounded. Anyone who says otherwise that the aforementioned variables do not need to be controlled for literally have no idea what they’re talking about.
Lastly, a few more things must be addressed. In PumpkinPerson’s most recent article “Racial differences in testosterone“, he seems to now disavow testosterone as a useful measure since it can fluctuate due to winning sports games, to marriage, to anticipating confrontation, to being in a relationship, to honor culture. Now his thing is exposure to androgens (testosterone) in the womb. Supposedly, blacks have the lowest digit ratio, and low digit ratios signify higher levels of testosterone in vitro (Manning et al, 2004). However, the relationship is far from proven (Koehler, Simmons, and Rhodes, 2004; Yan et al, 2008, Medland et al, 2010) with these results, leaning towards no. Koheler, Simmons, and Rhodes (2004) failed to find any significant correlation between 2d/4d ratio and traits mediated by testosterone. So this is another claim that’s been put to bed as well.
There is a lot of bullshit to sift through out there; some things may seem simple at face value, but if you dig into it, it’s much more likely to be more complex than what it looks to be on the surface. This one singular variable (testosterone) is one of them. It does not differ between the races; exposure to androgens in the womb doesn’t affect digit ratio; high testosterone does not cause prostate cancer, but low testosterone does (Morgentaler, Brunning, and DeWolf, 1996) ; you can literally inject exogenous testosterone into a man with PCa and not effect this malady (Eisenberg, 2015; Boyle et al, 2016)
I hope this is the last time I have to say this: testosterone does not differ between blacks and whites. Testosterone is not the cause of differences in mortality rate in regards to PCa. (Diet is the much more likely factor) People still regurgitate Rushton’s bullshit from REB; Ross et al (1986) still gets cited to this day, when there are much more robust studies and samples that controlled for the relevant confounds that Ross et al (1986) did not control for. This is why that study is garbage and should not be looked at when assessing testosterone differences between the races. Much larger and robust samples show that there is no testosterone difference when the WC is controlled for.
Testosterone is not higher in black Americans; the data shows either an extremely negligible difference or no difference, not the huge 13 and 21 percent difference in free and total testosterone in black Americans in Ross et al (1986).
For the last time, and say it with me: testosterone does NOT differ by race, 2d:4d ratio is NOT influenced by androgens in vitro, and testosterone does NOT influence PCa rates. Testosterone is an extremely important hormone for vital functioning, so whoever believes the myths of the high testosterone savage and LIKES having low testosterone, have fun with a slew of maladies later in life.
References
Boyle, P., Koechlin, A., Bota, M., Donofrio, A., Zaridze, D. G., Perrin, P., . . . Boniol, M. (2016). Endogenous and exogenous testosterone and the risk of prostate cancer and increased prostate-specific antigen (PSA) level: a meta-analysis. BJU International,118(5), 731-741. doi:10.1111/bju.13417
Dobs, A., & Morgentaler, A. (2008). Does Testosterone Therapy Increase the Risk of Prostate Cancer? Endocrine Practice,14(7), 904-911. doi:10.4158/ep.14.7.904
Eisenberg, M. L. (2015). Testosterone Replacement Therapy and Prostate Cancer Incidence. The World Journal of Men’s Health, 33(3), 125–129. http://doi.org/10.5534/wjmh.2015.33.3.125
Gapstur, S. M., Kopp, P., Gann, P. H., Chiu, B. C., Colangelo, L. A., & Liu, K. (2006). Changes in BMI modulate age-associated changes in sex hormone binding globulin and total testosterone, but not bioavailable testosterone in young adult men: the CARDIA Male Hormone Study. International Journal of Obesity. doi:10.1038/sj.ijo.0803465
Koehler, N., Simmons, L. W., & Rhodes, G. (2004). How well does second-to-fourth-digit ratio in hands correlate with other indications of masculinity in males? Proceedings of the Royal Society B: Biological Sciences,271(Suppl_5). doi:10.1098/rsbl.2004.0163
Lopez, D. S., Peskoe, S. B., Joshu, C. E., Dobs, A., Feinleib, M., Kanarek, N., . . . Platz, E. A. (2013). Racial/ethnic differences in serum sex steroid hormone concentrations in US adolescent males. Cancer Causes & Control,24(4), 817-826. doi:10.1007/s10552-013-0154-8
Michaud, J. E., Billups, K. L., & Partin, A. W. (2015). Testosterone and prostate cancer: an evidence-based review of pathogenesis and oncologic risk. Therapeutic Advances in Urology,7(6), 378-387. doi:10.1177/1756287215597633
Medland, S. E., Zayats, T., Glaser, B., Nyholt, D. R., Gordon, S. D., Wright, M. J., . . . Evans, D. M. (2010). A Variant in LIN28B Is Associated with 2D:4D Finger-Length Ratio, a Putative Retrospective Biomarker of Prenatal Testosterone Exposure. The American Journal of Human Genetics,86(4), 519-525. doi:10.1016/j.ajhg.2010.02.017
Manning, J., Stewart, A., Bundred, P., & Trivers, R. (2004). Sex and ethnic differences in 2nd to 4th digit ratio of children. Early Human Development,80(2), 161-168. doi:10.1016/j.earlhumdev.2004.06.004
Morgentaler, A., Brunning, C. O., 3rd, & DeWolf, W. C. (1996). Occult Prostate Cancer in Men With Low Serum Testosterone Levels. JAMA: The Journal of the American Medical Association,276(23), 1904. doi:10.1001/jama.1996.03540230054035
Nyhan, B., & Reifler, J. (2010). When Corrections Fail: The Persistence of Political Misperceptions. Political Behavior,32(2), 303-330. doi:10.1007/s11109-010-9112-2
Richard, A., Rohrmann, S., Zhang, L., Eichholzer, M., Basaria, S., Selvin, E., . . . Platz, E. A. (2014). Racial variation in sex steroid hormone concentration in black and white men: a meta-analysis. Andrology,2(3), 428-435. doi:10.1111/j.2047-2927.2014.00206.x
Rohrmann, S., Nelson, W. G., Rifai, N., Brown, T. R., Dobs, A., Kanarek, N., . . . Platz, E. A. (2007). Serum Estrogen, But Not Testosterone, Levels Differ between Black and White Men in a Nationally Representative Sample of Americans. The Journal of Clinical Endocrinology & Metabolism,92(7), 2519-2525. doi:10.1210/jc.2007-0028
Wang, C., Jackson, G., Jones, T. H., Matsumoto, A. M., Nehra, A., Perelman, M. A., … Cunningham, G. (2011). Low Testosterone Associated With Obesity and the Metabolic Syndrome Contributes to Sexual Dysfunction and Cardiovascular Disease Risk in Men With Type 2 Diabetes. Diabetes Care, 34(7), 1669–1675. http://doi.org/10.2337/dc10-2339
Yan RHY, Malisch JL, Hannon RM, Hurd PL, Garland T Jr (2008) Selective Breeding for a Behavioral Trait Changes Digit Ratio. PLoS ONE 3(9): e3216. https://doi.org/10.1371/journal.pone.0003216
Dinosaurs, Brains, and ‘Progressive’ Evolution: Part II
1700 words
In part I, I showed how Dale Russel’s contention that the troodon would have evolved into a bipedal ‘dinosauroid’ with human locomotion and a human-sized brain was pure fantasy. I ordered the book of his that Rushton cited in his book Race, Evolution, and Behavior and I finally received it last week. When I read the relevant parts, I yawned because it’s the same old stuff that I’ve covered here on this blog numerous times. Since literally the only relevant part in the book about the troodon is the final 7 pages, that’s what I will cover today—along with a few more lines of evidence that large brains lie outside reptilian design (Gould, 1989).
First off, all of Rushton’s contentions in the final pages of his book (Rushton, 1997) need to be rebutted. Rushton (1997: 294) writes that dinosaur brains were ‘progressing’ in size for 140 million years, but neither of Russel’s writings that I have (Russel 1983; 1989) have the statement in them.
In the book Up From Dragons: The Evolution of Human Intelligence neuroscientist, evolutionary psychologist John Skoyles and science writer Dorian Sagan—the son of Carl Sagan—speak briefly about reptilian intelligence and why they wouldn’t have reached our levels of intellect:
But cold-bloodedness is a dead-end for the great story of this book—the evolution of intelligence. Certainly reptiles could evolve huge sizes, as they did over vast sweeps of Earth as dinosaurs. But they never could have evolved our quick-witted and smart brains. Being tied to the sun restricts their behavior: Instead of being free and active, searching and understanding the world, they spend too much time avoiding getting too hot or too cold. (Skoyles and Sagan, 2002: 12)
Hopson (1977: 443) writes:
I would argue, as does Feduccia (44), that the mammalian/avian levels of activity claimed by Bakker for dinosaurs should be correlated with a great increase in motor and sensory control and this should be reflected in increased brain size. Such an increase is not indicated by most dinosaur endocasts.
Most importantly, if some dinosaurs DID have bird-sized brains, the above contention would still hold. Hopson concludes that, except for coelurosaurs “the range of behaviors that existed in dinosaurs, as inferred from trackways and skeletal morphology, may not have lain much outside the observed range in ectothermic crocodilians” (Hopson, 1977: 444).
Since the conjecture/’thought experiment’ of the troodon was rebutted last week, it’s pretty conclusive that large brains lie outside of reptilian design; they need to spend so much time avoiding getting too hot or cold—as well as hunt and eat—so exploring the world and learning was not possible for them—along with the fact that they didn’t have a primate morphology and thus didn’t have the ability to fully manipulate their environment as we do which would further select for larger brains. However, as Hopson (1977) notes, animals with higher metabolic rates had larger brains; coelurosaurs had high metabolic rates and the largest dinosaur brains (Russel, 1983; 1989)—but that doesn’t mean they would have eventually evolved human-like intelligence, bipedalism or brain size and to say otherwise is fantasy.
Furthermore, there is large variation in encephalization and encephalization is not universal in mammals (Shultz and Dunbar, 2010).
Here is the thing about brain size increases: it is a local level trend. A local level trend is a trend that occurs within one or a few related species. This is exactly what characterized brain evolution; there is large variation depending on what the environment calls for (Boddy et al, 2012; Montgomery et al, 2012; see also island gigantism; Bromham and Cardillo, 2007; Welch, 2009; and also see the deep sea rule; Mcclain, Boyer, and Rosenberg, 2006). So these local trends differ by species—even one population split by, say, 50 miles of water will speciate and become evolve a completely different phenotype due to the environment of time. That is evolution by natural selection; local change, not any inherent or intrinsic ‘progress’ (Gould, 1996).
The same local level trend occurs with parasites. Now think about parasites. The get selected for ‘complexity’ or a decrease in ‘complexity’ depending on what occurs in their host. Now, looking at it from this perspective, the body is the host’s environment while the earth is ours; so my example for an environmental change would be, as usual, the asteroid impact hitting the earth blocking out the sun and decreasing high-quality food all throughout the earth. Surely I don’t need to tell you what would occur…
Russel (1989) writes:
Examples of evolutionary changes that occured at ever-increasing speeds include the initial diversification of animals in the sea 650 and 550 million years ago, the attainment of tree stature in land plants between 410 and 360 million years ago, and the diversification of mammals between 200 million years ago and the present. Changes like this have resulted in increased organismal complexity, which, in combination with a general increase in number of species, has made the biosphere of the modern Earth so much richer than it was several hundred million years ago. It is reasonable to suppose that animals living in a complex environment might find it advantageous to possess complex nervous systems in order to have access to a greater variety of responses. Indeed, the largest proportion of brain weight in an animal has also increased at an ever-increasing rate across geological time. The brain has become evidently larger in animals as diverse as insects, mollusks, and backboned creatures. Relative brain size can be taken as an indication of biotic interactions.
He references time periods that correspond with decimations (mass extinctions). Decimations lead to diversification. Think back to the Cambrian Explosion. During the Cambrian Explosion, many more lifeforms existed than can be currently classified. Therefore, according to the decimation and diversification model, greater diversity of life existed in the past. When decimations (defined as a reduction in the anatomical forms of life from mass extinction) occur, the niches that become extinct quickly become filled.
The time periods that Russel references are when mass extinctions occurred. This is how diversification occurs. What allowed for this ‘organismal complexity’ and increase in the number of species (though body plans are limited due to the Burgess Decimation) is due to the decimations. Decimation and diversification proves that evolution is not progressive.
A ‘trend’ in biology is directional change in a group stat using the mean, median or mode. Any existence of a trend from the mean (‘progress’) tells us nothing about the underlying mechanisms behind it.
To wrap this all up, even if a trend in X were to be discovered, it still wouldn’t tell us a thing about the underlying mechanisms causing it, nor will it tell us about any increasing tendency.
The analogy of the drunkard’s walk (Gould, 1996) is why ‘progress’ doesn’t make sense. Further, niche construction matters as well. When organisms construct their own niches, change occurs based on those niche constructions. Milk-drinking 8kya in Europe and African farmers diverting water for their crops having mosquitoes come by and gaining a resistance to malaria are two examples of niche construction (Laland et al, 2009). That’s another barrier to progress!
In sum, Dale Russel says nothing I’ve not heard before in regards to ‘progressive’ evolution. He only describes ever-increasing ‘complexity’ which is due to decimations and further diversification by organisms to fill empty niches. Any type of ‘progress’ would have been stymied by mass extinctions.
Further, the fact that species can consciously—in a way—guide their own evolution through the manipulation of the environment once again shows how evolution doesn’t mean progress—it literally only means local change and any type of local change, no matter to what type of environment, will cause concurrent increases/decreases on whichever relevant traits that will give the organism the best chance for survival in that environment.
This is why evolution is not progressive; and even if scientists were to identify one thing, still, a causal mechanism won’t be able to be inferred. Ruseel (1989) describes right and left walls of complexity—nothing more. Dinosaurs didn’t have the body plans to have our brain size, bipedalism and intelligence, nor did they have the right type of blood, nor did they have the time to search and learn about the world due to being constrained to their cold-blooded system—being a slave to the sun, always attempting to avoid overheating or getting too cold (Skoyles and Sagan, 2002). The so-called ‘dinosauroid’ is an impossibility and implies a teleological lean to evolution—as if our morphology (or something similar from an unrelated organism) will always evolve if we replay the tape of life again (Gould, 1989; 1996). This is what Russel is pretty much arguing, and he is 100 percent wrong as noted above.
References
Bromham, L., & Cardillo, M. (2007). Primates follow the ‘island rule’: implications for interpreting Homo floresiensis. Biology Letters,3(4), 398-400. doi:10.1098/rsbl.2007.0113
Boddy, A. M., Mcgowen, M. R., Sherwood, C. C., Grossman, L. I., Goodman, M., & Wildman, D. E. (2012). Comparative analysis of encephalization in mammals reveals relaxed constraints on anthropoid primate and cetacean brain scaling. Journal of Evolutionary Biology,25(5), 981-994. doi:10.1111/j.1420-9101.2012.02491.x
Gould, S. J. (1989). Wonderful life: the Burgess Shale and the nature of history. New York: Norton.
Gould, S. J. (1996). Full House: The Spread of Excellence from Plato to Darwin. New York: Harmony Books.
Hopson, J. A. (1977). Relative Brain Size and Behavior in Archosaurian Reptiles. Annual Review of Ecology and Systematics,8(1), 429-448. doi:10.1146/annurev.es.08.110177.002241
Laland, K. N., Odling-Smee, J., Feldman, M. W., & Kendal, J. (2009). Conceptual Barriers to Progress Within Evolutionary Biology. Foundations of Science, 14(3), 195–216. http://doi.org/10.1007/s10699-008-9153-8
Mcclain, C. R., Boyer, A. G., & Rosenberg, G. (2006). The island rule and the evolution of body size in the deep sea. Journal of Biogeography,33(9), 1578-1584. doi:10.1111/j.1365-2699.2006.01545.x
Montgomery, S. H., Capellini, I., Barton, R. A., & Mundy, N. I. (2010). Reconstructing the ups and downs of primate brain evolution: implications for adaptive hypotheses and Homo floresiensis. BMC Biology,8(1), 9. doi:10.1186/1741-7007-8-9
Russell, D. A. (1983). Exponential evolution: Implications for intelligent extraterrestrial life. Advances in Space Research,3(9), 95-103. doi:10.1016/0273-1177(83)90045-5
Russell, D. A. (1989). An Odyssey in Time: The Dinosaurs of North America. Minocqua, WI: Published by NorthWord Press in association with National Museum of Natural Sciences.
Rushton J P (1997). Race, Evolution, and Behavior. A Life History Perspective (Transaction, New Brunswick, London).
Shultz, S., & Dunbar, R. (2010). Encephalization is not a universal macroevolutionary phenomenon in mammals but is associated with sociality. Proceedings of the National Academy of Sciences,107(50), 21582-21586. doi:10.1073/pnas.1005246107
Skoyles, J. R., & Sagan, D. (2002). Up From Dragons: The Evolution of Human Intelligence. New York: McGraw-Hill.
Welch, J. J. (2009). Testing the island rule: primates as a case study. Proceedings of the Royal Society B: Biological Sciences,276(1657), 675-682. doi:10.1098/rspb.2008.1180
