
Home » 2018
Yearly Archives: 2018
Muscle Fibers, Obesity, Cardiometabolic Disorders, and Race
2650 words
The association between muscle fiber typing obesity and race is striking. It is well-established that blacks have a higher proportion of type II skeletal muscle fibers than whites and these higher proportions of these specific types of muscle fibers lead to physiological differences between the two races which then lead to differing health outcomes between them—along with differences in athletic competition. Racial differences in health are no doubt complex, but there are certain differences between the races that we can look at and say that there is a relationship here that warrants further scrutiny.
Why is there an association between negative health outcomes and muscle phsyiology? The answer is very simple if one knows the basics of muscle physiology and how and why muscles contract (it is worth noting that out of a slew of anatomic and phsyiologic factors, movement is the only thing we can consciously control, compare to menstration and other similar physiologic processes which are beyond our control). In this article, I will describe what muscles do, how they are controlled, muscle physiology, the differences in fiber typing between the races and what it means for health outcomes between them.
Muscle anatomy and physiology
Muscle fiber number is determined by the second trimester. Bell (1980) noted that skeletal muscle fiber in 6 year olds is not different from normal adult tissue, and so, we can say that between the time in the womb and age 6, muscle fiber type is set and cannot be changed (though training can change how certain fibers respond, see below).
Muscle anatomy and physiology is interesting because it shows us how and why we move the way we do. Tendons attach muscle to bone. Attached to the tendon is the muscle belly. The muscle belly is made up of facsicles and the fascicles are made up of muscle fibers. Muscle fibers are made up of myofibrils and myofibrils are made up of myofilaments. Finally, myofilaments are made up of proteins—specifically actin and myosin, this is what makes up our muscles.

(Image from here.)
Muscle fibers are encased by sarcolemma which contains cell components such as sarcoplasm, nuclei, and mitochondria. They also have other cells called myofibrils which contain myofilaments which are then made up of actin (thin filaments) and mysoin (thick filaments). These two types of filaments form numerous repeating sections within a myofibril and each repeating section is known as a sarcomere. Sarcomeres are the “functional” unit of the muscle, like the neuron is for the nervous system. Each ‘z-line’ denotes another sarcomere across a myofibril (Franzini-Armstrong, 1973; Luther, 2009).
Other than actin and myosin, there are two more proteins important for muscle contraction: tropomyosin and troponin. Tropomyosin is found on the actin filament and it blocks myosin binding sites which are located on the actin filament, and so it keeps myosin from attaching to muscle while it is in a relaxed state. On the other hand, troponin is also located on the actin filament but troponin’s job is to provide binding sites for calcium and tropomyosin when a muscle needs to contract.
So the structure of skeletal muscle can be broken down like so: epymyseum > muscle belly > perimyseum > fascicle > endomyseum > muscle fibers > myofibrils > myofilaments > myosin and actin. Note diagram (C) from above; the sarcomere is the smallest contractile unit in the myofibril. According to sliding filament theory (see Cook, 2004 for a review), a sarcomere shortens as a result of the ‘z-lines’ moving closer together. The reason these ‘z-lines’ converge is because myosin heads attach to the actin filament which asynchronistically pulls the actin filament across the myosin, which then results in the shortening of the muscle fiber. Sarcomeres are the basic unit controlling changes in muscle length, so the faster or slower they fire depends on the majority type of fiber in that specific area.
But the skeletal muscle will not contract unless the skeletal muscles are stimulated. The nervous system and the muscular system communicate, which is called neural activiation—defined as the contraction of muscle generated by neural stimulation. We have what are called “motor neurons”—neurons located in the CNS (central nervous system) which can send impulses to muscles to move them. This is done through a special synapse called the neuromuscular junction. A motor neuron that connects with muscle fibers is called a motor unit and the point where the muscle fiber and motor unit meet is callled the neuromuscular junction. It is a small gap between the nerve and muscle fiber called a synapse. Action potentials (electrical impulses) are sent down the axon of the motor neuron from the CNS and when the action potential reaches the end of the axon, hormones called neurotransmitters are then released. Neurotransmitters transport the electrical signal from the nerve to the muscle.
Muscle fiber types
The two main categories of muscle fiber are type I and type II—‘slow’ and ‘fast’ twitch, respectively. Type I fibers contain more blood cappilaries, higher levels of mitochondria (which transforms food into ATP) and myoglobin which allows for an improved delivery of oxygen. Since myoglobin is similar to hemoglobin (the red pigment which is found in red blood cells), type I fibers are also known as ‘red fibers.’ Type I fibers are also smaller in diameter and slower to produce maximal tension, but are also the most fatigue-resistant type of fiber.
Type II fibers have two subdivisions—IIa and IIx—based on their mechanical and chemical properties. Type II fibers are in many ways the opposite of type I fibers—they contain far fewer blood cappilaries, mitochondria and myoglobin. Since they have less myoglobin, they are not red, but white, which is why they are known as ‘white fibers.’ IIx fibers have a lower oxidative capacity and thusly tire out quicker. IIa, on the other hand, have a higher oxidative capacity and fatigue slower than IIx fibers (Herbison, Jaweed, and Ditunno, 1982; Tellis et al, 2012). IIa fibers are also known as intermediate fast twitch fibers since they can use both anarobic and aerobic metabolism equally to produce energy. So IIx fibers are a combo of I and II fibers. Type II fibers are bigger, quicker to produce maximal tension, and tire out quicker.
Now, when it comes to fiber typing between the races, blacks have a higher proportion of type II fibers compared to whites who have a higher proportion of type I fibers (Ama et al, 1986; Ceaser and Hunter, 2015; see Entine, 2000 and Epstein, 2014 for reviews). Higher proportions of type I fibers are associated with lower chance of cardiovascular events, whereas type II fibers are associated with a higher risk. Thus, “Skeletal muscle fibre composition may be a mediator of the protective effects of exercise against cardiovascular disease” (Andersen et al, 2015).
Now that the basics of muscle anatomy and physiology are apparent, hopefully the hows and whys of muscle contraction and what different muscle fibers do are becoming clear, because these different fibers are distributed between the races in uneven frequencies, which then leads to differences in sporting performance but also differents in health outcomes.
Muscle fibers and health outcomes
We now know the physiology and anatomy of muscle and muscle fiber typing. We also know the differences between each type of skeletal muscle fiber. Since the two races do indeed differ in the percentage of skeletal muscle fiber possessed on average, we then should find stark differences in health outcomes, part of the reason being these differences in muscle fiber typing.
While blacks on average have a higher proportion of type II muscle fibers, whites have a higher proportion of type I muscle fibers. Noting what I wrote above about the differences between the fiber types, and knowing what we know about racial differences in disease outcomes, we can draw some inferences on how differences in muscle fiber typing between races/individuals can then affect disease seriousness/acquisition.
In their review of black-white differences in muscle fiber typing, Ceaser and Hunter (2015) write that “The longitudinal data regarding the rise in obesity indicates obesity rates have been highest among non-Hispanic Black women and Hispanic women.” And so, knowing what we know about fiber type differences between races and how these fibers act when they fire, we can see how muscle fiber typing would contribute to differences in disease acquisition between groups.
Tanner et al (2001) studied 53 women (n=28, lean women; and n=25, obese women) who were undergoing an elective abdominal surgery (either a hysterectomy or gastric bypass). Their physiologic/anatomic measures were taken and they were divided into races: blacks and whites, along with their obesity status. Tanner et al found that the lean subjects had a higher proportion of type I fibers and a lower proportion of type IIx fibers whereas those who were obese were more likely to have a higher proportion of type IIb muscle fibers.
Like other analyses on this matter, Tanner et al (2001) showed that the black subjects had a higher proportion of type II fibers in comparison to whites who had a higher proportion of type I fibers (adiposity was not taken into account). Fifty-one percent of the fiber typing from whites was type I whereas for blacks it was 43.7 pervent. Blacks had a higher proportion of type IIx fibers than whites (16.3 percent for whites and 23.4 for blacks). Lean blacks and lean whites, though, had a similar percentage of type IIx fibers (13.8 percent for whites and 15 percent for blacks). It is interesting to note that there was no difference in type I fibers between lean whites and blacks (55.1 percent for whites and 54.1 percent for blacks), though muscle fibers from obese blacks contained far fewer type I fibers compared to their white counterparts (48.6 percent for whites and 34.5 for blacks). Obese blacks’ muscle fiber had a higher proportion of type IIx fibers than obese whites’ fiber typing (19.2 percent for whites and 31 percent for blacks). Lean blacks and lean whites had a higher proportion of type I fibers than obese blacks and obese whites. Obese whites and obese blacks had more type IIx fibers than lean whites and lean blacks.
So, since type II fibers are insulin resistant (Jensen et al, 2007), then they should be related to glucose intloerance—type II diabetes—and blacks with ancestry from West Africa should be most affected. Fung (2016, 2018) shows that obesity is a disease of insulin resistance, and so, we can bring that same rationale to racial differences in obesity. Indeed, Nielsen and Christensen (2011) hypothesize that the higher prevalence of glucose intolerance in blacks is related to their lower percentage of type I fibers and their higher percentage of type II fibers.
Nielsen and Christensen (2011) hypothesize that since blacks have a lower percentage of type I fibers (the oxidative type), this explains the lower fat oxidation along with lower resting metabolic rate, sleeping metabolic rate, resting energy expenditure and Vo2 max in comparison to whites. Since type I fibers are more oxidative over the glycolitic type II fibers, the lower oxidative capacity in these fibers “may cause a higher fat storage at lower levels of energy intake than in individuals with a higher oxidative capacity” (Nielsen and Christensen, 2011: 611). Though the ratio of IIx and IIa fibers are extremely plastic and affected by lifestyle, Nielsen and Christensen do note that individuals with different fiber typings had similar oxidative capacity if they engaged in physical activity. Recall back to Caesar and Hunter (2015) who note that blacks have a lower maximal aerobic capacity and higher proportion of type II fibers. They note that lack of physical activity exacerbates the negative effects that a majority type II fibers has over majority type I. And so, some of these differences can be ameliorated between these two racial groups.
The point is, individuals/groups with a higher percentage of type II fibers who do not engage in physical activity have an even higher risk of lower oxidative capacity. Furthermore, a higher proportion of type II fibers implies a higher percentage of IIx fibers, “which are the least oxidative fibres and are positively associated with T2D and obesity” (Nielsen and Christensen, 2011: 612). They also note that this may explain the rural-urban difference in diabetes prevalance, with urban populations having a higher proportion of type II diabetics. They also note that this may explain the difference in type II diabetes in US blacks and West African natives—but the reverse is true for West Africans in the US. There is a higher rate of modernization and, with that, a higher chance to be less physically active and if the individual in question is less physically active and has a higher proportion of type II fibers then they will have a higher chance of acquiring metabolic diseases (obesity is also a metabolic disease). Since whites have a higher proportion of type I fibers, they can increase their fat intake—and with it, their fat oxidation—but this does not hold for blacks who “may not adjust well to changes in fat intake” (Nielsen and Christensen, 2011: 612).
Nielsen and Christensen end their paper writing:
Thus, Blacks of West African ancestry might be genetically predisposed to T2D because of an inherited lower amount of skeletal muscle fibre type I, whereby the oxidative capacity and fat oxidation is reduced, causing increased muscular tissue fat accumulation. This might induce skeletal muscle insulin resistance followed by an induced stress on the insulin-producing beta cells. Together with higher beta-cell dysfunction in the West African Diaspora compared to Whites, this will eventually lead to T2D (an overview of the ‘skeletal muscle distribution hypothesis’ can be seen in Figure 2).
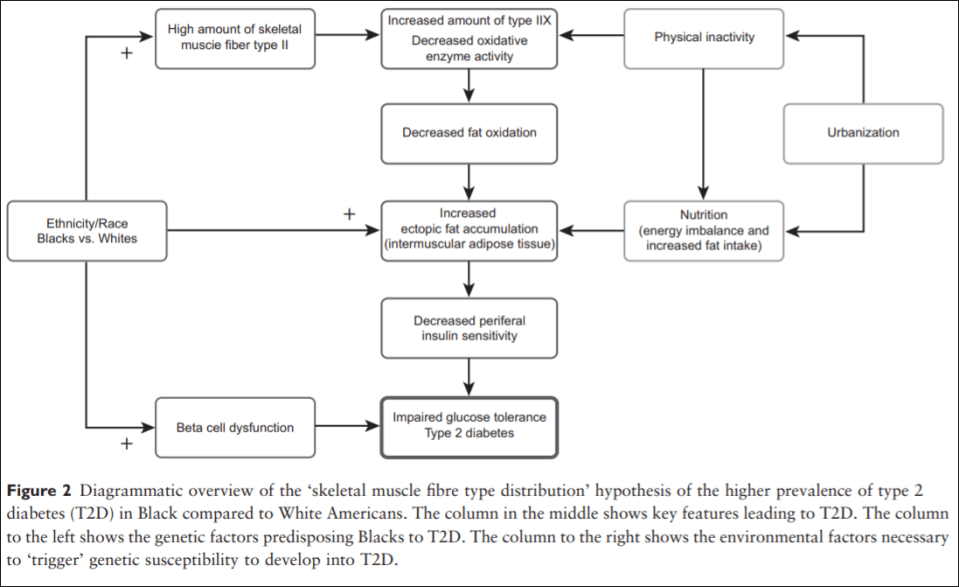
Lambernd et al (2012) show that muscle contractions eliminated insuin resistance by blocking pro-inflammatory signalling pathways: this is the mechanism by which physical activity decreases glucose intolerance and thusly improves health outcomes—especially for those with a higher proportion of type II fibers. Thus, it is important for individuals with type II fibers to exercise, since sedentariness is associated with an age-related insulin resistance due to impaired GLUT4 utilization (Bunprajun et al, 2013).
(Also see Morrison and Cooper’s (2006) hypothesis that “reduced oxygen-carrying capacity induced a shift to more explosive muscle properties” (Epstein, 2014: 179). Epstein notes that the only science there is on this hypothesis is one mouse and rat study showing that low hemoglobin can “induce a switch to more explosive muscle fibers” (Epstein, 2014: 178), but this has not been tested on humans to see if it would hold. If this is tested on humans and if it does hold, then that would lend credence to Morrison’s and Cooper’s (2006) hypothesis.)
Conclusion
Knowing what we know about muscle anatomy and physiology and how muscles act we can understand the influence the different muscle types have on disease and how they contribute to disease variation between race, sex and the individual level. Especially knowing how type II fibers act when the individual in question is insulin resistant is extremely important—though it has been noted that individuals who participate in aerobic exercise decrease their risk for cardiometabolic diseases and can change the fiber distribution difference between IIx and IIa fibers, lowering their risk for acquiring cardiometabolic diseases (Ceaser and Hunter, 2015).
Thinking back to sarcomeres (the smallest contractile unit in the muscle) and how they would act in type II fibers: they would obviously contract much faster in type II muscles over type I muscles; they would then obviously tear faster than type I muscles; since type II muscles are more likely to be insulin resistant, then those with a higher proportion of type II fibers need to focus more on aerobic activity to “balance out” type IIx and IIa fibers and decrease the risk of cardiometabolic disease due to more muscle contractions (Lambernd et al, 2012). Since blacks have a higher proportion of type II fibers and are more likely to be sedentary than whites, and since those who have a higher proportion of type II fibers are more likely to be obese, then it is clear that exercise can and will ameliorate some of the disparity in cardiometabolic diseases between blacks and whites.
Eugenics and the State
2450 words
The history of eugenics is intertwined with the history of genetics. The term “genetics” was established in 1905 by William Bateson. Half a century previously, Francis Galton coined the term “eugenics”, which is Greek for “well-born” or “good genesis”, though Galton did not attribute the genesis of the idea to the Greeks. Eugenic history began with the ancient Greeks, and, of course, is still around today. However, today we have technology that was previously not able to be conceptualized due to past people’s knowledge. This article will give a brief overview of eugenics throughout history up to the modern day.
Eugenics in ancient Greece
The idea of eugenics was first formulated by Aristotle. Aristotle imagined a rigged lottery (of course those who drew tickets did not know the lottery was rigged). The reasoning for rigging the lottery was simple: they could prevent so-called undesirable people from breeding while promoting births for people they want to breed. Plato only wanted those in the Greek upper-classes to mate (what was termed the “guardian class), men aged 25 and women aged 20—so they can birth children in their primes.
After the children were born, they were taken from their parents and held in special nurseries. Children who were Bron defective were” hidden away”, which could be a euphemism for infanticide (a form of which was reportedly practiced in Sparta where they left newborns at the city limits so the weak could die and the strong could live). Fit young men who chose not to breed had to pay a fine which went to the temple of Hera which was based on their class.
Aristotle had similar views to Plato. He, too, wanted the State to be in charge of the birthing process. He wanted both sexes to marry at their primes, in his eyes, aged 37 for men and 18 for women, while performing infanticide on babies born with deformities and aborting children if couples have too many. (See Galton, 1998 for a review of eugenic policies in ancient Greece.) (See also Genetics and the Decline of Sparta.)
The argument for autonomy
A simple argument can be erected against any and all State coercion to do something: the argument for the rejection of State authority (Wolff, 1970). The argument can be formulated thus:
P1. We have a higher-order interest in autonomy.
P2. If something promotes our higher-order interests, we have a duty to do it.
C1. We have a duty to be autonomous (modus ponens, P1, P2).
P3. If we have a duty to be autonomous, then autonomy requires that we decide what to do with ourselves.
C2. We should decide what to do for ourselves (modus ponens, C1, P3).
P4. If we accept the authority of others, then we are not autonomous.
C3. We should not accept the authority of others (modus tollens, C1, P4).
P5. If we accept the authority of law, then we accept the authority of others.
C4. We should not accept the authority of the law (modus tollens, C3, P5).
The argument is about rejecting State authority as a whole, but, of course, we can substitute. “If we accept the authority of [law X in regard to eugenic policy P] then we accept the authority of others[‘ thoughts of what to do with our lives]”. Thus we should disallow State eugenic policies.
Eugenics in ancient Rome
Like most other ancient civilizations, the ancient Romans practiced abortion and infanticide. Archaeological analyses uncovered a well, and in the well were dozens of little skeletons, implying that the Romans disposed of the babies of prostitutes (these were found in bordellos) because they were unwanted.
The Romans “deemed people with disabilities as sub-human“, with the abandonment of the baby being standard practice—which lends more credence to the baby wells of the Romans in the bordellos.
Eugenics in China
The Chinese Maternal and Infant Health Care Law states that if the screening of an embryo reveals that the couple in question has a chance of birthing a defective baby then they should agree to take contraceptive measures (see Guo, 2006 for a review). Further, if they already married then they must take long-term contraceptive measures. These measures are supported in China, since they have about 50 million disabled persons (see MacKeller and Bechtel, 2016).
Eugenics in Germany
The ideology of eugenics was present in Germany long before Hitler came to power. It was written about by numerous authors, including Nietzsche. “The first chair of eugenics was given in 1923 at the University of Munich to the geneticist Fritz Lenz” (MacKellar and Bechtel, 2016). Lenz promoted ‘racial hygeine’—he wanted Nordic traits to proliferate over others traits. He eventually joined the National Socialist party in 1937. The Nazis sterilized about 350,000 people between 1934 and 1939. This, then, led to the construction of a euthanasia program in 1939 called the “T-4 program”—‘Tiergartenstrasse 4″. The order allowed physicians to grant so-called ‘mercy killings’ to those deemed incurable as gauged by the current medical technologies. All euthanasia programs, however, were halted once word got out to the general German public. The National Socialists, thus, practiced both negative eugenics (selecting against people who they deemed unfit to have children) and positive eugenics (selecting for people who they deemed fit to have children).
The horrors of WWII Germany, though, caused great revile in every aspect of any kind of genetic modifications/selecting against/for any traits in humans. After the defeat of the Axis powers, in general, the public’s attitudes towards eugenics sharply changed.
In reaction to Nazi abuses, postwar politicians and members of the scientific community denounced the notion of any inherent inequality between human individuals.
[…]
After the Second World War, however, there was a significant drop of eugenic sterilizations of those with mental disabilities in response to a decline in support for eugenic policies as a whole. (MacKellar and Bechtel, 2016: 37-38)
Eugenics in Norway
In Norway, sterilization was first mentioned in 1927. They eventually enacted a law in 1934 based on the facts that parents could not look after their children and hereditary diseases. This continued all the way up until 1977, when the rights of the individual in question were considered. Though, unlike WWII Germany, there was no State-sanctioned eugenic policy, and each case was taken on a case-by-case basis.
Between the years 1934 and 1977, about 41,000 people were sterilized, with a majority consenting to the practice—about 75 percent were enacted on women (MacKellar and Bechtel, 2016: 39). However, between the years of 1943-1945, a law for the protection of the race was enacted in Norway while the Germans occupied the country—which was removed from law after Nazi Germany fell. However, in 1977, sterilizations changed from a way to control the people to a means of one’s ability to choose to limit their own reproduction.
Eugenics in Sweden
Sweden was the only Nordic country to have a national eugenics society. One of the leaders in eugenic thinking in Sweden in the early 1900s Arthur Engbert thought that we register pedigrees of our dogs and horses, so why not do so for ourselves (Swedes). Sweden, like Norway, enacted a sterilization act in 1934, though it only addressed sterilizations without the consent of the person; they did it to people who were legally incompetent. However, in 1941, this act was widened to include individuals with heritable physical disabilities. Women could also, of course, choose to sterilize themselves if they had too many children.
Thus, from 1935, when sterlizations were often being undertaken for eugenic reasons until 1975 when they were generally considered for medical reasons and the legislation was eventually overturned, Sweden sterlized nearly sixty-three thousand persons. Up to a quarter of these persons were mentally disabled. Though there was never a systematic sterilization program of mentally disabled individuals on the basis of political descisions or administrative instructions, the sterilizations seemed to develop on their own in the context of what was allowed. More than 90 per cent of these procedures were undertaken on women, but many individuals considered as having an antisocial way of life also felt the effects of somewhat coerced sterlizations. (MacKellar and Bechtel, 2016: 41)
Eugenics in the Soviet Union
Eugenics in the Soviet Union started with the biologists Aleksandr Serebrovskii in 1929. He recommended that artificial insemination be introduced to the populace in order to control who gives birth and who does not. He wrote (quoted in MacKellar and Bechtel, 2016: 41):
With the current state of artficial insemination technology…one talented and valuable produce could have up to one thousand children…In these conditions, human selection would make gigantic leaps forward. And various women and whole communes would then be proud…of their success and achievements in this undoubtedly most astonishing field — the production of new forms of human beings.
However, Stalin did not take to these proposals since, in his eyes, these types of eugenic policies contradicted Marxist ideology by claiming that human characters were determined by biology and not social/environmental ‘realities’. This, then, led to the deaths of many geneticists in the Soviet Union. (There are many good books on this matter, though my favorite is Lysenko’s Ghost.)
Eugenics in the United Kingdom
Sybil Gotto founded the Eugenics Education Society (EES) in 1907 in the UK to voice his concerns of the lower classes outbreeding the higher classes—what was perceived as a demographic problem. Francis Galton was the first Chair of this Society. So the EES proposed to prevent lower classes from breeding and promoting only breeding for the middle and higher classes, which would improve the whole society by excising the lower classes and replacing them with middle and upper classes. (There has to be a ‘bottom’ everywhere; so there still would exist ‘lower classes.’)
Winston Churchill was even quoted as saying persons in Britain “should, if possible, be segregated under proper conditions so that their curse diet with them and was not transmitted to future generations” (quoted in MacKellar and Bechtel, 2016: 43). After Churchill stated this, three years later, the UK Mental Deficiency Act of 1913 was established, and people deemed idiots, imbeciles, feebleminded or morally defective were detained in an effort to control the breeding of the population.
The Human Fertilisation and Embryology Act of 1990 was considered as allowing ‘soft’ forms of eugenics, it was stated that people would abort for even minor physical anomalies. Though, of course, this did end up occurring, lending credence to said ‘fears.’
Eugenics in America
Finally, the eugenics movement really became ‘scientific’ in America in the late 1800s to early 1900s. In fact, the first recorded eugenic experiment took place in America, at the so-called Perfectionist Community in Oneida, New York. The leader of this community was named John Noyes—a radical Christian, who believed that Christians had a moral responsibility to promote ‘moral perfection.’ He was influenced, of course, by Darwin and Galton. Between the years of 1869 and 1879, Noyes organized a campaign to get the so-called best to propagate. Members chosen to propagate were chosen on the basis of characters like ‘intelligence’, physical traits, and, of course, the commitment to Noyes’ vision (of a utopia). Fifty-eight children were born, but the project was soon abandoned.
Numerous US states passed laws barring certain people from marriage—imbeciles, epileptics, or those with feeble-minds. (See, for example, this article on Connecticut eugenic policies.) The Connecticut law was, in fact, the first eugenic policy passed in the country. These types of policies even were used to prevent immigration into America (see Dolmage, 2018 for a review).
Further, during the first 70 years of the 20th century, “eugenic policies affected up to sixty-four thousand Americans. But this happened primarily through measures such as forced sterilization. The 1907 Compulsory Sterilization Law of Indiana was the first to enact such legislation, though the public at large, was generally unaware of the initiative. According to this law, every institution that housed ‘confirmed criminals, idiots, rapists and imbeciles’ could authorize medical personnel ‘to perform such operation for the prevention of procreation’. This legislation was replicated by other states to such an extent that, by 1927, an estimated twenty-four states had enacted similar laws. Of these, the state of California was one of the most active, performing 4,636 sterilizations and castrations between 1907 and 1925, reaching a total of 9,930 by 1935″ (MacKellar and Bechtel, 2016: 46-47).
Then in 1927, a woman named Carrie Buck challenged the state of Virginia when a physician said that her sterilization would be for the ‘good of society.’ The high court of Virginia denied her claim, so she took it to the SCOTUS—known as Buck v. Bell. SCOTUS went 8-1 against Buck, saying that Buck “was a threat and danger to the genetic stability of society” (MacKellar and Bechtel, 2016: 47). So, the highest Court in the Land sided with the physician and not the woman. Eugenic policies were hardly opposed until 1974, with the case Relf v. Weinberger. The case came about due to to “the malicious, undesired sterilization of sisters Mary Alice and Minnie Reif” (MacKellar and Bechtel, 2016: 47). In the ruling, it was stated that mentally competent adults had to give their consent to sterilization, but this did not become standard until 1981.
Ultimately, formal acknowledgment of the ethically unacceptable abuse of sterilizations in the United States only came in 2003.
The international influence of American eugenic policies shpould not be underestimated. The Nazi government regularly cited a publication that touted favorable results of the sterilization policy in the state of California as evidence that wide-reaching sterilization programs were both feasible and humane. At the Nuremberg trials following the Second World War, Nazi administrators accused of war crimes actually justified the mass-sterilization of hundreds of thousands of people in less than a decade by referencing the United States success as their inspiration. (MacKellar and Bechtel, 2016: 47-48)
Conclusion
Eugenic thinking has been with us at least since the ancient Greeks and the idea has mutated over the times with different types of policies and measures each society has taken to ensure that their ‘genetic hygiene’ was as good as possible.
Eugenic thinking is, of course, still around today. Most recently, the Chinese using CRISPR—on twin girls—to create the first genetically modified humans, who were born this month. It is important to note that these claims have not been verified by other scientists nor have they been published yet. I am not too surprised at the outrage of this—if it did indeed happen (many people are skeptical that it did). I know some will say ‘Ethics don’t matter when the Chinese don’t care about ethics and look at what they’re doing!!!’ (If they truly did do this.) However, just because people disregard something doesn’t mean it does not exist. (The scientist, He Jiankui, is currently under investigation amidst questions of whether or not what he supposedly did was ethical and legal.)
Understanding the history of eugenics—the hows and whys—can prevent us from repeating past mistakes.
(For some info on the supposed Chinese gene-edited baby, see China’s gene-edited babies will push bioethics into a dark new era and A Sobering Moment for thoughts on the supposed gene-editing.)
Why Biological Racial Realism is True
1650 words
To be a realist about race is to hold that racial categories pick out real kinds in nature. (Smith, 2015: 43; Nature, Human Nature, and Human Difference Race in Early Modern Philosophy)
Claims from biological racial realists are simple: Racial categories pick out real kinds in nature. If racial categories pick out real kinds in nature, then race surely exists.
Racial groups—or groups taken to be racial groups—characterize three conditions from Hardimon (2017): C1: they are distinguished from other groups by patterns of visible physical features; C2: the members are linked by common ancestry which is peculiar to that group; and C3: they derive from a distinct geographic location.
Justifying C1 is simple: groups taken to be ‘racial’ are distinguished from other groups on the basis of physical characters. Someone from Europe looks different than someone from Africa; someone from Africa looks different than someone from Asia; someone from Asia looks different than someone from the Pacific Islands; someone from the Pacific Islands looks different than the Natives of America. Groups taken to be ‘racial’ have different facial features; they have different morphology. Thus, since there are heritable differences between groups taken to be ‘racial’, then this is evidence that race does indeed exist.
It’s important to also discuss what C1 does not demand: it does not demand that racial groups be distinguished by each of their visible physical features; it does not demand that each visible physical features of members of a race be identical; it allows skin color to vary just as much within race as it does between race; finally, it also allows great variation in hair color, skull morphology and skin color. Thus, since Hardimon’s concept is ‘vague’, then one might be able to say that it is “clinal” (that is, these differences vary by geography). But “Physical anthropologist Frank Livingstone’s well-known adage “There are no races, only clines” overlooks the possibility that, logically speaking, races might be clines” (Hardimon, 2017: 38). The claim “There are no races, only clines” is one that is oft-repeated against the reality of biological races.
C2, very simply, shows that differences in visible physical features are not the only things that delineate race: race is also defined in terms of ancestry and is therefore essential to the concept of race (I’d argue that ancestry is essential to any argument that attempts to establish races as biologically real). Races are, clearly, morphologically demarcated ancestry groups. The justification for C2 is thus: it is intuitive. Examples of race articulated in the past also bore this very basic concept: Linneus’ europeaus, asiaticus, afer, and americanus; Blumenbach’s Mongoloid, Caucasoid, Ethiopian, Malay and American; UNESCO’s Negroid, Mongoloid, and Caucasoid (deployed most famously by JP Rushton); and the Office of Management and Budget’s American Indian (or Alaskan native), black, Asian, whites, native Hawaiians (Pacific Islanders) (see Spencer, 2014 for a treatment of the OMB’s views on race and his ‘radical solution to the race problem’).
Now, finally, C3: the condition that groups taken to be ‘racial’ must derive from a distinct geographic location. Race, and the names used to refer to race, and so “The use of typonyms in the naming of racial groups suggests that the thinkers who chose these names were thinking of race as a geographical grouping” (Hardimon, 2017: 50). So, C1 and C2 have been established. This leaves us with C3. Races differ in patterns of visible physical features; these differences are explained by differences in geographic location. If race R1 derives from geographic location G1, and G1 is distinct from G2 which race R2 inhabits, then races R1 and R2 will look physically different.
Thus the groups that we think of when we think about race are groups that genetically transmit heritable characters to their offspring which then correspond to differences in geographic ancestry. So groups that satisfy C1-C3 are ‘races’, in the normal sense of the word. Groups that satisfy C1-C3 are articulated in Hardimon’s (2017) populationist race concept using Rosenberg et al’s (2002) data, and these are, largely, the same groups that Blumenbach pointed out centuries ago (Spencer, 2014).
The Visible Physical Features of Minimalist Race Are Racial
The visible physical features of minimalist race that correspond to geographical ancestry count as “racial” because they are defining features of minimalist races. They no more need to be correlated with normatively important features to be properly counted as racial then minimalist races need to be characterized by normatively important features to be properly counted as races. Just as the concept of minimalist race deflates the concept of RACE, so too it deflates the concept of RACIAL. Visible physical features that correspond to geographical ancestry are eo ipso racial. (Hardimon, 2017: 52)
Hardimon (2017: 99) also articulates one of the best definitions of race I have come across:
A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.
Since Hardimon’s views are new (published in 2017), there are no replies to his argument—excpet one, by Spencer (2018). Spencer doesn’t take to two of Hardimon’s claims: that (1) that the minimalist concept of race is the ordinary concept of race and (2) that minimalist races are biologically real. He grants (1) until Hardimon provides evidence that the minimalist concept of race is the ordinary concept of race. (2), on the other hand, Spencer attacks.
His objection to (2) comes down to the simple fact that 13/17 of the different conceptions of racial groups discussed by Linnaeus, Blumenbach, the OMB, and UNESCO do not fit C1-C3 (Blumenbach’s races do fit C1-C3). Spencer (2018) states that “there’s no ancestor that Eurasians share that’s not also shared by East Asians, Oceanians, and Native Americans […] there’s no ancestor that East Asians share that’s not also shared by Oceanians and Native Americans.” (See Duda and Zrzavy, 2016.) We need to be clear on what Hardimon means by “ancestry.” The dictionary definition of “ancestry” is thus: “one’s family or ethnic descent.” On this definition of “ancestry”, groups taken to be races do have “distinct ancestry“, so defined, and so, Hardimon’s (2017) C2 does indeed hold.
Biological racial realism “should” mean “race is a geniuine kind in biology.” Take the argument from Spencer (2011: 24):
(1) The meaning of ‘biological racial realism’ in the race debate should be a metaphysically minimal interpretation of important scientific kindhood that also does the most justice to what counts as an important scientific kind.
(2) A “metaphysically minimal” interpretation of important scientific kindhood is one that does not adopt unnecessary and contentious metaphysical assumptions.
(3) The interpretation of important scientific kindhood that does the most justice to what counts as an important scientific kind is the one that best captures epistemically important scientific kinds—or ‘EIS kinds’ for short.
(4) The candidates for important scientific kindhood in the race debate are natural kinds, naturali kinds, naturalu kinds, naturalp kinds, realp biological kinds, reali biological kinds, and geniuine kinds.
(5) No kind of kind in the race debate is both metaphysically minimal and does a better job of capturing EIS kindhood than genuine kinds.
(6) Therefore, the meaning of ‘biological racial realism’ in the race debate should be ‘race is a genuine kind in biology’.
Spencer has good critiques of Hardimon’s minimalist/populationist race view, but it does not hold.
Even if we allow Spencer’s views on Hardimon’s arguments for the existence of race to hold, Spencer himself has articulated a sound argument for the existence of race. In his 2014 paper A Radical Solution to the Race Problem, Spencer (2014) shows that Americans defer to the US Census on matters of race; the US Census defers to the OMB; the OMB refers to “sets of” populations—blacks, whites, Asians, Native Americans and Pacific Islanders; these “sets of” populations are not kinds (like what Hardimon argues his racial classifications are); therefore, races are not ‘kinds’, the term ‘race’ refers to sets of population groups. Thus, according to Spencer (2014), race refers to “proper names” for population groups, not “kinds”.
Both of Hardimon’s and Spencer’s arguments show that race is a biological reality; they both show that biological racial realism is true. Their concepts pick out real kinds in nature (Smith, 2016: 43).
Philosopher Justin Smith, in his book 2015 book Nature, Human Nature, and Human Difference Race in Early Modern Philosophy articulates a Hardimonian argument for the existence our habits of distinguishing between human populations:
Now if scientific taxonomy builds on folk taxonomy, and if racial classification builds on this in turn, there might be some basis for supposing that something about the modern habit of distinguishing between human groups on racial grounds is more deep-seated than we have acknowledged it to be. (Smith, 2015: 47)
The reason why there “might be some basis for supposing that something about the modern habit [which is not truly modern; Sarich and Miele, 2004; which I am sure that Smith knows due to the content of his book] of distinguishing between human groups on racial grounds is more deep-seated” than we have acknowledged because we are picking out real kinds that exist in nature.
Conclusion
Race, as a concept, is biologically real. Racial categories pick out real kinds in nature, as argued by Spencer (2014) and Hardimon (2017). Criticisms on Hardimon from Spencer or Spencer from Hardimon do not take away from this one fact: that race exists and is biologically real. Groups taken to be ‘racial’ look different from each other; they look different from each other due to their geographic locations and their ancestry (C1-C3). Since this is true, then race exists. We can argue this view simply:
P1. If groups of people look different from each other depending on where their ancestors evolved, then race exists.
P2. Groups of people look different from each other depending on where their ancestors evolved.
C. Therefore, race exists since people look different depending on where their ancestors evolved (modus ponens, P1, P2).
Biological racial realism is true.
An Argument For and Against Germline Editing
1200 words
In the past few weeks, talks of genetic modification have increased in the news cycle. Questions of whether or not to edit the genes of future people constantly arise. Should we edit genes and or the germline? If we edit the germline, what types of problems would occur? Is it moral to edit the germline of future people (babes are included in this as well) when they have no say? Is it moral to edit the genes of a baby that cannot consent to such editing? I will present one argument for and against editing the germline. This is a really big debate in current contemporary discourse; the argument for editing the germline rests on wanting the best for future people (which of course include our children) while the argument against editing the germline rests on the fact that said future people cannot consent to said germline editing so we should not edit the germline.
An argument for editing the germline
Germline editing is editing the germline in such a way that said edit is heritable—the modification to the germline is then acquired by the next generation of progeny. The rationale for editing the germline could be very simple:
Parents want what’s best for their children; since parents want what’s best for their children, then parents should edit their germline to rid their children of any disease and/or make them the best person they can possibly be, as is the job of all parents; therefore parents should edit the germline so said heritable changes can pass to the next generations since parents want what’s best for their children.
One may say that a babe has no choice in being born, naturally or artificially, and so since parents are able to choose the modifications, then this does consider the babe’s rights as a (future) person/human since it is, in theory, giving the babe the best possible chance at life with little, to no, diseases (that are noticed at conception). Parents can use new, up-and-coming genetic technology to attempt to give their child a head-start in life. They can edit their own germlines, and so, each change done to their germline would pass on to future progeny.
An argument against germline editing
Ethicist Walter Glannon articulates two great arguments against germline editing in his book Genes and Future People: Philosophical Issues in Human Genetics (2002). Glannon (2002: 89-90) writes:
Among other things, however, germ-line genetic alteration may not be desireable from an evolutionary perspective. Some genetic mutations are necessary for species to adapt to changing environmental conditions, and some genetic disorders involve alleles that confer a survival advantage on certain populations.
[…]
This raises the risk of whether or not we have a duty to prevent passing on altered genes with potentially harmful consequences to people who will exist in the distant future. It may recommend avoiding germ-line genetic manipulation altogether, which is supported by two related points. First, people existing in the future may be adversely affected by the consequences of a practice to which they did not consent. Second, because of the complex way in which genes interact, it would be difficult to weigh the probable health benefits of people in the present and near duture generations against the probable health burdens to people in the distant future. Because their interest in, and right to, not being harmed have just as much moral weight as those of the people who already exist or will exist in the near future, we would be well-advised to err on the side of caution. Indded, we would be morally obligated to do so, on the grounds of nonmaleficence. This would mean prohibiting germ-line genetic manipulation, or at least postponing it until further research can provide a more favorable assessment of its safety and efficacy.
I largely agree with Glannon here; though I will take his argument a step further: since future people literally cannot consent to germline genetic modificaitons then we should not edit our germline since we would be passing on the heritable changes to our descendants who did not ask for such changes.
The argument against germline editing is very simple:
(1) People should have a choice in whether or not their genes are modified.
(2) Since people should have a choice in whether or not their genes are modified, they then should be able to say “yes” or “no” to the modifications; though they cannot consent since they are not present to consent to the germline editing they will acquire in the future since they are not alive yet.
(3) Therefore we should not modify the germline without consent from future people, meaning that we should not edit the germline since there is a strong moral imperative to not do so since the future people in question cannot consent to the editing.
This argument against germline editing is a very strong moral argument: if one cannot consent to something, then that something should not be done. Future people cannot consent to germline editing. Therefore we should not edit the germline.
Another thing to think about is that if parents can edit the germline and genes of their children (future people), then it can be said that they would be more like “commodities”, like a handbag or whatnot, since they can make choices of what type of handbag they have, they would then make choices on what type of kid to have.
Hildt (2016) writes:
It is questionable whether there would be broader justifiable medical uses for germline interventions, especially in view of the availability of genetic testing and pre-implantation genetic diagnosis.
Gene editing can give us “designer babies” (though my argument presented above also is an argument against “designer babies” since they are future people, too); it can also put an end to many diseases that plague our society. However, there are many things we need to think about—both ethically/morally and empirically—before we even begin to think about editing our germline cells.
Conclusion
So, on the basis of (1) future people not being able to consent to said germline modifications and (2) us not knowing the future consequences of said germline editing, then we should not edit the germline. We, in fact, have a moral imperative to not do so since they cannot consent. The argument “for” germline editing, in my opinion, do not override the argument “against” germline editing. I am aware that most people would say “Who cares?” in regard to the arguments for or against germline modification, because people would “Just do it anyway.” Though, if there are laws against the editing of the germline, then germline editing cannot (should not) go through. Just because we *can* do something does not mean that we *should* do it.
We should not modify the germline because future people cannot consent to the changes. The moral argument provided here against germline editing is sound; the argument is a very strong moral one and since it is sound we should accept the argument’s conclusion that: “we should not modify the germline without consent from future people, meaning that we should not edit the germline since there is a strong moral imperative to not do so since the future people in question cannot consent to the editing.“
Arguments For and Against Preimplantation Genetic Diagnosis
1500 words
Like abortion, preimplantation genetic diagnosis (PGD) is feared. This is due, in part, to fears of eugenics coming back through a “backdoor” with the advent of new technology such as CRISPR/CAS9 and other types of tools we can use to genetically modify ourselves. The case of PGD—just like abortion—has been heavily debated in recent times, more so due to the recent strides in genomics we have made since the advent of the Human Genome Project.
PGD offers us a method to identify embryos with genetic diseases. Understandably, this has raised caution with some, due to the strong link with eugenic thinking/policies. See The Ethical Implications of Preimplantation Genetic Diagnosis. Thus, by scanning the genomes of fetuses, we can then see if they have a higher chance of acquiring any disease and select fetuses which have a lower to nonexistent chance of acquiring said disease.
An argument against PGD
In his paper Just diagnosis? Preimplantation genetic diagnosis and injustices to disabled people, Peterson (2005) presents one slippery slope argument against PGD (Freeman, 1996) (and later provides a refutation). The argument that Peterson (2005) presents is a “slippery slope” argument—that is, it’s an argument which argues that if we allow X, then since we allowed X, then horrible thing Y can and will follow. Peterson (2005) articulates the argument thus:
As situation A (the use of PGD to select against severe genetic diseases) is refined, “it will be difficult, if not impossible, to contain the uses of such research”. A will therefore bring about situation B, where PGD will be used to select against mild or perhaps non-medical conditions.
Besides the refinement of A, B will be brought about because “There will likely be an increasing pressure … on people to take advantage of these techniques, and not bring even a mildly disabled child into the world …”.
Finally, we could reach a morally abhorrent outcome Z, which is disturbingly close to eugenics, where our notion of the moral equality of all human beings, including those with disabilities, is undermined.
Z is so morally bad, that it outweighs the benefits of undertaking A.
Therefore, A should not be undertaken.
This argument, in my view, seems to be appealing to emotion by saying that since we can reach morally abhorrent outcome Z (a type of eugenics), then we should not continue with this practice. However, others argue that this discriminates against people with disabilities (see Katthab, 2009). Peterson (2005) argues that Freeman’s (1996) argument “lacks empirical support” and so it makes the conclusion difficult to assess; technology can and will be regulated which would quell any fears of possible use of this technology for any eugenic ideals; and, through using PGD, we can use it to “fight the obvious causes of discrimination, such as intolerance and egoism“, which would, in turn, reduce discrimination. Lastly, addressing Freeman’s (1996) concerns that PGD would lead to the discrimination of currently disabled persons, Peterson (2005) claims that “even if we accept that PGD will generate discrimination against disabled people, it is far from obvious that this is sufficient to warrant its moral condemnation.” Thus, Peterson (2005) concludes that Freeman’s (1996) argument is not sufficient to end the use of PGD technology. (Also see Robertson, 2003 for the view that “except for sex selection of the first child, most current extensions of PGD are ethically acceptable“.)
Many arguments against PGD rely on the concept of a fetus as a person and terminating any fetus with any prospective disease is paramount to killing a person. Others, of course, hinge on the fact that PGD does help reduce the risk of a babe being born with deleterious diseases, it does not completely ameliorate any generic risk for disease and so the fetus must be monitored through conception up until pregnancy to be sure that no disease appears during conception. And, of course, certain diseases that may hamper one’s quality of life may not appear until one reaches adolescence, adulthood, middle or old age. This is another fact against PGD: that even selecting embryos that apparently have no risk for disease, they may acquire diseases in older age which would not be seen since some diseases only generate symptoms at certain stages of life.
One final objection to PGD is also moral: it could, and will, send a message to any individuals currently alive that their lives are somehow “less” than others, since individuals with a chance to acquire said disease are selected against, as McConachy (2010) argues.
Lastly, Richardson (2017: 155-157) argues that the selection of embryos with so-called “potential” is ill-founded since they talk about vague concepts such as “egg quality”. Differences in potential lie, supposedly, in the “genetic blueprint” (see my article DNA Is not a Blueprint for arguments against that notion), though “That view implies that differences in individuals in important functions are largely due to differences in genes. As we have seen, though, things are far from being so simple” (Richardson, 2017: 156).
An argument for PGD
PGD can be used for many things; most importantly, screening the genome of a perspective fetus before IVF. Though, this has led some to worry that this could be a way in which eugenics can “sneak in through the backdoor” by virtue of making people with diseases more likely to be discriminated against since “disabled phenotypes” would slowly be phased out by PGD. One argument for could be:
Parents have rights; if parents have rights, then they have the right to do what they want with their children, and they want to do what is best for their children; therefore a parent should have the right to use PGD to select the best-possible embryo for implantation.
This is where we think of the implications of aborting a fetus, or not implanting a fetus that has a higher chance of acquiring any disease. There are, of course, certain people who would willingly select embryos which have a high chance to be disabled, because they themselves are disabled or they believe they “should be” disabled themselves and so want disabled children. Since parents have rights, as can be seen in the reasoning chain above, then parents should be able to choose the status of their babe. But if the babe’s quality of life is low, then is it ethical for that person to select an embryo with a high chance of acquiring a disease?
Another argument “for” PGD can be:
Humans should not suffer; if we can prevent human suffering with our current technology, then we have a moral imperative to do whatever is in our power to do so; if we can prevent low qualities of life for any embryo E, then we should do so; therefore, we should screen embryos for diseases that can and will lower their qualities of life and select against these embryos.
One may argue that a fetus may not have a “moral right” to life (see Tooley, 1972), though, if we know that a fetus has a high chance to have such a debilitating disease such that it lowers its quality of life, then it should be aborted/not implanted in the womb. Religious views, of course, come into play here but I am not worried about them; I am worried only about sound arguments for them. (See Fasoulioutis and Schenker, 1997 for these views.)
PGD may, of course, prevent abortions of said fetus since we know that the fetus in question may have a higher chance of acquiring a certain disease, so if one is against abortion, then they may be for the use of PGD to screen the fetuses’ genome to scan for any readily apparent problems in their genome in regard to the acquiring of certain genetic diseases.
Arguments for PGD hinge on parents wanting the best possible lives for the children they conceive and the arguments really rely on parental autonomy, the parent’s want to choose how their kid is born, if their chance for disease is high or not (which also would turn to “designer babies”; an argument against “designer babies” will be erected soon. If parents can do what’s best for an unborn child then, most would argue they have a moral imperative to give the babe the best possible life and so they should abort/select against certain embryos which have a high chance of acquiring diseases.
Conclusion
There do not seem to be as many strong arguments against PGD compared to abortion. Though one can use the basic blueprint of the argument against abortion and liken it to PGD. The PGD debate is similar to the abortion debate. One can use similar arguments against abortion to argue against PGD.
These debates are both ethical scientific: we have the ability to now do X which would stop suffering Y in embryo E. Just because we can do something, does that mean we should do so? Like with the editing of the germline, we don’t know what types of consequences would occur since we have, pretty much, no experience in editing the germline/genes of humans in a large-scale way.
An Argument For and Against Abortion
1350 words
Abortion is a touchy subject for many people. There are many different arguments for and against abortion, including, but not limited to, the woman’s right to do what she wants with her body on the pro-abortion side, to the right of a fetus to live a good life if there is little chance of the fetus developing a serious disease. In this article, I will provide two arguments: one for and one against abortion. The abortion debate is an ethical, not scientific, one, and so, we must use argumentation to see the best way to move forward in this debate.
An argument for abortion
Michael Tooley, in his paper Abortion and Infanticide, provides an argument not only for the abortion of fetuses, but the killing of infants and animals since they cannot conceive of continuing their selves. He argues that an organism only has a right o life of they can conceive of that right to life. His conclusion is that it should be morally permissible to end a baby’s life shortly after birth since it cannot conceive of wanting to live. The conclusion of the argument also includes—quite controversially, in fact—young infants and (nonhuman) animals. Ben Saunders articulates Tooley’s argument in Just the Arguments: 100 of the Most Important Arguments in Western Philosophy (2011: 284-286):
P1. If A has a morally serious right to X, then A must be able to want X.
P2. If A is able to want X, then A must be able to conceive if X.
C1. If A has a morally serious right to X, then A must be able to conceive of X (hypothetical syllogism, P1, P2).
P3. Fetuses, young infants, and animals cannot conceive of their continuing as subjects of mental states.
C2. Fetuses, young infants, and animals cannot want their continuance as subjects of mental states (modus tollens, P2, P3).
C3. Fetuses, young infants, and animals do not have morally serious rights to continue as subjects of mental states (modus tollens, P1, C2).
P4. If something does not have a morally serious right to life, then it is not morally wrong to kill it painlessly.
C4. It is not wrong to kill fetuses, young infants, or animals painlessly (modus ponens, C3, P4).
Of course, most people would seriously disagree with C4, since a babe’s life is one of the most precious things in the world— the protection of said babes is how we continue our species. However, the argument is deductively valid, and so one must show which premise is wrong and why. This argument—along with the one that will be presented below against abortion (of healthy fetuses)—is very strong. Thus, if a woman so pleases (along with her autonomy), she can choose to abort her fetus since it is not wrong to kill a fetus painlessly. (I am not aware if fetuses can feel pain or not, however. If they can, then the conclusion of this argument does not hold.)
Tooley’s argument regarding the killing of infants is similar to an argument made by Gibiulini and Minerva (2013) who argue that since fetuses and newborns don’t have the same moral status as actual persons, fetuses and infants can eventually become persons, and since adoption is not always in the best interests 9f people, then “‘after-birth abortion’ (killing a newborn) should be permissible in all the cases where abortion is, including cases where the newborn is not disabled” (Giubilini and Minerva, 2013).
An argument against abortion
One strong argument against abortion exists: Marquis’ (1989) argument in his paper Why Abortion Is Immoral. Women may want an abortion for many reasons: such as not wanting to carry a babe to term, to finding out that the babe has a serious genetic disorder. Though, what matters to this argument is not the latter, but the former: the mother wanting an abortion of a healthy fetus. Marquis’ argument is simple: killing is wrong; killing is wrong since killing ends one’s life, and ending one’s life means they won’t experience anything anymore, they won’t be happy anymore, they won’t be able to accomplish things, and this is one of the greatest losses that can be suffered; abortions of a healthy fetus cause the loss of experiences, activities, and enjoyment to the fetus; thus, the abortion of a healthy fetus is not only ethically wrong, but seriously wrong. Marquis’ (1989) argument is put succinctly by Leslie Burkholder in the book Just the Arguments: 100 of the Most Important Arguments in Western Philosophy (2011: 282-283):
P1. Killing this particular adult human being or child would be seriously wrong.
P2. What makes it so wrong is that it causes the loss of this individual’s future experiences, activities, projects, and enjoyments, and this loss is one of the greatest losses that can be suffered.
C1. Killing this adult human being or child would be seriously wrong, and what makes it so wrong is that it causes the loss of this individual’s future experiences, activities, projects, and enjoyments, and this loss is one of the greatest losses that can be suffered (conjunction, P1, P2).
P3. If killing this particular adult human being or child would be seriously wrong and what makes it so wrong is that it causes the loss of all this individual’s experiences, activities, projects, and enjoyments, and this loss is one of the greatest losses that can be suffered, then anything that causes to any individual the loss of all future experiences, activities, projects, and enjoyments is seriously wrong.
C2. Anything that causes to any individual the loss of all future experiences, activities, projects, and enjoyments is seriously wrong (modus ponens, C1, P3).
P4. All aborting of any healthy fetus would cause the loss to that individual of all its future experiences, activities, projects, and enjoyments.
C3. If A causes to individual F the loss of all future experiences, activities, projects, and enjoyments, then A is seriously wrong (particular instantiation, C2).
C4. If A is an abortion of healthy fetus F, then A causes to individual F the loss of all future experiences, activities, projects, and enjoyments (particular instantiation, P4).
C5. If A is an abortion of a healthy fetus F, then A is seriously wrong (hypothetical syllogism, C3, C4).
C6. All aborting of any healthy fetus is seriously wrong (universal generalization, C5).
In this case, the argument is about abortion in regard to healthy fetuses. This argument, like the one for abortion, is also deductively valid. (Arguments for and against the abortion of unhealthy fetuses will be covered in the future.) Thus, if a fetus is healthy then it should not be aborted since doing so would cause the individual to lose their future experiences, enjoyments, activities, and projects. Thus, the abortion of a healthy fetus is seriously and morally wrong. This argument clearly establishes the fetuses’ right to life if it is healthy.
Conclusion
Both of these arguments for and against abortion are strong; on the “for” side, we have the apparent facts that fetuses, infants, and (nonhuman) animals cannot want their continuance of their mental states since they cannot conceive of their continuance and want of mental states, so if they cannot want their continuance of their mental states they do not have a morally serious right to life and it is, therefore, morally right to kill them painlessly. On the “against” side, we have the facts that aborting healthy fetuses will cause the loss of all future experiences, enjoyments, activities, and projects, and so, the abortion of these healthy fetuses is both seriously and morally wrong.
I will cover these types of arguments—and more—in the future. However, if one is against genetic modification, embryo selection, preimplantation genetic diagnosis (PGD) and ‘eugenics’, then one must, logically, be against the abortion of healthy fetuses as well. These two arguments, of course, have implications for any looming eugenic policies as well, which I will cover in the future.
(I, personally, lean toward the “against” side in this debate; though, of course, the argument presented in this article on the “for” side is strong as well.)
The Physical Reality of Race
1600 words
Rational people can just look at people of different ancestries and see that there is something to what we call “race.” We notice that others look different based on where their ancestors came from and we classify people into different races on the basis of their physical appearance. Anti-biological racial realists may point to the fact that there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016; Hardimon, 2017). While this is true, this does not mean that race is “just a social construct” (a phrase used to deflate the meaning of “race”); it is both a social construct and a biological reality.
The definition of race is simple—a group of populations which genetically transmit heritable characteristics which correspond to that group’s geographic ancestry who also belong to a biological line of descent which was initiated by a geographically isolated and reproductively isolated founding populations (Hardimon, 2017). Note how this definition says nothing about differences in allele frequencies between populations between populations—because, for these purposes, they’re irrelevant for the argument being made. The fact of the matter is, the reality of race hinges on two things: (1) the heritable differences between population groups which were geographically/reproductively isolated and (2) our ability to discern these population groups by their phenotype.
A great book on the history of race, its meaning and how the term was used over the ages is Race: The Reality of Human Differences by Sarich and Miele (2004). For the purposes of this piece, the first two chapters are the most important, since they touch on aspects of race that I have in the past—mainly the fact that we only need phenotype to discern one’s race. People from Europe look phenotypically different from people from Africa who look phenotypically different from people from Asia etc. These differences between these groups are evidence that race exists—these racial differences in phenotype are due, in part, to the climate they evolved in while geographically and reproductively isolated (two conditions for racehood).
Sarich and Miele (2004: 29) write:
Vince [Sarich; one of the authors of the book] naively asked for the legal definition of “race” and was told there wasn’t one.
[…]
As we began working on this book, we discussed the issue of the legal definition of “race” … He informed us that there is still no legal definition of “race”; nor, as far as we know, does it appear that the legal system feels the need for one. Thus, it appears that the most adversarial part of our complex society, the legal system, not only continues to accept the existence of “race” but also relies on the ability of the average individual to sort people into races. Our legal system treats “racial identification” as self-evident …
[…]
The courts have come to accept the commonsense definition of race, and it is this commonsense view that, as we show, best conforms to reality. A look at two recent (2000) cases is illustrative. In both Rice v. Office of Hawaiian Affairs and in Hank v. Rochester School District, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race (even if the racial categories are euphemistically termed “peoples” or “populations”). No special expertise was assumed or granted in defining or recognizing race other than the everyday commonsense usage, as given in the Oxford English Dictionary, that a race is “a group of persons connected by common descent” or “a tribe, nation, or people, regarded as common stock.” The courts and the contending parties, in effect, accepted the existence of race and the ability of the ordinary person to distinguish between races based on a set of physical features.
In Rice v. Office of Hawaiian Affairs, Rice challenged the state of Hawaii since they did not allow him to vote—on the basis that he was not a native Hawaiian, and that the electoral system of Hawaii is for the benefit of Hawaiians and Hawaiians only. Everyone agreed that Rice was a Hawaiian citizen—but he did not have Hawaiian ancestry, so he could not be recognized as “Hawaiian” under state law. However, the SCOTUS overturned the ruling (that Rice should not be allowed to vote on the basis of not having Hawaiian ancestry) 7-2, citing the 15th amendment: “The right of the citizens of the United States to vote shall not be denied or abridged by the United States or any State on account of race, color, or previous condition of servitude.” Sarich and Miele (2004: 31) write “The 15th amendment is explicit—race means what the average person thinks it means—and the majority of the Supreme Court read it that way.” (Also see Hong, 2008 for an overview of the case.)
On the other hand, in Haak v. Rochester School District, the Second Circuit Court of Appeals ruled that a white fourth-grade student named Jessica Haak could transfer from her current district to another district (full of whites) since the transfer program was initiated with the idea of lessening the racial isolation of the adjoining districts. Jessica’s mother cited the 14th amendement, and a district court ruled in their favor but the Second Circuit Court of Appeals overturned the decision. “A “minority pupil” was defined as “a pupil who is of Black or Hispanic origin or is a member of another minority group that historically has been the subject of discrimination” (Sarich and Miele, 2004: 31).
The critical points here are that in both Rice and Haak, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race. No special expertise was assumed or granted in defining or recognizing race other than the everyday usage of the term. In Rice, the court, in effect, took judicial notice of the commonsense definition of race. In Haak, the court accepted physical appearance as a valid means by which the average citizen can recognize races and distinguish among them.
In short, the courts accepted the existence of race, even if the legislature was afraid to use the offending word.
Despite the fact that Sarich and Miele (2004) claim that there is no legal definition of race, Cornell Law School has one definition stating that “the term “racial group” means a set of individuals whose identity as such is distinctive in terms of physical characteristics or biological descent.” While the Law Dictionary, citing the 15th amendment writes that race is “A tribe, people, or nation, belonging or supposed to belong to the same stock or lineage. “Race, color, or previous condition of servitude.” Const U. S., Am. XV.” (Also see Hoffman, 2004 who argues that “race” should not be used in the legal system.)
Notice how Sarich and Miele’s (2004) description of “race” and what “race” is almost—word-for-word—like Spencer’s Blumenbachian partitions (Spencer, 2014). Americans defer to the US Census Bureau on matters of race; the US Census Bureau defers to the Office of Management and Budget (OMB) who speak of sets of populations; these sets of populations correspond to geographic clusters who have distinct phenotypes based on their geographic ancestry, which the average American can discern; therefore race exists. Spencer states that when Americans refer to “race” that Americans refer to both a social construct and a biological reality—that is, Americans socially construct race (think of how Hardimon’s minimalist concept of race is related to the concept of socialrace) but these social constructs do have biological underpinnings which can be discerned in two ways: (1) just observation of phenotypes and (2) looking into the genomes of genetically related individuals who make up these population groups.
Even the ancients distinguished races and sorted them on the basis of hair color/type, skin color, physiognomy etc. “[The Egyptians, Greeks, Romans, Indians, and Chinese] sorted [broad racial groups] based upon the same set of characteristics—skin color, hair form, and head shape” while “it is evident that they relied upon a set of observable features (skin color and form, body build, facial features) quite similar to those used in the commonsense notion of race and the racial classifications of nineteenth-century anthropology to sort the many diverse groups they encountered into a smaller number of categories” (Sarcih and Miele, 2004: 42).
It is very clear that, ever since antiquity at the very least, we have been classifying racial groups on the basis of phenotype—and, come to find out, this is one of the best ways to sort people—and you don’t even need to look at genetic differences between groups. Phenotype is clearly enough to delineate racial groupings, you don’t need genes to delineate race. We only need to recognize that (1) people look different on the basis of where they (or their ancestors) came from; (2) observe that these physical differences between people who come from different places are between real and existing groups; (3) people have common ancestry with others; (4) people derive from distinct geographic locations; so (5) we can infer that race exists.
Race is very clearly a reality—both biologically and socially. At least three sound arguments exist for the existence of race (Sarich and Miele, 2004; Spencer, 2014; Hardimon, 2017; see Hardimon’s and Spencer’s arguments at length). Even those in antiquity delineated races on the basis of physical features—exactly what has been argued by Spencer and Hardimon. Race is physically real—people look different from each other individually, ethnically, and racially.
Biological racial realism is true, and if biological racial realism is true then race exists.
(1) If groups of people look different from each other depending on where their ancestors evolved, then race exists.
(2) Groups of people look different from each other depending on where their ancestors evolved.
(3) Therefore, race exists since people look different depending on where their ancestors evolved.
My Reading List
In no particular order, below is my reading list. This is not an extensive list of the books I’ve read in my life, only what I’ve read (bought) in the past three years
(1) Race: The Reality of Human Differences (Sarich and Miele, 2004)
(2) Beyond Versus: The Struggle to Understand the Interaction Between Nature and Nurture (Tabery, 2014)
(3) Out of Our Heads: Why You Are Not Your Brain, and Other Lessons from the Biology of Consciousness (Noë, 2014)
(4) Limits of Science? Important things we do not know about everything (Beerbower, 2016)
(5) Evolutionary Biology: Conceptual, Ethical, and Religious Issues (Edited by Thompson and Walsh, 2014)
(6) Genes and Future People: Philosophical Issues in Human Genetics (Glannon, 2002)
(7) The Skull Collectors: Race, Science, and America’s Unburied Dead (Fabian, 2010)
(8) The Ontogeny of Information: Developmental Systems and Evolution (Oyama, 1985)
(9) Evolution’s Eye: A System’s View of the Biology-Culture Divide (Oyama, 2000)
(10) I am Not a Brain: Philosophy of Mind for the 21st Century (Gabriel, 2017)
(11) Agents and Goals in Evolution (Okasha, 2018)
(12) Getting Darwin Wrong: Why Evolutionary Psychology Won’t Work (Wallace, 2010)
(13) Sex and Death: An Introduction to Philosophy of Biology (Sterelny and Griffiths, 1999)
(14) What Darwin Got Wrong (Fodor and Piatteli-Palmarini, 2010)
(15) The Sports Gene: Inside the Science of Extraordinary Athletic Performance (Epstein, 2014)
(16) Dance to the Tune of Life: Biological Relativity (Noble, 2017
(17) Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life (Jablonka and Lamb, 2005)
(18) Making Sense of Genes (Kampourakis, 2017)
(19) Did My Neurons Make Me Do It?: Philosophical and Neurobiological Perspectives on Moral Responsibility and Free Will (Murphy and Brown, 2009)
(20) The Essential Davidson (Introduction by Lepore and Ludwig, 2006)
(21) Evolutionary Psychology as Maladapted Psychology (Richardson, 2007)
(22) Ingenious Genes: How Gene Regulation Networks Evolve to Control Development (Sansom, 2011)
(23) Mind and Cosmos: Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False (Nagle, 2012)
(24) Mental Causation: The Mind-Body Problem (Dardis, 2008)
(25) Paleofantasy: What Evolution Really Tells Us About Sex, Diet, and How We Live (Zuk, 2014)
(26) Logic and Philosophy: An Integrated Introduction (Brenner, 1993)
(27) Why Gould Was Wrong (Oeijord, 2003)
(28) Not in Your Genes: The Real Reason Why Children Are Like Their Parents James, 2016)
(29) Evolving Human Nutrition: Implications for Public Health (Ulijaszek, Mann, and Elton, 2012)
(30) The Philosophy of Human Evolution (Ruse, 2012)
(31) Genetics and Philosophy: An Introduction (Griffiths and Stotz, 2013)
(32) What is Philosophy For? (Midgley, 2018)
(33) A Brief History of Everyone Who Ever Lived: The Human Story Retold Through Our Genes (Rutherford, 2017)
(34) Genes, Polymorphisms, and the Making of Societies: How Genetic Behavioral Traits Influence Human Cultures (Kiaris, 2012)
(35) The Trouble with Twin Studies: A Reassessment of Twin Research in the Social and Behavioral Sciences (Joseph, 2015)
(36) Race Unmasked: Biology and Race in the Twentieth Century (Yudell, 2014)
(37) Inventing Intelligence: How America Came to Worship IQ (Castles, 2012)
(38) Beyond Human Nature: How Culture and Experience Shape the Human Mind (Prinz, 2014)
(39) Darwin Deleted: Imagining a World without Darwin (Bowler, 2013)
(40) Getting Science Wrong: Why the Philosophy of Science Matters (Dicken, 2018)
(41) The Philosophy of Cognitive Science (Cain, 2015)
(42) Social By Nature: The Promise and Peril of Sociogenomics (Bliss, 2018)
(43) Extended Heredity: A New Understanding of Inheritance and Evolution (Bonduriansky and Day, 2018)
(44) Genes: A Philosophical Inquiry (Graham, 2002)
(45) Philosophy of Microbiology (O’Malley, 2014)
(46) Genes, Cells, and Brains: The Promethean Promises of the New Biology (Rose and Rose, 2013)
(47) The Genome Factor: What the Social Genomics Revolution Reveals About Ourselves, Our History, and the Future (Coney and Fletcher, 2017)
(48) Endless Forms Most Beautiful: The New Science of Evo Devo (Carroll, 2006)
(49) The Diabetes Code: Prevent and Reverse Type 2 Diabetes Naturally (Fung, 2018)
(50) The Developing Genome: An Introduction to Behavioral Epigenetics (Moore, 2015)
(51) The Epigenetics Revolution: How Biology is Rewriting Our Understanding of Genetics, Disease, and Inheritance (Carey, 2012)
(52) Darwin’s Athletes: How Sport Has Damaged Black America and Preserved the Myth of Race (Hoberman, 1996)
(53) Psychology and Race (Edited by Watson, 1973)
(54) The Human Superorganism: How the Microbiome is Revolutionizing the Pursuit of a Healthy Life (Dietert, 2016)
(55) Reliability in Cognitive Neuroscience: A Meta-Meta-Analysis (Uttal, 2012)
(56) Studying Human Behavior: How Scientists Investigate Aggression and Sexuality (Longino, 2013)
(57) Philosophy of Mind: An Introduction (Jaworski, 2011)
(58) Straightening the Bell Curve: How Stereotypes about Black Masculinity Drive Research on Race and Intelligence (Hilliard, 2012)
(59) Schizophrenia and Genetics: The End of an Illusion (Joseph, 2017)
(60) Disabled Upon Arrival: Eugenics, Immigration, and the Construction of Race and Disability (Dolmage, 2018)
(61) Living Color: The Biological and Social Meaning of Skin Color (Jablonski, 2012)
(62) The Evolved Apprentice: How Evolution Made Humans Unique (Sterelny, 2012)
(63) Who We Are and How We Got Here: Ancient DNA and the New Science of the Human Past (Reich, 2018)
(64) The Gene: From Genetics to Postgenomics (Rheinberger and Müller-Wille, 2018)
(65) Misbehaving Science: Controversy and the Development of Behavior Genetics (Panofsky, 2014)
(66) Postgenomics: Perspectives on Biology After the Genome (Edited by Richardson and Stevens, 2015)
(67) Behave: The Biology of Humans at Our Best and Worst (Sapolsky, 2017)
(68) The Century of the Gene (Keller, 2000)
(69) My Cells Made Me Do It: The Story of Cellular Determinism (Hayes, 2015)
(70) Intelligence and How to Get It: Why Schools and Cultures Count (Nisbett, 2010)
(71) The Obesity Epidemic: Why Diets and Exercise Don’t Work—And What Does (Toomath, 2017)
(72) Genes, Brains, and Human Potential: The Science and Ideology of Intelligence (Richardson, 2017)
(73) Rethinking Race: The Case for Deflationary Realism (Hardimon, 2017)
(74) DNA Is Not Destiny: The Remarkable, Completely Misunderstood Relationship Between You and Your Genes (Heine, 2017)
(75) Everyone Is African: How Science Explodes the Myth of Race (Fairbanks, 2015)
(76) The Story of the Human Body: Evolution, Health, and Disease (Lieberman, 2013)
(77) The End of Overeating: Taking Back the Insatiable American Appetite (Kessler, 2009)
(78) Full House: The Spread of Excellence from Plato to Darwin (Gould, 1996)
(79) Catching Fire: How Cooking Made Us Human (Wrangham, 2009)
(80) J. Phillipe Rushton: A Life History Perspective (Dutton, 2018)
(81) Why Diets Make Us Fat: The Unintended Consequences of Our Obsession with Weight Loss (Aamodt, 2016)
(82) The Blank Slate: The Modern Denial of Human Nature (Pinker, 2003)
(83) Understanding Biology (Mason et al, 2018)
(84) Up From Dragons: The Evolution of Human Intelligence (Skoyles and Sagan, 2002)
(85) Anatomy and Physiology: The Unity of Form an Function (Saladin, 2010)
(86) The Anatomy of Violence: The Biological Roots of Crime (Raine, 2014)
(87) The Selfish Gene (Dawkins, 1976)
(88) Why We Get Fat and What to Do About It (Taubes, 2011)
(89) The Obesity Code: Unlocking the Secrets of Weight Loss (Fung, 2016)
(90) A New History of Life: The Radical New Discoveries About the Origins and Evolution of Life on Earth (Ward and Kirschvink, 2016)
(91) A Troublesome Inheritance: Genes, Race, and Human History (Wade, 2014)
(92) Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It (Entine, 2000)
(93) Monad to Man: The Concept of Progress in Evolutionary Biology (Ruse, 1996)
(94) Descent of Man, and Selection in Relation to Sex (Darwin, 1871)
(95) On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life (Darwin, 1859)
(96) Guns, Germs, and Steel: The Fates of Human Societies (Diamond, 1997)
(97) This is Your Brain on Parasites: How Tiny Creatures Manipulate Our Behavior and Change Our Society (McAuliffe, 2016)
(98) Secrets from the Eating Lab: The Science of Weight Loss, the Myth of Willpower, and Why You Should Never Diet Again (Mann, 2015)
(99) Darwin’s Unfinished Symphony: How Culture Made the Human Mind (Laland, 2017)
(100) Wonderful Life: The Burgess Shale and Natural History (Gould, 1989)
(101) The Evolution of Beauty: How Darwin’s Forgotten Theory of Mate Choice Shapes the Animal World—And Us (Prum, 2017)
(102) Arrival of the Fittest: How Nature Innovates (Wagner, 2015)
(103) This Idea Must Die: Scientific Theories That Are Blocking Progress (Edited by Brockman, 2015)
(104) The Righteous Mind: Why Good People are Divided by Politics and Religion (Haidt, 2012)
(105) Humankind: How Biology and Geography Shape Diversity (Harcourt, 2016)
(106) Rethinking Thin: The New Science of Weight Loss–and the Realities of Dieting (Kolata, 2008)
(107) Why Evolution Is True (Coyne, 2010)
(108) The Greatest Show on Earth: The Evidence for Evolution (Dawkins, 2009)
(109) The Complete Guide to Fasting: Heal Your Body Through Intermittent, Alternate-Day, and Extended Fasting (Fung and Moore, 2016)
(110) Altruism, Socialization, and Society (Rushton, 1980)
(111) The Paradox of Evolution: The Strange Relationship Between Natural Selection and Reproduction (Rothman, 2015)
(112) The Meaning of Human Existence (Wilson, 2015)
(113) The Red Queen: Sex and the Evolution of Human Nature (Ridley, 1993)
(114) The Evolution Delusion: A Scientific Study of Creation and Evolution (Kirkwood, 2015)
(115) The Ten Thousand Year Explosion: How Civilization Accelerated Human Evolution (Cochran and Harpending, 2009)
(116) Moral Tribes: Emotion, Reason, and the Gap Between Us and Them (Greene, 2013)
(117) The God Delusion (Dawkins, 2006)
(118) The Human Advantage: A New Understanding of How Our Brain Became Remarkable (Herculano-Houzel, 2016)
(119) The Testosterone Hypothesis: How Hormones Regulate the Lifecycles of Civilizations (Barzilai, 2015)
(120) Population Wars: A New Perspective on Competition and Coexistence (Graffin, 2016)
(121) An Odyssey in Time: The Dinosaurs of North America (Russell, 1992)
(122) Race, Evolution, and Behavior: A Life History Perspective (Rushton, 1995)
(123) The g Factor: The Science of Mental Ability (Jensen, 1998)
(124) IQ and Human Intelligence (Mackintosh, 1998)
(125) Good Calories, Bad Calories: Fats, Carbs, and the Controversial Science of Diet and Health (Taubes, 2008)
(126) Intelligence in the Flesh: Why Your Mind Needs Your Body Much More Than It Thinks (Claxton, 2015)
(127) The Bell Curve: Intelligence and Class Structure in American Life (Murray and Herrnstein, 1994)
(128) The Genius in All of Us: New Insights into Genetics, Talent, and IQ (Shenk, 2010)
On Individual Variation in Response to Exercise
1500 words
On Twitter, JayMan wrote: “Not talked about much by fitness buffs (a world that’s full of BS anyway): a fair fraction of people respond little to even *negatively* to exercise“. This is the same person that thinks behavior genetics is a science, and that is a field “that’s full of BS anyway”, too. Anyway, the article that JayMan cited was from the website Stronger by Science, titled Hardgainers? What We Know About Non-Responders by Greg Nuckols.
First off, JayMan’s comment that “a fair fraction of people respond … *negatively* to exercise” is, on its face, already false. Most everyone in the study referenced by Nuckols (There Are No Nonresponders to Resistance-Type Training in Older Men and Women; Churchward-Venne et al, 2015) gained strength, but some people’s muscle fibers did not grow, and some apparently shrank (that is, their muscle cross-section area; CSA). But the important thing to note is that ALL gained strength, which implies physiologic adaptation to the stressor placed on the body (something that is overlooked).
Though, even if some people do not respond to certain programs or weight/rep schemes, does not mean that they are “non-responders”. All that needs to be done is to change the program if one “does not respond” to the program created. All exercise programs should be tailored to the individual and their own specific goals. There is no “one-size-fits-all” exercise program, as can be seen from these studies on so-called “hardgainers.”
The best study for this matter, though, is the HERITAGE (HEalth, RIsk factors, exercise, Training, And GEnetics) study, carried out by five universities in Canada and the US, who enlisted 98 two-generation families and then subject each member to five months of the same stationary bike training regimen—three workouts per week with increasing intensity. Each of the 482 individuals in the study was assayed, and so we would also see which genes would play a role in how fit one person would be in comparison to another.
David Epstein, author of The Sports Gene, writes (pg 85):
Despite the fact that every member of the study was on an identical exercise program, all four sites saw a vast and similar spectrum of aerobic capacity improvement, from about 15 percent of participants who showed little or no gain whatsoever after five months of training all the way up to 15 percent of participants who improved dramatically, increasing the amount of oxygen their bodies could use by 50 percent or more.
Amazingly, the amount of improvement that any one person experienced had nothing to do with how good they were to start. In some cases, the poor got relatively poorer (people who started with a low aerobic capacity and improved little); in others, the oxygen rich got richer (people who started with high aerobic capacity and improved rapidly); with all manner of variation in between—exercisers with a high baseline aerobic capacity and little improvement and others with meager starting aerobic capaacity whose bodies transformed drastically.
Though, contrary to JayMan’s claims, “Fortunately, every single HERITAGE subject experienced health benefits from exercise. Even those who did not improve at all in aerobic capacity improved in some other health parameter, like blood pressure, cholesterol, or insulin sensitivity” (Epstein, 2014: 88).
Epstein also writes about another study, undertaken at the University of Alabama-Birmingham’s Core Muscle and Research Laboratory, writing:
Sixty-six people of varying ages were put on a four-month strength training plan—squats, leg press, and leg lifts—all matched for effort level as a percentage of the meximinum they could lift. (A typical set was eleven reps at 75 percent of the maxmimum that could be lifted for a single rep.) At the end of the trainin, the sibjects fell rather neatly into three groups: those whose thigh muscle fibers grew 50 percent in size; those whose fibers grew 25 percent; and those who had no increased in muscle size at all.
[…]
Seventeen weight lifters were “extreme responders” who added muscle furiously; thirty-two were moderate responders, who had decent gains; and seventeen were nonresponders, whose muscle fibers did not grow.* (pg 110)
* “It’s important to keep in mind that the harder the training, the less likely there are to be “nonresponders.” The harder the work, the more likely a subject will get at least some response, even if it is less than her peers” (pg 376).
Those who responded the most to the regimen had the most satellite cells in their quads which were waiting to be activated by training. When one becomes stronger from hypertrophy, the muscle thickness correlates to muscle CSA (Franchi et al, 2018). When one performs a repetition, the muscle fibers break down—this leads to trauma of the cellular proteins in the muscles which must then go under repair. Numerous growth factors influence the growth of skeletal muscle, such as GH (growth hormone), testosterone, protein and carb intake. Skeletal muscle adapts almost immediately after a bout of exercise, but the apparent changes to the muscle (both in the mirror and seeing large gains in strength on any particular movement) will take weeks and months.
There’s one thing about the claims of “exercise nonresponders” that really gets me: everyone responds positively to exercise, even if it’s not the same exact response to another individual doing the same—or different—exercise! I don’t know who made the claim that “people respond the same to any exercise program”, but that’s a claim that hbdchick made, writing “plenty of the “fitness buffs” do [make the claim that everyone would respond the same to the same exercise regimen]. I then asked her, and JayMan, to name three people who made this outrageous claim: but, of course, I got no answer.
Not to mention that Nuckols ended the article writing:
… there were way fewer nonresponders when people were put on personalized training programs instead of one-size-fits-all standardized programs. This study was primarily looking at aerobic fitness, but it also examined strength measures (bench press and leg press 5RM). It found that all the subjects on personalized programs got stronger, while only 64.3% of the subjects on standardized programs got stronger. This gives us more evidence that “nonresponders” in scientific studies aren’t necessarily “true” nonresponders.
Take two people who have similar measures and, say, start at the same weight on one exercise. In 6 months, all else being equal with regard to lifestyle, there will be a difference in strength gained on that particular exercise. However, an increase from t he baseline from when both individuals began, to the 6-month point, shows that they did, indeed, respond to the exercise program at least in some way (see above quotes from Epstein). Thus, the claim that “there are nonresponders to exercise” makes no sense, on the basis that people necessarily respond physiologically to the stressors placed on them, and so, if they do more (and they will) than they did previously from their baseline, then they did adapt to the protocol, implying that they are not “nonresponders” to exercise. It does not matter if Person B does not catch up to Person A on all variables: the fact that there was a difference in each individual from the baseline all the way to 6 months on a specific regimen implies adaptation to the stressors—which implies that there is no such thing “nonresponders”.
JayMan also has views similar to this, which I have responded to last year in the articles Diet and Exercise: Don’t Do It? and Diet and Exercise: Don’t Do It? Part II. Eating well and exercising—although benefits are not the same for each individual (and I do not know who made the claim this was the case)—does ameliorate numerous diseases and can extend lifespan, contrary to the results of certain studies (e.g., the Look AHEAD study; Annuzzi et al 2014).
Claims from people like JayMan who do not know the first thing about dieting and exercise are dangerous—though, all one has to do is have a basic understanding of physiology to understand that the claim “a fair fraction of people respond little to even *negatively* to exercise” is false, since everyone who does something for the first few times will ALWAYS be better in the months after learning the specific movement, implying that there are no nonresponders to exercise.
Of course everyone does not respond the same to exercise regimen A. Other studies found that increasing the frequency, reps, and set scheme lead to changes in the so-called “nonresponders.” Different individuals respond differently to different training programs [be it, strength, conditioning, cardio, plyometrics, balance, and stabilization etc. But it must be stressed that, although not everyone has the same potential for muscle-building/strength-gaining as, say, the IFBB pros or strongmen/powerlifters, everyone can and does benefit from NOT being sedentary, that much is most definitely clear. These studies that show “nonresponders” run people through the same exercise regimen. Anyone with an iota of experience in this industry knows that people do not respond the same to any and every exercise regimen and, so, the program must be tailored to that specific individual. Though, people like JayMan read this stuff and, without understanding what they’re talking about, jump to brash conclusions that are not supported by reality.
Gene-Selectionism vs. Developmental Systems Theory
2300 words
Two dominant theories exist in regard to development, the “gene’s eye view—gene selectionism (GS)—and the developmental view—developmental systems theory (DST). GS proposes that there are two fundamental processes in regard to evolution: replication and interaction. Replicators (the term was coined by Dawkins) are anything that is copied into the next generation whereas interactors (vehicles) are things that only exist to ensure the replicators’ survival. Thus, Dawkins (1976) proposes a distinction between the “vehicle” (organism) and its “riders/replicators” (the genes).
Gene selectionism
Gene selectionists propose a simple hypothesis: evolution through the differential survival of genes, its main premise being that the “gene” is “the ultimate, fundamental unit of natural selection.” Dusek (1999: 156) writes that “Gene selection claims that genes, not organisms, groups of organisms or species, are selected. The gene is considered to be the unit of selection.” The view of gene selectionists is best—and most popularly put—by Richard Dawkins’ seminal book The Selfish Gene (1976), in which he posits that genes “compete” with each other, and that our “selfish actions” are the result of our genes attempting to replicate to the next generation, relegating our bodies to disposable “vehicles” that only house the “replicators” (or “drivers).
Though, just because one is a gene selectionist does not necessarily mean that they are a genetic determinist (both views will be argued below). Gene selectionists are comitted to the view that genes make a distinctive contribution toward building interactors. Dawkins (1982) claims that genetic determinism is not a problem in regard to gene selectionism. Replicators (genes) have a special status to gene selectionists. Gene selectionists argue that adaptive evolution only occurs through cumulative selection, while only the replicators persist through the generations. Gene selectionists do not see organisms as replicators since genes—and not organisms—are what is replicated according to the view.
The gene selectionist view (Dawkins’ 1976 view) can also be said to apply what Okasha (2018) terms “agential thinking”. “Agential thinking” is “treating an evolved organism as if it were an agent pursuing a goal, such as survival or reproduction, and treating its phenotypic traits, including its behaviour, as strategies for achieving that goal, or furthering its biological interests” (Okasha, 2018: 12). Dawkins—and other gene selectionists—treat genes as if they have agency, speaking of “intra-genomic conflict”, as if genes are competing with each other (sure, it’s “just a metaphor”, see below).
Okasha (2018: 71) writes:
To see how this distinction relates to agential thinking, note that every gene is necessarily playing a zero-sum game against other alleles at the same locus: it can only spread in the population if they decline. Therefore every gene, including outlaws, can be thought of as ‘trying’ to outcompete alleles, or having this as its ultimate goal.
Selfish genes also have intermediate goals, which are to maximize fitness, which is done through expression in the organismic phenotype.
Thus, according to Okasha (2018: 73), “… selfish genetic elements have phenotypic effects which can be regarded as adaptations, but only if we apply the notions of agent, benefit, and goal to genes themselves”, though “… only in an evolutionary context [does] it [make] sense to treat genes as agent-like and credit them with goals and interests.” It does not “make sense to treat genes as even “agent-like and credit them with goals and interests since they can only be attributed to humans.
Other genes have as their intermediate goal to enhance the fitness of their host organism’s relatives, by causing altruistic behaviour [genes can’t cause altruistic behavior; it is an action]. However, a small handful of genes have a different intermediate goal, namely to increase their own transmission in their host organism’s gametes, for example, by biasing segregation in their favour, or distorting the sex-ratio, or transposing to new sites in the genome. These are outlaws, or selfish genetic elements.If oulaws are absent or are effectively suppressed, then the genes within a single organism have a common (intermediate) goal, so will cooperate: each gene can onluy benefit by itself by benefiting the whole organism. Agential thinking then can be applied to the organism itself. The organism’s goal—maximizing its fitness—then equates to the intermediate goal of each of the genes within it. (Okasha, 2018: 72)
Attributing agential thinking to anything other than humans is erroneous, since genes are not “selfish.”
The selfish gene is one of the main theories that define the neo-Darwinian paradigm and it is flat out wrong. Genes are not ultimate causes, as the crafters of the neo-Darwinian Modern Synthesis (MS) propose, genes are resources in a dynamic system and can thusly only be seen as causes in a passive, not active, sense (Noble, 2011).
Developmental systems
The alternative to the gene-centric view of evolution is that of developmental systems theory (DST), first proposed by Oyama (1985).
The argument for DST is simple:
(1) Organisms obviously inherit more than DNA from their parents. Since organisms can behave in ways that alter the environment, environments are also passed onto offspring. Thus, it can be said that genes are not the only things inherited, but a whole developmental matrix is.
(2) Genes, according to the orthodox view of the MS, interact with many other factors for development to occur, and so genes are not the only thing that help ‘build’ the organism. Genes can still play some “privileged” role in development, in that they “control”, “direct” or “organize” everything else, but this is up to gene-selectionists to prove. (See Noble, 2012.)
(3) The common claim that genes contain “information” (that is, context-independent information) is untenable, since every reconstruction of genes contain development about information applies directly to all other developmental outcomes. Genes cannot be singled out as privileged causes in development.
(4) Other attempts—such as genes are copied more “directly—are mistaken, since they draw a distinction between development and other factors but fail.
(5) Genes, then, cannot be privileged in development, and are no different than any other developmental factor. Genes, in fact, are just passive templates for protein construction, waiting to be used by the system in a context-dependent fashion (see Moore, 2002; Schneider, 2007). The entire developmental system reconstructs itself “through numerous independent causal pathways” (Sterelny and Griffiths, 1999: 109).
DNA is not the only thing inherited, and the so-called “famed immortality of DNA is actually a property of cells [since] [o]nly cells have the machinery to correct frequent faults that occur in DNA replication.” The thing about replication, though, is that “DNA and the cell must replicate together” (Noble, 2017: 238). A whole slew of developmental tools are inherited and that is what constructs the organism; organisms are, quite obviously, constructed not by genes alone.
Developmental systems, as described by Oyama (1985: 49) do not “have a final form, encoded before its starting point and realized at maturity. It has, if one focuses finely enough, as many forms as time has segments.” Oyama (1985: 61) further writes that “The function of the gene or any other influence can be understood only in relation to the system in which they are involved. The biological relevance or any influence, and therefore the very “information” it conveys, is jointly determined, frequently in a statistically interactive, not additive, manner, by that influence and the system state it influences.”
DNA is, of course, important. For without it, there would be nothing for the cell to read (recall how the genome is an organ of the cell) and so no development would occur. DNA is only “information” about an organism only in the process of cellular functioning.
The simple fact of the matter is this: the development of organs and tissues are not directly “controlled” by genes, but by the exchange signals of the cells. “Details notwithstanding, what is important to note is that whatever kinds of signals it sends out depends on the kind of signals it receives from its immediate environment. Therefore, neighboring cells are interdependent, and its local interactions among cells that drive the developmental processes” (Kampourakis, 2017: 173).
The fact of the matter is that whether or not a trait is realized depends on the developmental processes (and the physiologic system itself) and the environment. Kampourakis, just like Noble (2006, 2012, 2017) pushes a holistic view of development and the system. Kampourakis (2017: 184) writes:
What genetics research consistently shows is that biological phenomena should be approached holistically. at various levels. For example, as genes are expressed and produce proteins, and some of these proteins regulate or affect gene expression, there is absolutely no reason to privilege genes over proteins. This is why it is important to consider developmental processes in order to undertand how characters and disease arise. Genes cannot be considered alone but only in the broader context (cellular, organismal, environmental) in which they exist. And both characters and disease in fact develop; they are not just produced. Therefore, reductionism, the idea that genes provide the ultimate explanation for characters and disease, is also wrong. In order to understand such phenomena, we need to consider influence at various levels of organization, both bottom-up and top-down. This is why current research has adopted a systems biology approach (see Noble, 2006; Voit, 2016 for accessible introductions).
All this shows that developmental processes and interactions play a major role in shaping characters. Organisms can respond to changing environments through changes in their development and eventually their phenotypes. Most interestingly, plastic responses of this kind can become stable and inherited by their offspring. Therefore, genes do not predetermine phenotypes; genes are implicated in the development of phenotypes only through their products, which depends on what else is going on within and outside cells (Jablonka, 2013). It is therefore necessary to replacr the common representation of gene function presented in Figure 9.6a, which we usually find in the public sphere, with others that consider development, such as the one in figure 9.6b. Genes do not determine characters, but they are implicated in their development. Genes are resources that provide cells with a generative plan about the development of the organism, and have a major role in this process through their products. This plan is the resouce for the production of robust developmental outcomes that are at the same time plastic enough to accomodate changes stemming from environmental signals.
Figure 9.6 (a) The common representation of gene function: a single gene determines a single phenotype. It should be clear by what has been present in the book so far that is not accurate. (b) A more accurate representation of gene function that takes development and environment into account. In this case, a phenotype is propduced in a particular environment by developmental processes in which genes are implicated. In a different environment the same genes might contribute tothe development of a different phenotype. Note the “black box” of development.
[Kampourakis also writes on page 188, note 3]
In the original analogy, Wolpert (2011, p. 11) actually uses the term “program.” However, I consider the term “plan” as more accurate and thus more appropriate. In my view, the term “program” impies instructions and their implimentation, whereas the term “plan” is about instructions only. The notion of a genetic program can be very misleading because it implies that, if it were technically feasible, it would be possible to compute an organism by reading the DNA sequence alone (see Lewontin, 2000, pp. 17-18).
Kampourakis is obviously speaking of a “plan” in a context-dependent manner since that is the only way that genes/DNA contain “information” (Moore, 2002; Schneider, 2007). The whole point is that genes, to use Noble’s terminology, are “slaves” to the system, since they are used by and for the (physiological) system. Developmental systems theory is a “wholeheartedly epigenetic approach to development, inheritance and evolution” (Hochman and Griffiths, 2015).
This point is driven home by Richardson (2017:111):
And how did genes eventually become established? Probably not at all as the original recipes, designers, and controllers of life. Instead they arose as templates for molecular components used repeatedly in the life of the cell and the organism: a kind of facility for just-in-time production of parts needed on a recurring basis. Over time, of course, the role of these parts themselves evolved to become key players in the metabolism of the call—but as part of a team, not the boss.
[…]
It is not surprising, then, that we find that variation in form and function has, for most traits, only a tenuous relationship with variation in genes.
[And also writes on page 133]:
There is no direct command line between environments and genes or between genes and phenotypes. Predictions and decisions about form and variation are made through a highly evolved dynamical system. That is why ostensibly the same environment, such as hormonal signal, can initiate a variaety of responses like growth, cell division, differentiation, and migration, depending on deeper context. This reflects more than fixes responses from fixed information in genes, something fatally overlooked in the nature-nurture debate
(Also read Richardson’s article So what is a gene?)
Conclusion
The gene-selectionist point-of-view entails too many (false) assumptions. The DST point of view, on the other hand, does not fall prey to the pitfalls of the gene-selectionist POV; Developmental systems theorists look at the gene, not as the ultimate causes of development—and, along with that, only changes in gene frequency driving evolutionary change—but only as products to be used by and for the system. Genes can only be looked at in terms of development, and in no other way (Kamporuakis, 2017; Noble, 2017). Thus, the gene-selectionists are wrong; the main tenet of the neo-Darwinian Modern Synthesis, gene-selectionism—the selfish gene—has been refuted (Jablonka and Lamb, 2005; Noble, 2006, 2011). The main tenets of the neo-Darwinian Modern Synthesis have been refuted, and so it is now time to replace the Modern Synthesis with a new view of evolution: one that includes the role of genes and development and the role of epigenetics on the developmental system. The gene-selectionist view champions an untenable view of the gene: that the gene is priviliged above any other developmental variables, but Noble and Kampourakis show that this is not the case, since DNA is inherited with the cell; the cell is what is “immortal” to use the language of Dawkins—not DNA itself.
A priori, there is no privileged level of causation, and this includes the gene, which so many place at the top of the hierarchy (Noble, 2012).
