
Is Racial Superiority in Sports a Myth? A Response to Kerr (2010)
2750 words
Racial differences in sporting success are undeniable. The races are somewhat stratified in different sports and we can trace the cause of this to differences in genes and where one’s ancestors were born. We can then say that there is a relationship between them since, they have certain traits which their ancestors also had, which then correlate with geographic ancestry, and we can explain how and why certain populations dominate (or would have the capacity to based on body type and physiology) certain sporting events. Critiques of Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It are few and far between, and the few that I am aware of are alright, but this one I will discuss today is not particularly good, because the author makes a lot of claims he could have easily verified himself.
In 2010, Ian Kerr published The Myth of Racial Superiority in Sports, who states that there is a “dark side” to sports, and specifically sets his sights on Jon Entine’s (2000) book Taboo. In this article, Kerr (2010) makes a lot of, in my opinion, giant claims which provide a lot of evidence and arguments in order to show their validity. I will discuss Kerr’s views on race, biology, the “environment”, “genetic determinism”, and racial dominance in sports (which will have a focus on sprinting/distance running in this article).
Race
Since establishing the reality and validity of the concept of race is central to proving Entine’s (2002) argument on racial differences in sports, then I must prove the reality of race (and rebut what Kerr 2010 writes about race). Kerr (2010: 20) writes:
First, it is important to note that Entine is not working in a vacuum; his assertions about race and sports are part of a larger ongoing argument about folk notions of race. Folk notions of race founded on the idea that deep, mutually exclusive biological categories dividing groups of people have scientific and cultural merit. This type of thinking is rooted in the notion that there are underlying, essential differences among people and that those observable physical differences among people are rooted in biology, in genetics (Ossorio, Duster, 2005: 2).
Dividing groups of people does have scientific, cultural and philosophical merit. The concept of “essences” has long been discarded by philosophers. Though there are differences in both anatomy and physiology in people that differ by geographic location, and this then, at the extreme end, would be enough to cause the differences in elite sporting competition that is seen.
Either way, the argument for the existence of race is simple: 1) populations differ in physical attributes (facial, morphological) which then 2) correlate with geographic ancestry. Therefore, race has a biological basis since the physical differences between these populations are biological in nature. Now that we have established that race exists using only physical features, it should be extremely simple to show how Kerr (2010) is in error with his strong claims regarding race and the so-called “mythology” of racial superiority in sports. Race is biological; the biological argument for race is sound (read here and here, and also see Hardimon, 2017).
Genetic determinism
True genetic determinism—as is commonly thought—does not have any sound, logical basis (Resnick and Vorhaus, 2006). So Kerr’s (2010) claims in this section need to be dissected here. This next quote, though, is pretty much imperative to the soundness and validity of his whole article, and let’s just say that it’s easy to rebut and invalidates his whole entire argument:
Vinay Harpalani is one of the most outspoken critics of using genetic determinism to validate notions of inferiority or the superiority of certain groups (in this case Black athletes). He argues that in order for any of Entine’s claims to be valid he must prove that: 1) there is a systematic way to define Black and White populations; 2) consistent and plausible genetic differences between the populations can be demonstrated; 3) a link between those genetic differences and athletic performance can be clearly shown (2004).
This is too easy to prove.
1) While I do agree that the terminology of ‘white’ and ‘black’ are extremely broad, as can be seen by looking at Rosenberg et al (2002), population clusters that cluster with what we call ‘white’ and ‘black’ exist (and are a part of continental-level minimalist races). So is there a systematic way to define ‘Black’ and ‘White’ populations? Yes, there is; genetic testing will show where one’s ancestors came from recently, thereby proving point 1.
2) Consistent and plausible genetic differences between populations can be demonstrated. Sure, there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016). Even these small between-continent/group differences would have huge effects on the tail end of said distribution.
3) I have compiled numerous data on genetic differences between African ethnies and European ethnies and how these genetic differences then cause differences in elite athletic performance. I have shown that Jamaicans, West Africans, Kenyans and Ethiopians (certain subgroups of the two aforementioned countries) have genetic/somatypic differences that then lead to differences in these sporting competitions. So we can say that race can predict traits important for certain athletic competitions.
1) The terminology of ‘White’ and ‘Black’ are broad; but we can still classify individuals along these lines; 2) consistent and plausible genetic differences between races and ethnies do exist; 3) a link between these genetic differences between genes/athletic differences between groups can be found. Therefore Entine’s (2002) arguments—and the validity thereof—are sound.
Kerr (2010) then makes a few comments on the West’s “obsession with superficial physical features such as skin color”, but using Hardimon’s minimalist race concept, skin color is a part of the argument to prove the existence and biological reality of race, therefore skin color is not ‘superficial’, since it is also a tell of where one’s ancestors evolved in the recent past. Kerr (2010: 21) then writes:
Marks writes that Entine is saying one of three things: that the very best Black athletes have an inherent genetic advantage over the very best White athletes; that the average Black athlete has a genetic advantage over the average White athlete; that all Blacks have the genetic potential to be better athletes than all Whites. Clearly these three propositions are both unknowable and scientifically untenable. Marks writes that “the first statement is trivial, the secondly statistically intractable, and the third ridiculous for its racial essentialism” (Marks, 2000: 1077).
The first two, in my opinion (the very best black athletes have an inherent genetic advantage over the very best white athletes and the average black athlete has a genetic advantage over the average white athlete), are true, and I don’t know how you can deny this; especially if you’re talking about AVERAGES. The third statement is ridiculous, because it doesn’t work like that. Kerr (2010), of course, states that race is not a biological reality, but I’ve proven that it is so that statement is a non-factor.
Kerr (2010) then states that “ demonstrating across the board genetic variations between
populations — has in recent years been roundly debunked“, and also says “ Differences in height, skin color, and hair texture are simply the result of climate-related variation.” This is one of the craziest things I’ve read all year! Differences in height would cause differences in elite sporting competition; differences in skin color can be conceptualized as one’s ancestors’ multi-generational adaptation to the climate they evolved in as can hair texture. If only Kerr (2010) knew that this statement here was the beginning of the end of his shitty argument on Entine’s book. Race is a social construct of a biological reality, and there are genetic differences between races—however small (Risch et al, 2002; Tang et al, 2005) but these small differences can mean big differences at the elite level.
The “environment” and biological variability
Kerr (2010) then shifts his focus over to, not genetic differences, but biological differences. He specifically discusses the Kenyans—Kalenjin—stating that “height or weight, which play an instrumental role in helping define an individual’s athletic prowess, have not been proven to be exclusively rooted in biology or genetics.” While estimates of BMI and height are high (both around .8), I think we can disregard the numbers since they came from highly flawed twin studies, since molecular genetic evidence shows lower heritabilities. Either way, surely height is strongly influenced by ‘genes’. Another important caveat is that Kenya has one of the lowest BMIs in the world, 20.7 for Kenyan men, which also is part of the cause of why certain African ethnies dominate running competitions.
I don’t disagree with Kerr (2010) here too much; many papers show that SES/cultural/social factors are very important to Kenyan runners (Onywera et al, 2006; Wilbur and Pistiladis, 2012; Tucker, Onywera, and Santos-Concejero, 2015). You can have all of the ‘physical gifts’ in the world, if it’s not combined with the will to want to do your best, along with cultural and social factors you won’t succeed. But having an advantageous genotype and physique are useless without a strong mind (Lippi, Favaloro, and Guidi, 2008):
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.”
Dissecting this, though, is tougher. Because being born at certain altitudes will cause certain advantageous traits, such as a larger lung capacity (and you will have an advantage in lung capacity when competing at lower altitudes), but certain subpopulations live in these high-altitude areas, so what is it? Genetic? Cultural? Environmental? All three? Nature vs nurture is a false dichotomy; so it is a mixture of the three.
How does one explain, then, the athlete who trains countless hours a day fine-tuning a jump shot, like LeBron James or shaving seconds off sub-four minute miles like Robert Kipkoech Cheruiyot, a four time Boston Marathon winner?
Literally no one denies that elite athletes put in insane amounts of practice; but if everyone has the same amount of practice they won’t have similar abilities.
He also briefly brings up muscle fibers, stating:
These include studies on African fast twitch muscle fibers and development of motor skills. Entine includes these studies to demonstrate irrevocable proof of embedded genetic differences between populations but refuses to accept the fact that any differences may be due to environmental factors or training.
This, again, shows ignorance of the literature. An individual’s muscle fibers are formed during development from the fusion of several myoblasts, with differentiation being completed before birth. Muscle fiber typoing is also set at age 6, no difference in skeletal muscle tissue was found when comparing 6-year-olds and adults, therefore we can state that muscle fiber typing is set by age 6 (Bell et al, 1980). You can, of course, train type II fibers to have similar aerobic capacity to type I fibers, but they’ll never be fully similar. This is something that Kerr (2010) obviously is ignorant to because he’s not well-read on the literature which causes him to make dumb statements like “any differences [in muscle fiber typing] may be due to environmental factors or training“.
Black domination in sports
Finally, Kerr (2010) discusses the fact that whites dominated certain running competitions in the Olympics and that before the 1960s, a majority of distance-running gold medals went to white athletes. He then states that the 2008 Boston Marathon winner was Kenyan; but the next 4 behind him were not. Now, let’s check out the 2017 Marathon winners: Kenya, USA, Japan for the top 3; while 5 Kenyans/Ethiopians are in the top 15 while the same is also true of women; a Kenyan winner, with Kenyans/Ethiopians taking 5 of the top 15 spots. The fact that whites used to do well in running sports is a non-factor; Jesse Owens blew away the competition in the Games in Germany, which showed how blacks would begin to dominate in the US decades later.
Kerr (2010) then ends the article with a ton of wild claims; the wildest one, in my opinion, being that “Kenyans are no more genetically different from any other African or European population on average“, does anyone believe this? Because I have data to the contrary. They have a higher Vo2 max, which of course is trainable but with a ‘genetic’ component (Larsen, 2003), while other authors argue that genetic differences between populations account for differences in success in running competition between populations (Vancini et al, 2014), while male and female Kenyan and Ethiopian runners are the fastest in the half and full marathon (Knechtle et al, 2016). There is a large amount of data out there that speaks about Kenyan/Ethiopian and others’ dominance in running; it seems Kerr (2010) just ignored the data. I agree with Kerr that Kenyanholos show that humans can adapt to their environment; but his conclusion here:
The fact that runners coming from Kenya do so well in running events attests to the fact the combination of intense high altitude training, consumption of a low-fat, high protein diet, and a social and cultural expectation to succeed have created in recent decades an environment which is highly conducive to producing excellent long-distance runners.
is very strong, and while I don’t disagree at all with anything here, he’s disregarding how somatype and genes differ between Kenyans and other populations that compete in these sports that then lead to differences in elite sporting competitions.
Elite sporting performance is influenced by myriad factors, including psychology, ‘environment’, and genetic factors. Something that Kerr (2010) doesn’t understand—because he’s not well-read on this literature—is that many genetic factors that influence sporting performance are known. The ability to become elite depends on one’s capacity for endurance, muscle performance, the ability of the tendons and ligaments to withstand stress and injury, and the attitude to train and push above and beyond what normal people can do (Lippi, Longo, and Maffulli, 2010). We can then extend this to human races; some are better-equipped to excel in running competitions than others.
On its face, Kerr’s (2010) claim that there are no inherent differences between races is wrong. Races differ in somatype, which is due to evolution in different geographic locations for tens of thousands of years. The human body is perfectly adapted to for long distance running (Murray and Costa, 2012), and since our capabilities for endurance running evolved in Africa and they, theoretically, have a musculoskeletal structure similar to the Homo sapiens that left Africa around 70 kya, then it’s only logical to state that African’s, on average, have an inherent ability in running competitions (West and East Africans, while North Africans fare very well in middle distance running, which, again, comes down to living in higher altitudes like Kenyans and Ethiopians).
Wagner and Heyward (2000) reviewed many studies on the physiological differences between blacks and whites. Blacks skew towards mesomorphy; black youths had smaller billiac and bitrochanteric width (the widest measure of the pelvis at the outer edges and the flat process on the femur, respectively), and black infants had longer extremities than white infants (Wagner and Heyward, 2000). We have anatomic evidence that blacks are superior runners (in an American context). Mesomorphic athletes are more likely to be sprinters (Sands et al, 2005; which is also seen in prepubescent children: Marta et al, 2013) Kenyans are ecto-dominant (Vernillo et al, 2013) which helps to explain their success at long-distance running. So just on only looking at the phenotype (a marker for race with geographic ancestry, proving the biological existence of race) we can confidently state, on average just by looking at an individual or a population, how they will fare in certain competitions.
Conclusion
Kerr’s (2010) arguments leave a ton to be desired. Race exists and is a biological reality. I don’t know why this paper got published since it was so full of errors; his arguments were not sound and much of the literature contradicts his claims. What he states at the end about Kenyans is not wrong at all, but to not even bring up genetic/biologic differences as a factor influencing their performance is dishonest.
Of course, a whole slew of factors, be they biological, cultural, psychological, genetic, socioeconomic, anatomic, physiologic etc influence sporting performance, but certain traits are more likely to be found in certain populations, and in the year 2018 we have a good idea of what influences elite sporting performance and what does not. It just so happens that these traits are unevenly distributed between populations, and the cause is evolution in differing climates in differing geographic locations.
Race exists and is a biological reality. Biological anatomic/physiological differences between these races then manifest themselves in elite sporting competition. The races differ, on average, in traits important for success in certain competitions. Therefore, race explains some of the variance in elite sporting competition.
Does Playing Violent Video Games Lead to Violent Behavior?
1400 words
President Trump was quoted the other day saying “We have to look at the Internet because a lot of bad things are happening to young kids and young minds and their minds are being formed,” Trump said, according to a pool report, “and we have to do something about maybe what they’re seeing and how they’re seeing it. And also video games. I’m hearing more and more people say the level of violence on video games is really shaping young people’s thoughts.” But outside of broad assertions like this—that playing violent video games cause violent behavior—does it stack up to what the scientific literature says about it? In short, no, it does not. (A lot of publication bias exists in this debate, too.) Why do people think that violent video games cause violent behavior? Mostly due to the APA and their broad claims with little evidence.
Just doing a cursory Google search of ‘violence in video games pubmed‘ brings up 9 journal articles, so let’s take a look at a few of those.
The first article is titled The Effect of Online Violent Video Games on Levels of Aggression by Hollingdale and Greitemeyer (2014). They took 101 participants and randomized them to one of four experimental conditions: neutral, offline; neutral online; (Little Big Planet 2) violent offline; and violent online video games (Call of Duty: Modern Warfare). After they played said games, they answered a questionnaire and then measured aggression using the hot sauce paradigm (Lieberman et al, 1999) to measure aggressive behavior. Hollingdale and Greitemeyer (2014) conclude that “this study has identified that increases in aggression are not more pronounced when playing a violent video game online in comparison to playing a neutral video game online.”
Staude-Muller (2011) finds that “it was not the consumption of violent video games but rather an uncontrolled pattern of video game use that was associated with increasing aggressive tendencies.” Przybylski, Ryan, and Rigby (2009) found that enjoyment, value, and desire to play in the future were strongly related to competence in the game. Players who were high in trait aggression, though, were more likely to prefer violent games, even though it didn’t add to their enjoyment of the game, while violent content lent little overall variance to the satisfactions previously cited.
Tear and Nielsen (2013) failed to find evidence that violent video game playing leads to a decrease in pro-social behavior (Szycik et al, 2017 also show that video games do not affect empathy). Gentile et al (2014) show that “habitual violent VGP increases long-term AB [aggressive behavior] by producing general changes in ACs [aggressive cognitions], and this occurs regardless of sex, age, initial aggressiveness, and parental involvement. These robust effects support the long-term predictions of social-cognitive theories of aggression and confirm that these effects generalize across culture.” The APA (2015) even states that “scientific research has demonstrated an association between violent video game use and both increases in aggressive behavior, aggressive affect, aggressive cognitions and decreases in prosocial behavior, empathy, and moral engagement.” How true is all of this, though? Does playing violent video games truly increase aggression/aggressive behavior? Does it have an effect on violence in America and shootings overall?
No.
Whitney (2015) states that the video-games-cause-violence paradigm has “weak support” (pg 11) and that, pretty much, we should be cautious before taking this “weak support” as conclusive. He concludes that there is not enough evidence to establish a truly causal connection between violent video game playing and violent and aggressive behavior. Cunningham, Engelstatter, and Ward (2016) tracked the sale of violent video games and criminal offenses after those games were sold. They found that violent crime actually decreased the weeks following the release of a violent game. Of course, this does not rule out any longer-term effects of violent game-playing, but in the short term, this is good evidence against the case of violent games causing violence. (Also see the PsychologyToday article on the matter.)
We seem to have a few problems here, though. How are we to untangle the effects of movies and other forms of violent media that children consume? You can’t. So the researcher(s) must assume that video games and only video games cause this type of aggression. I don’t even see how one can logically state that out of all other types of media that violent video games—and not violent movies, cartoons, TV shows etc—cause aggression/violent behavior.
Back in 2011, the Supreme Court case Brown vs. Entertainment Merchants Association concluding that since the effects on violent/aggressive behavior were so small and couldn’t be untangled from other so-called effects from other violent types of media. Ferguson (2015) found that violent video game playing had little effect on children’s mood, aggression levels, pro-social behavior or grades. He also found publication bias in this literature (Ferguson, 2017). Contrary to what those say about video games causing violence/aggressive behavior, video game playing was associated with a decrease in youth crime (Ferguson, 2014; Markey, Markey, and French, 2015 which is in line with Cunningham, Engelstatter, and Ward, 2016). You can read more about this in Ferguson’s article for The Conversation, along with his and others’ responses to the APA who state that violent video games cause violent behavior (with them stating that the APA is biased). (Also read a letter from 230 researchers on the bias in the APA’s Task Force on Violent Media.)
How would one actually untangle the effects of, say, violent video game playing and the effects of such other ‘problematic’ forms of media that also show aggression/aggressive acts towards others and actually pinpoint that violent video games are the culprit? That’s right, they can’t. How would you realistically control for the fact that the child grows up around—and consumes—so much ‘violent’ media, seeing others become violent around him etc; how can you logically state that the video games are the cause? Some may think it logical that someone who plays a game like, say, Call of Duty for hours on end a day would be more likely to be more violent/aggressive or more likely to commit such atrocities like school shootings. But none of these studies have ever come to the conclusion that violent video games may/will cause someone to kill or go on a shooting spree. It just doesn’t make sense. I can, of course, see the logic in believing that it would lead to aggressive behavior/lack of pro-social behavior (let’s say the kid played a lot of games and had little outside contact with people his age), but of course the literature on this subject should be enough to put claims like this to bed.
It’s just about impossible to untangle the so-called small effects of video games on violent/aggressive behavior from other types of media such as violent cartoons and violent movies. Who’s to say it’s not just the violent video games and not the violent movies and violent cartoons, too, that ’cause’ this type of behavior? It’s logically impossible to distinguish this, so therefore the small relationship between video games and violent behavior should be safely ignored. The media seems to be getting this right, which is a surprise (though I bet if Trump said the opposite—that violent video games didn’t cause violent behavior/shootings—that these same people would be saying that they do), but a broken clock is right twice a day.
So Trump’s claim (even if he didn’t outright state it) is wrong, along with anyone else who would want to jump in and attempt to say that video games cause violence. In fact, the literature shows a decrease in violence after games are released (Ferguson, 2014; Markey, Markey, and French, 2015; Cunningham, Engelstatter, and Ward, 2016). The amount of publication bias (also see Copenhaver and Ferguson, 2015 where they show how the APA ignores bias and methodological problems regarding these studies) in this field (Ferguson, 2017) should lead one to question the body of data we currently have, since studies that find an effect are more likely to get published than studies that find no effect.
Video games do not cause violent/aggressive behavior/school shootings. There is literally no evidence that they are linked to the deaths of individuals, and with the small effects noted on violent/aggressive behavior due to violent video game playing, we can disregard those claims. (One thing video games are good for, though, is improving reaction time (Benoit et al, 2017). The literature is strong here; playing these so-called “violent video games” such as Call of Duty improved children’s reaction time, so wouldn’t you say that these ‘violent video games’ have some utility?)
Lead, Race, and Crime
2500 words
Lead has many known neurological effects on the brain (regarding the development of the brain and nervous system) that lead to many deleterious health outcomes and negative outcomes in general. Including (but not limited to) lower IQ, higher rates of crime, higher blood pressure and higher rates of kidney damage, which have permanent, persistent effects (Stewart et al, 2007). Chronic lead exposure, too, can “also lead to decreased fertility, cataracts, nerve disorders, muscle and joint pain, and memory or concentration problems” (Sanders et al, 2009). Lead exposure in vitro, in infancy, and in childhood can also lead to “neuronal death” (Lidsky and Schneider, 2003). While epigenetic inheritance also plays a part (Sen et al, 2015). How do blacks and whites differ in exposure to lead? How much is the difference between the two races in America, and how much would it contribute to crime? On the other hand, China has high rates of lead exposure, but lower rates of crime, so how does this relationship play out with the lead-crime relationship overall? Are the Chinese an outlier or is there something else going on?
The effects of lead on the brain are well known, and numerous amounts of effort have been put into lowering levels of lead in America (Gould, 2009). Higher exposure to lead is also found in poorer, lower class communities (Hood, 2005). So since higher levels of lead exposure are found more often in lower-class communities, then blacks should have higher blood-lead levels than whites. This is what we find.
Blacks had a 27 percent higher concentration of lead in their tibia, while having significantly higher levels of blood lead, “likely because of sustained higher ongoing lead exposure over the decades” (Theppeang et al, 2008). Other data—coming out of Detroit—shows the same relationships (Haar et al, 1979; Talbot, Murphy, and Kuller, 1982; Lead poisoning in children under 6 jumped 28% in Detroit in 2016; also see Maqsood, Stanbury, and Miller, 2017) while lead levels in the water contribute to high levels of blood-lead in Flint, Michigan (Hanna-Attisha et al, 2016; Laidlaw et al, 2016). Cassidy-Bushrow et al (2017) also show that “The disproportionate burden of lead exposure is vertically transmitted (i.e., mother-to-child) to African-American children before they are born and persists into early childhood.”
Children exposed to lead have lower brain volumes as children, specifically in the ventrolateral prefrontal cortex, which is the same region of the brain that is impaired in antisocial and psychotic persons (Cecil et al, 2008). The community that was tested was well within the ‘safe’ range set by the CDC (Raine, 2014: 224), though the CDC says that there is no safe level of lead exposure. There is a large body of studies which show that there is no safe level of lead exposure (Needleman and Landrigan, 2004; Canfield, Jusko, and Kordas, 2005; Barret, 2008; Rossi, 2008; Abelsohn and Sanborn, 2010; Betts, 2012; Flora, Gupta, and Tiwari, 2012; Gidlow, 2015; Lanphear, 2015; Wani, Ara, and Usmani, 2015; Council on Environmental Health, 2016; Hanna-Attisha et al, 2016; Vorvolakos, Aresniou, and Samakouri, 2016; Lanphear, 2017). So the data is clear that there is absolutely no safe level of lead exposure, and even small effects can lead to deleterious outcomes.
Further, one brain study of 532 men who worked in a lead plant showed that those who had higher levels of lead in their bones had smaller brains, even after controlling for confounds like age and education (Stewart et al, 2008). Raine (2014: 224) writes:
The fact that the frontal cortex was particularly reduced is very interesting, given that this brain region is involved in violence. This lead effect was equivalent to five years of premature aging of the brain.
So we have good data that the parts of the brain that relate to violent tendencies are reduced in people exposed to more lead had the same smaller parts of the brain, indicating a relationship. But what about antisocial disorders? Are people with higher levels of lead in their blood more likely to be antisocial?
Needleman et al (1996) show that boys who had higher levels of lead in their blood had higher teacher ratings of aggressive and delinquent behavior, along with higher self-reported ratings of aggressive behavior. Even high blood-lead levels later in life is related to crime. One study in Yugoslavia showed that blood lead levels at age three had a stronger relationship with destructive behavior than did prenatal blood lead levels (Wasserman et al, 2008); with this same relationship being seen in America with high blood lead levels correlating with antisocial and aggressive behavior at age 7 and not age 2 (Chen et al 2007).
Nevin (2007) showed a strong relationship between preschool lead exposure and subsequent increases in criminal cases in America, Canada, Britain, France, Australia, Finland, West Germany, and New Zealand. Reyes (2007) also shows that crime increased quicker in states that saw a subsequent large decrease in lead levels, while variations in lead levels within cities correlating with variations in crime rates (Mielke and Zahran, 2012). Nevin (2000) showed a strong relationship between environmental lead levels from 1941 to 1986 and corresponding changes to violent crime twenty-three years later in the United States. Raine (2014: 226) writes (emphasis mine):
So, young children who are most vulnerable to lead absorption go on twenty-three years later to perpetrate adult violence. As lead levels rose throughout the 1950s, 1960s, and 1970s, so too did violence correspondingly rise in the 1970s, 1980s and 1990s. When lead levels fell in the late 1970s and early 1980s, so too did violence fall in the 1990s and the first decade of the twenty-first century. Changes in lead levels explained a full 91 percent of the variance in violent offending—an extremely strong relationship.
[…]
From international to national to state to city levels, the lead levels and violence curves match up almost exactly.
But does lead have a causal effect on crime? Due to the deleterious effects it has on the developing brain and nervous system, we should expect to find a relationship, and this relationship should become stronger with higher doses of lead. Fortunately, I am aware of one analysis, a sample that’s 90 percent black, which shows that with every 5 microgram increase in prenatal blood-lead levels, that there was a 40 percent higher risk of arrest (Wright et al, 2008). This makes sense with the deleterious developmental effects of lead; we are aware of how and why people with high levels of lead in their blood show similar brain scans/brain volume in certain parts of the brain in comparison to antisocial/violent people. So this is yet more suggestive evidence for a causal relationship.
Jennifer Doleac discusses three studies that show that blood-lead levels in America need to be addressed, since they are related strongly to negative health outcomes.Aizer and Curry (2017) show that “A one-unit increase in lead increased the probability of suspension from school by 6.4-9.3 percent and the probability of detention by 27-74 percent, though the latter applies only to boys.” They also show that children who live nearer to roads have higher blood-lead levels, since the soil near highways was contaminated decades ago with leaded gasoline. Fiegenbaum and Muller (2016) show that cities’ use of lead pipes increased murder rates between the years o921 and 1936. Finally, Billings and Schnepnel (2017: 4) show that their “results suggest that the effects of high levels of [lead] exposure on antisocial behavior can largely be reversed by intervention—children who test twice over the alert threshold exhibit similar outcomes as children with lower levels of [lead] exposure (BLL<5μg/dL).”
A relationship with lead exposure in vitro and arrests at adulthood. The sample was 90 percent black, with numerous controls. They found that prenatal and post-natal blood-lead exposure was associated with higher arrest rates, along with higher arrest rates for violent acts (Wright et al, 2008). To be specific, for every 5 microgram increase in prenatal blood-lead levels, there was a 40 percent greater risk for arrest. This is direct causal evidence for the lead-causes-crime hypothesis.
One study showed that in post-Katrina New Orleans, decreasing lead levels in the soil caused a subsequent decrease in blood lead levels in children (Mielke, Gonzales, and Powell, 2017). Sean Last argues that, while he believes that lead does contribute to crime, that the racial gaps have closed in the recent decades, therefore blood-lead levels cannot be a source of some of the variance in crime between blacks and whites, and even cites the CDC ‘lowering its “safe” values’ for lead, even though there is no such thing as a safe level of lead exposure (references cited above). White, Bonilha, and Ellis Jr., (2015) also show that minorities—blacks in particular—have higher rates of lead in their blood. Either way, Last seems to downplay large differences in lead exposure between whites and blacks at young ages, even though that’s when critical development of the mind/brain and other important functioning occurs. There is no safe level of lead exposure—pre- or post-natal—nor are there safe levels at adulthood. Even a small difference in blood lead levels would have some pretty large effects on criminal behavior.
Sean Last also writes that “Black children had a mean BLL which was 1 ug/dl higher than White children and that this BLL gap shrank to 0.9 ug/dl in samples taken between 2003 and 2006, and to 0.5 ug/dl in samples taken between 2007 and 2010.” Though, still, there are problems here too: “After adjustment, a 1 microgram per deciliter increase in average childhood blood lead level significantly predicts 0.06 (95% confidence interval [CI] = 0.01, 0.12) and 0.09 (95% CI = 0.03, 0.16) SD increases and a 0.37 (95% CI = 0.11, 0.64) point increase in adolescent impulsivity, anxiety or depression, and body mass index, respectively, following ordinary least squares regression. Results following matching and instrumental variable strategies are very similar” (Winter and Sampson, 2017).
Naysayers may point to China and how they have higher levels of blood-lead levels than America (two times higher), but lower rates of crime, some of the lowest in the world. The Hunan province in China has considerably lowered blood-lead levels in recent years, but they are still higher than developed countries (Qiu et al, 2015). One study even shows ridiculously high levels of lead in Chinese children “Results showed that mean blood lead level was 88.3 micro g/L for 3 – 5 year old children living in the cities in China and mean blood lead level of boys (91.1 micro g/L) was higher than that of girls (87.3 micro g/L). Twenty-nine point nine one per cent of the children’s blood lead level exceeded 100 micro g/L” (Qi et al, 2002), while Li et al (2014) found similar levels. Shanghai also has higher levels of blood lead than the rest of the developed world (Cao et al, 2014). Blood lead levels are also higher in Taizhou, China compared to other parts of the country—and the world (Gao et al, 2017). But blood lead levels are decreasing with time, but still higher than other developed countries (He, Wang, and Zhang, 2009).
Furthermore, Chinese women, compared to American women, had two times higher BLL (Wang et al, 2015). With transgenerational epigenetic inheritance playing a part in the inheritance of methylation DNA passed from mother to daughter then to grandchildren (Sen et al, 2015), this is a public health threat to Chinese women and their children. So just by going off of this data, the claim that China is a safe country should be called into question.
Reality seems to tell a different story. It seems that the true crime rate in China is covered up, especially the murder rate:
In Guangzhou, Dr Bakken’s research team found that 97.5 per cent of crime was not reported in the official statistics.
Of 2.5 million cases of crime, in 2015 the police commissioner reported 59,985 — exactly 15 less than his ‘target’ of 60,000, down from 90,000 at the start of his tenure in 2012.
The murder rate in China is around 10,000 per year according to official statistics, 25 per cent less than the rate in Australia per capita.
“I have the internal numbers from the beginning of the millennium, and in 2002 there were 52,500 murders in China,” he said.Instead of 25 per cent less murder than Australia, Dr Bakken said the real figure was closer to 400 per cent more.”
Guangzhou, for instance, doesn’t keep data for crime committed by migrants, who commit 80 percent of the crime in this province. Out of 2.5 million crimes committed in Guangzhou, only 5,985 crimes were reported in their official statistics, which was 15 crimes away from their target of 6000. Weird… Either way, China doesn’t have a similar murder rate to Switzerland:
The murder rate in China does not equal that of Switzerland, as the Global Times claimed in 2015. It’s higher than anywhere in Europe and similar to that of the US.
China also ranks highly on the corruption index, higher than the US, which is more evidence indicative of a covered up crime rate. So this is good evidence that, contrary to the claims of people who would attempt to downplay the lead-crime relationship, that these effects are real and that they do matter in regard to crime and murder.
So it’s clear that we can’t trust the official Chinese crime stats since there much of their crime is not reported. Why should we trust crime stats from a corrupt government? The evidence is clear that China has a higher crime—and murder rate—than is seen on the Chinese books.
Lastly, effects of epigenetics can and do have a lasting effect on even the grandchildren of mothers exposed to lead while pregnant (Senut et al, 2012; Sen et al, 2015). Sen et al (2015) showed lead exposure during pregnancy affected the DNA methylation status of the fetal germ cells, which then lead to altered DNA methylation on dried blood spots in the grandchildren of the mother exposed to lead while pregnant.—though it’s indirect evidence. If this is true and holds in larger samples, then this could be big for criminological theory and could be a cause for higher rates of black crime (note: I am not claiming that lead exposure could account for all, or even most of the racial crime disparity. It does account for some, as can be seen by the data compiled here).
In conclusion, the relationship between lead exposure and crime is robust and replicated across many countries and cultures. No safe level of blood lead exists, even so-called trace amounts can have horrible developmental and life outcomes, which include higher rates of criminal activity. There is a clear relationship between lead increases/decreases in populations—even within cities—that then predict crime rates. Some may point to the Chinese as evidence against a strong relationship, though there is strong evidence that the Chinese do not report anywhere near all of their crime data. Epigenetic inheritance, too, can play a role here mostly regarding blacks since they’re more likely to be exposed to high levels of lead in the womb, their infancy, and childhood. This could also exacerbate crime rates, too. The evidence is clear that lead exposure leads to increased criminal activity, and that there is a strong relationship between blood lead levels and crime.
Minimalist Races Exist and are Biologically Real
3050 words
People look different depending on where their ancestors derived from; this is not a controversial statement, and any reasonable person would agree with that assertion. Though what most don’t realize, is that even if you assert that biological races do not exist, but allow for patterns of distinct visible physical features between human populations that then correspond with geographic ancestry, then race—as a biological reality—exists because what denotes the physical characters are biological in nature, and the geographic ancestry corresponds to physical differences between continental groups. These populations, then, can be shown to be real in genetic analyses, and that they correspond to traditional racial groups. So we can then say that Eurasian, East Asian, Oceanian, black African, and East Asians are continental-level minimalist races since they hold all of the criteria needed to be called minimalist races: (1) distinct facial characters; (2) distinct morphologic differences; and (3) they come from a unique geographic location. Therefore minimalist races exist and are a biological reality. (Note: There is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016), but this does not mean that the minimalist biological concept of race has no grounding in biology.)
Minimalist race exists
The concept of minimalist race is simple: people share a peculiar geographic ancestry unique to them, they have peculiar physiognomy (facial features like lips, facial structure, eyes, nose etc), other physical traits (hair/hair color), and a peculiar morphology. Minimalist races exist, and are biologically real since minimalist races can survive findings from population genetics. Hardimon (2017) asks, “Is the minimalist concept of race a social concept?” on page 62. He writes that social concepts are socially constructed in a pernicious sense if and only if it “(i) fails to represent any fact of the matter and (ii) supports and legitimizes domination.” Of course, populations who derive from Africa, Europe, and East Asia have peculiar facial morphology/morphology unique to that isolated population. Therefore we can say that minimalist race does not conform to criteria (i). Hardimon (2017: 63) then writes:
Because it lacks the nasty features that make the racialist concept of race well suited to support and legalize domination, the minimalist race concept fails to satisfy condition (ii). The racialist concept, on the other hand, is socially constructed in the pernicious sense. Since there are no racialist races, there are no facts of the matter it represents. So it satisfies (i). To elaborate, the racialist race concept legtizamizes racial domination by representing the social hierarchy of race as “natural” (in a value-conferring sense): as the “natural” (socially unmediated and inevitable) expression of the talent and efforts of the inidividuals who stand on its rungs. It supports racial domination by conveying the idea that no alternative arrangment of social institutions could possibly result in racial equality and hence that attempts to engage in collective action in the hopes of ending the social hierarchy of race are futile. For these reasons the racialist race concept is also idealogical in the prejorative sense.
Knowing what we know about minimalist races (they have distinct physiognomy, distinct morphology and geographic ancestry unique to that population), we can say that this is a biological phenomenon, since what makes minimalist races distinct from one another (skin color, hair color etc) are based on biological factors. We can say that brown skin, kinky hair and full lips, with sub-Saharan African ancestry, is African, while pale/light skin, straight/wavy/curly hair with thin lips, a narrow nose, and European ancestry makes the individual European.
These physical features between the races correspond to differences in geographic ancestry, and since they differ between the races on average, they are biological in nature and therefore it can be said that race is a biological phenomenon. Skin color, nose shape, hair type, morphology etc are all biological. So knowing that there is a biological basis to these physical differences between populations, we can say that minimalist races are biological, therefore we can use the term minimalist biological phenomenon of race, and it exists because there are differences in the patterns of visible physical features between human populations that correspond to geographic ancestry.
Hardimon then talks about how eliminativist philosophers and others don’t deny that above premises above the minimalist biological phenomenon of race, but they allow these to exist. Hardimon (2017: 68-69) then quotes a few prominent people who profess that there are, of course, differences in physical features between human populations:
… Lewontin … who denies that biological races exist, freely grants that “peoples who have occupied major geographic areas for much of the recent past look different from one another. Sub-Saharan Africans have dark skin and people who have lived in East Asia tend to have a light tan skin and an eye color and eye shape that is difference from Europeans.” Similarly, population geneticist Marcus W. Feldman (final author of Rosenberg et al., “Genetic Stucture of Human Populations” [2002]), who also denies the existence of biological races, acknowledges that “it has been known for centuries that certain physical features of humans are concentrated within families: hair, eye, and skin color, height, inability to digest milk, curliness of hair, and so on. These phenotypes also show obvious variation among people from different continents. Indeed, skin color, facial shape, and hair are examples of phenotypes whose variation among populations from different regions is noticeable.” In the same vein, eliminative anthropologist C. Loring Brace concedes, “It is perfectly true that long term residents of various parts of the world have patterns of features that we can identify as characteristic of they area from which they come.”
So even these people who claim to not believe in “biological races”, do indeed believe in biological races because what they are describing is biological in nature and they, of course, do not deny that people look different while their ancestors came from different places so therefore they believe in biological races. We can then use the minimalist biological phenomenon of race to get to the existence of minimalist races.
Hardimon (2017: 69) writes:
Step 1. Recognize that there are differences in patterns of visible physical features of human beings that correspond to their differences in geographic ancestry.
Step 2. Observe that these patterns are exhibited by groups (that is, real existing groups).
Step 3. Note that the groups that exhibit these patterns of visible physical features correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race.
Step 4. Infer that minimalist race exists.
Those individuals mentioned previously who deny biological races but allow that people with ancestors from differing geographic locales look differently do not disagree with step 1, nor does anyone really disagree with step 2. Step 4’s inference immediately flows from the premise in step 3. “Groups that exhibit patterns or visible physical features that correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race. Call (1)-(4) the argument from the minimalist biological phenomenon of race” (Hardimon, 2017: 70). Of course, the argument does not identify which populations may be called races (see further below), it just shows that race is a biological reality. Because if minimalist races exist, then races exist because minimalist races are races. Minimalist races exist, therefore biological races exist. Of course, no one doubts that people come from Europe, sub-Saharan Africa, East Asia, the Americas, and the Pacific Islands, even though the boundaries between them are ‘blurry’. They exhibit patterns of visible physical characters that correspond to their differing geographic ancestry, they are minimalist races therefore minimalist races exist.
Pretty much, the minimalist concept of race is just laying out what everyone knows and arguing for its existence. Minimalist races exist, but are they biologically real?
Minimalist races are biologically real
Of course, some who would assert that minimalist races do not exist would say that there are no ‘genes’ that are exclusive to one certain population—call them ‘race genes’. Of course, these types of genes do not exist. Whether or not one individual is a part of one race or not does not rest on the basis of his physical characters, but is determined by who his parents are, because one of the three premises for the minimalist race argument is ‘must have a peculiar geographic ancestry’. So it’s not that members of races share sets of genes that other races do not, it’s based on the fact that they share a distinctive set of visible physical features that then correspond with geographic ancestry. So of course if the minimalist concept of race is a biological concept then it entails more than ‘genes for’ races.
Of course, there is a biological significance to the existence of minimalist biological races. Consider that one of the physical characters that differ between populations is skin color. Skin color is controlled by genes (about half a dozen within and a dozen between populations). Lack of UV rays for individuals with dark skin will lead to diseases like prostate cancer, while darker skin is a protectant against UV damage to human skin (Brenner and Hearing, 2008; Jablonksi and Chaplin, 2010). Since minimalist race is biologically significant and minimalist races are partly defined by differences in skin color between populations then skin color has both medical and ecological significance.
(1) Consider light skin. People with light skin are more susceptible to skin cancer since they evolved in locations with poor UVR radiation (D’Orazio et al, 2013). The body needs vitamin D to absorb and use calcium for maintaining proper cell functioning. People who evolved near the equator don’t have to worry about this because the doses of UVB they absorb are sufficient for the production of enough previtamin D. While East Asians and Europeans on the other hand, became adapted to low-sunlight locations and therefore over time evolved lighter skin. This loss of pigmentation allowed for better UVB absorption in these new environments. (Also read my article on the evolution of human skin variation and also how skin color is not a ‘tell’ of aggression in humans.)
(2) While darker-skinned people have a lower rate of skin cancer “primarily a result of photo-protection provided by increased epidermal melanin, which filters twice as much ultraviolet (UV) radiation as does that in the epidermis of Caucasians” (Bradford, 2009). Dark skin is thought to have evolved to protect against skin cancer (Greaves, 2014a) but this has been contested (Jablonski and Chaplin, 2014) and defended (Greaves, 2014b). So therefore, using (1) and (2), skin color has evolutionary signifigance.
So as humans began becoming physically adapted to their new niches they found themselves in, they developed new features distinct from the location they previously came from to better cope with the new lifestyle due to their new environments. For instance “Northern Europeans tend to have light skin because they belong to a morphologically marked ancestral group—a minimalist race—that was subject to one set of environmental conditions (low UVR) in Europe” (Hardimon, 2017: 81). Of course explaining how human beings survived in new locations falls into the realm of biology, while minimalist races can explain why this happened.
Minimalist races clearly exist since minimalist races constitute complex biological patterns between populations. Hardimon (2017: 83) writes:
It [minimalist race] also enjoys intrinsic scientific interest because it represents distinctive salient systematic dimension of human biological diversity. To clarify: Minimalist race counts as (i) salient because human differences of color and shape are striking. Racial differences in color and shape are (ii) systematic in that they correspond to differences in geographic ancestry. They are not random. Racial differences are (iii) distinctive in that they are different from the sort of biological differences associated with the other two salient systematic dimensions of human diversity: sex and age.
[…]
An additional consideration: Like sex and age, minimalist race constitutes one member of what might be called “the triumverate of human biodiversity.” An account of human biodiversity that failed to include any one of these three elements would be obviously incomplete. Minimalist race’s claim to be biologically real is as good as the claim of the other members of the triumverate. Sex is biologically real. Age is biologically real. Minimalist race is biologically real.
Real does not mean deep. Compared to the biological associated with sex (sex as contrasted with gender), the biological differences associated with minimalist race are superficial.
Of course, the five ‘clusters’ and ‘populations’ identified by Rosenberg et al’s (2002) K=5 graph, which told structure to produce 5 genetic clusters, corresponds to Eurasia, Africa, East Asia, Oceania, and the Americas, are great candidates for minimalist biological races since they correspond to geographic locations, and even corroborates what Fredrich Blumenbach said about human races back in the 17th century. Hardimon further writes (pg 85-86):
If the five populations corresponding to the major areas are continental-level minimalist races, the clusters represent continental-level minimalist races: The cluster in the mostly orange segment represents the sub-Saharan African continental-level minimalist race. The cluster in the mostly blue segment represents the Eurasian continental-level minimal race. The cluster in the mostly pink segment represents the East Asian continental-level minimalist race. The cluster in the mostly green segment represents the Pacific Islander continental-level minimalist race. And the cluster in the mostly purple segment represents the American continental-level minimalist race.
[…]
The assumption that the five populations are continental-level minimalist races entitles us to interpret structure as having the capacity to assign individuals to continental-level minimalist races on the basis of markers that track ancestry. In constructing clusters corresponding to the five continental-level minimalist races on the basis of objective, race-neutral genetic markers, structure essentially “reconstructs” those races on the basis of a race-blind procedure. Modulo our assumption, the article shows that it is possible to assign individuals to continental-level races without knowing anything about the race or ancestry of the individuals from whose genotypes the microsattelites are drawn. The populations studied were “defined by geography, language, and culture,” not skin color or “race.”
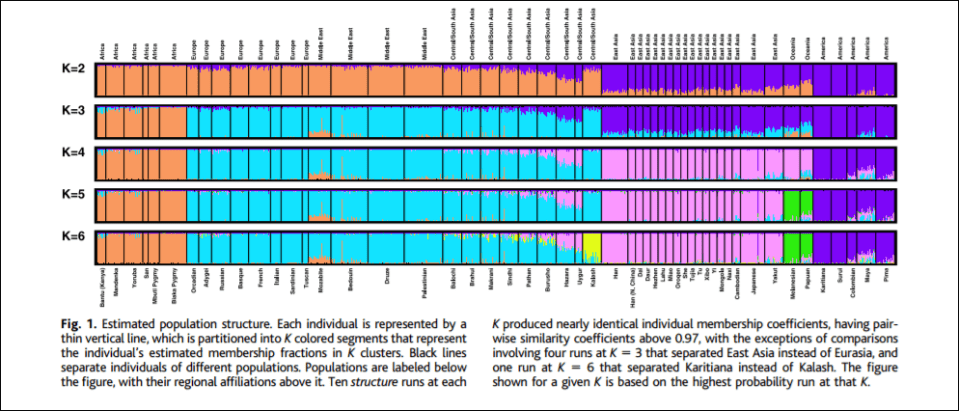
Of course, as critics note, the researchers predetermine how many populations that structure demarcates, for instance, K=5 indicates that the researchers told the program to delineate 5 clusters. Though, these objections do not matter. For the 5 populations that come out in K=5 “are genetically structured … which is to say, meaningfully demarcated solely on the basis of genetic markers” (Hardimon, 2017: 88). K=6 brings one more population, the Kalash, a group from northern Pakistan who speak an Indo-European language. Though “The fact that structure represents a population as genetically distinct does not entail that the population is a race. Nor is the idea that populations corresponding to the five major geographic areas are minimalist races undercut by the fact that structure picks out the Kalash as a genetically distinct group. Like the K=5 graph, the K=6 graph shows that modulo our assumption, continental-level races are genetically structured” (Hardimon, 2017: 88).
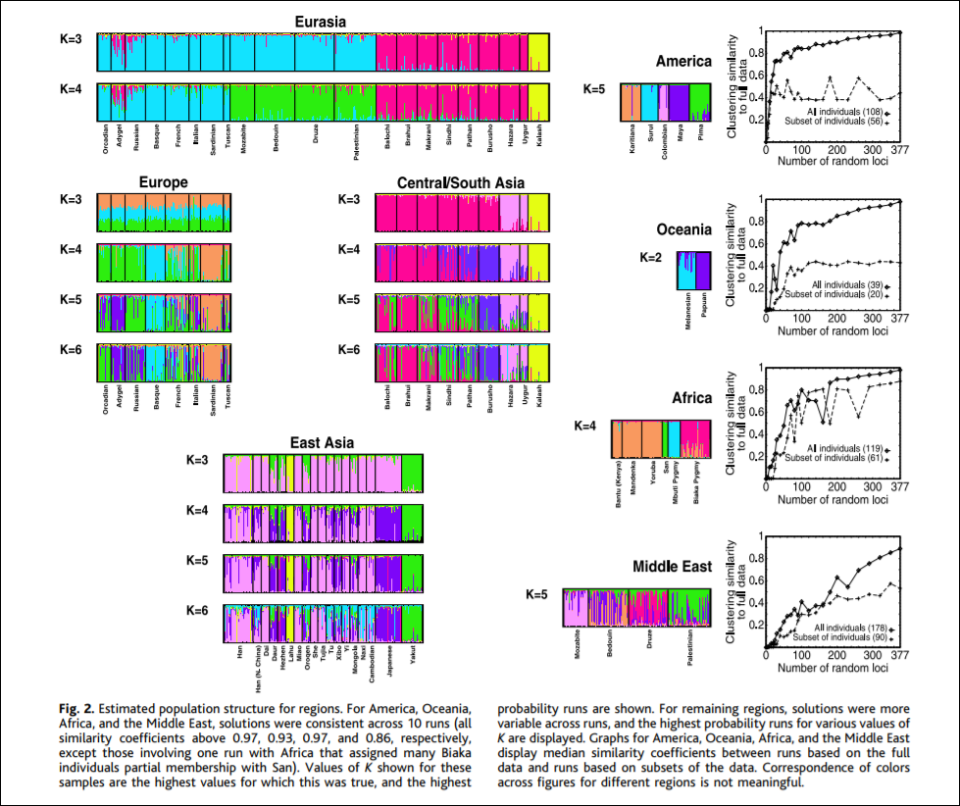
Though of course there are naysayers. Svante Paabo and David Serre, Hardimon writes, state that when individuals are sampled from homogeneous populations from around the world, the gradients of the allele frequencies that are found are distributed randomly across the world rather than clustering discretely. Though Rosenberg et al responded by verifying that the clusters they found are not artifacts of sampling as Paabo and Serre imply, but reflect features of underlying human variation. Though Rosenberg et al agree with Paabo and Serre in that that human genetic diversity consists of clines in variation in allele frequencies (Hardimon, 2017: 89). Other naysayers also state that all Rosenberg et al show is what we can “see with our eyes”. Though a computer does not partition individuals into different populations based on something that can be done with eyes, it’s based on an algorithm.
Hardimon also accepts that black Africans, Caucasians, East Asians, American Indians and Oceanians can be said to be races in the basic sense because “they constitute a partition of the human species“, and that they are distinguishable “at the level of the gene” (Hardimon, 2017: 93). And of course, K=5 shows that the 5 races are genetically distinguishable.
Hardimon finally discusses some medical significance for minimalist races. He states that if you are Caucasian that it is more likely that you have a polymorphism that protects against HIV compared to a member of another race. Meanwhile, East Asians are more likely to carry alleles that make them more susceptible to Steven-Johnson syndrome or another syndrome where their skin falls off. Though of course, the instances where this would matter in a biomedical context are rare, but still should be at the back of everyone’s mind (as I have argued), even though instances where medical differences between minimalist races are rare, there are times where one’s race can be medically significant.
Hardimon finally states that this type of “metaphysics of biological race” can be called “deflationary realism.” Deflationary because it “consists in the repudiation of the ideas that racialist races exist and that race enjoys the kind of biological reality that racialist race was supposed to have” and realism which “consists in its acknowledgement of the existence of minimalist races and the genetically grounded, relatively superficial, but still significant biological reality of minimalist race” (Hardimon, 2017: 95-96).
Conclusion
Minimalist races exist. Minimalist races are a biological reality because distinct visible patterns show differences between geographically isolated populations. This is enough for the classification of the five classic races we know of to be called race, be biologically real, and have a medical significance—however small—because certain biological/physical traits are tied to different geographic populations—minimalist races.
Hardimon (2017: 97) shows an alternative to racialism:
Deflationary realism provides a worked-out alternative to racialism—it it a theory that represents race as a genetically grounded, relatively superficial biological reality that is not normatively important in itself. Deflationary realism makes it possible to rethink race. It offers the promise of freeing ourselves, if only imperfectly, from the racialist background conception of race.
It is clear that minimalist races exist and are biologically real. You do not need to speak about supposed mental traits between these minimalist races, they are irrelevant to the existence of these minimalist biological races. As Hardimon (2017: 67) writes: “No reference is made to normatively important features such as intelligence, sexuality, or morality. No reference is made to essences. The idea of sharp boundaries between patterns of visible physical features or corresponding geographical regions is not invoked. Nor again is reference made to the idea of significant genetic differences. No reference is made to groups that exhibit patterns of visible physical features that correspond to geographic ancestry.”
The minimalist biological concept of race stands up to numerous lines of argumentation, therefore we can say without a shadow of a doubt that minimalist biological race exists and is real.
Do pigmentation and the melanocortin system modulate aggression and sexuality in humans as they do in other animals? A Response to Rushton and Templer (2012)
2100 words
Rushton et al have kept me pretty busy over the last year or so. I’ve debunked many of their claims that rest on biology—such as testosterone causing crime and aggression. The last paper that Rushton published before he died in October of 2012 was an article with Donald Templer—another psychologist—titled Do pigmentation and the melanocortin system modulate aggression and sexuality in humans as they do in other animals? (Rushton and Templer, 2012) and they make a surfeit of bold claims that do not follow. They review animal studies on skin and fur pigmentation and show that the darker an animal’s skin or fur, the more likely they are to be aggressive and violent. They then conclude that, of course (it wouldn’t be a Rushton article without it), that the long-debunked r/K ‘continuum’ explains the co-variation between human populations in birth rate, longevity, violent crime, infant mortality and rate and acquisition of AIDS/HIV.
In one of the very first articles I wrote on this site, I cited Rushton and Templer (2012) favorably (back when I had way less knowledge of biology and hormones). I was caught by biases and not knowing anything about what was discussed. After I learned more about biology and hormones over the years, I came to find out that the claims in the paper are wrong and that they make huge, sweeping conclusions based on a few correlations. Either way, I have seen the error of my ways and the biases that lead me to the beliefs I held, and when I learned more about hormones and biology I saw how ridiculous some of the papers I have cited in the past truly were.
Rushton and Templer (2012) start off the paper by discussing Ducrest et al (2008) who state that within each species studied, darker-pigmented individuals of said species exhibited higher rates of aggression, sexuality and social dominance (which is caused by testosterone) than lighter-pigmented individuals in that same species. They state that this is due to pleiotropy—when a single gene has to or more phenotypic effects. They then refer to Rushton and Jensen (2005) to reference the claim that low IQ is correlated with skin color (skin color doesn’t cause IQ, obviously).
They then state that in 40 vertebrate species that within each that the darker-pigmented members had higher levels of aggression and sexual activity along with a larger body size, better stress resistance, and are more physically active while grooming (Ducrest, Keller, and Roulin, 2008). Rushton and Templer (2012) then state that this relationship was ‘robust’ across numerous species, specifically 36 species of birds, 4 species of fish, 3 species of mammals, and 4 species of reptiles.
Rushton and Templer (2012) then discuss the “Validation of the pigmentation system as causal to the naturalistic observations was demonstrated by experimentally manipulating pharmacological dosages and by studies of cross-fostering“, citing Ducrest, Keller, and Roulin (2008). They even state that ‘Placing darker versus lighter pigmented individuals with adoptive parents of the opposite pigmentation did not modify offspring behavior.” Seems legit. Must mean that their pigmentation caused these differences. They then state something patently ridiculous: “The genes that control that balance occupy a high level in the hierarchical system of the genome.” Though, unfortunately for their hypothesis, there is no privileged level of causation (Noble, 2016; also see Noble, 2008), so this is a nonsense claim. Genes are not ‘blueprints’ or ‘recipes’ (Oyama, 1985; Schneider, 2007).
They then refer to Ducrest, Keller and Roulin (2008: 507) who write:
In this respect, it is important to note that variation in melanin-based coloration between human populations is primarily due to mutations at, for example, MC1R, TYR, MATP and SLC24A5 [29,30] and that human populations are therefore not expected to consistently exhibit the associations between melaninbased coloration and the physiological and behavioural traits reported in our study.
This quote, however, seems to be ignored by Rushton and Templer (2012) throughout the rest of their article, and so even though they did a brief mentioning of the paper and how one should be ‘cautious’ in interpreting the data in their study, it seems like they just brush it under the rug to not have to contend with it. Rushton and Templer (2012) then cite the famous silver fox study, where tame foxes were bred. They lost their dark fur and became lighter and, apparently, were less aggressive than their darker-pigmented kin. These animal studies are, in my useless when attempting to correlate skin color and the melanocortin system in the modulation of aggressive behavior, so let’s see what they write about human studies.
It’s funny, because Rushton and Templer (2012) cite Ducrest, Keller, and Roulin (2008: 507) to show that caution should be made when assessing any so-called differences in the melanocortin system between human races. They then disregard that by writing “A first examination of whether melanin based pigmentation plays a role in human aggression and sexuality (as seen in non-human animals), is to compare people of African descent with those of European descent and observe whether darker skinned individuals average higher levels of aggression and sexuality (with violent crime the main indicator of aggression).” This is a dumb comparison. Yes, African nations commit more crime than European nations, but does this mean that the skin color (or whatever modulates skin color/melanocortin system) is the cause for this? No. Not at all.
There really isn’t anything to discuss here, though, because they just run through how different African nations have higher levels of crime than European and East Asian nations, how blacks report having more sex and feel less guilty about it. Rushton and Templer (2012) then state that one study “asked married couples how often they had sex each week. Pacific Islanders and Native Americans said from 1 to 4 times, US Whites answered 2–4 times, while Africans said 3 to over 10 times.” They then switch over to their ‘replication’ of this finding, using the data from Alfred Kinsey (Rushton and Bogaert, 1988). Though, unfortunately for Rushton and Bogaert, there are massive problems with this data.
Though, the Kinsey data can hardly be seen as representative (Zuckerman and Brody, 1988), and it is also based on outdated, non-representative, non-random samples (Lynn, 1989). Rushton and Templer (2012) also discuss so-called differences in penis size between races, too. But I have written two response articles on the matter and shown that Rushton used shoddy sources like ‘French Army Surgeon who contradicts himself: “Similarly, while the French Army surgeon announces on p. 56 that he once discovered a 12-inch penis, an organ of that size becomes “far from rare” on p. 243. As one might presume from such a work, there is no indication of the statistical procedures used to compute averages, what terms such as “often” mean, how subjects were selected, how measurements were made, what the sample sizes were, etc” (Weizmann et al, 1990: 8).
Rushton and Templer (2012) invoke, of course, Rushton’s (1985; 1995) r/K selection theory as applied to human races. I have written numerous articles on r/K selection and attempts at reviving it, but it is long dead, especially as a way to describe human populations (Anderson, 1991; Graves, 2002). The theory was refuted in the late 70s (Graves, 2002), and replaced with age-specific mortality (Reznick et al, 2002). Some of his larger claims I will cover in the future (like how r/K relates to criminal activity), but he just goes through all of the same old motions he’s been going through for years, bringing nothing new to the table. In all honesty, testosterone is one of the pillars of Rushton’s r/K selection theory (e.g., Lynn, 1990; Rushton, 1997; Rushton, 1999; Hart, 2007; Ellis, 2017; extensive arguments against Ellis, 2017 can be found here). If testosterone doesn’t do what he believes it does and the levels of testosterone between the races are not as high as believed/non-existent (Gapstur et al, 2002; read my discussion of Gapstur et al 2002; Rohrmann et al, 2007; Richard et al, 2014. Though see Mazur, 2016 and read my interpretation of the paper) then we can safely disregard their claims.
Rushton and Templer (2012: 6) write:
Another is that Blacks have the most testosterone (Ellis & Nyborg, 1992), which
helps to explain their higher levels of athletic ability (Entine, 2000).
As I have said many times in the past, Ellis and Nyborg (1992) found a 3 percent difference in testosterone levels between white and black ex-military men. This is irrelavent. He also, then cites John Entine’s (2002) book Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It, but this doesn’t make sense. Because he literally cites Rushton who cites Ellis and Nyborg (1992) and Ross et al (1986) (stating that blacks have 3-19 percent higher levels of testosterone than whites, citing Ross et al’s 1986 uncorrected numbers)—and I have specifically pointed out numerous flaws in their analysis and so, Ross et al (1986) cannot seriously be used as evidence for high testosterone differences between the races. Though I cited Fish (2013), who wrote about Ellis and Nyborg (1992):
“These uncorrected figures are, of course, not consistent with their racial r- and K-continuum.”
Rushton and Templer (2012) then state that testosterone acts like a ‘master switch’ (Rushton, 1999), implicating testosterone as a cause for aggression, though I’ve shown that this is not true, and that aggression causes testosterone production, testosterone doesn’t cause aggression. Testosterone does control muscle mass, of course. But Rushton’s claim that blacks have deeper voices due to higher levels of testosterone, but this claim does not hold in newer studies.
Rushton and Templer (2012) then shift gears to discuss Templer and Arikawa’s (2006) study on the correlation between skin color and ‘IQ’. However, there is something important to note here from Razib:
we know the genetic architecture of pigmentation. that is, we know all the genes (~10, usually less than 6 in pairwise between population comparisons). skin color varies via a small number of large effect trait loci. in contrast, I.Q. varies by a huge number of small effect loci. so logically the correlation is obviously just a correlation. to give you an example, SLC45A2 explains 25-40% of the variance between africans and europeans.
long story short: it’s stupid to keep repeating the correlation between skin color and I.Q. as if it’s a novel genetic story. it’s not. i hope don’t have to keep repeating this for too many years.
Rushton and Templer (2012: 7) conclude:
The melanocortin system is a physiological coordinator of pigmentation and life history traits. Skin color provides an important marker placing hormonal mediators such as testosterone in broader perspective.
I don’t have a problem with the claim that the melanocortin system is a physiological coordinator of pigmentation, because it’s true and we have a great understanding of the physiology behind the melanocortin system (see Cone, 2006 for a review). EvolutionistX also has a great article, reviewing some studies (mouse studies and some others) showing that increasing melatonin appears to decreases melanin.
Rushton and Templer’s (2012) make huge assumptions not warranted by any data. For instance, Rushton states in his VDare article on the subject, J. Phillipe Rushton Says Color May Be More Than Skin Deep, “But what about humans? Despite all the evidence on color, aggression, and sexuality in animals, there has been little or no discussion of the relationship in people. Ducrest & Co. even warned that genetic mutations may make human populations not exhibit coloration effects as consistently as other species. But they provided no evidence.” All Rushton and Templer (2012) do in their article is just restating known relationships with crime and race, and then attempting to implicate the melanocortin system as a factor driving this relationship, literally off of a slew of animal studies. Even then, the claim that Ducrest, Keller, and Roulin (2008: 507) provide no evidence for their warning is incorrect, because before they stated that, they wrote “In this respect, it is important to note that variation in melanin-based coloration between human populations is primarily due to mutations at, for example, MC1R, TYR, MATP and SLC24A5 [29,30]. . .” Melanin does not cause aggression, it does not cause crime. Rushton and Templer just assume too many things based on no evidence in humans, while their whole hypothesis is structured around a bunch of animal studies.
In conclusion, it seems like Rushton and Templer don’t know anything about the physiology of the melanocortin system if they believe that pigmentation and the melanocortin system modulates aggression and sexual behavior in humans. I know of no evidence (studies, not Rushton and Templer’s 2012 relationships with crime and then asserting that, because these relationships are seen in animals, that it must mean that the melanocortin system in humans modulates the relationships too) for these assertions by Rushton and Templer (2012). The fact that they think that restating relationships between crime and race, country of origin and race, supposed correlations with testosterone and crime and blacks supposedly having higher testosterone than whites, among other things, shows that the proposed relationships are caused by the melanocortin system and Lift History Theory shows their ignorance of the human physiological system.
Height, Longetivity, and Aging
1700 words
Humans reach their maximum height at around their mid-20s. It is commonly thought that taller people have better life outcomes, and are in general healthier. Though this misconception stems from misconceptions about the human body. In all reality, shorter people live longer than taller people. (Manlets of the world should be rejoicing; in case anyone is wondering I am 5’10”.) This flies in the face about what people think, and may be counter-intuitive to some but the logic—and data—is sound. I will touch on mortality differences between tall and short people and at the end talk a bit about shrinking with age (and studies that show there is no—or little—decrease in height due to self-reports, the study is flawed).
One reason why the misconception of taller people living longer, healthier lives than shorter people is the correlation between height and IQ—people assume that they are traits that are ‘similar’ in that they become ‘stable’ at adulthood—but one way to explain that relationship is that IQ is correlated with height because higher SES people can afford better food and thus be better nourished. Either way, it is a myth that taller people have lower rates of all-cause mortality.
The truth of the matter is this: smaller bodies live longer lives, and this is seen in the animal kingdom and humans—larger body size independently reduces mortality (Samaras and Elrick, 2002). They discuss numerous lines of evidence—from human to animal studies—and show that smaller bodies have a lower chance of all-cause mortality, the reasoning being (one of the reasons, anyway) that larger bodies have more cells which then would, in turn, be more subject to carcinogens and, obviously, would have higher rates of cancer which would then, too, lower mortality rates. Samaras (2012) also has another paper where the implications are reviewed for this, and other causes are proposed for this observation. Causes are reduced cell damage, lower DNA damage, and lower cancer incidence; with other, hormonal differences, between tall and short people that explain more of the variation between them.
One study found a positive linear correlation between height and cancer mortality. Lee et al (2009) write:
A positive linear association was observed between height and cancer mortality. For each standard deviation greater height, the risk of cancer was increased by 5% (2–8%) and 9% (5–14%) in men and women, respectively.
One study suggests that “variations in adult height (and, by implication, the genetic and other determinants of height) have pleiotropic effects on several major adult-onset diseases” (The Emerging Risk Factors Collaboration, 2012). Taller people also are at greater risk for heart attack (Tamaras, 2013). The cause for this, Tamaras writes, is “including reduced telomere shortening, lower atrial fibrillation, higher heart pumping efficiency, lower DNA damage, lower risk of blood clots, lower left ventricular hypertrophy and superior blood parameters.” Height, though, may be inversely associated with long-term incidence of fatal stroke (Goldbourt and Tanne, 2002). Schmidt et al (2014) conclude: “In conclusion, short stature was a risk factor for ischemic heart disease and premature death, but a protective factor for atrial fibrillation. Stature was not substantially associated with stroke or venous thromboembolism.” Cancer incidence also increases with height (Green et al, 2011). Samaras, Elrick, and Storms (2003) suggest that men live longer than women live longer than men due to the height difference between them, being about 8 percent taller than women but having a 7.9 percent lower life expectancy at birth.
Height at mid-life, too, is a predictor of mortality with shorter people living longer lives (He et al, 2014). There are numerous lines of evidence that shorter people—and people of shorter ethnies, too—live longer lives if they are vertically challenged. One study on patients undergoing maintenance hemodialysis stated that “height was directly associated with all-cause mortality and with mortality due to cardiovascular events, cancer, and infection” (Daugirdas, 2015; Shapiro et al, 2015). Even childhood height is associated with prostate cancer acquisition (Aarestrup et al, 2015). Even men who are both tall and have more adipose tissue (body fat) are more likely to die younger and that greater height was associated with a higher risk of acquiring prostate cancer (Perez-Cornago et al, 2017). Short height is a risk factor for death for hemodyalisis patients (Takenaka et al, 2010). Though there are conflicting papers regarding short height and CHD, many reviews show that shorter people have better health outcomes than taller people.
Sohn (2016) writes:
An additional inch increase in height is related to a hazard ratio of death from all causes that is 2.2% higher for men and 2.5% higher for women. The findings are robust to changing survival distributions, and further analyses indicate that the figures are lower bounds. This relationship is mainly driven by the positive relationship between height and development of cancer. An additional inch increase in height is related to a hazard ratio of death from malignant neoplasms that is 7.1% higher for men and 5.7% higher for women.
[…]
It has been widely observed that tall individuals live longer or die later than short ones even when age and other socioeconomic conditions are controlled for. Some researchers challenged this position, but their evidence was largely based on selective samples.
Four additional inches of height in post-menopausal women coincided with an increase in all types of cancer risk by 13 percent (Kabat et al, 2013), while taller people also have less efficient lungs (Leon et al, 1995; Smith et al, 2000). Samaras and Storms (1992) write “Men of height 175.3 cm or less lived an average of 4.95 years longer than those of height over 175.3 cm, while men of height 170.2 cm or less lived 7.46 years longer than those of at least 182.9 cm.”
Lastly, regarding height and mortality, Turchin et al (2012) write “We show that frequencies of alleles associated with increased height, both at known loci and genome wide, are systematically elevated in Northern Europeans compared with Southern Europeans.” This makes sense, because Southern European populations live longer (and have fewer maladies) than Northern European populations:
Compared with northern Europeans, shorter southern Europeans had substantially lower death rates from CHD and all causes.2 Greeks and Italians in Australia live about 4 years longer than the taller host population … (Samaras and Elrick, 2002)
So we have some data that doesn’t follow the trend of taller people living shorter lives due to maladies they acquire due to their height, but most of the data points in the direction that taller people live shorter lives, higher rates of cancer, lower heart pumping efficiency (the heart needs to pump more blood through a bigger body) etc. It makes logical sense that a shorter body would have fewer maladies, and would have higher heart pumping efficiency, lower atrial fibrillation, lower DNA damage, lower risk of blood clotting (duh) when compared to taller people. So it seems that, if you’re a normal American man, then if you want to live a good, long life then you’d want to be shorther, rather than taller.
Lastly, do we truly shrink as we age? Steve Hsu has an article on this matter, citing Birrell et al (2005) which is a longitudinal study in Newcastle, England which began in 1947. The children were measured when full height was expected to be acheived, which is about 22 years of age. They were then followed up at age 50. Birrell et al (2005) write:
Height loss was reported by 57 study members (15%, median height loss: 2.5 cm), with nine reporting height loss of >3.5 cm. However, of the 24 subjects reporting height loss for whom true height loss from age 22 could be calculated, assuming equivalence of heights within 0.5 cm, 7 had gained height, 9 were unchanged and only 8 had lost height. There was a poor correlation between self-reported and true height loss (r=0.28) (Fig. 1).
In this population, self-reported height was off the mark, and it seems like Hsu takes this conclusion further than he should, writing “Apparently people don’t shrink quite as much with age as they think they do.” No no no. This study is not good. We begin shrinking at around age 30:
Men gradually lose an inch between the ages of 30 to 70, and women can lose about two inches. After the age of 80, it’s possible for both men and women to lose another inch.
The conclusion from Hsu on that study is not warranted. To see this, we can look at Sorkin, Muller, and Andres (1999) who write:
For both sexes, height loss began at about age 30 years and accelerated with increasing age. Cumulative height loss from age 30 to 70 years averaged about 3 cm for men and 5 cm for women; by age 80 years, it increased to 5 cm for men and 8 cm for women. This degree of height loss would account for an “artifactual” increase in body mass index of approximately 0.7 kg/m2 for men and 1.6 kg/m2 for women by age 70 years that increases to 1.4 and 2.6 kg/m2, respectively, by age 80 years.
So, it seems that Hsu’s conclusion is wrong. We do shrink with age for myriad reasons, including discs between the vertebrae and spine decompress and dehydrate, the aging spine becomes more curved due to loss of bone density, and loss of torso muscle could contribute to the differing posture. Either way, these are preventable, but some height decrease will be notable for most people. Either way, Hsu doesn’t know what he’s talking about here.
In conclusion, while there is some conflicting data on whether tall or short people have lower all-cause mortality, the data seems to point to the fact that shorter people live longer due since they have lower atrial fibrillation, higher heart pumping efficiency, low DNA damage, lower risk for blood clots (since the blood doesn’t have to travel too far in shorter people), along with superior blood parameters etc. With the exception of a few diseases, shorter people do have a higher quality of life and higher lung efficiency. We do get shorter as we age—though with the right diet we can ameliroate some of those effects (for instance keeping calcium high). There are many reasons why we shrink due to age, and the study that Hsu cited isn’t good compared to the other data we have in the literature on this phenomenon. All in all, shorter people live longer for myriad reasons and we do shrink as we age, contrary to Steve Hsu’s claims.
Happy Darwin Day: The Modern Synthesis Has Causation in Biology Wrong
2800 words
The first Darwin Day I started writing just for this day, I wrote about (and defended Darwin’s words) how both Creationists and evolutionists who are themselves evolutionary progressionists twist Darwin’s words for their own gain. Darwin never wrote in The Descent of Man that the ‘higher races’ would take out ‘the lower races’, but that doesn’t stop Creationists and evolutionists—who I presume have not read one sentence in Darwin’s words from one of his books—from taking what Darwin meant out of context and attributing to him beliefs he does not hold. This year, though, I am going in a different direction. The Modern Synthesis (MS) has causation in biology wrong. The MS upholds the ‘gene’ as one of the highest seats in evolutionary biology, with a sort of ‘power’ to direct. Though, as I will show, genes do nothing unless transcribed by the system. Since the MS has causation in biology wrong, then we either need to extend or replace the MS.
To begin, Darwin, without knowledge of genes or other hypothesized units of inheritance, had a theory of inheritance in which things called ‘gemmules’ (what Darwin called heritable molecules) were transmitted to offspring (Choi and Mango, 2014). It’s ironic, because Darwin’s theory of inheritance was one of the more Lamarckian theories of inheritance in his day, and Darwin himself sympathized with the Lamarckian view of evolution—he most definitely did not discard it like modern-day Darwinists do. Darwin suggested that these gemmules circulated in the body and that some were used for the regeneration of some bodily tissues, but most aggregated in the reproductive organs (Jablonka and Lamb, 2015: 23). Further, according to Darwin, gemmules were not always immediately used but could reappear later in life or even be used in future generations. Darwin even said that “inheritance must be looked at as a form of growth” (Darwin, 1883, vol 2, p. 398; quoted by Jablonka and Lamb, 2015: 24).
The crux of the MS is the selfish gene theory of Dawkins (1976). Dawkins (1976, 2006) writing “They are in you and me; they created us, body and mind; and their preservation is the ultimate rationale for our existence.” “They”, of course, being genes. The gene has been given a sort of power that it does not have, but has been placed on it by overzealous people, quick to jump to conclusions while we still have yet to understand what ‘genes’ do. The MS—with the selfish gene theory—is at the forefront of the neo-Darwinist revolution, that evolution is gene-centered, with genes playing the starring role in the evolutionary story.
Though, numerous researchers are against such simplistic and reductionist viewpoints of evolution, mainly the gene-centered view of evolution pushed by the MS. There is no privileged level of causation in biology (though I will state later in this article that I think ATP comes close to it) (Noble, 2016).
Neo-Darwinists, like Richard Dawkins, overstate natural selection’s importance regarding evolution. They elevate the gene’s overall importance. In the quote from Dawkins above, where he stated that “they” (genes) “created us, body and mind”, he is implying that genes are a sort of ‘blueprint’, like a ‘plan’ or ‘recipe’ for the form of the organism. But this was taken care of by Susan Oyama in her 1985 book The Ontogeny of Information where she writes on pages 77:
“Though a plan implies action, it does not itself act, so if the genes are a blueprint, something else is the constructor-construction worker. Though blueprints are usually contrasted with building materials, the genes are quite easily conceptualized as templates for building tools and materials; once so utilized, of course, they enter the developmental process and influence its course. The point of the blueprint analogy, though, does not seem to be to illuminate developmental processes, but rather to assume them and, in celebrating their regularity, to impute cognitive functions to genes. How these functions are exercised is left unclear in this type of metaphor, except that the genetic plan is seen in some peculiar way to carry itself out, generating all the necessary steps in the necessary sequence. No light is shed on multiple developmental possibilities, species-typical or atypical.“
The genes-as-blueprints canard is one that is heavily used by proponents of the MS. Oyama also writes on page 53 “Just as traditional thought placed biological forms in the mind of God, so modern thought finds many ways of endowing the genes with ultimate formative power, a power bestowed by Nature over countless millennia.” This same sentiment from Oyama is also echoed by developmental systems theorist and psychologist David Moore in his book The Dependent Gene: The Fallacy of “Nature VS. nurture”, where he writes:
Such contextual dependence renders untenable the simplistic belief that there are coherent, long-lived entities called “genes” that dictate instructions to cellular machinery that merely constructs the body accordingly. The common belief that genes contain context-independent “information”—and so are analogous to “blueprints” or “recipes”—is simply false. (p. 81) (Quoted from Schneider, 2007)
Environmental factors are imperative in determining which protein-coding exons get read from a cistron, when and how often. So the very concept of a gene depends on the environment and environmental inputs, and thusly gene ABC does not code for trait T on its own.
When it comes to epigenetics (defined here as inherited changes in gene expression with no genetic change to the genome), this completely changes how we view evolution.
The underlying nucleotide sequence stays the same but differences are inherited due to environmental stressors. I’ve stated in the past that these inherited marks on the genome (through histone modification, DNA methylation, which then alter the chromatin structure of the DNA. Further, this would show up on heritability estimates as ‘genetic’ when the ’cause’ was ‘environmental’ in nature (which is also yet another reason that heritability estimates are inflated).
DNA methylation, histone modification and noncoding RNA all can affect the structure of chromatin. As of now, the mechanisms of mitotic inheritance aren’t too well known, but advances in the field are coming.
If you want to talk the P and F1 generations regarding transgenerational epigenetics, then you must realize that these changes do not occur on the genome, the genome remains the same, just certain genes are expressed differently (as I’m sure you know). Though mi-MRNA signals can change the DNA methylation patterns in the F2 sperm which then is replicated in meiotic and mitotic cycles (Trerotola et al, 2015).
For another similar process on how DNA methylation persists, this (semiconservative) replication of DNA methylation occurs on both strands of the DNA which then become hemimethylated DNA which can then become fully methylated by methylase maintenance. So chromatin structure affects the genetic expression of the eukaryotic genome which then becomes the basis for epigenetic effects. Xist RNA also mediates the X-chromosome deactivation. This doesn’t even get into how and why the microbiome can also affect gene expression (which has also been called ‘the second genome’ (Zhu, Wang, and Li, 2010) with other authors calling it an ‘organ’ (Clarke et al, 2014; Brown and Hazen, 2015) this can also affect gene expression and heritable variation that becomes the target of selection (along with the other modes of selection) (Maurice, Haiser, and Turnbaug, 2014; Byrd and Segre, 2015). This shows that gene expression in the F2 and F3 generations is not so simple, and that other factors such as our gut microbiota can also affect gene expression and stressors experienced by parents and grandparents can also be passed to future generations, and may have a chance of becoming part of heritable variation that natural selection then acts on (Jablonka and Lamb, 2015).
The point of the debate with neo-Darwinists is over causation: do genes hold this ‘ultimate formative power’ as people like Dawkins contest? Or are genes nothing but ‘slaves’, passive, not active, causes as Denis Noble writes in his 2016 book Dance to the Tune of Life. (Noble, 2008 discusses genes and causation, again showing that there is no true causation, but getting technical, ATP is up there in the ‘chain’, if you want to get literal. The point is that genes do not have the ‘power’ that the neo-Darwinists think they do, they’re just slaves for the intelligent physiological system.)
When discovering the structure of DNA, Francis Crick famously announced to his drinking companions in a Cambridge tavern that he had discovered ‘the secret of life’. The director of his Institute, Max Perutz, was rather more careful than Crick when he said that DNA was the ‘score of life’. That is more correct since a musical score does nothing until it is played, DNA does nothing until activated to do so.
[…]
Recent experimental work in biological science has deconstructed the idea of a gene, and an important message of this book is that it has thereby drthroned the gene as a uniquely privileged level of causation. As we will see, genes, defined as DNA sequences, are indeed essential, but not in the way in which they are often portrayed. (Noble, 2016: 53)
A 2017 paper titled Was the Watchmaker Blind? Or Was She One-Eyed?, Noble and Noble (2017) write that organisms and their interacting populations have evolved mechanisms so that they can harness blind stochasticity, thereby generating functional changes to the phenotype as to better respond to environmental challenges. They put forth a good argument, though it really makes me think because I’ve been such a staunch critic against evolution having a ‘direction’ and against the ‘teleological view’ of evolution: “If organisms have agency and, within obvious limits, can choose their lifestyles, and if these lifestyles result in inheritable epigenetic changes, then it follows that organisms can at least partially make choices that can have long-term evolutionary impact.”
Noble and Noble (2017) argue (using Dawkins’ analogy of the Blind Watchmaker) that humans are the only Watchmakers that we know of. Humans evolved from other organisms. The ability to become a Watchmaker has evolved. Ergo, there is no surprise that there is directed agency for other organisms that directs their evolution too. There are several processes, they conclude, that could account for directed evolutionary change which are “targeted mutation, gene transposition, epigenetics, cultural change, niche construction and adaptation” (Noble and Noble, 2017). Niche construction, for instance, is heavily pushed by Kevin Laland, author of the book Darwin’s Unfinished Symphony: How Culture Made the Human Mind who has a few papers and featured it heavily in his new book. Either way, these ways in which organisms can in a way direct their own evolution are not covered by the MS.
Though I couldn’t end this article without, of course, discussing Jerry Coyne who goes absolutely crazy at people pushing to either extend or replace the MS. His most recent article is about Kevin Laland and how he is “at it again” touting “a radically different view of evolution”. It seems as Coyne has made up his mind and that the MS is all there is—he believes it is no problem for our current understanding of evolutionary theory to absorb things such as niche construction, epigenetic inheritance, stochasticity, and even (way more controversially) directed mutations. Coyne has also criticized Noble’s attacks on the MS, though Noble came back and responded to Coyne during a video presentation.
Lastly, Portin and Wilkins (2017) review the history of the gene, and go through different definitions it has been given over the decades. They conclude in this paper that they “will propose a definition that we believe comes closer to doing justice to the idea of the “gene,”
in light of current knowledge. It makes no reference to “the unit of heredity”—the long-standing sense of the term—because we feel that it is now clear that no such generic universal unit exists.” Writing on page 1361-1362:
A gene is a DNA sequence (whose component segments do not necessarily need to be physically contiguous) that specifies one or more sequence-related RNAs/proteins that are both evoked by GRNs and participate as elements in GRNs, often with indirect effects, or as outputs of GRNs, the latter yielding more direct phenotypic effects. [GRNs are genetic regulatory networks]
This is similar to what Jablonka and Lamb (2015: 17) write:
Although many psychiatrists, biochemists, and other scientists who are not geneticists (yet express themselves with remarkable facility on genetic issues) still use the language of genes as simple causal agents, and promise their audience rapid solutions to all sorts of problems, they are no more than propagandists whose knowledge or motives must be suspect. The geneticists themselves now think and talk (most of the time) in terms of genetic networks composed of tens or hundreds of genes and gene products, which interact with each other and together affect the development of a particular trait. They recognize that whether or not a trait (a sexual preference, for example) develops does not depend, in the majority of cases, on a difference in a single gene. It involves interactions among many genes, many proteins and other types of molecule, and the environment in which an individual develops.
The gene as an active causal actor has been definitively refuted. Genes on their own do nothing at all, until they are transcribed by the intelligent physiological system. Noble likens genes as slaves that are used by the system to carry out processes by and for the system. So genes are caused to give their information by and to the system that activates them (Noble, 2011). Noble’s slave metaphor makes much more sense than Dawkins’ selfish metaphor, since genes are used like slaves by the system, the genes are then caused to give their information by and to the system that activates them, which shows how they are a passive, not active, cause, completely upending the MS and how it views causation in biology. Indeed, Jablonka and Lamb state that one of their problems with Dawkins is that “Dawkins assumes that the gene is the only biological (noncultural) hereditary unit. This simply is not true. There are additional biological inheritance systems, which he does not consider, and these have properties different from those we see in the genetic system. In these systems his distinction between replicator and vehicle is not valid.”
So, both Gould and Dawkins overlooked the inheritance of acquired characters, as Jablonka and Lamb write in their book. They argue that inherited variation had a large effect on the evolution of species, but admit that evidence for the view is scant. They write on page 145 “If you accept that heritable epigenetic variation is possible, self-evidently some of the variants will have an advantage relative to other variants. Even if all epigenetic variations were blind, this would happen, and it’s very much more likely if we accept that a lot of them are induced and directed.” Not everything that is inherited is genetic.
DNA is found in the cell, and what powers the cell? ATP (adenosine triphosphate). Cells use and store ATP to carry out their functions (Khakh and Burnstock, 2016). Cells produce ATP from ADP and Pi. Cells use exergonic reactions to provide the energy needed to synthesize ATP from ADP and Pi. The hydrolysis of ATP provides the energy needed to drive endergonic actions.So the cells continuously produced more ATP from ADP and Pi to then carry out diverse functionings across the body. So, in a way, you can argue that one of the ultimate causes is ATP since it has to power the cell, then you can look at all of the other reactions that occur before ATP is created and privilege that part of the chain, but there will never be some ultimate causation, since, as Noble argues in his book Dance to the Tune of Life, there is no privileged causation in biology.
In conclusion, evolution, development, and life, in general, is extremely complex. Paradigms like the selfish gene—a largely reductionist paradigm—do not account for numerous other factors that drive the evolution of species, such as targeted mutation, niche construction etc. An extended evolutionary synthesis that integrates these phenomena will better be able to describe what occurs to drive the evolution of species, and if the directed mutation idea has any weight, then it will be interesting to see how and why certain organisms have evolved this ability. It’s ironic how the MS is being defended as if it is infallible—like it can do no wrong and that it does not need to be added to/extended or replaced by something else that incorporates the phenomena brought up in this article.
Either way, a revolution in modern biology is coming, and Darwin would have it no other way. The Modern Synthesis has causation in biology wrong: the gene is not an active agent in evolution, it only does what it is told by the intelligent physiological system, and so we must look at whole organisms and not reduce organisms down to genes, but we must look at the whole organism—a holistic view of the organism, not one that is reduced down to just ‘the genes’, because there is no privileged level of causation in biology (Noble, 2016).
Is There Really More Variation Within Races Than Between Them?
1500 words
In 1972 Richard Lewontin, studying the blood groups of different races, came to the conclusion that “Human racial classification is of no social value and is positively destructive of social and human relations. Since such racial classification is now seen to be of virtually no genetic or taxonomic significance either, no justification can be offered for its continuance” (pg 397). He also found that “the difference between populations within a race account for an additional 8.3 percent, so that only 6.3 percent is accounted for by racial classification.” This has lead numerous people to, along with Lewontin, conclude that race is ‘of virtually no genetic or taxonomic significance’ and conclude that, due to this, race does not exist.
Lewontin’s main reasoning was that since there is more variation within races than between them (85 percent of differences were within populations while 15 percent was within them) then since a lion’s share of human diversity is distributed within races, not between them, then race is of no genetic nor taxonomic use. Lewontin is correct that there is more variation within races than between them, but he is incorrect that this means that racial classification ‘is of no social value’, since knowing and understanding the reality of race (even our perceptions of them, whether they are true or not) influence things such as medical outcomes.
Though, like Lewontin, people have cited this paper as evidence against the existence of human races, for if there is way more genetic variation between races, and that most human genetic variation is within races, then race cannot be of any significance for things such as medical outcomes since most genetic variation is within races not between them.
Rosenberg et al (2002) also confirmed and replicated Lewontin’s analysis, showing that within-population genetic variation accounts for 93-95 percent of human genetic variation, while 3 to 5 percent of human genetic variation lies between groups. Philosopher Michael Hardimon (2017) uses these arguments to buttress his point that ‘racialist races’ (as he calls them) do not exist. His criteria being:
(a) The fraction of human genetic diversity between populations must exceed the fraction of diversity between them.
(b) The fraction of human genetic diversity within populations must be small.
(c) The fraction of diversity between populations must be large.
(d) Most genes must be highly differentiated by race.
(e) The variation in genes that underlie obvious physical differences must be typical of the genome in general.
(f) There must be several important genetic differences between races apart from the genetic differences that underlie obvious physical differences.
Note: (b) says that racialist races are genetically racially homogeneous groups; (c)-(f) say that racialist races are distinguised by major biological differences.
Call (a)-(f) the racialist concept of race’s genetic profile. (Hardimon, 2017: 21)
He clearly strawmans the racialist position, but I’ll get into that another day. Hardimon writes about how both of these studies lend credence to his above argument on racialist races (pg 24):
Rosenberg and colleagues also confirm Lewontin’s findings that most genes are not highly differentiated by race and that the variation in genes that underlie obvious physical differences is not typical of the variation of the genome in general. They also suggest that it is not the case that there are many important genetic differences between races apart from the genetic differences that underlie the obvious physical differences. These considerations further buttress the case against the existence of racialist races.
[…]
The results of Lewontin’s 1972 study and Rosenberg and colleagues’ 2002 study strongly suggest that it is extremely unlikely that there are many important genetic differences between races apart from the genetic differences that underlie the obvious physical differences.
(Hardimon also writes on page 124 that Rosenberg et al’s 2002 study could also be used as evidence for his populationist concept of race, which I will return to in the future.)
Though, my reasoning for writing this article is to show that the findings by Lewontin and Rosenberg et al regarding more variation within races than between them are indeed true, despite claims to the contrary. There is one article, though, that people cite as evidence against the conclusions by Lewontin and Rosenberg et al, though it’s clear that they only read the abstract and not the full paper.
Witherspoon et al (2007) write that “sufficient genetic data can permit accurate classification of individuals into populations“, which is what the individuals who cite this study as evidence for their contention mean, though they conclude (emphasis mine):
The fact that, given enough genetic data, individuals can be correctly assigned to their populations of origin is compatible with the observation that most human genetic variation is found within populations, not between them. It is also compatible with our finding that, even when the most distinct populations are considered and hundreds of loci are used, individuals are frequently more similar to members of other populations than to members of their own population. Thus, caution should be used when using geographic or genetic ancestry to make inferences about individual phenotypes.
Witherspoon et al (2007) analyzed the three classical races (Europeans, Africans and East Asians) over thousands of loci and came to the conclusion when genetic similarity is measured over thousands of loci, the answer to the question “How often is a pair of individuals from one population genetically more dissimilar than two individuals chosen from two different populations?” is “never“.
Hunley, Cabana, and Long (2016: 7) also confirm Lewontin’s analysis, writing “In sum, we concur with Lewontin’s conclusion that Western-based racial classifications have no taxonomic significance, and we hope that this research, which takes into account our current understanding of the structure of human diversity, places his seminal finding on firmer evolutionary footing.” But the claim that “racial classifications have no taxonomic significance” is FALSE.
This is a point that Edwards (2003) rebutted in depth. While he did agree with Lewontin’s (1972) analysis that there was more variation within races than between them (which was confirmed through subsequent analysis), he strongly disagreed with Lewontin’s conclusion that race is of no taxonomic significance. Richard Dawkins, too disagreed with Lewontin, though as Dawkins writes in his book The Ancestors Tale: “Most of the variation among humans can be found within races as well as between them. Only a small admixture of extra variation distinguishes races from each other. That is all correct. What is not correct is the inferene that race is therefore a meaningless concept.” The fact that there is more variation within races than between them is irrelevant to taxonomic classification, and classifying races by phenotypic differences (morphology, and facial features) along with geographic ancestry shows that just by looking at the average phenotype that race exists, though these concepts make no value-based judgements on anything you can’t ‘see’, such as mental and personality differences between populations.
Though while some agree with Edwards’ analysis of Lewontin’s argument about race’s taxonomic significance, they don’t believe that he successfully refuted Lewontin. For instance, Hardimon (2017: 22-23) writes that Lewontin’s argument against—what Hardimon (2017) calls ‘racialist race’ (his strawman quoted above)—the existence of race because the within-race component of genetic variation is greater than the genetic variation between races “is untouched by Edwards’ objections.”
Though Sesardic (2010: 152) argues that “Therefore, contra Lewontin, the racial classification that is based on a number of genetic differences between populations may well be extremely reliable and robust, despite the fact that any single of those genetic between-population differences\ remains, in itself, a very poor predictor of racial membership.” He also states that the 7 to 10 percent difference between populations “actually refers to the inter-racial portion of variation that is averaged over the separate contributions of a number of individual genetic indicators that were sampled in different studies” (pg 150).
I personally avoid all of this talk about genes/allele frequencies between populations and jump straight to using Hardimon’s minimalist race concept—a concept that, according to Hardimon is “stripped down to its barest bones” since it captures enough of the racialist concept of race to be considered a race concept.
In sum, variation within races is greater than variation between races, but this does not mean anything for the reality of race since race can still be delineated based on peculiar physical features and peculiar geographic ancestry to that group. Using a few indicators (morphology, facial features such as nose, lips, cheekbones, facial structure, and hair along with geographic ancestry), we can group races based on these criteria and we can show that race does indeed exist in a physical—not social—sense and that these categories are meaningful in a medical context (Hardimon, 2013, 2017). So even though genetic variation is greater within races than between them, this does not mean that there is no taxonomic significance to race, as other authors have argued. Hardimon (2017: 23) agrees, writing (emphasis his) “… Lewontin’s data do not preclude the possibility that racial classification might have taxonomic significance, but they do preclude the possibility that racialist races exist.”
Hardimon’s strawman of the racialist concept notwithstanding (which I will cover in the future), his other three race concepts (minimalist, populationist and socialrace concepts) are logically sound and stand up to a lot of criticism. Either way, race does exist, and it does not matter if the apportionment of human genetic diversity is greatest within races than between them.
Delaying Gratification and Social Trust
1900 words
Tests of delayed gratification, such as the Marshmallow Experiment, show that those who can better delay their gratification have better life outcomes than those who cannot. The children who succumbed to eating the treat while the researcher was out of the room had worse life outcomes than the children who could wait. This was chalked up to cognitive processes by the originator of the test, while individual differences in these cognitive processes also were used as explanations for individual differences between children in the task. However, it doesn’t seem to be that simple. I did write an article back in December of 2015 on the Marshmallow Experiment and how it was a powerful predictor, but after extensive reading into the subject, my mind has changed. New research shows that social trust has a causal effect on whether or not one would wait for the reward—if the individual trusted the researcher he or she was more likely to wait for the other reward than if they did not trust the researcher, in which they were more likely to take what was offered in the first place.
The famous Marshmallow Experiment showed that children who could wait with a marshmallow or other treat in front of them while the researcher was out of the room, they would get an extra treat. The children who could not wait and ate the treat while the researcher was out of the room had worse life outcomes than the children who could wait for the other treat. These lead researchers to the conclusion that the ability to delay gratification depended on ‘hot’ and ‘cold’ cognitive processes. According to Walter Mischel, the originator of the study method, the ‘cool’ system is the thinking one, the cognitive system, which reminds you that you get a reward if you wait, while the ‘hot’ system is the impulsive system, the system that makes you want the treat now and not want to wait for the other treat (Metcalfe and Mischel, 1999).
Some of these participants were followed up on decades later, and those who could better delay their gratification had lower BMIs (Schlam et al, 2014); scored better on the SAT (Shoda, Mischel, and Peake, 1990) and other tests of educational attainment (Ayduk et al, 2000); along with other positive life outcomes. So it seems that placing a single treat—whether it be a marshmallow or another sweet treat—would predict one’s success, BMI, educational attainment and future prospects in life and that there are underlying cognitive processes, between individuals that lead to differences between them. But it’s not that simple.
After Mischel’s studies in the 50s, 60s and 70s on delayed gratification and positive and negative life outcomes (e.g., Mischel, 1958; Mischel, 1961; Mischel, Ebbeson, and Zeiss, 1972) it was pretty much an accepted fact that delaying gratification somehow was related to these positive life outcomes, while the negative life outcomes were partly a result of the lack of ability to delay gratification. Though in 2014, a study was conducted showing that ability to delay gratification depends on social trust (Michaelson et al, 2013).
Using Amazon’s Mechanical Turk, (n = 78, 34 male, 39 female and 5 who preferred not to state their gender) completed online surveys and read three vignettes in order—trusty, untrustworthy and neutral—while using a scale of 1-7 to note how likeable, trustworthy, and how sharing their likelihood of sharing. Michaelson et al (2013) write:
Next, participants completed intertemporal choice questions (as in Kirby and Maraković, 1996), which varied in immediate reward values ($15–83), delayed reward values ($30–85), and length of delays (10–75 days). Each question was modified to mention an individual from one of the vignettes [e.g., “If (trustworthy individual) offered you $40 now or $65 in 70 days, which would you choose?”]. Participants completed 63 questions in total, with 21 different questions that occurred once with each vignette, interleaved in a single fixed but random order for all participants. The 21 choices were classified into 7 ranks (using the classification system from Kirby and Maraković, 1996), where higher ranks should yield higher likelihood of delaying, allowing a rough estimation of a subject’s willingness to delay using a small number of trials. Rewards were hypothetical, given that hypothetical and real rewards elicit equivalent behaviors (Madden et al., 2003) and brain activity (Bickel et al., 2009), and were preceded by instructions asking participants to consider each choice as if they would actually receive the option selected. Participants took as much time as they needed to complete the procedures.
When one’s trust was manipulated in the absence of a reward, within the group of subjects influenced their ability to delay gratification, along with how trustworthy one was perceived to be, influenced their ability to delay gratification. So this suggests that, in the absence of rewards, when social trust is reduced, ability to delay gratification would be lessened. Due to the issues of social trust manipulation due to the order of how the vignettes were read, they did a second experiment using the same model using 172 participants (65 males, 63 females, and 13 who chose not to state their gender). Though in this experiment, a computer-generated trustworthy, untrustworthy and neutral face was presented to the participants. They were only paid $.25 cents, though it has been shown that the compensation only affects turnout, not data quality (Burhmester, Kwang, and Gosling, 2011).
In this experiment, each participant read a vignette and there was a particular face attached to it (trustworthy, untrustworthy and neutral), which were used in previous studies on this matter. They found that when trust was manipulated in the absence of a reward between the subjects, this influenced the participants’ willingness and to delay gratification along with the perceived trustworthiness influencing it as well.
Michaelson et al (2013) conclude that the ability to delay gratification is predicated on social trust, and present an alternative hypothesis for all of these positive and negative life outcomes:
Social factors suggest intriguing alternative interpretations of prior findings on delay of gratification, and suggest new directions for intervention. For example, the struggles of certain populations, such as addicts, criminals, and youth, might reflect their reduced ability to trust that rewards will be delivered as promised. Such variations in trust might reflect experience (e.g., children have little control over whether parents will provide a promised toy) and predisposition (e.g., with genetic variations predicting trust; Krueger et al., 2012). Children show little change in their ability to delay gratification across the 2–5 years age range (Beck et al., 2011), despite dramatic improvements in self-control, indicating that other factors must be at work. The fact that delay of gratification at 4-years predicts successful outcomes years or decades later (Casey et al., 2011; Shoda et al., 1990) might reflect the importance of delaying gratification in other processes, or the importance of individual differences in trust from an early age (e.g., Kidd et al., 2012).
Another paper (small n, n = 28) showed that the children’s perception of the researchers’ reliability predicted delay of gratification (Kidd, Palmeri, and Aslin, 2012). They suggest that “children’s wait-times reflected reasoned beliefs about whether waiting would ultimately pay off.” So these tasks “may not only reflect differences in self-control abilities, but also beliefs about the stability of the world.” Children who had reliable interactions with the researcher waited about 4 times as long—12 minutes compared to 3 minutes—if they thought the researcher was trustworthy. Sean Last over at the Alternative Hypothesis uses these types of tasks (and other correlates) to show that blacks have lower self-control than whites, citing studies showing correlations with IQ and delay of gratification. Though, as can be seen, alternative explanations for these phenomena make just as much sense, and with the new experimental evidence on social trust and delaying gratification, this adds a new wrinkle to this debate. (He also shortly discusses ‘reasons’ why blacks have lower self-control, implicating the MAOA alleles. However, I have already discussed this and blaming ‘genes for’ violence/self-control doesn’t make sense.)
Michaelson and Munakata (2016) show more evidence for the relationship between social trust and delaying gratification. When children (age 4 years, 5 months, n = 34) observed an adult as trustworthy, they were able to wait for the reward, compared to when they observed the adult as untrustworthy they ate the treat thinking that, since they observed the adult as untrustworthy, they were not likely to get the second marshmallow than if they waited for the adult to return if they believed him to be untrustworthy. Ma et al (2018) also replicated these findings in a sample of 150 Chinese children aged 3 to 5 years old. They conclude that “there is more to delay of gratification than cognitive capacity, and they suggest that there are individual differences in whether children consider sacrificing for a future outcome to be worth the risk.” Those who had higher levels of generalized trust waited longer, even when age and level of executive functioning were controlled for.
Romer et al (2010) show that people who are more willing to take risks may be more likely to engage in risky behavior that provides insights to that specific individual on why delaying gratification and having patience leads to longer-term rewards. This is a case of social learning. However, people who are more willing to take risks have higher IQs than people who do not. Though SES was not controlled for, it is possible that the ability to delay gratification in this study came down to SES, with lower class people taking the money, while higher class people deferred. Raine et al (2002) showed a relationship between sensation seeking in 3-year-old children from Mauritius, which then was related to their ‘cognitive scores’ at age 11. As usual, parental occupation was used as a measure of ‘social class’, and since SES does not capture all aspects of social class then controlling for the variable does not seem to be too useful. Because a confound here could be that children from higher classes have more of a chance to sensation seek which may cause higher IQ scores due to cognitive enrichment. Either way, you can’t say that IQ ’causes’ delayed gratification since there are more robust predictors such as social trust.
Though the relationship is there, what to make of it? Since exploring more leads to, theoretically, more chances to get things wrong and take risks by being impulsive, those who are more open to experience will have had more chances to learn from their impulsivity, and so learn to delay gratification through social learning and being more open. ‘IQ’ correlating with it, in my opinion, doesn’t matter too much; it just shows that there is a social learning component to delaying gratification.
In conclusion, there are alternative ways to look at the results from Marshmallow Experiments, such as social trust and social learning (being impulsive and seeing what occurs when an impulsive act is carried out may have one learn, in the future, to wait for something). Though these experiments are new and the research is young, it’s very promising that there are other explanations for delayed gratification that don’t have to do with differences in ‘cognitive ability’, but depend on social trust—trust between the child and the researcher. If the child sees the researcher is trustworthy, then the child will wait for the reward, whereas if they see the researcher is not trustworthy, they ill take the marshmallow or whatnot, since they believe the researcher is not trustworthy and therefore won’t stick to their word. (I am also currently reading Mischel’s 2014 book Marshmallow Test: Mastering Self-Control and will have more thoughts on this in the future.)
Steroid Mythconceptions and Racial Differences in Steroid Use
2000 words
Steroids get a bad reputation. It largely comes from movies and people’s anecdotal experiences and repeating stories they hear from the media and other forms of entertainment, usually stating that there is a phenomenon called ‘roid rage’ that makes steroid users violent. Is this true? Are any myths about steroids true, such as a shrunken penis? Are there ways to off-set it? Steroids and their derivatives are off-topic for this blog, but it needs to be stressed that there are a few myths that get pushes about steroids and what it does to behavior, its supposed effects on aggression and so forth.
With about 3 million AAS (ab)users (anabolic-androgenic steroids) in America (El Osta et al, 2016), knowing the effects of steroids and similar drugs such as Winny (a cutting agent) would have positive effects, since, of course, athletes mostly use them.
Shrunken testicles
This is, perhaps, one of the most popular. Though the actual myth is that AAS use causes the penis to shrink (which is not true), in reality, AAS use causes the testicles to shrink by causing the Leydig cells to decrease natural testosterone production which then decreases the firmness and shape of the testicles which then results in a loss of size.
In one study of 772 gay men using 6 gyms between the months of January and February (and you need to think of the type of bias there that those people who are ‘Resolutioners’ would be more likely to go to the gym those months), a questionnaire was given to the men. 15 .2 percent of the men had used, with 11.7 percent of them injecting within the past 12 months. HIV positive men were more likely to have used in the past compared to negative men (probably due to scripts). Fifty-one percent of them reported testicular atrophy, and they were more likely to report suicidal thoughts (Bolding, Sherr, and Elford, 2002). They conclude:
One in seven gay men surveyed in central London gyms in 2000 said they had used steroids in the previous 12 months. HIV positive men were more likely to have used steroids than other men, some therapeutically. Side effects were reported widely and steroid use was associated with having had suicidal thoughts and feeling depressed, although cause and effect could not be established. Our findings suggest that steroid use among gay men may have serious consequences for both physical and mental health.
Of course, those who (ab)use substances have more psychological problems than those who do not. Another study of 203 bodybuilders found that 8 percent (n = 17) found testicular atrophy (for what it’s worth, it was an internet survey of drug utilization) (Perry et al, 2005). Another study found that out of 88 percent of individuals who abused the drug complained of side-effects of AAS use, about 40 percent described testicular atrophy (Evans, 1997), while testicular atrophy was noted in about 50 percent of cases (sample size n = 24) (Darke et al, 2016).
Sperm production
One study of steroid users found that only 17 percent of them had normal sperm levels (Torres-Calleja et al, 2001), this is because exogenous testosterone will result in the atrophy of germinal cells which cause a decrease in spermatogenesis. Though, too, increased AAS (ab)use later into life may lead to infertility later in life. Knuth et al (1989) also studied 41 bodybuilders with an average age of 26.7. They went through a huge laundry list of different types of steroids they have taken over their lives. Nineteen of the men were still using steroids at the time of the investigation (group I), whereas 12 of them (group II) stopped taking steroids 3 months prior, while 10 of them (group III) stopped steroid use 4 to 24 months prior.
They found that only 5 of them had sperm counts below the average of 20 million sperm per square ml, while 24 of the bodybuilders showed these symptoms. No difference between group I and II was noticed and group III (the group that abstained from use for 4 to 24 months) largely had sperm levels in the normal range. So, the data suggests that even in cases of severe decrease of sensitivity to androgens due to AAS (ab)use, spermatogenesis may still continue normally in some men, even when high levels of androgens are administered exogenously, while even after prolonged use it seems it is possible for sperm levels to go back to the normal range (Knuth et al 1989).
Aggression and crime
Now it’s time for the fun part and my reason for writing this article. Does (ab)using steroids cause someone to go into an uncontrollable rage, a la the Incredible Hulk when they inject themselves with testosterone? The media has latched into the mind of many, with films and TV shows showing the insanely aggressive man who has been (ab)using AAS. But how true is this? A few papers have shown that this phenomenon is indeed true (Konacher and Workman, 1989; Pope and Katz, 1994), but how true is it on its own, since AAS (ab)users are known to use multiple substances???
Konacher and Workman (1989) is a case study done on one man who had no criminal history, who began taking AASs three months before he murdered his wife, and they conclude that AAS can be said to be a ‘personality changer’. Piacetino et al (2015) conclude in their review of steroid use and psychopathology in athletes that “AAS use in athletes is associated with mood and anxiety disturbances, as well as reckless behavior, in some predisposed individuals, who are likely to develop various types of psychopathology after long-term exposure to these substances. There is a lack of studies investigating whether the preexistence of psychopathology is likely to induce AAS consumption, but the bulk of available data, combined with animal data, point to the development of specific psycho-pathology, increased aggressiveness, mood destabilization, eating behavior abnormalities, and psychosis after AAS abuse/dependence.” I, too, would add that since most steroid abuse are polysubstance abusers (they use multiple illicit drugs on top of AAS), that the steroids per se are not causing crime or aggressive behavior, it’s the other drugs that the steroid (ab)user is also taking. And there is evidence for this assertion.
Lundholm et al (2015) showed just that: that AAS (ab)use was confounded with other substances used while the individual in question was also taking AAS. They write:
“We found a strong association between self-reported lifetime AAS use and violent offending in a population-based sample of more than 10,000 men aged 20-47 years. However, the association decreased substantially and lost statistical significance after adjusting for other substance abuse. This supports the notion that AAS use in the general population occurs as a component of polysubstance abuse, but argues against its purported role as a primary risk factor for interpersonal violence. Further, adjusting for potential individual-level confounders initially attenuated the association, but did not contribute to any substantial change after controlling for polysubstance abuse.“
Even The National Institute of Health (NIH) writes: “In summary, the extent to which steroid abuse contributes to violence and behavioral disorders is unknown. As with the health complications of steroid abuse, the prevalence of extreme cases of violence and behavioral disorders seems to be low, but it may be underreported or underrecognized.” We don’t know whether steroids cause aggression or more aggressive athletes are more likely to use the substance (Freberg, 2009: 424). Clearly, the claims of steroids causing aggressive behavior and crime are overblown and there has yet to be a scientific consensus on the matter. A great documentary on the matter is Bigger, Stronger, Faster, which goes through the myths of testosterone while chronicling the use of illicit drugs in bodybuilding and powerlifting.
This, too, was even seen in one study where men were administered supraphysiologic doses of testosterone to see its effects on muscle size and strength since it had never been tested; no changes in mood or behavior occurred (Bhasin et al, 1996). Furthermore, injecting individuals with supraphysiological doses of testosterone as high as 200 and 600 mg per week does not cause heightened anger or aggression (Tricker et al, 1996; O’Connor et, 2002). Testosterone is one of the most abused AASs around, and if a heightened level of T doesn’t cause crime, nor can testosterone levels being higher this week compared to last seem to be a trigger for crime, we can safely disregard any claims of ‘roid rage’ since they coincide with other drug use (polysubstance abuse). So since we know that supraphysiologic doses of testosterone don’t cause crime nor aggression, we can say that AAS use, on its own (and even with other drugs) does not cause crime or heightened aggression since aggression elevates testosterone secretion, testosterone doesn’t elevate aggression.
One review also suggests that medical issues associated with AAS (ab)use are exaggerated to deter their use by athletes (Hoffman and Ratamess, 2006). They conclude that “Existing data suggest that in certain circumstances the medical risk associated with anabolic steroid use may have been somewhat exaggerated, possibly to dissuade use in athletes.”
Racial differences in steroid use
Irving et al (2002) found that 2.1 percent of whites used steroids, whereas 7.6 percent of blacks did; 6.1 percent of ‘Hispanics’ use them within the past 12 months, and a whopping 14.1 percent of Hmong Chinese used them; 7.9 percent of ‘other Asians’ used them, and 3,1 percent of ‘Native Americans’ did with 11.3 percent of mixed race people using them within the past 12 months to gain muscle. Middle schoolers were more likely to use than high schoolers, while people from lower SES brackets were more likely to use than people in higher SES brackets.
Stilger and Yesalis (1999: 134) write (emphasis mine):
Of the 873 high school football players participating in the study, 54 (6.3%) reported having used or currently using AAS. Caucasians represented 85% of all subjects in the survey. Nine percent were African-American while the remainder (6%) consisted of Hispanics, Asian, and other. Of the AAS users, 74% were Caucasian, 13% African American, 7% Hispanic, and 3% Asian, x2 (4,854 4) 4.203, p 4 .38. The study also indicated that minorities are twice as likely to use AAS as opposed to Caucasians. Cross tabulated results indicate that 11.2% of all minorities use/used AAS as opposed to 6.5% of all Caucasians (data not displayed).
One study even had whites and blacks reporting the same abuse of steroids in their sample (n = 10,850 ‘Caucasians’ and n = 1,883 black Americans), with blacks reporting, too, lower levels of other drug abuse (Green et al, 2001). Studies indeed find higher rates of drug use for white Americans than other ethnies, in college (McCabe et al, 2007). Black Americans also frequently underreport and lie about their drug use (Ledgerwood et al, 2008; Lu et al, 2001). Blacks are also more likely to go to the ER after abusing drugs than whites (Drug Abuse Warning Network, 2011). Bauman and Ennett (1994) also found that blacks underreport drug use whereas whites overreport.
So can we really believe the black athletes who state that they do not (ab)use AAS? No, we cannot. Blacks like about any and all drug use, so believing that they are being truthful about AAS (ab)use in this specific instance is not called for.
Conclusion
Like with all things you use and abuse, there are always side-effects. Though, the media furor one hears regarding AAS and testosterone (ab)use are largely blown out of proportion. The risks associated with AAS (ab)use are ‘transient’, and will subside after one discontinues using the drugs. Blacks seem to take more AAS than whites, even if they do lie about any and all drug use. (And other races, too, seem to use it at higher rates than whites.) Steroid use does not seem to be ‘bad’ if one knows what they’re doing and are under Doctor’s supervision, but even then, if you want to know the truth about AAS, then you need to watch the documentary Bigger, Stronger, Faster. I chalk this up to the media themselves demonizing testosterone itself, along with the ‘toxic masculinity’ and the ‘toxic jock effect‘ (Miller, 2009; Miller, 2011). Though, if you dig into the literature yourself you’ll see there is scant evidence for AAS and testosterone (ab)use causing crime, that doesn’t stop papers like those two by Miller talking about the effects of ‘toxic jocks’ and in effect, deriding masculine men and with it the hormone that makes Men men: testosterone. If taken safely, there is nothing wrong with AAS/testosterone use.
(Note: Doctor’s supervision only, etc)
