
2300 words
Two dominant theories exist in regard to development, the “gene’s eye view—gene selectionism (GS)—and the developmental view—developmental systems theory (DST). GS proposes that there are two fundamental processes in regard to evolution: replication and interaction. Replicators (the term was coined by Dawkins) are anything that is copied into the next generation whereas interactors (vehicles) are things that only exist to ensure the replicators’ survival. Thus, Dawkins (1976) proposes a distinction between the “vehicle” (organism) and its “riders/replicators” (the genes).
Gene selectionism
Gene selectionists propose a simple hypothesis: evolution through the differential survival of genes, its main premise being that the “gene” is “the ultimate, fundamental unit of natural selection.” Dusek (1999: 156) writes that “Gene selection claims that genes, not organisms, groups of organisms or species, are selected. The gene is considered to be the unit of selection.” The view of gene selectionists is best—and most popularly put—by Richard Dawkins’ seminal book The Selfish Gene (1976), in which he posits that genes “compete” with each other, and that our “selfish actions” are the result of our genes attempting to replicate to the next generation, relegating our bodies to disposable “vehicles” that only house the “replicators” (or “drivers).
Though, just because one is a gene selectionist does not necessarily mean that they are a genetic determinist (both views will be argued below). Gene selectionists are comitted to the view that genes make a distinctive contribution toward building interactors. Dawkins (1982) claims that genetic determinism is not a problem in regard to gene selectionism. Replicators (genes) have a special status to gene selectionists. Gene selectionists argue that adaptive evolution only occurs through cumulative selection, while only the replicators persist through the generations. Gene selectionists do not see organisms as replicators since genes—and not organisms—are what is replicated according to the view.
The gene selectionist view (Dawkins’ 1976 view) can also be said to apply what Okasha (2018) terms “agential thinking”. “Agential thinking” is “treating an evolved organism as if it were an agent pursuing a goal, such as survival or reproduction, and treating its phenotypic traits, including its behaviour, as strategies for achieving that goal, or furthering its biological interests” (Okasha, 2018: 12). Dawkins—and other gene selectionists—treat genes as if they have agency, speaking of “intra-genomic conflict”, as if genes are competing with each other (sure, it’s “just a metaphor”, see below).
Okasha (2018: 71) writes:
To see how this distinction relates to agential thinking, note that every gene is necessarily playing a zero-sum game against other alleles at the same locus: it can only spread in the population if they decline. Therefore every gene, including outlaws, can be thought of as ‘trying’ to outcompete alleles, or having this as its ultimate goal.
Selfish genes also have intermediate goals, which are to maximize fitness, which is done through expression in the organismic phenotype.
Thus, according to Okasha (2018: 73), “… selfish genetic elements have phenotypic effects which can be regarded as adaptations, but only if we apply the notions of agent, benefit, and goal to genes themselves”, though “… only in an evolutionary context [does] it [make] sense to treat genes as agent-like and credit them with goals and interests.” It does not “make sense to treat genes as even “agent-like and credit them with goals and interests since they can only be attributed to humans.
Other genes have as their intermediate goal to enhance the fitness of their host organism’s relatives, by causing altruistic behaviour [genes can’t cause altruistic behavior; it is an action]. However, a small handful of genes have a different intermediate goal, namely to increase their own transmission in their host organism’s gametes, for example, by biasing segregation in their favour, or distorting the sex-ratio, or transposing to new sites in the genome. These are outlaws, or selfish genetic elements.If oulaws are absent or are effectively suppressed, then the genes within a single organism have a common (intermediate) goal, so will cooperate: each gene can onluy benefit by itself by benefiting the whole organism. Agential thinking then can be applied to the organism itself. The organism’s goal—maximizing its fitness—then equates to the intermediate goal of each of the genes within it. (Okasha, 2018: 72)
Attributing agential thinking to anything other than humans is erroneous, since genes are not “selfish.”
The selfish gene is one of the main theories that define the neo-Darwinian paradigm and it is flat out wrong. Genes are not ultimate causes, as the crafters of the neo-Darwinian Modern Synthesis (MS) propose, genes are resources in a dynamic system and can thusly only be seen as causes in a passive, not active, sense (Noble, 2011).
Developmental systems
The alternative to the gene-centric view of evolution is that of developmental systems theory (DST), first proposed by Oyama (1985).
The argument for DST is simple:
(1) Organisms obviously inherit more than DNA from their parents. Since organisms can behave in ways that alter the environment, environments are also passed onto offspring. Thus, it can be said that genes are not the only things inherited, but a whole developmental matrix is.
(2) Genes, according to the orthodox view of the MS, interact with many other factors for development to occur, and so genes are not the only thing that help ‘build’ the organism. Genes can still play some “privileged” role in development, in that they “control”, “direct” or “organize” everything else, but this is up to gene-selectionists to prove. (See Noble, 2012.)
(3) The common claim that genes contain “information” (that is, context-independent information) is untenable, since every reconstruction of genes contain development about information applies directly to all other developmental outcomes. Genes cannot be singled out as privileged causes in development.
(4) Other attempts—such as genes are copied more “directly—are mistaken, since they draw a distinction between development and other factors but fail.
(5) Genes, then, cannot be privileged in development, and are no different than any other developmental factor. Genes, in fact, are just passive templates for protein construction, waiting to be used by the system in a context-dependent fashion (see Moore, 2002; Schneider, 2007). The entire developmental system reconstructs itself “through numerous independent causal pathways” (Sterelny and Griffiths, 1999: 109).
DNA is not the only thing inherited, and the so-called “famed immortality of DNA is actually a property of cells [since] [o]nly cells have the machinery to correct frequent faults that occur in DNA replication.” The thing about replication, though, is that “DNA and the cell must replicate together” (Noble, 2017: 238). A whole slew of developmental tools are inherited and that is what constructs the organism; organisms are, quite obviously, constructed not by genes alone.
Developmental systems, as described by Oyama (1985: 49) do not “have a final form, encoded before its starting point and realized at maturity. It has, if one focuses finely enough, as many forms as time has segments.” Oyama (1985: 61) further writes that “The function of the gene or any other influence can be understood only in relation to the system in which they are involved. The biological relevance or any influence, and therefore the very “information” it conveys, is jointly determined, frequently in a statistically interactive, not additive, manner, by that influence and the system state it influences.”
DNA is, of course, important. For without it, there would be nothing for the cell to read (recall how the genome is an organ of the cell) and so no development would occur. DNA is only “information” about an organism only in the process of cellular functioning.
The simple fact of the matter is this: the development of organs and tissues are not directly “controlled” by genes, but by the exchange signals of the cells. “Details notwithstanding, what is important to note is that whatever kinds of signals it sends out depends on the kind of signals it receives from its immediate environment. Therefore, neighboring cells are interdependent, and its local interactions among cells that drive the developmental processes” (Kampourakis, 2017: 173).
The fact of the matter is that whether or not a trait is realized depends on the developmental processes (and the physiologic system itself) and the environment. Kampourakis, just like Noble (2006, 2012, 2017) pushes a holistic view of development and the system. Kampourakis (2017: 184) writes:
What genetics research consistently shows is that biological phenomena should be approached holistically. at various levels. For example, as genes are expressed and produce proteins, and some of these proteins regulate or affect gene expression, there is absolutely no reason to privilege genes over proteins. This is why it is important to consider developmental processes in order to undertand how characters and disease arise. Genes cannot be considered alone but only in the broader context (cellular, organismal, environmental) in which they exist. And both characters and disease in fact develop; they are not just produced. Therefore, reductionism, the idea that genes provide the ultimate explanation for characters and disease, is also wrong. In order to understand such phenomena, we need to consider influence at various levels of organization, both bottom-up and top-down. This is why current research has adopted a systems biology approach (see Noble, 2006; Voit, 2016 for accessible introductions).
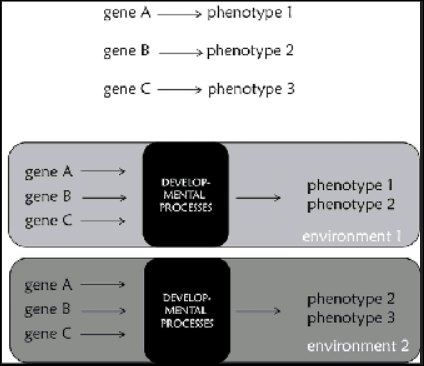
All this shows that developmental processes and interactions play a major role in shaping characters. Organisms can respond to changing environments through changes in their development and eventually their phenotypes. Most interestingly, plastic responses of this kind can become stable and inherited by their offspring. Therefore, genes do not predetermine phenotypes; genes are implicated in the development of phenotypes only through their products, which depends on what else is going on within and outside cells (Jablonka, 2013). It is therefore necessary to replacr the common representation of gene function presented in Figure 9.6a, which we usually find in the public sphere, with others that consider development, such as the one in figure 9.6b. Genes do not determine characters, but they are implicated in their development. Genes are resources that provide cells with a generative plan about the development of the organism, and have a major role in this process through their products. This plan is the resouce for the production of robust developmental outcomes that are at the same time plastic enough to accomodate changes stemming from environmental signals.
Figure 9.6 (a) The common representation of gene function: a single gene determines a single phenotype. It should be clear by what has been present in the book so far that is not accurate. (b) A more accurate representation of gene function that takes development and environment into account. In this case, a phenotype is propduced in a particular environment by developmental processes in which genes are implicated. In a different environment the same genes might contribute tothe development of a different phenotype. Note the “black box” of development.
[Kampourakis also writes on page 188, note 3]
In the original analogy, Wolpert (2011, p. 11) actually uses the term “program.” However, I consider the term “plan” as more accurate and thus more appropriate. In my view, the term “program” impies instructions and their implimentation, whereas the term “plan” is about instructions only. The notion of a genetic program can be very misleading because it implies that, if it were technically feasible, it would be possible to compute an organism by reading the DNA sequence alone (see Lewontin, 2000, pp. 17-18).
Kampourakis is obviously speaking of a “plan” in a context-dependent manner since that is the only way that genes/DNA contain “information” (Moore, 2002; Schneider, 2007). The whole point is that genes, to use Noble’s terminology, are “slaves” to the system, since they are used by and for the (physiological) system. Developmental systems theory is a “wholeheartedly epigenetic approach to development, inheritance and evolution” (Hochman and Griffiths, 2015).
This point is driven home by Richardson (2017:111):
And how did genes eventually become established? Probably not at all as the original recipes, designers, and controllers of life. Instead they arose as templates for molecular components used repeatedly in the life of the cell and the organism: a kind of facility for just-in-time production of parts needed on a recurring basis. Over time, of course, the role of these parts themselves evolved to become key players in the metabolism of the call—but as part of a team, not the boss.
[…]
It is not surprising, then, that we find that variation in form and function has, for most traits, only a tenuous relationship with variation in genes.
[And also writes on page 133]:
There is no direct command line between environments and genes or between genes and phenotypes. Predictions and decisions about form and variation are made through a highly evolved dynamical system. That is why ostensibly the same environment, such as hormonal signal, can initiate a variaety of responses like growth, cell division, differentiation, and migration, depending on deeper context. This reflects more than fixes responses from fixed information in genes, something fatally overlooked in the nature-nurture debate
(Also read Richardson’s article So what is a gene?)
Conclusion
The gene-selectionist point-of-view entails too many (false) assumptions. The DST point of view, on the other hand, does not fall prey to the pitfalls of the gene-selectionist POV; Developmental systems theorists look at the gene, not as the ultimate causes of development—and, along with that, only changes in gene frequency driving evolutionary change—but only as products to be used by and for the system. Genes can only be looked at in terms of development, and in no other way (Kamporuakis, 2017; Noble, 2017). Thus, the gene-selectionists are wrong; the main tenet of the neo-Darwinian Modern Synthesis, gene-selectionism—the selfish gene—has been refuted (Jablonka and Lamb, 2005; Noble, 2006, 2011). The main tenets of the neo-Darwinian Modern Synthesis have been refuted, and so it is now time to replace the Modern Synthesis with a new view of evolution: one that includes the role of genes and development and the role of epigenetics on the developmental system. The gene-selectionist view champions an untenable view of the gene: that the gene is priviliged above any other developmental variables, but Noble and Kampourakis show that this is not the case, since DNA is inherited with the cell; the cell is what is “immortal” to use the language of Dawkins—not DNA itself.
A priori, there is no privileged level of causation, and this includes the gene, which so many place at the top of the hierarchy (Noble, 2012).

what is the developmental system? you mean multiple interaction levels? the post doesn’t define what the developmental system is, only that it is what replicates itself. you say “the system” uses genes to replicate itself but what is the system? i think you mean the entire process (in the same way, say, societies “replicate” themselves by creating a new generation of members)?
i don’t think you really deal well with the “it’s a metaphor” argument – it refers to the statistical process, not that genes have minds and will. genes that don’t statistically proliferate will just disappear. e.g eusocial species have most members reproduce indirectly.
LikeLike