
Home » Darwin
Category Archives: Darwin
Mechanisms that Transcend Natural Selection in the Evolutionary Process: Alternatives to Natural Selection
2250 words
Fodor’s argument was a general complaint against adaptationism. Selection can’t be the mechanism of evolution since it can’t distinguish between causes and correlates of causes—so it thusly can’t account for the creation (arrival) of new species. Here, I will provide quotes showing that the claim that natural selection is a mechanism is ubiquitous in the literature—claims that either Darwin discovered the mechanism or claims that it is a mechanism—and that’s what Fodor was responding to. I will then provide an argument combining saltation, internal physiological mechanisms and decimationism and the EES into a coherent explanatory framework to show that there are alternatives to Darwinian evolution, and that these thusly explain speciation and the proliferation of traits while natural selection can’t since it isn’t a mechanism.
Grant and Grant, 2007: “the driving mechanism of evolutionary change was natural selection”
American Museum of Natural History: “Natural selection is a simple mechanism that causes populations of living things to change over time.”
Andrews et al, 2010: “Natural selection is certainly an important mechanism of allele-frequency change, and it is the only mechanism that generates adaptation of organisms to their environments.”
Pianka: “Natural selection is the only directed evolutionary mechanism resulting in conformity between an organism and its environment”
Cottner and Wassenberg, 2020: “This mechanism is natural selection: individuals who inherit adaptations simply out-compete (by out-surviving and out-reproducing) individuals that do not possess the adaptations.”
So natural selection is seen as the mechanism by which traits become fixed in organisms and how speciation happens. Indeed, Darwin (1859: 54) wrote in On the Origin of Species:
“From these several considerations I think it inevitably follows, that as new species in the course of time are formed through natural selection, others will become rarer and rarer, and finally extinct.”
[And some more of the same from authors in the modern day]
“The role of natural selection in speciation, first described by Darwin, has finally been widely accepted” (Via, 2009)
“Selection must necessarily be involved in speciation” (Barton, 2010)
“Darwin’s theory shows how some natural phenomena may be explained (including at least adaptations and speciation)” (SEP, Natural Selection)
“Natural selection has always been considered a key component of adaptive divergence and speciation (2, 15–17)” (Schneider, 2000)
“Natural selection plays a prominent role in most theories of speciation” (Schulter and Nagel, 1995)
So quite obviously, natural selection is seen as a mechanism, and this mechanism supposedly explains speciation of organisms. But since Fodor (2008) and Fodor and Piattelli-Palmarini (2010) showed that natural selection isn’t a mechanism and can’t explain speciation, then there are obviously other ways that evolution happened. There are alternatives to natural selection, and that’s where I will now turn. I will discuss saltation, internal physiological mechanisms and decimationism and then cohere them into a framework that shows how species can arise sans selection.
Explaining speciation
Saltation is the concept of abrupt and substantial changes which lead to the creation of new species, and it challenges phyletic gradualism through natural selection. Instances of sudden genetic alterations along with other goings-on in the environment that lead to things such as directed mutation can eventually result in the emergence of distinct species. Saltation, therefore, challenges Darwinism showing that certain traits can arise quickly, which lead to the emergence of new species within a short time frame. We also have internal physiological mechanisms which play a role in speciation while influencing the development and divergence of traits within biological populations. They don’t rely on external selective pressures—although goings-on in the environment of course can affect physiology—this emphasizes internal factors like developmental constraints, epigenetic modifications and genetic regulatory networks. These can then lead to the expression of novel traits and then on to speciation without the need for external selective forces. And finally decimationism—which emphasizes periodic mass extinction as drivers of evolutionary change—offers another alternative.
Catastrophic events create holes in ecological niches which then allow for the rapid adaptation and diversification of surviving species. So the decimation and recurrent re-colonizing of ecological niches can then lead to the establishment of distinct lineages (species), which then highlight the role of external and non-selective factors in the process of evolution.
So the interaction between saltation, internal physiological mechanisms, and decimationism thusly provides a novel and comprehensive framework for understanding speciation. Sudden genetic changes and other changes to the system can the initiate the development of unique physiological traits (due to the interaction of the developmental resources, and so any change to one resource would cause a cascading change to the system), while internal mechanisms then ensure the stabilization and heritability of the traits within the population. And when this is coupled with environmental upheaval caused by decimation leading to mass extinctions, these processes then contribute to the formation of new species which then offers a framework and novel perspective of the ARRIVAL of the fittest (Darwin’s theory said nothing about arrival, only the struggle for existence), which extends beyond the concept of natural selection.
So if abrupt genetic and other internal changes (saltation) can passively respond to external stimuli and/or environmental pressures, leading to the emergence of distinct traits within a population, and if internal physiological mechanisms influence the expression and development of these traits, then it follows that saltation, coupled with internal physiological mechanisms, can explain and contribute to the rise of new species. If periodic mass extinctions (decimationism) create ecological vacuums and opportunities for adaptive radiation, and if internal physiological mechanisms play a role in the heritability and stability of traits, then it follows that decimationism in conjunction with internal physiological mechanisms can contribute to the speciation of surviving lineages. Also note that all of this is consistent with Gould’s punctuated equilibrium (PE) model.
Punctuated equilibrium was proposed by Gould and Eldgridge as an alternative to phyletic gradualism (Eldgridge and Gould, 1971). It proposes that species evolve rapidly and not gradually. A developmental gene hypothesis also exists for PE (Casanova and Conkel, 2020).
One prediction of PE is rapid speciation events. During periods of punctuated equilibrium, there will be relatively short intervals of rapid speciation which then result in the emergence of new species. This follows from the theory in that it posits that speciation occurs rapidly, concentrated in short bursts, which lead to the prediction that distinct species should emerge more quickly during these punctuated periods. So if species undergo long periods of stasis with occasional rapid change, then it logically follows that new species should arise quickly during these punctuated periods. Seeing that the PE model was developed to explain the lack of transitional fossils, it proposes that species undergo a long period of morphological stasis, with evolutionary changes occurring in short bursts during speciation events, which therefore provides a framework that accounts for the intermittent presence of transitional fossils in the fossil record.
Another prediction is that during periods of stasis (equilibrium), species will exhibit stability in terms of morphology and adaptation. This follows from the theory in that PE posits that stability characterizes a majority of a species existence and that change should occur in quick bursts. Thus, between these bursts, there should be morphological stability. So the prediction is that observable changes are concentrated in specific intervals.
The epigenome along with transposable elements have been argued to be at the heart of PE, and that “physiological stress, associated with major climatic change or invasion of new habitats, disrupts epigenetic silencing, resulting in TE reactivation, increased TE expression and/or germ-line infection by exogenous retroviruses” (Zeh, Zeh, and Ishida, 2009: 715). Further, this hypothesis—that the epigenetic regulation of transposable elements regulates PE—makes testable predictions (Zeh, Zeh and Ishida, 2009: 721). This is also a mechanism to further explain how stress-induced directed mutations occur. Thus, there is an epigenetic basis for the rapid transformation of species which involves the silencing of transposable elements. So calls for an epigenetic synthesis have been made (Crews and Gore, 2012). We, furthermore, know that Lamarckian inheritance is a major mechanism of evolution (Koonin, 2014). We also know that epigenetic processes like DNA methylation contribute to the evolutionary course (Ash, Colot, and Oldroyd, 2021). Such epigenetic mechanisms have been given solid treatment in West-Eberhard’s (2003) Developmental Plasticity and Evolution. (See also West-Eberhard, 2005 on how developmental plasticity leads to the origin of species differences and Wund, 2015 on the impact of phenotypic plasticity on the evolutionary process.)
Integrating the mechanisms into the EES
So in integrating saltation, internal physiological mechanisms, decimationism, epigenetic processes, phenotypic evaluation and directed mutations into the EES (extended evolutionary synthesis), we can then get a more comprehensive framework. Phenotypic plasticity allows organisms to exhibit various phenotypes in response to various environmental cues, so this introduces a broader aspect of adaptability that go beyond genetic change while emphasizing the capacity of populations to change based on what is going on in the immediate environment during development.
Generic drift and neutral evolution also at a role. So beyond the selective pressures emphasized by the modern synthesis, the EES recognizes that genetic changes can occur through stochastic mechanisms which then influence the genetic constitution of a population. Evo-devo then contributes to the synthesis by highlighting the role of developmental processes in evolutionary outcomes. Thus, by understanding how changes in gene regulation during development contribute to morphological diversity, evo-devo therefore provides insight into evolutionary mechanisms which transcend so-called natural selection.
Moreover, the integration of epigenetic inheritance and cultural evolution also extends the scope of the EES. Epigenetic mechanisms can influence gene expression without a change to the DNA sequence, and can contribute to heritability and adaptability. Cultural evolution, then, while acknowledging the power of transmitted knowledge and practices on adaptive success, also broadens our understanding of evolution beyond biological factors. Thus, by incorporating all of the discussed mechanisms, the EES fosters a unique approach in integrating numerous different mechanisms while recognizing that the evolutionary process is influenced by a mixture of biological, environmental, cultural and developmental factors. There is also the fact that the EES has better predictive and explanatory power than the modern synthesis—it also makes novel predictions (Laland et al, 2015).
Conclusion
This discussion has delved into diverse facets of evolutionary theory, showed that natural selection is seen as a mechanism in the modern day, that Darwin and modern day authors see natural selection as the mechanism of speciation, and has considered a few mechanisms of evolution beyond natural selection. Fodor’s argument was introduced to question the applicability of “selection-for” traits, and challenged the notion of natural selection as a mechanism of evolutionary change. Fodor’s argument therefore paved the way for the mechanisms I discussed and opened the door for the reevaluation of saltation, internal physiological mechanisms, decimationism and the EES more broadly in explaining the fact of evolution. So this discussion has shown that we have to think about evolution not as selection-centric, but in a more holistic manner.
There are clearly epigenetic mechanisms which influence speciation on a PE model, and these epigenetic mechanisms then also contribute to the broader understanding of evolution beyond PE. In the PE model, where speciation events are characterized by rapid and distinct changes, epigenetic mechanisms play a crucial role in influencing the trajectory of evolutionary transitions. These epigenetic mechanisms, then, continue to the heritability of traits and the adaptability of populations. These epigenetic mechanisms also extend beyond their impact of speciation within the PE model. So by influencing gene expression in response to environmental cues, epigenetic changes then provide a dynamic layer to the evolutionary process which allow populations to adapt more rapidly to changing conditions. Therefore, epigenetic mechanisms become integral components in explaining evolutionary dynamics which then align with the principles of the EES.
The integration of these concepts into the EES then further broadens our understanding of evolution. So by incorporating genetic drift, phenotypic plasticity, evo-devo, epigenetic inheritance, directed mutation, and cultural evolution, the EES provides a comprehensive framework which recognizes the complexity of evolutionary process beyond mere reductive genetic change. Phenotypic plasticity allows organisms to be adaptively plastic to respond to cues during development and change the course of their development to respond to what is occurring in the environment without relying solely on genetic changes. Genetic drift then introduces stochastic processes and neutral evolution. Evo-devo then contributes to the synthesis by highlighting the role of developmental processes in evolutionary outcomes. Epigenetic inheritance also brings a non-genetic layer to heritability, acknowledging the impact of environmentally responsive gene regulation. Cultural evolution then recognizes the transmission of knowledge and practices within populations as a factor which influences adaptive success. So putting this all together, these integrations then suggests that evolution is a multifaceted interplay of irreducible levels (Noble, 2012) which then challenges natural selection as a a primary or sole mechanism of evolution and as a mechanism at all, since we can explain what natural selection purports to explain without reliance on it.
So if evolutionary processes encompass mechanisms beyond natural selection like saltation, internal physiological mechanisms, decimationism, punctuated equilibrium, and phenotypic plasticity, and if we are to reject natural selection as an explanation for trait fixation and speciation based on Fodor’s argument, and if these mechanisms are an integral part of the EES, then the EES offers a more comprehensive framework in understanding evolution. Evolutionary processes do encompass mechanisms beyond natural selection as evidenced by critiques of selection-centric views and those views that are seen as alternatives to natural selection like saltation, internal physiological mechanisms and decimationism. Thus, by incorporating the aforementioned mechanisms, we will have a better understanding evolution than if merely relying on the non-mechanism of natural selection to explain trait fixation and sp
Directed Mutations, Epigenetics and Evolution
2400 words
A mutation can be said to be directed if it arises due to the needs of the developing organism, and they occur at higher frequencies if it is beneficial (Foster, 2000; Saier et al, 2017). If there is some sort of stress, then an adaptive mutation would occur. The existence of this kind of mechanism has been debated in the literature, but its existence spells trouble for neo-Darwinian theory, whose proponents claim that mutations are random and then “selected-for” in virtue of their contributions to fitness. Indeed, this concept challenges a core tenet of neo-Darwinism (Sarkar, 1991). I will argue that directed mutation/non-random mutation/stress-directed adaptation (DM, directed mutation for short) spells trouble for the neo-Darwinian paradigm.
The issue at hand
The possibility of DMs were argued for by Cairns, Overbaugh, and Miller (1988), where they argue that environmental pressure can cause adaptive changes to genes that would be beneficial to the organism. This then spurred a long debate about whether or not such mutations were possible (see Sarkar, 1991; Fox Keller, 1992; Brisson, 2003; Jablonka and Lamb, 2014). Although Cairns, Overbaugh, and Miller were wrong—that is, they were not dealing with mutations that were due to the environmental disturbances they posed (Jablonka and Lamb, 2014: 84)—their paper did bring up the possibility that some mutations could be a direct consequence of environmental disturbances which would then be catapulted by the homeodynamic physiology of the organism.
Saier et al (2017) state the specific issue with DM and its existence:
Recently, strong support for directed mutation has emerged, not for point mutations as independently proposed by Cairns, Hall and their collaborators, but for transposon-mediated mutations (12, 13). If accepted by the scientific community, this concept could advance (or revise) our perception of evolution, allowing increased rates of mutational change in times of need. But this concept goes against the current dogma that states that mutations occur randomly, and only the beneficial ones are selected for (14, 15). The concept of directed mutation, if established, would require the reversal of a long accepted precept.
This is similar to the concept of phenotypic plasticity. It is the phenomenon of a given genotype expressing different phenotypes due to environmental factors. This concept is basically a physiological one. When talking about how plastic a phenotype is, its relation to the physiology of the organism is paramount. We know that physiological changes are homeodynamic. That is, changes in physiology are constantly happening due to the effects of the environment the organism finds itself in. For example, acute changes in heart rate occur due to what happens in the environment, like say a predator chase it’s prey. The heart rates of both predator and prey increases as blood flow increases due to stress hormones. I will discuss phenotypic plasticity on its own in the future, but for now I will just note that genetic and environmental factors influence the plasticity of phenotypes (Ledon-Rettig and Ragsdale, 2021) and that phenotypic plasticity and development play a role in evolution (West-Eberhard, 2003, 2005; Wund, 2015
The fact of the matter is, phenotypic plasticity is directly related to the concept of directed mutation, due to DM being a largely physiological concept. I will argue that this refutes a central Darwinian premise. Namely that since directed mutations are possible, then they are not random. If they are not random, then due to what occurs during the development of an organism, a directed mutation could be adaptive. This, then, is the answer to how phenotypic traits become fixed in the genome without the need for natural selection.
Directed mutations
Sueoka (1988) showed that basically all organisms are subject to directed mutations. It has been noted by mathematicans that on a purely random mutational model, that there would not be enough time to explain all of the phenotypic diversity we see today (Wright, 2000). Doubt is placed on three principles of neo-Darwinism: mutations occur independently of the environment the organism is in (this is empirically false); mutations are due to replication errors (this is true, but not always the case) and mutation rates are constant (Brisson, 2003).
One of the main claims of the neo-Darwinian paradigm is that mutations occur at random, and the mutation is selected-for or against based on its relationship to fitness. Fodor’s argument has refuted the concept of natural selection, since “selection-for” is an intensional context and so can’t distinguish between correlated traits. However, we know now that since physiology is sensitive to the environment, and since adaptive changes to physiology would occur not only in an organism but during its development, it then follows that directed mutations would be a thing, and so they wouldn’t be random as neo-Darwinian dogma would claim.
In her review Stress-directed adaptive mutations and evolution, Wright (2004) concludes:
In nature, where cell division must often be negligible as a result of multiple adverse conditions, beneficial mutations for evolution can arise in specific response to stressors that target related genes for derepression. Specific transcription of these genes then results in localized DNA secondary structures containing unpaired bases vulnerable to mutation. Many environmental stressors can also affect supercoiling and [stress-directed mutation] directly.
But what are the mechanisms of DMs? “Mechanism” in this meaning would “refer to the circumstances affecting mutation rates” (Wright, 2000). She also defines what “random” means in neo-Darwinian parlance: “a mutation is random if it is unrelated to the metabolic function of the gene and if it occurs at a rate that is undirected by specific selective conditions of the environment.” Thus, the existence of DMs would then refute this tenet of neo-Darwinism. Two of the mechanisms of such DMs are transcriptional activation and supercoiling. Transcriptional activation (TA) can cause changes to single-stranded DNA (ssDNA) and also supercoiling (the addition of more coils onto DNA). TA can be caused by either derepression (which is a mechanism which occurs due to the absence of some molecule) or induction (the activation of an inactive gene which then becomes transcribed). Thus, knowing this, “genetic derepression may be the only mechanism by which particular environmental conditions of stress target specific regions of the genome for higher mutation rates (hypermutation)” (Wright, 2000). Such responses rely on a quick response, and this is due to the plastic phenotypes of the organism which then allow such DMs to occur. It then follows that stress-induced changes would allow organisms to survive in new environments, without a need for neo-Darwinian “mechanisms”—mainly natural selection. Thus, the biochemical mechanism for such mutations is transcriptional activation. Such stress-directed mutation could be seen as “quasi-Lamarckian” (Koonin and Wolf, 2009).
In nature, nutritional stress and associated genetic derepression must be rampant. If mutation rates can be altered by the many variables controlling specific, stress-induced transcription, one might reasonably argue that many mutations are to some extent directed as a result of the unique metabolism of every organism responding to the challenges of its environment. (Wright, 2000)
This is noted wonderfully by Jablonka and Lamb (2014: 92) in Evolution in Four Dimensions:
No longer can we think about mutation solely in terms of random failures in DNA maintenance and repair. We now know that stress conditions can affect the operation of the enzyme systems that are responsible for maintaining and repairing DNA, and parts of these systems sometimes seem to be coupled with regulatory elements that control how, how much, and where DNA is altered.
Jablonka and Lamb present solid evidence that mutations are semi-directed. Such mutations, as we have seen, are able to be induced by the environment in response to stress, which is due to our plastic, homeodynamic physiology. They discuss “four dimensions” of evolution which are DNA, epigenetic, behavioral and cultural. Their works (including their Epigenetic Inheritance and Evolution: The Lamarckian Dimension; see Jablonka and Lamb, 2015) provide solid evidence and arguments against the neo-Darwinian view of evolution. The fact of the matter is, there are multiple inheritance systems over and above DNA, which then contribute to nonrandom, directed mutations. The fact of the matter is, Lamarckism wasn’t wrong and Jablonka and Lamb have strongly argued for that conclusion. Epigenetics clearly influences evolution, and this therefore vindicates Lamarckism. Epigenetic variation can be inherited too (Jablonka and Lamb, 1989). Since phenotypic plasticity is relevant in how organisms adapt to their environment, then epigenetic mechanisms contribute to evolution (Ashe, Colot, and Oldroyd, 2021). Such changes that arise due to epigenetic mechanisms can indeed influence mutation (Meyer, 2015), and I would say—more directly—that certain epigenetic mechanisms play a part in how an adaptive, directed mutation would arise during the development of an organism. Stochastic epigenetic variation can indeed become adaptive (Feinberg and Irizarry, 2010).
Non-random mutations have been known to be pretty ubiquitous (Tsunoyama, Bellgard, and Gojobori, 2001). This has even been shown in the plant Arabidopis (Monroe et al, 2022), which shows that basically, mutations are not random (Domingues, 2023). A similar concept to DMs is blind stochasticity. Noble and Noble (2017, 2018; cf Noble, 2017) have shown that organisms harness stochastic processes in order to adapt to the environment—to harness function. A stochastic process is a state of a system that cannot be predicted even knowing the current state of said system.
Even all the way back in 1979, such changes were beginning to be noticed by evolutionists, such as Ho and Saunders (1979) who write that variations in the phenotype
are produced by interactions between the organism and the environment during development. We propose, therefore, that the intrinsic dynamical structure of the epigenetic system itself, in its interaction with the environment, is the source of non-random variations which direct evolutionary change, and that a proper study of evolution consists in the working out of the dynamics of the epigenetic system and its response to environmental stimuli as well as the mechanisms whereby novel developmental responses are canalized.
The organism participates in its own evolution (as considerations from niche construction show), and “evolutionary novelties” can and do arise nonrandomly (Ho, 2010). This is completely at-odds with the neo-Darwinian paradigm. Indeed, the creators of the Modern Synthesis ignored developmental and epigenetic issues when it came to formulating their theory. Fortunately, in the new millennium, we have come to understand and appreciate how development and evolution occur and how dynamic the physiological system itself truly is.
There have been critical takes on the concept of DM (Lenski and Mittler, 2003; Charlesworth, Barton, and Charlesworth, 2017; see Noble and Shapiro, 2021 for critique), like for example Futuyama (2017) who claims that DM is “groundless.” However, James Shapiro’s (1992; 2013, 2014) concept of natural genetic engineering states that cells can restructure their genomes so this “means viewing genetic change as a coordinated cell biological process, the reorganization of discrete genomic modules, resulting in the formation of new DNA structures” (Shapiro, 1993). DNA is harnessed by and for the physiological system to carry out certain tasks. Since development is self-organizing and dynamic (Smith and Thelen, 2003; Saetzler, Sonnenschein, and Soto, 2012) and since development is spurred on by physiological processes, along with the fact that physiology is sensitive to the goings-on of the environment that the developing organism finds itself in, then it follows that mutations can and would arise due to need, which would refute claims from neo-Darwinians who claim that mutations arise due to chance and not need.
Conclusion
It is clear that mutations can be (1) adaptive and (2) environmentally-induced. Such adaptive mutations, clearly, arise due to need and not chance. If they arise due to need and not chance, then they are directed and adaptive. They are directed by the plastic physiology of the organism which constructs the phenotype in a dialectical manner, using genes as its passive products, not active causes. This is because biological causation is multi-leveled, not one-way (Noble, 2012). There is also the fact of the matter that “genetic change is far from random and often not gradual” (Noble, 2013).
As can be seen in this discussion, adaptive, directed mutations are a fact of life, and so, one more domino of neo-Darwinism has fallen. Berkley claims that “The genetic variation that occurs in a population because of mutation is random“; “mutations are random“, but as we’ve seen here, this is not the case. Through the biological process of physiology and its relationship to the ebbs and flows of the environment, the organism’s phenotype that is being constructed by the self-organizing system can respond to changes in the cellular and overall environment and thusly direct changes in the phenotype and genes which would then enhance survival due to the environmental insult.
Lamarckism has been vindicated over the past 25 or so years, and it’s due to a better understanding of epigenetic processes in evolution and in the developing organism. Since what Lamarck is known for is the claim that the environment can affect the phenotype in a heritable manner, and since we now know that DNA is not the only thing inherited but epigenetically-modified DNA sequences are too, it follows that Lamarck was right. What we need to understand development and evolution is the Extended Evolutionary Synthesis, which does make novel predictions and predictions that the neo-Darwinian paradigm doesn’t (Laland et al, 2015).
Such directed changes in the genome which are caused by the physiological system due to the plastic nature of organismal construction refute a main premise of the neo-Darwinian paradigm. This is the alternative to neo-Darwinian natural selection, as Fodor noted in his attack on neo-Darwinism:
The alternative possibility to Darwin’s is that the direction of phenotypic change is very largely determined by endogenous variables. The current literature suggests that alterations in the timing of genetically controlled developmental processes is often the endogenous variable of choice; hence the ‘devo’ in ‘evo-devo’.
Darwin got quite a bit wrong, and it’s of no fault of his own. But those who claim that Darwin discovered mechanisms or articulated the random process of mutations quite obviously need to update their thoughts in the new millennium on the basis of new information informed by systems biologists and epigeneticists. The process of the construction of organisms is dynamic and self-organizing, and this is how phenotypic traits become fixed in populations of organisms. Plasticity is in fact a major driver of evolution along with the concept of genetic assimilation, which results in the canalization of the plastic trait which then eliminates the plastic response from the environment (Sommer, 2020). Phenotypic plasticity can have adaptive traits arise, but natural selection can’t be the mechanism of evolution due to Fodor’s considerations. Development can lead to evolution, not only evolution leading to development (West-Eberhard, 2003). In fact, development in many cases precedes evolution.
Natural Selection is not an Explanatory Mechanism
2450 words
Darwin proposed, back in 1859, that species arose due to natural selection—the pruning of deleterious genetic variations in a population, which led to the thinking that the “inherent design” in nature, formerly thought to be due to a designer (“God”) was due to a force Darwin called “natural selection” (NS). The line of reasoning is thus: (1) two individuals of the same population are mostly the same genetically/phenotypically, but have small differences between them, and one of the small differences is a difference in a trait needed for survival. (2) But both traits can contribute to fitness, how does NS ‘know’ to select for either coextensive trait? Now think about two traits: trait T and trait T’. What would explain the fixation of either trait in the population we are discussing? NS is not—cannot—be the mechanism of evolution.
In 2010, philosopher Jerry Fodor and cognitive scientist Massimo Piattelli-Palmarini, wrote a book titled “What Darwin Got Wrong“, which argued that NS is not a causal mechanism in regard to the formation of new species. Their argument is (pg 114):
- Selection-for is a causal process.
- Actual causal relations aren’t sensitive to counterfactual states of affairs: if it wasn’t the case that A, then the fact that it’s being A would have caused its being B doesn’t explain its being the case that B.
- But the distinction between traits that are selected-for and their free-riders turns on the truth (or falsity) of relevant counterfactuals.
- So if T and T’ are coextensive, selection cannot distinguish the case in which T free-rides on T’ from the case that T’ free-rides on T.
- So the claim that selection is the mechanism of evolution cannot be true.
This argument is incredibly strong. If it is true, then NS cannot be the mechanism by which evolution occurs; NS is not—nor can it be—the mechanism of evolution. So, regarding the case of two traits that are coextensive with each other, it’s not possible to ascertain which trait was selected-for and which trait was the free-rider. NS cannot distinguish between two locally coextensive traits, so, therefore, it is not an explanatory mechanism and does not explain the evolution of species, contra Darwin. It cannot be the mechanism that connects phenotypic variation with fitness variation.
The general adaptationist argument is: “(1) the claim that evolution is a process in which creatures with adaptive traits are selected and (2) the claim that evolution is a process in which creatures are selected for their adaptive traits” (Fodor and Piattelli-Palmarini, 2010: 13). Darwinists are committed to inferring (2) from (1), though it is fallacious. It is known as the intensional fallacy.
“Due to the intensionality of “select-for” and “trait”, one cannot infer from ‘Xs have trait t and Xs were selected’ to ‘Xs were selected for having trait t’” (Fodor and Piattelli-Palmarini, 2010: 139). How does one distinguish from a trait that was selected-for and a free-rider that hitched a ride on the truly adaptive trait for the organism in question? The argument provided above shows that it is not possible. “Darwinists have a crux about free-riding because they haven’t noticed the intensionality of selection-for and the like; and when it is brought to their attention, they haven’t the slightest idea what to do about it” (Fodor and Piattelli-Palmarini, 2010: 16).
No observation can show whether or not trait T or T’ was selected-for in virtue of its contribution to fitness in a given population; favoring one story over another in regard to the adaptation of a trait in question, therefore, does not make any logical sense due to the problem of free-riders (and, also, favoring one story over another is due to bias for the like of the specific adaptive just-so story in question). For if two traits are coextensive—meaning that traits coincide with one another—then how can NS—which does not have a mind—‘know’ to “select-for” whichever trait contributes to fitness in the population in question? Breeders are the perfect example.
Breeders have minds and can therefore select for certain traits and against undesirable traits; however, of course, since NS does not have a mind, this is not the case when it comes to naturally selected traits (so-called), since NS does not have a mind. NS cannot explain the distribution of phenotypic traits throughout the world; there is no agent of NS nor are there ‘laws of selection’, therefore NS is not an explanatory mechanism. Explanations based on NS are based only on correlations with traits and fitness, not on causes themselves (this critique can be extended to numerous other fields, too). The problem with relying only on correlations between traits and fitness is two-fold: (1) the trait in question can be irrelevant to fitness and (2) the trait in question can be a free-rider.
Creatures have traits that increase fitness because they were selected-for, the story goes. NS explains why the creature in question has trait T, which increases fitness in environment E. One can then also make the claim that the selection of the trait in question was due to the increased fitness it gave the creature. However, if this claim is made, “then the theory of natural selection would reduce to a trait’s being a cause of reproductive success [which then] explains its being a cause of reproductive success which explains nothing (and isn’t true).”
So since genetically-linked traits are coextensive with an infinitude of different possible outcomes, then the hypothesis that trait X is an adaptation is underdetermined by all possible observations, which means that NS cannot explain how and why organisms have the traits they do, since NS cannot distinguish between two coextensive traits, since NS lacks a mind and agency.
NS can be said to be an explanation if and only if two conditions are met: (1) if NS can be understood as acting on counterfactuals and (2) if NS can be said to be acting on any physical evolutionary laws.
(1) A counterfactual is an “if-clause”, which is contrary to a fact. A counterfactual is a statement that cannot be true, for example, “I hear but I have no ears” or “I see but I have no eyes.” Thus, if it were possible for NS to be an explanation for the continuance of a specific trait that is linked to other traits (that is, they are coextensive) in a given population, it would need to—necessarily—invoke a counterfactual about NS. It would need to be the case that the trait in question would still be selected for in the absence of free-riders. As an example from Fodor and Piattelli-Palmarini (2010: 103) a heart pumps blood (what it was selected-for) and makes pumping sounds (its linked free-rider). Thus, if the pumping of blood and the sound that blood-pumping makes were not coextensive, then the pumping, not the pumping sounds, get selected for.
There is a huge problem, though. Counterfactuals are intentional statements; they refer to concepts found in our minds, not any physical things. NS does not have a mind and thus lacks the ability to “select-for” since “selecting-for” is intentional. Therefore NS does not act on counterfactuals; it is blind to the fact of counterfactuals since it does not have a mind.
(2) It does not seem likely that there are “laws of selection”. Clearly, the adaptive value of any phenotype depends on the environment that the organism is in. Fodor and Piattelli-Palmarini (2010: 149) write (emphasis theirs):
The problem is that it’s unlikely that there are laws of selection. Suppose that P1 and P2 are coextensive but that, whereas the former is a property that affects fitness, the latter is merely a correlate of a property that does. The suggestion is that all this comes out right if the relation between P1 and fitness is lawful, and the relation between P2 and fitness is not. …it’s just not plausible that there are laws that relate phenotypic traits per se to fitness. What (if any) effect a trait has on fitness depends on what kind of phenotype is embedded in, and what ecology the creature that has the trait inhabits. This is to say that, if you wish to explain the effects that a phenotypic trait has on a creature’s fitness, what you need is not its history of selection but its natural history. And natural history offers not laws of selection but narrative accounts of causal chains that lead to the fixation of phenotypic traits. Although laws support counterfactuals, natural histories do not; and, as we’ve repeatedly remarked, it’s counterfactual support on which distinguishing the arches from the spandrels depends.
There is, too, a simple example regarding coextensive traits and selection. Think of the lactase gene. It is well-known that we humans are adapted to drink milk—and the cause is gene-culture coevolution that occurred at around the time of cow domestication (Beja-Perreira et al, 2003; Gerbalt et al, 2011). No one disputes the fact that gene-culture coevolution is how and why we can drink milk. But what people do dispute is the adaptive just-so story (Gould and Lewontin, 1976; Lloyd, 1999; Richardson, 2007) that was made to explain how and why the trait went to fixation in certain human populations. Nielsen (2009) writes (emphasis mine):
The difference in lactose intolerance among human geographic groups, is caused by a difference in allele frequencies in and around the lactase gene (Harvey et al. 1998; Hollox et al. 2001; Enattah et al. 2002; Poulter et al. 2003). The cause for the difference in allele frequencies is primarily natural selection emerging about the same time as dairy farming evolved culturally (Bersaglieri et al. 2004). Together, these observations lead to a compelling adaptive story of natural selection favoring alleles causing lactose tolerance. But even in this case we have not directly shown that the cause for the selection is differential survival due to an ability/inability to digest lactose. We must acknowledge that there could have been other factors, unknown to us, causing the selection acting on the region around the Lactase gene. Even if we can argue that selection acted on a specific mutation, and functionally that this mutation has a certain effect on the ability to digest lactose, we cannot, strictly speaking, exclude the possibility that selection acted on some other pleiotropic effect of the mutation. This argument is not erected to dispute the adaptive story regarding the lactase gene, the total evidence in favor of adaptation and selection related to lactose tolerance is overwhelming in this case, but rather to argue that the combination of a functional effect and selection does not demonstrate that selection acted on the specific trait in question.
Selection could have acted on a free-rider that is coextensive with the lactase gene, and just because “the story fits the data” well (that’s a necessary truth; of course the story can fit the data because any story can be formulated for any data) does not mean that it is true, that the reason for trait T is reason R since they “fit the data so well.”
Of course, this holds for EP, evolutionary anthropology, and my favorite theory for the evolution of human skin color, the vitamin D hypothesis. I do not, of course, deny that light skin is needed in order to synthesize vitamin D in climates with low UVB; that is a truism. What is denied is the fact that selection acted on light skin (and its associated/causal genes); what is denied is the combination of functional effect and selection. Just-so stories are necessarily true; they, of course, fit any data because one can formulate any story to fit any data points they have. Thus, Darwinists are just storytellers who have a bunch of data; there is no way to distinguish between the selection of a trait because it increased fitness and the selection of a free-rider that is “just there” that does not increase fitness, but the thing that increases fitness is what the free-rider “rode in on.”
NS is not and cannot be an explanatory mechanism. Darwinism has already been falsified (Jablonka and Lamb, 2005; Noble, 2011; Noble, 2012; Noble, 2017) and so, this is yet another nail-in-the-coffin for Darwinism. The fact that traits that are coextensive means that NS would have to “know” which trait to act on; NS cannot “know” which of the coextensive traits to act on (because it has no mind) and, NS cannot be a general mechanism that connects phenotypic variation to variation in fitness. NS does not explain the evolution of species, nor can NS distinguish between two locally coextensive traits—traits T and T’—because NS has no agency and does not have a mind. Therefore NS is not an explanatory mechanism. Just invoking NS to explain the continuance of any trait fails to explain the survival of the trait because NS cannot distinguish between traits that enhance an organism’s fitness and free-riders which are irrelevant to survival but are coextensive with the selected-for trait, as long as the traits in question are coextensive.
P1) If there is selection for T but not T’, various counterfactuals must be true.
P2) If the counterfactuals are true, then NS must be an intentional-agent, or there must be laws about “selection-for”.
P3) NS is mindless.
P4) There are no laws for “selection-for”.
∴ It is false that selection for T but not T’ occurs in a population.
One then has two choices:
(1) Argue that NS has a mind and therefore that it can “select for” certain traits that are adaptable in a given population of organisms in the environment in question. “Select-for” implies intention. Intentional acts only occur in organisms with minds. Intentional states are only possible if something has a mind. Humans are the only organisms with minds. Humans are the only organisms that can act intentionally. NS does not have a mind. (Animal breeder’s are an example that can select-for desirable traits and against undesirable traits because animals breeder’s are humans and humans can act intentionally.) Therefore NS does not act intentionally since it does not have a mind. I don’t think anyone would argue that NS has a mind and acts intentionally as an agent, therefore P3 is true.
(2) Argue that there are laws for “selection-for” phenotypic traits related to fitness. But it’s not possible that there are laws that relate to the selection of a phenotype, per se, in a given population. The effect of a trait depends on the ecology of the organism in question as well as its natural history. Therefore, to understand the effects of a phenotypic trait on the fitness of an organism we must understand its natural history, not its selection history (so-called). Therefore P4 is true.
There are no laws for “selection-for”, nor does NS have a mind that can select a trait that lends to an organism’s fitness and not a trait that’s just correlated with the trait in question
There is No ‘Marching Up the Evolutionary Tree’
2000 words
The notion that there is any ‘progress’ to evolution is something that I have rebutted countless times on this blog. My most recent entry being Marching Up the ‘Evolutionary Tree’? which was a response to Pumpkin Person’s article Marching up the evolutionary tree. Of course, people never ever change their views in a discussion (I have seen it, albeit it is rare) due, mainly to, in my opinion, ideology. People have so much time invested in their little pet theories that they cannot possibly fathom at the thought of being wrong or being led astray by shoddy hypotheses/theories that confirm their pre-existing beliefs. I will quote a few comments from Pumpkin Person’s blog where he just spews his ‘correlations with brain size and ‘splits’ on the ‘evolutionary tree” that ‘proves that evolution is progressive’, then I will touch on two papers (I will cover both in great depth in the future) that directly rebut his idiotic notion that so-called brain size increases across our evolutionary history (and even before we became humans) are due to ‘progress in evolution’
One of my co-bloggers Phil wrote:
I think you mistyped that, but i see your point. Problem, however, most of your used phylogenies were unbalanced.
Based on the definition you provided, but not based on any meaningful definition. To me, an unbalanced tree is . . .
This is literally meaningless. Keep showing that you’ve never taken a biology class in your life, it really shows.
All it is is ignorance to basic biological thinking, along with an ideology to prove his ridiculous Rushtonian notion that ‘brain size increases prove that evolution is progressive’.
You have yet to present ANY scientific logic, and my argument about taxonomic specificity is clearly beyond you.
Scientific logic?! Scientific logic?! Please. Berkely has a whole page on misconceptions on evolution that directly rebut his idiotic, uneducated views on evolution. It doesn’t help that his evolution education most likely comes from psychologists. Nevertheless, PP’s ‘argument’ is straight garbage. Taxonomic specificity’ is meaningless when you don’t have an understanding of basic biological concepts and evolution. (I will have much more to say on his ‘taxonomic specificity’ below.)
PP writes:
Was every tree perfect? No, but most were pretty close, and keep in mind that any flawed trees would have the effect of REDUCING the correlation between brain size/encephalization and branching, because random error is a source of statistical noise which obscures any underlying relationship. So the fact that I repeatedly found such robust correlation in spite of alleged problems with my trees, makes my conclusions stronger, not weaker.
The fact that you ‘repeatedly’ found ‘correlations’ in spite of the ‘problems’ with your trees makes your ‘conclusions’ weaker. Comparing organisms over evolutionary time and you notice a ‘trend’ in brain size. Must mean that evolution is progressive and brain size is its calling card!!
PP writes:
I’m right and all the skeptics you cite are wrong.
Said like a true idealogue.
Here is where PP’s biggest blunder comes in:
It’s not how many splits they have that I’ve been measuring, it’s how many splits occur on the tree before they branch off. Here’s a source from 2017:
Eukaryotes represent a domain of life, but within this domain there are multiple kingdoms. The most common classification creates four kingdoms in this domain: Protista, Fungi, Plantae, and Animalia.
So you needed ‘a source from 2017’ to tell you something that is literally taught on the first day of biology 101? Keep showing how uneducated you are here.
Nothing fallacious about a correlation between number of splits and brain size/encephalization.
Post hoc, ergo propter hoc is a Latin phrase for “after this, therefore, because of this.” The term refers to a logical fallacy that because two events occurred in succession, the former event caused the latter event.[1][2]
…
Magical thinking is a form of post hoc, ergo propter hoc fallacy, in which superstitions are formed based on seeing patterns in a series of coincidences. For example, “these are my lucky trousers. Sometimes good things happen to me when I wear them.”
P1: X happened before Y.
P2: (unstated) Y was caused by something (that happened before Y).
C1: Therefore, X caused Y.
Here is PP’s (fallacious) logic:
P1: splits (X) happened before Y (brain size increase)
P2: (unstated) brain size increase was caused by something (that happened before brain size increaes [splits on the tree])
C1: therefore, splits caused brain size increase
Now, I know that PP will argue that ‘splits on the evolutionary tree’ denote speciation which, in turn, denotes environmental change. This is meaningless. You’re still stating that Y was caused by something (that happened before Y) and therefore inferring that X caused Y. That is the fallacy (which a lot of HBD theories rest on).
PP writes:
You don’t get it. Even statistically insignificant correlations become significant when you get them FIVE TIMES IN A ROW. If you want to believe it was all a coincidence, then fine.
Phylogenies are created from shared derived factors. Berkely is the go-to authority here on this matter. (No that’s not appeal to authority.) Biologists collect information about a given animal and then infer the evolutionary relationship. Furthermore, PP’s logic is, again, fallacious. Berkely also has tips for tree reading, which they write:
Trees depict evolutionary relationships, not evolutionary progress. It’s easy to think that taxa that appear near one side of a phylogenetic tree are more advanced than other organisms on the tree, but this is simply not the case. First, the idea of evolutionary “advancement” is not a particularly scientific idea. There is no unbiased, universal scale for “advancement.” Second, taxa with extreme versions of traits (which might be perceived as more “advanced”) may occur on any terminal branch. The position of a terminal taxon is not an indication of how adaptive, specialized, or extreme its traits are.
He may emphatically argue (as I know he will) that he’s not doing this. But, as can be seen from his article, X is ‘less advanced’ than Y, therefore splits, brain size, correlation=progress. This is dumb.
For anyone who wants to know how (and how not to) read phylogenies, read Gregory (2008). These idotic notions that PP espouses are what Freshman in college believe due to ‘intuitiveness’ about evolution. It’s so rampant that biologists have writen numerous papers on the matter. But some guy with a blog and no science background (and an ideology to hammer) must know more than people who do this for a living (educate people on phylogenies).
On Phil’s response to see the Deacon paper that I will discuss below, PP writes:
That’s not a rebuttal.
Yes it is, as I will show shortly.
The first paper I will discuss is Deacon’s (1990) paper Fallacies of Progression in Theories of Brain-Size Evolution. This is a meaty paper with a ton of great ideas about phylogenies, along with numerous fallacies that people go to when reading trees (my favorite being the Numerology fallacy, which PP uses, see below).
Deacon argues that since people fail to analyze allometry, this anatomists have mistaken artifacts for evolutionary trends. He also argues that many structural’brain size increases’ from ‘primitive to advanced forms’ (take note here, because this is what PP did and this is what discredits his idiotic ideology) are the result of allometric processes.
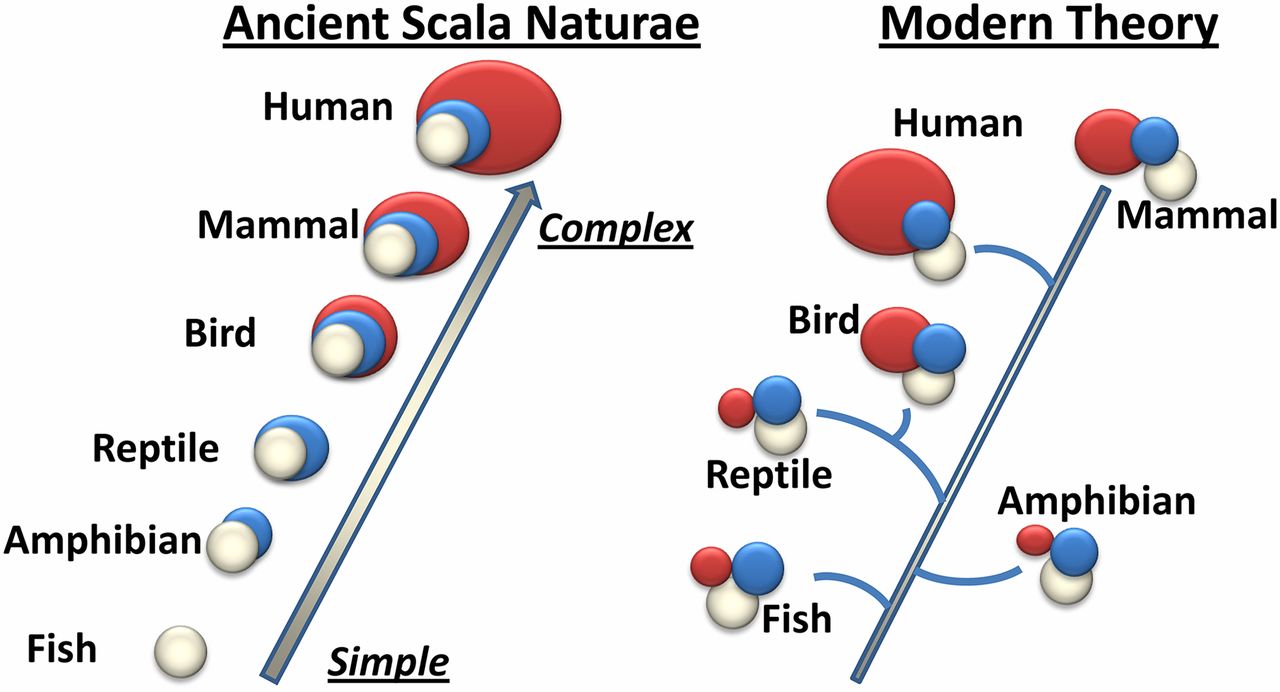
Source: Evolution of consciousness: Phylogeny, ontogeny, and emergence from general anesthesia Mashour and Alkire (2013)
This paper (and picture) show it all. This notion of scala naturae (which Rushton (2004) attempted to revive with r/K selection theory has been rebutted by me) was first proposed by Aristotle. We now know how the brain structure evolved, so the old ‘simple scala naturae‘ is, obviously, out of date in the study of brain evolution.
This paper is pretty long and I don’t have time to discuss all of it so I will just provide one quote that disproves PP’s ‘study’:
Whenever a method is discovered for simplifying the representation of a complex or apparently nonsystematic numerical relationship, the method of simplification itself provides new insight into the phenomenon under study. But reduction of a complex relationship to a simple statistic makes it far easier to find spurious relationships with other simple statistics. Numerology fallacies are apparent correlations that turn out to be artifacts of numerical oversimplification. Numerology fallacies in science, like their mystical counterparts, are likely to be committed when meaning is ascribed to some statistic merely by virtue of its numeric similarity to some other statistic, without supportive evidence from the empirical system that is being described.
Deacon also writes in another 1990 article titled Commentary on Ilya I. Glezer, Myron So Jacobs, and Peter J Morgane (1988) Implications of the “initial brain’9 concept for brain evolution in Cetacea:
The study of brain evolution is one of the last refuges for theories of progressive evolution in biology, but in this field its influence is still pervasive. To a great extent the apparent “progress” of mammalian brain evolution vanishes when the effects of brain size and functional specialization are taken into account.
(It’s worth noting that in the author’s response to Deacon, he did not have any qualms about ‘progressive brain-size’.)
In regards to PP’s final ‘correlation’ on human races and brain-size, this is a perfect quote from McShea (1994: 1761):
If such a trend [increase in brain size leading to ‘intelligence’] in primates exists and it is driven, that is, if the trend is a direct result of concerted forces acting on most lineages across the intelligence spectrum, then the inference is justified. But if it is passive, that is, forces act only on lineages at the low-intelligence end, then most lineages will have no increasing tendency. In that case, most primate species—especially those out on the right tail of the distribution like ours—would be just as likely to lose intelligence as to gain it in subsequent evolution (if they change at all).
The ‘trend’ is passive. Homo floresiensis is the best example. We are just as likely to lose our ‘intellect’ and our ‘big brains’ as we are to ‘get more intelligent’ and ‘smaller brains’. The fact of the matter is this: environment dictates brain size/whatever other traits an organism has. Imagine a future environment that is a barren wasteland. Kilocalories are scarce; do you think that humans would keep their big brains—which are two percent of their body weight accounting for a whopping 25 percent of total daily energy needs—without enough high-quality energy? When brain size supposedly began to increase in our taxa is when erectus learned to control fire and cook meat (Hlublik et al, 2017).
All in all, there is no ‘progress’ to evolution and, as Deacon argues, so-called brain-size increases across evolutionary time disappear after adjustments for body size and functional specialties are taken into account. However, for the idealogue who looks for everything they can to push their ideology/worldview, things like this are never enough. “No, that wasn’t a rebuttal! YOU’RE WRONG!!” Those are not scientific arguments. If one believes in ‘evolutionary progress’ and that brain-size increases are the proof in the pudding that evolution is ‘progressive’ (re has a ‘direction’), then they must rebut Deacon’s arguments on allometry and his fallacies in his 1990 paper. Stop equating evolution with ‘progress’. Though, I can’t fault laymen for believing that. I can, however, fault someone who supposedly enjoys the study of evolution. You’re wrong. The people you cite (who are out of their field of expertise) are wrong.
Evolution is an amazing process. To equate it with ‘progress’ does not allow one to appreciate the beauty of the process. Evolution does carry baggage with it, and if I weren’t so used to the term I would use Descent by Modification (DbM, which is what Darwin used). Nevertheless, progressionists will hide out in whatever safehold they can to attempt to push their idealogy that is not based on science.
(Also read Rethinking Mammalian Brain Evolution by Terrence Deacon. I go more in depth on these three articles in the future.)
Happy Darwin Day, Heathens
1650 words
Today is Darwin’s 208th birthday and the 158th year since the publication of On the Origin of Species by Means of Natural Selection or the Preservation of Favoured Races in the Struggle of Life. So many people get Darwin wrong. They either have never read his books, or are taking a secondhand account. You can tell who has never read his writings in his own words and who takes snippets of his writings to use them for ideological purposes. I was going to wait until I finished The Descent of Man (Darwin, 1871) until I wrote this article but Darwin Day seems like the best time to do it.
People call themselves ‘Darwinists’ when it’s clear they’ve never read his writings. And due to this, people have large misconceptions on cherry-picked quotes and then use it for their pet ideology—not even attempting to understand the context around what he wrote. One large misquotation you may see around the Internet may put charges of ‘racism’ on Darwin since he ‘believed’ that the ‘higher’ races of Man will one day exterminate the ‘lower’ races. PumpkinPerson is guilty of this, writing in his article Darwin’s terrifying prediction:
Sadly, if HBD is correct, I think there probably will be natural selection favoring higher IQ populations, in fact it’s already happening. In sub-Saharan Africa, we see the more primitive cultures like pygmies and Bushmen losing more and more territory and their populations declining.
Which is based on a (misinterpreted) Darwin quote from his book Descent of Man (1871 (2004): 132-3); note: I have the Barnes n Noble edition):
The great break in the organic chain between man and his nearest allies, which cannot be bridged over by any extinct or living species, has often been advanced as a grave objection to the belief that man is descended from some lower form; but this objection will not appear of much weight to those who, convinced by general reasons, believe in the general principle of evolution. Breaks incessantly occur in all parts of the series, some being wide, sharp and defined, others less so in various degrees; as between the orang and its nearest allies—between the Tarsius and the other Lemuridae—between the elephant and in a more striking manner between the Ornithorhynchus or Echidna, and other mammals.
But all these breaks depend merely on the number of related forms which have become extinct. At some future period, not very distant as measured by centuries, the civilised races of man will almost certainly exterminate and replace throughout the world the savage races. At the same time the anthropomorphous apes, as Professor Schaaffhausen has remarked, will no doubt be exterminated. The break will then be rendered wider, for it will intervene between man in a more civilised state, as we may hope, than the Caucasian, and some ape as low as a baboon, instead of as at present between the negro or Australian and the gorilla.
This is the big quote. The quote that supposedly what paints Darwin as a ‘racist’ and one of the many, many instances of quote-mining from Creationists attempting to discredit his theory of evolution through natural selection. But here’s the thing that people fail to realize: without the rest of the context, you won’t know what he’s saying because the very next paragraph writes (pg 132):
With respect to the absence of fossil remains, serving to connect man with his ape-like progenitors, no one will lay much stress on this fact who reads Sir C. Lyell’s discussion, where he shows that in all the vertebrate classes the discovery of fossil remains has been a very slow and fortuitous process. Nor should it be forgotten that those regions which are the most likely to afford remains connecting man with some extinct ape-like creature, have not as yet been searched by geologists.
So, the whole quote taken in context, it seems he was defending his theory showing that even though no there was an “absence of fossil remains” connecting us to our apelike ancestors.
This book was written 12 years after On the Origin, so knowing that and then seeing the rest of the omitted context behind the controversial quote (and, of course, how Creationists quote-mine and attempt to twist and turn words), what do you think he was saying? To me, it looks like he was defending his theory and addressing critics who said that the fossil record does not support his claims. In fact, Darwin and other Naturalists of the time didn’t separate culture and biology and thus used a blend of both. Darwin was simply observing that a slight advantage between races of men would, after time, lead to the creation of a new species. You’d have to have actually read his books to know that, though.
PP’s other post on Darwin, Did Darwin believe in HBD? he writes (referring to the previous quote-mine):
What it looks like is Darwin describing an evolutionary hierarchy: Caucasian > negro/Australoid > gorilla > baboon.
If you’re looking for something, you’re going to find it. Complete misrepresentation of Darwin’s words, and just reading Descent of Man will let you know how grossly incorrect this interpretation really is.
Darwin only meant that Caucasians would replace savage races because of their cultural superiority; biological superiority had nothing to do with it. And are we also supposed to believe that Darwin’s predicted demise of gorillas was also for cultural, not biological reasons?
PP, read the whole context and tell me if that’s how you still interpret it. It is worth noting that the quotes are taken from a part of the book that has the subsection: On the Birthplace and Antiquity of Man, which lends more credence to the fact that he was defending his theory from detractors (due to the names he brought up and his prose, in context) who needed to see ‘transitional’ fossils between ape and man.
Further, since PP is using a Creationist quotation, then a Creationist rebuttal is apt here:
First of all, Darwin is making a technical argument as to the “reality” of species, particularly Homo sapiens in this case, and why there should still be apparently distinct species, if all the different forms of life are related by common descent through incremental small changes. His answer is that competition against those forms with some, even small, advantage tends to eliminate closely related forms, giving rise to an apparent “gap” between the remaining forms. Whether or not Darwin was right about that is irrelevant to the use of this quote mine, of course, since that is part of the context that the creationists using it have assiduously removed.
Irony aside that an atheist is using a Creationist quote-mine to prove biological differences, this shows how people who’ve never read his writing can misinterpret what he really meant.
Darwin was also a huge abolitionist, which is never brought up in these discussions. He argued, for his whole life, that slavery should be abolished. He also came from an extremely abolitionist family, so any charges of ‘racism’ to Darwin seem pretty far off the mark.
PP says:
According to liberals, Darwin only meant that Caucasians would replace savage races because of their cultural superiority; biological superiority had nothing to do with it. And are we also supposed to believe that Darwin’s predicted demise of gorillas was also for cultural, not biological reasons?
According to people that know what they’re talking about, Darwin meant that closely related organisms even will a small advantage will replace the other, and that will give rise to a ‘gap’ between organisms. Learn the context behind the whole quote, instead of what Creationists quote-mine. And biological superiority doesn’t exist.
Never mind that Darwin’s theory of natural selection was actually based on biology, not culture.
Can natural selection NOT occur because of cultural differences? Say, two genetically similar populations and one has the native culture and the other with a new, alien culture and they have to use it to adapt to a new environment. Would that be an example of culture and its effect on natural selection?
Never mind that Darwin’s own cousin (Francis Galton) was the father of HBD.
Nothing to do with Darwin himself.
Never mind that Darwin’s own book on natural selection was subtitled The Preservation of Favoured Races in the Struggle for Life.
Which of the great cabbage races will survive? He used race as a term for varieties.
Why let facts get in the way of a convenient rationalization.
Why don’t you tell me?
They just come up with increasingly creative rationalizations to deny the truth, and the effort this takes makes them more and more psychologically invested in denying inconvenient realities.
Ironic….
People who quote Darwin should most definitely read his works, as if they’re quoting him—especially in these contexts—they should really know the whole context behind the quote and not rely on a Creationist quote-mine which is easily dismantled.
And the way it’s going now, the savage races are outbreeding the civilized races—so how do you see (your interpretation of) Darwin’s theory coming to pass? How will your race war fantasy with each of the macro-races genociding the rest of the ethnies in their group and form one ethnicitu of that racial group? PP believes that eventually it’ll be Ashkenazi Jews vs. East Asians for East Asia. Except Ashkenazi Jews frequently breed with gentiles, and in 100 years there will be very few Ashkenazi Jews left. Japan is having a huge population decline, which is partly biological and partly cultural/environmental in nature. One of the so-called ‘most evolved’ ethnies isn’t able to reign superior over the rest of the inferior ethnies/races due to low birthrates? As I said last night: civilization is dysgenic and leads to low birth rates. So how will the civilized races exterminate the savage races, if the civilized races hardly breed because they get too civilized?
