
Home » Race Realism (Page 12)
Category Archives: Race Realism
Social Constructivists about Race are Realists about Race
1750 words
“Race is just a social construct!” is a claim that one who ventures on this side of the internet is likely to hear. People who believe that race is a biological—and social—reality would then say that they are denying the reality of race—that they are saying that race does not exist. But think about the phrase “Race is just a social construct.” In the phrase, we should not assume that there is anything in it which states that the one who is using it is saying that “Race does not exist.” On the contrary: they ARE being realists about race, but in a social, not biological, manner.
Even those who drafted the new AAPA Statement on Race and Racism state that ““race” as a social reality — as a way of structuring societies and experiencing the world — is very real.” They further state that (my emphasis):
… race has become a social reality that structures societies and how we experience the world. In this regard, race is real, as is racism, and both have real biological consequences.
What people do not understand is that just by saying that something is “socially constructed” it does not mean that something “is not real.” For, if social constructivists about race did not believe in the reality of race, what would they have to talk about if what they were talking about did not exist?
Taking a social constructivist approach to race does not mean that one denies its reality. In fact, one of the most impactful arguments for the existence of race identifies race as a social construct of a biological reality (Spencer, 2014). We socially construct races based on what we see—what we see correlates to geographic locales. However, note the socially construct part of the previous phrase. Just that one part can denote so much when it comes to race and its reality.
Social constructivists about race deny that race exists as a biological reality but admit that it exists as a social reality. This then allows them to argue that “racial inequality becomes embodied—literally—in the biological well-being of racialized groups and individuals” (Gravlee, 2009). Gravlee (2009) explains how race becomes biologicized through people’s social expectations of certain races. So, as Gravlee argues, there are biological consequences of racializing social groups.
This is just as Kaplan (2010) argues: although the concept of folk races is false (I disagree), the fact that this concept is still deployed in American thought implies that this is part of what drives social inequalities between social groups that are racialized to be races (what Hardimon, 2017 terms “socialraces“). So, these socially/historically constructed races can and do mean big things when it comes to biology. We can biologicize social groups that we take to be racial groups (i.e., make them socialraces), and this can then drive health disparities between the now-racialized groups.
Now, when people state that “Race is a social construct without biological backing”, they are not denying that what we call “races” captures a portion of genetic diversity—it does, capturing about 4.3 percent of genetic variation between humans is accounted for by folk races (Rosenberg et al, 2002).
Spencer (2011) even identifies social constructivisits about race as realists about race in his Table 1.:
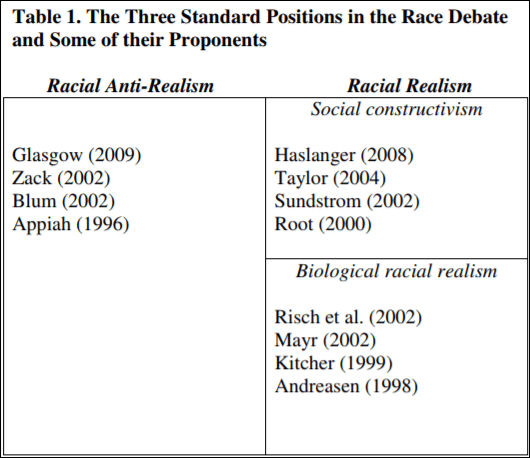
Kaplan and Winther (2014) even identify social constructivists as racial realists, writing:
Social racial realism defends the existence of distinct human groups in our ordinary discourse and social interactions. Such groups are often identified and stabilized by “surface” factors such as skin color or facial features.
(I would be the first of their racial realists, the biogenomic cluster/race, while Jensen, Lynn, and Rushton would be what they term biological racial realists. I would then put Kaplan (2010) and Gravlee (2009) into the social racial realist category.) Kaplan and Winther (2014) are antirealists about biological race. When racialists hear people denying that race exists, that’s what they mean—they are antirealists about biological race.
So, race can exist as a social and political reality, but not exist as the biological racial realists would claim. That is because what we—as a society—impute onto racialized groups, i.e., social races, does have make a difference in regard to the type of health outcomes, for example, that constructivists about race care about. So while they are antirealist about biological races, they hold that those who uphold the existence of biological races do, in part, drive negative health disparities between racialized groups.
Social constructivists about race talk about race in a social manner. When it comes to Hardimon’s (2017) concept of socialrace, it picks out three different but related social kinds: a kind of social group, a social position, and a social system. To Hardimon (2017: 131):
The concept SOCIALRACE refers to
(1) a social group that is taken to be a racialist tace,
(2) the social position occupied by a particular social group that is a socialrace, or
(3) the system of social psoitions that are socialraces.
Socialraces are social groups because they are developed by social relations and defined by social properties. In this way, socialraces are not biological at all because they are not developed by biological relations and defined by biological properties. In this way, the concept of socialrace is distinct from all of Hardimon’s (2017) other race concepts—it, and it alone, constitutes the social dimension of race (this is the difference between Hardimon’s and Spencer’s (2014) concept of race—Hardimon separates the social and biological, Spencer combines both). And so, a socialrace is a social group that is taken to be a racialist race
Hardimon (2017: 132) writes:
Socialrace (the phenomenon) is social in something like the sense in which money and marital status are. Without the requisite social practices, there would be no money, no marriage, no socialrace. Spcoa;race differs straightforwardly social instutions, however, in that its sociality is disguised. The concept SOCIALRACE represents socialraces as social groups whose sociality is hidden—not from the critical theorist already in possession of the concept SOCIALRACE, of course, but from the ordinary individual in society. Socialraces are social groups that appear to be biological. The fact that they are social races goes unseen.
So socialraces are hierarchical in nature—some socialraces, such as whites, are dominant whereas other socialraces, such as blacks, are subordinate. So members of one dominant socialrace can oppress members of another subordinate socialrace. The fact that socialraces are taken to be racialist races—that the belief that people are members of biological races is necessary for the existence of socialraces is true.
Hardimon (2017: 132) writes
SOCIALRACE is strongly nonbiological. It is logically possible tht socialreaces could be found in a world in which the apparent differences in human skin color were actually the result of the nocturnal activity of race fairies, spray-painting hues of black, brown, yellow, red, and pink on the skin of sleeping souls in the dead of night. What makes a person a member of a socialrace are social practices (which include the local application of specific criteria for individual racial classifications), not the possession of biological features. The concept SOCIALRACE is free of any implication that there are even partially biological races. It does not represent socialrace as a “social-natural” kind, representing it instead as a social kinf that falsely purports to be a biological kind.
So we can see that, in Hardimon’s view, socialraces are groups taken to be racialist races—they are falsely thought of as being biologicized, and so, the “social” aspect of these groups is hidden, and only the “apparent” biological nature of these groups is seen. Though do note how SOCIALRACE cannot be both biological and social—it can only be one and it is, of course, social.
Further, minimalist race is a correlate of socialrace. However, just because minimalist race is a correlate of socialrace does not mean that there is a corresponding minimalist race for every socialrace. The best example is “Latinos/Hispanics.” They are a group that are racialized and can be said to be a socialrace, but they have no minimalist race correlate since they are an amalgamation of different races/ethnic groups. No minimalist race functions as the correlate of “Latinos/Hispanics.” “The Irish were a socialrace at one point bu are probably not a minimalist race. We can say that Jews have been a socialrace without committing to ourselves to the claim that they are a minimalist race” (Hardimon, 2017: 136).
So Hardimon’s socialrace concept is a distinctive race concept and satisfies a genuine representational need. Hardimon’s concept puts forth a way of thinking about social groups which are then racialized as biological racial groups, the way that these socialraces occupy social positions, and the system of social structures of which those positions are parts (of which those parts are socialraces). Hardimon’s concept, indeed, does not show that race “does not exist” or “is not real”, since he even says that “‘Constructivisim’ is often used in such a way that to say X is socially constructed is to say that X is unreal. As I use the term, ‘constructivism’ carries no such implication” (Hardimon, 2017: 136). Of course, there could carry that very implication, that whatever is socially constructed does not exist outside of what is doing the socially constructing.
In sum, social constructivists about race are realists about race. Indeed, they need to be realists about race in order for their proposed solutions to work. For if they were not realists about race, what are they fighting for? When people say “Those who say ‘race is a social construct’ deny the existence of race”, they mean to bring up antirealists about biological race, not social constructivists about race. Race, whether you take the biological or social/political view, clearly exists. And accepting one concept over the other dictates what you think about the so-called racial groups in question.
While Spencer’s (2014) race concept already has the social and biological parts inherent in them, Hardimon (2017) separates them—from racialist races, to minimalist race, to populationist race and finally socialrace. Socialraces have a minimalist race correlate—though not all of them do. In any case, socialraces have certain institutions that they are more likely to inhabit. The way that these socialraces occupy social positions, and the system of social structures of which those positions are part are what the concept of socialrace elaborates on.
Therefore, social constructivists about race are realists about race—and there can be no other way for them to be about race. They may be antirealists about biological race, but they are not antirealists about race all the way—they have to believe that race exists, or else what they are doing holds no weight in the world.
The White-Black Swimming/Drowning Gap and Disparities in Elite Running and Swimming Competitions
1600 words
Blacks are better sprinters and whites are better swimmers. Why is this? A whole slew of factors influence this—social, physiological, anatomic. However, there is a stereotype about blacks that has been repeated since I was a child: that blacks can’t swim. How true is this? If it is true, what explains it? It is my opinion that it is true, and that social, cultural, and anatomic and physiologic factors account for this. The same for whites and running. Black children drown at a rate of about 3 times higher than white children. About 70 percent of black children cannot swim, compared to 60 percent of “Hispanic” children and 40 percent of white children. Why is that? Well, one of the most telling answers why is anatomic. Irwin et al (2011) note in their study that blacks are more likely to be “aquaphobes”—having a fear of water—compared to whites.
Almost three years ago, I wrote White Men Can’t Jump? That’s OK, Black Men Can’t Swim. In the article, I explain how and why blacks have a harder time swimming than whites. One anatomic reason is their chest cavity. Compared to whites, blacks have a narrower chest cavity. They have denser, shallower chests. This is a burden while swimming, since those who have a wider chest can take longer strides with their arms while swimming. Blacks have denser bones than whites (Ettinger et al, 1997), Swimmers have lower bone density than non-swimmers (Gomez-Bruton et al, 2013), and so, high bone density is not conducive to swimming success, either.
The first black man to make the swim team for America in the Olympics was Anthony Ervin in 2000. (Funny story. In a class I took a few years ago, racial differences in sports came up. I brought up race differences in swimming. A black guy behind me said “My grandfather was the first to qualify for the Olympics.” I said “Yea? Your grandfather is Anthony Ervin?” He didn’t say anything, it seemed like he got mad at me for calling him out.) That it took this long for a black man to qualify for the US in swimming is telling, and anatomy and physiology, in my opinion, are how we can explain the observed disparity,
So, blacks have lower body fat (on average), and narrower chest cavities. These two things play a role in why blacks are not good swimmers. Yet another role-player, could be, the fact that black women don’t want their hair to get wet and so never taught their children how to swim, parental encouragement, to “swimming is something that white people do” (Wiltse, 2014). Who knows? Maybe in the coming years, blacks could match whites at swimming. Though, with what we know about anatomy and physiology of elite swimmers, this is highly unlikely. It’s like saying “Who knows? Maybe in the coming years, whites could match blacks at running.” Our knowledge of anatomy and physiology throws a wrench in claims like that.
The phenomenon of fast black sprinters and fast white swimmers is predictable through physics (Bejan, Jones, and Charles, 2010). The finalists of running competitions are continuously black, whereas in the swimming competitions they are continuously white. What accounts for this? Well, other than the factors discussed above, there is one more: center of mass.
It is well-known that different races have different anatomic measurements. Blacks have longer limbs than whites (Gerace et al, 1994; Wagner and Heyward, 2000) and longer legs and smaller muscle circumferences (e.g., calves, arms), then they have a higher center of mass than an individual of the same height. So since Asians and whites have long torsos (i.e., since they are endomorphic), they have a lower centers of mass. Asians have the tallest sitting heights, matched with people of the same height, and so we would expect them to be exceptional swimmers. However, since they are not as tall as whites, they do not set records. Blacks, on the other hand, have a lower sitting height when matched with someone of the same height—3 cm shorter. Whites’ sitting height was lower than Asians, but whites are taller so whites dominate swimming compared to Asians because of their average center of mass. See Table 3 from Bejan, Jones, and Charles (2010).

So the difference between blacks’ and whites’ center of mass is 3 percent. This 3 percent difference can account for why the two races excel in running and swimming. When it comes to the runners (blacks), the 3 percent increase in center of mass translates to a 1.5 percent increase in winning speed for the 100 m dash, and a 1.5 percent decrease in winning time, from 10 s to 9.85 s, for example. So the 3 percent difference in running is a huge advantage for blacks.
When it comes to whites, the same holds, except for swimming. So the 3 percent increase in correct length for whites translates over to a 1.5 increase in winning speed and a 1.5 decrease in winning time.
So for taller athletes, mass that falls from a higher altitude falls faster, down and forward; speed increases with larger physiques. So since blacks have larger physiques than whites, then, at the extremes of elite sports (running), their mass allows them to fall down forward, faster and since they have larger physiques, they are faster. So world records are set by athletes with different centers of mass: black athletes in running and white athletes in swimming.
Shifting away from physics, we will now discuss the cultural/social component. The fact that many blacks do not know how to swim became apparent after the Red River drownings of 2010 (Wiltse, 2014). Wiltse (2014) notes three reasons why blacks may be bad swimmers compared to whites: (1) white swimmers denied blacks access to pools; (2) cities provided few pools to black communities and the pools they did provide were small; and (3) the cities closed many public pools after desegregation occurred. White parents taught their children how to swim, but black parents hardly ever did. As this occurred as swimming became popular in American culture, this could be one reason why blacks aren’t as represented in swimming when compared to whites.
Wiltse’s (2014) argument is that past discrimination to blacks from whites when it came to swimming explains the drowning disparity between the races. Whites passed down their swimming knowledge, whereas blacks had little to no chance to pass theirs down—if they even knew how to swim, that is. This type of cultural transmission could explain most—if not all—of the disparity in drowning between the races. It is simple: to address the disparity, the claim that swimming is “what white people do” needs to be addressed. I would assume that this claim grew from the 60s and desegregation from when blacks were barred from swimming pools, as Wiltse (2014) notes. While the swimming and drowning gap can be closed, the elite sports (running and swimming) gap cannot be—as most of what drives the relationship between race and those sports are anatomic and physiologic in nature, combined with numerous other irreducible variables.
However, pointing to these types of cultural/social causes can be reversed. We can say that since white parents don’t teach their children how to sprint and thus they have not taught their children how to sprint for successive generations, then if white parents did just that then whites would begin to close the gap when it comes to sprinting. While I do not deny that we would have more black swimmers had these types of discriminatory acts had not occurred, it is ridiculous to claim that the two races can and will become equal if this were to occur. It’d be like saying that if we train this person from birth to become an elite sprinter then they would be. Though the right analogy would be that since there are fewer whites than blacks in elite running sports, then what explains the disparity is that they are not trained that way from pretty much conception. However this betrays the systems view of athleticism, and while there are necessary factors in regard to running success, the whole system must be looked at when assessing what makes an elite athlete.
In conclusion, there are many anatomic and physiologic reasons why blacks and whites differ in running and swimming sports. Anatomic differences, such as center of mass, explain the disparities in swimming and running. Blacks’ morphology—long limbs and short torso—is conducive to running success. They can take longer strides and take fewer strides a race compared to someone of the same size that does not have the same limb length. When it comes to white swimmers, where the altitude is set by the body rising out of the water, whites hold a 1.5 percent speed advantage in swimming.
Though there are these anatomic differences that lend themselves to differences in elite sporting competitions, these differences do not lend themselves do the swimming and drowning gap in regard to blacks and whites. What explains those gaps is generational access to swimming pools; blacks were barred access to swimming pools just as they started to become popular in America, after the 60s when the country was desegregated. This led to swimming being looked at as “something that white people do”, and so, fewer and fewer black parents taught their children how to swim. Further cultural and social factors explain this, too. While I would assume that some of these aforementioned factors would then play a role in the black-white swimming/drowning gaps, I doubt that it would count for a super-majority of it. Thus, the gap can be closed by ridding the stigma that swimming is “something that white people do”.
The elite sporting gap in running and swimming, however, cannot be closed.
Usain Bolt—Anatomy and Physiology of a Champion: Bolt’s Record Times Can Be Better
1100 words
Usain Bolt is one of—if not the—fastest men who has ever lived. At age 12 he was already the fastest boy in his school (Irving, 2010: 54). At the 2009 Berlin Olympics, he set the world record for 100 m race (Bolt also holds the world record for the 200 m dash, at 19.19s), clocking in at 9.58 seconds. His average speed was 27.8 mph with an average speed of 23.5 mph. Why is Bolt so fast? Of course, there are multiple interacting factors that contribute to Bolt’s world record times. Bolt’s somatotype, muscle fibers, will to win, intense training, mind, etc all contribute to his world record—along with the type of athlete he is. In this article I will discuss what Bolt does, his anatomy and physiology, what lead up to his record-breaking time, and a possible challenge to his record.
Usain Bolt is tall, as far as sprinters go, with a height of 6’5”. Since he is so tall, compared to other sprinters, his average stride length is at the extreme upper-limit of modern sprinters. So what makes Bolt unique as a sprinter is his stride length (Shinabarger, Hellrich, and Baker, 2010). So Bolt has to take fewer strides than other sprinters, which, in part, explains his success.
During Bolt’s record-setting 9.69 s dash in 2008, during the last 2 seconds—when 20 meters were left to run—Bolt looked to the side and started celebrating (Eriksen et al, 2009). Bolt’s coach claimed that he would have shattered even his future record-setting performance of 9.58 s running 9.52 s or better. The runner-up of this race was Richard Thompson. By 4 s, Bolt and Thompson were neck-and-neck, so Bolt’s medal was won between 4 and 8 s. After 8 s, Bolt considerably decelerated while Thompson equalizes and surpasses Bolt. Thompson could not match Bolt’s speed, though, and slows down after 8.5 s. Then, to answer the question “How fast would Bolt have run had he not celebrated the last 2 s?”, Eriksen et al (2009: 226) make two assumptions:
Assumption 1: Bolt matches Thompson’s speed at up to 8 s.
Assumption 2: Bolt maintains a 0.5 m/s2 higher acceleration than Thompson at 8.5 s.
Of course the justification for A1 is obvious: Bolt outran Thompson between 4 and 8 s. But in regard to A2, it is difficult to quantify exactly how much stronger Bolt was than Thompson, since Bolt is a 200m specialist, they take the 0.5 m/s2. So, in two scenarios that Eriksen et al (2009) put forth, the world record would either be 9.61 s or 9.55 s. Eriksen et al (2009: 228) conclude “that a new world record of less than 9.5 s is within reach by Usain Bolt in the near future.” And what do you know: a year later, Bolt ran the 100 m at 9.59 s.
Bolt has a slow reaction to the gun—that is, he has more moving to do to get to the sprinting start since he is so tall. His reaction time at the Beijing Olympic Final was 0.165 s. So if he could reduce his reaction to the gun by .3 s then he would have beaten his world record of 9.58 s to 9.56 s. If he could get it down to 0.12 s then he would be looking at a 9.55 s time, and if he could get it down to as fast as the rules allow—at 0.10 s—then his time would have been 9.53 seconds, almost right there by his coach’s prediction had he not celebrated during his record-setting run (Darrow, 2012).
Since Bolt is so tall—taller than his competitors—he can take fewer steps per 100 m. For instance, he set his record time in 2009 taking 41 steps to win, whereas his competitors took 45 steps (Beneke and Taylor, 2010). The average sprinter has a higher proportion of type II fibers compares to type I fibers (Zierath and Hawley, 2004). So one thing that separates Bolt from his contemporaries is superior biomechanical efficiency along with relative power generated per-step (Beneke and Taylor, 2010; Coh et al, 2018). So Bolt’s record-setting performances comes down to anthropometric characters, coordinated motor abilities, his ability to generate power, and an effective running technique. Sprint performance on the force generated during ground contact.
Bolt has an ectomorphic-dominant somatotype. Since he is ecto-dominant, this gives him certain advantages over more endo- and meso-dominant competitors. Furthermore, along with his body type, Bolt is Jamaican. Most of the ancestry found in Jamaicans is derived from West Africa. Jamaicans are more likely to have the RR ACTN3 genotype (Scott et al, 2010), while the RR genotype—along with type II fibers (with a greater cross-section area) contributes to whole muscle performance during high-velocity contractions (Broos et al, 2016). I am not aware of any analyses of Bolt’s genotype, but I would bet what’s in my bank account that he has the RR genotype—that he has two copies of ACTN3.
Tyson Gay then emerged as a challenger to Bolt (in 2013, Gay gave a dirty urine for PEDs, performance-enhancing drugs, and Bolt said that Gay should be “kicked out of the sport“). Varlet et al (2015) state that Bolt and Gay influenced how fast the other ran in Berlin, 2009. Both Bolt’s and Gay’s steps were pretty much synchronized with each other. Though since Gay was slightly behind Bolt in the race, he had the better chance to synchronize his movement with Bolt’s. However, Blikslager and de Poel (2017) argue against this: they state that there is no sufficient evidence for the claim that Bolt and Gay had synchronized movements.
The center of mass in blacks is around 3 percent higher in blacks than it is in whites. This 3 percent difference in center of mass between whites and blacks leads to them doing better in one sport over another: sprinting for blacks and swimming for whites (this is one reason why blacks are worse swimmers than whites). Further, for runners, the 3 percent increase in center of height translates over to a 1.5 percent increase in running speed, translating to a difference of 10 s compared to 9.85 s (Bejan, Jones, and Charles, 2010). So the change is 0.15 s for runners. This is yet another reason why Bolt excels: he is exceptionally tall.
Bolt is really tall compared to his contemporaries; Bolt goes through insane training (as do his contemporaries). Of course, the explanation for Bolt’s running success is due to numerous factors, including (but not limited to) his height, leg length/stride length, running economy, Vo2 max, training, where he grew up, and a whole slew of other—irreducible—factors. The fact that Bolt could have set an even more unbelievable record had he not celebrated with 2 s—or 20 meters—left during his record-setting run is incredible. That he can even hit at or near to what his coach predicted that he would have gotten had he not celebrated, while getting his reaction time better is even more incredible. Bolt does not even need to improve his running skill to become better—just improve his reaction to the gun and he will, in my opinion—set records that no one wull ever break.
Kenyan Marathon Runners
1850 words
The Boston Marathon is one of the oldest continuous running marathons around. The 122nd just finished today, and—surprise surprise—a Kenyan man and Ethiopian woman took first place. For the men, Lawrence Chereno (time at 2:07: 57) barely edged out the second place winner Lelisa Desisa Benti (2:07:59; an Ethiopian) while for the women Worknesh Degefa (2:23:31) beat Edna Kiplagat (2:24:13; a Kenyan). For the men, all 5 of the top placers were East African, whereas for the women all 3 were East African. What explains Kenyan marathon success? Incredibly, from 1992 onward—with the exception of 2001 and 2018—East Africans have won the Boston Marathon. We know that athleticism is irreducible to biology, and while genes do play a part in morphology and other things that are conducive to running success, they do not—of course—tell the whole story. A whole slew of factors needs to come together to make an elite athlete, while one thing does not fully explain marathon success.
Back in September of 2017, I covered many factors that make both elite marathoners and sprinters. All of the factors that make an elite athlete combine, no one factor is more important than any other, but if one does not have the will to train and win, of course, they will not do well.
When it comes to Kenyans, a small tribe in Kenya explain the success of—the Kalenjin, most specifically, the Nandi sub-tribe and a complex interaction of genotype, phenotype, and socioeconomic factors explain their success (Tucker, Onywera, and Santos-Concejero, 2015). The Kalenjin account for a whopping 84% of Kenya’s Olympic and world championship medals, 79 percent of Kenya’ top 25 marathon performances, contributing to 34%. Kenyans have won 152 medals, compared with 145 with other African countries—42-61% being Ethiopian—while the rest of the world combined won 153 medals. The Nandi sub-tribe has won 72 medals, accounting for 47& of the total for Kenya. What accounts for the insane disparity between East African marathoners (specifically Kalenjin, and a more specific sub-tribe at that) and the rest?
In his book The Genius in All of Us, David Shenk (2010: 102) writes:
Take the running Kenyans. Relatively new to the international competitions, Kenyans have in recent years become overwhelmingly dominant in middle- and long-distance races. “It’s pointless for me to run on the pro-circuit,” complained American 10,000 meter champion Mike Mykytok to the New York Times in 1998. “With all of the Kenyans, I could set a personal best time, and still only place 12th and win $200.”
The Kenyan-born journalist John Manners describes a just-so story to explain how and why Kenyans dominate these competitions: The best young men who were the fastest and had more endurance acquired more cattle, and those who acquired more cattle could then get a bride and have more children, Shenk explains. “It is not hard to imagine that such a reproductive advantage might cause a significant shift in a group’s genetic makeup over the course of a few centuries” (John Manners, quoted in Shenk, 2010: 103).
However, no matter what the origin of Kenyan running success is, the Kalenjin have a passionate dedication to running. Kipchoge Keino was the one who put Kenya on the map regarding distance running. Shenk quotes Keino saying:
I used to run from the farm to school and back … we didn’t have a water tap in the house, so you run to the river, take your shower, run home, change [run] to school . . . Everything is running.
However, when Keino entered 1968 Olympics in Mexico, he came down with gallstones and his doctor told him not to race. However, he took a cab to Aztec Stadium, and when he get caught in traffic he ran the last mile to the stadium and barely got there before the race started. Even though Keino was sick, he destroyed the then-world record by 6 seconds.
Sports geographers don’t point to one variable that explains Kenyan running success—because they all interact. They train at high altitude—and while high altitude is not the only factor regarding long-distance running success, it is crucial. Because training at a high altitude and then running at a lower altitude can change running time by a large amount. One with a normal running economy who goes by the mantra “live high, train low” can shave off about 8 minutes of their time in a 26.2-mile marathon (Chapman and Levine, 2007). Further, socioeconomic variables also explain the success, with it being part of what drives them to succeed, along with favorable morphology, a strong running economy, high intensity training (living at and training at high altitude) and a slew of psychological factors related to social status and socioeconomic factors (Wilbur and Pitsiladis, 2012). This paper speaks perfectly to the slew of variables that need to come together to make an elite athlete.
Shenk (2010: 108) then reverses John Manner’s just-so story:
… it’s an entertaining theory that fits well with the popular gene-centric view of natural selection [it fits well because it’s selected to be so]. But developmental biologists would point out that you could take exactly the same story line and flip the conclusion on its head: the fastest man earns the most wives and has the most kids—but rather than passing on quickness genes, he passes on crucial ingredients. such as the knowledge and means to attain maximal nutrition, inspiring stories, the most propitious attitude and beliefs, access to the best trainers, the most leisure time to pursue training, and so on. This nongenetic aspect of inheritance is often overlooked by genetic determinists: culture, knowledge, attitudes, and environments are also passed on in many different ways.
Further, Shenk also cites sports scientist Tim Noakes who states that the best Kenyan runners cover 230 km (about 143 miles) a week at 6,000 feet in altitude—and this, of course, would be conducive to running success when the event is held at lower altitudes.
David Epstein wrote a solid book on athleticism in 2014—The Sports Gene. Chapters 12 and 13 are pivotal for this discussion. Chapter 12 titled Can Every Kalenjin Run? In this chapter, Epstein, too, cites John Manners, explaining the same thing that Shenk did, but adds this:
In the next breath of the very same chapter [after describing the just-so story about cattle-gathering and wife-acquisiton], though, Manners seems to doubt the suggestion as soon as he raises it. “The idea just occurred to me, so I just put it in.” (pg 184)
Manners came to see his just-so story as less powerful since, over the years as he interviewed Kalenjin runners because “other “hot spots” of endurance running talent have materialized in East Africa, and the athletes responsible are also from traditionally pastoralist cultures that once practiced cattle raiding” (Epstein, 2014: 184-185).
Epstein then discusses how 17 American men in history have run a marathon better than 2:10—or 4:58 per mile—while 32 Kalenjin men did it in October of 2011 alone. Five American high-schoolers have run a sub-4 minute mile, while one high-school in Kenya alone produced 4 sub-4 mile runners!
Kenyan runners have long legs for their height, along with “upper leg length, total leg length and total leg length to body height ratio were correlated with running performance” (Mooses et al, 2014)—which means that they can cover more distance than one with shorter legs. This is critical for running success—of any kind. Kenyans have a high number of type I muscle fibers, but, of course, this alone does not explain their running success. Elite Kenyan distance runners are characterized by low BMI, low fat mass and slim limbs (Kong and de Heer, 2008).
So now let’s discuss altitude adaptation. One objection to this variable—out of many others, of course—that are conducive to running success is why are Tibetans and Andeas not succeeding in these types of competitions as well as the Kalenjin? The answer is simple—because they do not have the long, ecto-dominant (Vernillo et al, 2013) body types. There is also another, perhaps more critical, component to altitude training—hemoglobin, since the amount of oxygen one has in their blood is dictated by two factors—how much hemoglobin one has in their blood and the amount of oxygen the hemoglobin carries. Altitude increases the number of red blood cells in the body, since it is a good way to get oxygen in an environment with less oxygen.
Epstein (2014: 208) writes:
Preferable to moving to altitude to rain is being born there. Altitude natives who are born and go through dilchood at elevation tend to have proportionally larger lungs than sea-level natives, and large lungs have large surface ares that permit more oxygen to pass from the lungs into the blood. This cannot be the result of altitude ancestry that has altered the genes over generations, because it occurs not only in natives of the Himalayas, but also among American children who do not have altitude ancestry but who grow up in the Rockies. Once childhood is gone, though, so too is the chance for this adaptation. It is not genetic, but neither is it alterable after adolescence.
Epstein (2014: 213) quotes the first man to run a sub-4 minute mile, Roger Bannister who says:
The human body is centuries in advance of the physiologist, and can perform an integration of heart, lungs, and muscles which is too complex for the sciencist to analyze.
This, of course, is a hard pill to swallow for some people, who may not believe this. I believe this is true—though we can point to certain factors, each individual’s trajectory into X is unique, and so, explaining Y for all will be close to impossible.
Finally, Epstein (2014: 214) cites Claudio Berardelli:
Berardelli believes that Kenyans are, in general, more likely to be gifted runners. But he also knows that no matter their talent or body type or childhood environment or country of origin, 2:05 marathon runners do not fall from the sky. Their gifts must be coupled with herculean will.
Although that, too, is not entirely seperable from innate [whatever that means] talent.
Hamilton (2000) concludes that:
It seems that the presumed causes of such domination are often recycled, out of date, and based on misinformation and myth.
This, however, betrays understanding of a systems view of running success. Just because North Africans are beginning to show up in these types of competitions it does not mean that the systems view of athleticism is false.
Of course, the East African running advantage is more than ‘genetic’, it is also cultural—which, rightly, shows how every part of the system interacts to produce an elite athletic phenotype. As Louis (2014: 41) notes “The analysis and explanation of racial athleticism is therefore irreducible to biological or socio-cultural determinants and requires a ‘biocultural approach’ (Malina, 1988; Burfoot, 1999; Entine, 2000) or must account for environmental factors (Himes, 1988; Samson and Yerl`es, 1988).” Genetics alone cannot explain the running success of East Africans.
In sum, what explains the success of East African runners? A whole slew of factors that are irreducible, since the whole system interacts. Of course, I do not deny the role that physiological and anatomic factors have on running performance—they are crucial, but not the only, determinant for running success. Reducing a complex bio-system to X, Y, or Z does not make any sense, as every factor interacts to create the elite athlete. East African dominance in middle- and long-distance running will, of course, continue, since they have the right mix of factors that all interact with each other.
Snapchat Dysmorphia
1450 words
I’m not really one for social media (the only social media I use is Twitter and this blog) and so I don’t keep up with the new types of social media that continuously pop up. Snapchat has been around since 2011. It’s a type of social media where users can share pictures before they are then unavailable to the user they sent it to. I don’t understand the utility of media like this but maybe that’s because I’m not the target demographic.
In any case, I’m not going to talk about Snapchat in that way today, because that’s not what this blog is about. What I will talk about today, though, is the rise of “Snapchat dysmorphia.” “Dysmorphia” is defined by the Merriam Webster Dictionary as “characterized by malformation.” “Dysphoria”, according to the Merriam Webster Dictionary, is defined as “a state of feeling very unhappy, uneasy, or dissatisfied.” This is a type of body dysmorphia. The two terms—dysphoria and dysmorphia—are similar, in any case.
So where does Snapchat come into play here? Well, there are certain functions that one can do with their pictures—and I’m sure they can do the same with other applications as well. There are what I would term “picture editors” which can change how a person looks. From changing the background you’re in, to changing your facial features, there are a wide range of things these kinds of filters can generate onto photographs/videos.
Well, of course, with the rise of social media and people constantly being glued to their phones day in and day out—along with pretty much living their entire lives on social media—people get sucked into the digital world they make for themselves. People constantly send pictures to others about what they’re doing on that day—they don’t have a chance to live in the moment because they’re always trying to get the “best picture” of the moment they’re in, since they’re trying to get the best picture for their followers on social media. In any case, this is where the problem with these kinds of filters come in—and how Snapchat is driving these problems.
So people use these filters on their pictures. They then get used to seeing themselves as they see themselves in the filtered pictures. Since they spend so much time on social media, constantly filtering their pictures, they—and their social media followers—get used to seeing their filtered photos and not how they really like. This, right here, is the problem.
People then become dysphoric—they become unsatisfied with their appearance due to how they look in their filtered photos. This has lead numerous people to do what I believe is insane—they go and get plastic surgery to look like their Snapchat selves. This is, in part, what the meteoric rise of social media has done to the minds of the youth. They give them unrealistic expectations—through their filters—and then, since they spend so much time Snapchatting and whatever else they do, seeing their filtered pictures, they then get sad that they do not look like they do in their filtered pictures in their digital world, which causes them to become dysphoric about their facial features since they do not look like their Snapchat selves.
One Snapchat user said to Vice:
We’d rather have a digitally obscured version of ourselves than our actual selves out there. It’s honestly sad, but it’s a bitter reality. I try to avoid using them as much as I can because they seriously cause an unhealthy dysphoria.
Therein lies the problem: people become used to what I would say are “idealized versions” of themselves. Some of these filters completely change how one’s facial structure is; some of them give bigger or smaller eyes; others change the shape of the jawline and cheekbones; others give fuller lips. So now, instead of people bringing photographs of celebrities to plastic surgeons and saying to them “This is what I want to look like”, they’re bringing their edited Snapchat pictures to plastic surgeons and telling them that they want that look.
So it’s no wonder that people become dysphoric about their facial features when they pretty much live on social media. They constantly play around with this filter and that filter, and they become used to what then becomes an idealized version of themselves. These types of picture filters have been argued to be bad for self-esteem, and it’s no wonder why they are, given the types of things these filters can do to radically change the appearances of the users who use them.
There has been a rise in individuals bringing in their filtered photos to plastic surgeons, telling them that they want to look like the filtered picture. Indeed, some of the before and afters I have seen bear striking similarities to the filtered photo.
The term “Snapchat dysmorphia” has even made it into the journal JAMA in an article titled Selfies—living in the era of filtered photographs (Rajanala, Maymobe, and Vashi, 2018). They write that:
Previously, patients would bring images of celebrities to their consultations to emulate their attractive features. A new phenomenon, dubbed “Snapchat dysmorphia,” has patients seeking out cosmetic surgery to look like filtered versions of themselves instead, with fuller lips, bigger eyes, or a thinner nose.
Ramphus and Mejias (2018) state that while it may be too early to employ the term “Snapchat dysmorphia”, it is imperative to realize the reasons why many young people are thinking and getting plastic surgery. Indeed, a few plastic surgeons have stated that the type of alterations that patients describe to them, indeed, are what are found with Snapchat facial edits.
Ramphul and Mejias (2018) also write:
There are already some ongoing legal issues about the use of Snapchat in the operating room by some plastic surgeons but none currently involving any patients accusing Snapchat of giving them a false perception of themselves yet. The proper code of ethics among plastic surgeons should be respected and an early detection of associated symptoms in such patients might help provide them with the appropriate counseling and help they need.
Clearly, this issue is now becoming large enough that medical journals are now employing the term in their articles.
McLean et al’s (2015) results “showed that girls who regularly shared self-images on social media, relative to those who did not, reported significantly higher overvaluation of shape and weight, body dissatisfaction, dietary restraint, and internalization of the thin ideal. In addition, among girls who shared photos of themselves on social media, higher engagement in manipulation of and investment in these photos, but not higher media exposure, were associated with greater body-related and eating concerns, including after accounting for media use and internalization of the thin ideal.” This seems intuitive: the more time one spends on social media, sharing images, overvalues certain things. And putting this into context in regard to Snapchat dysmorphia, girls spending too much time on these types of applications that can change their appearance, may also develop eating disorders.
Ward et al (2018) report that in 2014, about 93 million selfies were taken per day. With the way selfies are taken—up close—this then distorts the nasal dimensions, increasing them (Ward et al, 2018). Although this is only tangentially related to the issue of Snapchat dysmorphia, it will also increase the chance of people seeking plastic surgery, since a lot of people spend so much time on social media, taking selfies and eventually idealizing their selves with the angles they take the pictures in.
Although there are only 2 pages on Google scholar when you search “Snapchat dysmorphia”, we can expect the number of journal articles and references to the term to increase in the coming years due to people basically living most of their lives on social media. This is troubling: that young people are spending so much time on social media, editing their photos and acquiring dysmorphia due to the types of edits that are possible with these applications is an issue that we will need to soon address. Quite obviously, getting plastic surgery to look more like the idealized Snapchat photo is not the solution to the problem—something more like counseling or therapy would seem to address the issue. Not pretty much telling people “If you have the money and the time to get this surgery done then you should, to look like how you idealize yourself.”
Should people get plastic surgery to fix their selves, or should they get counseling? People who look to, or get, survey to fix dysmorphic issues they have with themselves will never be satisfied. They will always see a blemish, an imperfection to fix. For this reason, getting surgery in an attempt “fix” yourself if addicted to your looks while using these picture filters won’t work, as the deeper problem isn’t addressed—which I would claim is rampant social media use.
Vegans/Vegetarians vs. Carnivores and the Neanderthal Diet
2050 words
The vegan/vegetarian-carnivore debate is one that is a false dichotomy. Of course, the middle ground is eating both plants and animals. I, personally, eat more meat (as I eat a high protein diet) than plants, but the plants are good for a palate-switch-up and getting other nutrients in my diet. In any case, on Twitter, I see that there is a debate between “carnivores” and “vegans/vegetarians” on which diet is healthier. I think the “carnivore” diet is healthier, though there is no evolutionary basis for the claims that they espouse. (Because we did evolve from plant-eaters.) In this article, I will discuss the best argument for ethical vegetarianism and the evolutionary basis for meat-eating.
Veganism/Vegetarianism
The ethical vegetarian argument is simple: Humans and non-human animals deserve the same moral consideration. Since they deserve the same moral consideration and we would not house humans for food, it then follows that we should not house non-human animals for food. The best argument for ethical vegetarianism comes from Peter Singer from Unsanctifying Animal Life. Singer’s argument also can be extended to using non-human animals for entertainment, research, and companionship.
Any being that can suffer has an interest in avoiding suffering. So the equal consideration of interests principle (Guidi, 2008) asserts that the ability to suffer applies to both human and non-human animals.
Here is Singer’s argument, from Just the Arguments: 100 of the Most Important Arguments in Western Philosophy (pg. 277-278):
P1. If a being can suffer, then that being’s interests merit moral consideration.
P2. If a being cannot suffer, then that beings interests do not merit moral consideration.
C1. If a being’s interests merit moral consideration, then that being can suffer (transposition, P2).
C2. A being’s interests merit moral consideration if and only if that being can suffer (material equivalence, P1, C1).
P3. The same interests merit the same moral consideration, regardless of what kind of being is the interest-bearer (equal consideration of interests principle).
P4. If one causes a being to suffer without adequate justification, then one violates that being’s interests.
P5. If one violates a being’s interests, then one does what is morally wrong.
C3. If one causes a being to suffer without adequate justification, then one does what is morally wrong (hypothetical syllogism, P4, P5).
P6. If P3, then if one kills, confines, or causes nonhuman animals to experience pain in order to use them as food, then one causes them to suffer without adequate justification.
P7. If one eats meat, then one participates in killin, confining, and causing nonhuman animals to experience pain in order to use them as food.
C4. If one eats mea, then one causes nonhuman animals to suffer without adequate justification (hypothetical syllogism, P6, P7).
C5. If one eats meat, the one does what is morally wrong (hypothetical syllogism, C3, C4).
This argument is pretty strong, indeed it is sound. However, I personally will never eat a vegetarian/vegan diet because I love eating meat too much. (Steak, turkey, chicken.) I will do what is morally wrong because I love the taste of meat.
In an evolutionary context, the animals we evolved from were plant-eaters. The amount of meat in our diets grew as we diverged from our non-human ancestors; we added meat through the ages as our tool-kit became more complex. Since the animals we evolved from were plant-eaters and we added meat as time went on, then, clearly, we were not “one or the other” in regard to diet—our diet constantly changed as we migrated into new biomes.
So although Singer’s argument is sound, I will never become a vegan/vegetarian. Fatty meat tastes too good.
Nathan Cofnas (2018) argues that “we cannot say decisively that vegetarianism or veganism is safe for children.” This is because even if the vitamins and minerals not gotten through the diet are supplemented, the bioavailability of the consumed nutrients are lower (Pressman, Clement, and Hayes, 2017). Furthermore, pregnant women should not eat a vegan/vegetarian diet since vegetarian diets can lead to B12 and iron deficiency along with low birth weight and vegan diets can lead to DHZ, zinc, and iron deficiencies along with a higher risk of pre-eclampsia and inadequate fetal brain development (Danielewicz et al, 2017). (See also Tan, Zhao, and Wang, 2019.)
Carnivory
Meat was important to our evolution, this cannot be denied. However, prominent “carnivores” take this fact and push it further than it goes. Yes, there is data that meat-eating allowed our brains to grow bigger, trading-off with body size. Fonseca-Azevedo and Herculano-Houzel (2012) showed that metabolic limitations resulting from hours of feeding and low caloric yield explain the body/brain size in great apes. Plant foods are low in kcal; great apes have large bodies and so, need to eat a lot of plants. They spend about 10 to 11 hours per day feeding. On the other hand, our brains started increasing in size with the appearance of erectus.
If erectus ate nothing but raw foods, he would have had to eat more than 8 hours per day while hominids with neurons around our level (about 86 billion; Herculano-Houzel, 2009). Thus, due to the extreme difficulty of attaining the amount of kcal needed to power the brains with more neurons, it is very unlikely that erectus would have been able to survive on only plant foods while eating 8+ hours per day. Indeed, with the archaeological evidence we have about erectus, it is patently ridiculous to claim that erectus did eat for that long. Great apes mostly graze all day. Since they graze all day—indeed, they need to as the caloric availability of raw foods is lower than in cooked foods (even cooked plant foods would have a higher bioavailability of nutrients)—then to afford their large bodies they need to basically do nothing but eat all day.
It makes no sense for erectus—and our immediate Homo sapiens ancestors—to eat nothing but raw plant foods for what amounts to more than a work day in the modern world. If this were the case, where would they have found the time to do everything else that we have learned about them in the archaeological record?
There is genetic evidence for human adaptation to a cooked diet (Carmody et al, 2016). Cooking food denatures the protein in it, making it easier to digest. Denaturation is the alteration of the protein shape of whatever is being cooked. Take the same kind of food. That food will have different nutrient bioavailability depending on whether or not it is cooked. This difference, Herculano-Houzel (2016) and Wrangham (2009) argue is what drove the evolution of our genus and our big brains.
Just because meat-eating and cooking was what drove the evolution of our big brains—or even only allowed our brains to grow bigger past a certain point—does not mean that we are “carnivores”; though it does throw a wrench into the idea that we—as in our species Homo sapiens—were strictly plant-eaters. Our ancestors ate a wide-range of foods depending on the biome they migrated to.
The fact that our brain takes up around 20 percent of our TDEE while representing only 2 percent of our overall body mass, the reason being our 86 billion neurons (Herculano-Houzel, 2011). So, clearly, as our brains grew bigger and acquired more neurons, there had to have been a way for our ancestors to acquire the energy need to power their brains and neurons and, as Fonseca-Azevedo and Herculano-Houzel (2012) show, it was not possible on only a plant diet. Eating and cooking meat was the impetus for brain growth and keeping the size of our brains.
Take this thought experiment. An asteroid smashes into the earth. A huge dust cloud blocks out the sun. So the asteroid would have been a cause of lowering food production. This halting of food production—high-quality foods—persisted for hundreds of years. What would happen to our bodies and brains? They would, of course, shrink depending on how much and what we eat. Food scarcity and availability, of course, do influence the brain and body size of primates (Montgomery et al, 2010), and humans would be no different. So, in this scenario I have concocted, in such an event, we would shrink, in both brain and body size. I would imagine in such a scenario that high-quality foods would disappear or become extremely hard to come by. This would further buttress the hypothesis that a shift to higher-quality energy is how and why our large brains evolved.
Neanderthal Diet
A new analysis of the tooth of a Neanderthal apparently establishes that they were mostly carnivorous, living mostly on horse and reindeer meat (Jaouen et al, 2019). Neanderthals did indeed have a high-meat diet in northerly latitudes during the cold season. Neanderthals in Southern Europe—especially during the warmer seasons—however, ate a mixture of plants and animals (Fiorenza et al, 2008). Further, there was a considerable plant component to the diet of Neanderthals (Perez-Perez et al, 2003) (with the existence of plant-rich diets for Neanderthals being seen mostly in the Near East; Henry, Brooks, and Piperno, 2011) while the diet of both Neanderthals and Homo sapiens varied due to climatic fluctuations (El Zataari et al, 2016). From what we know about modern human biochemistry and digestion, we can further make the claim that Neanderthals ate a good amount of plants.
Ulijaszek, Mann, and Elton (2013: 96) write:
‘Absence of evidence’ does not equate to ‘evidence of absence,’ and the meat-eating signals from numerous types of data probably swamp the plant-eating signlas for Neanderthals. Their dietary variability across space and time is consistent with the pattern observed in the hominin clade as a whole, and illustrates hominin dietary adaptatbility. It also mirrors trends observed in modern foragers, whereby those populations that live in less productive environments have a greater (albeit generally not exclusive) dependance on meat. Differences in Neanderthal and modern human diet may have resulted from exploitation of different environments: within Europe and Asia, it has been argued that modern humans exploited marginal areas, such as steppe environments, whereas Neanderthals may have preferred more mosaic, Mediterranean-type habitats.
Quite clearly, one cannot point to any one study to support an (ideologically driven) belief that our genus or Neanderthals were “strictly carnivore”, as there was great variability in the Neanderthal diet, as I have shown.
Conclusion
Singer’s argument for ethical vegetarianism is sound; I personally can find no fault in it (if anyone can, leave a comment and we can discuss it, I will take Singer’s side). Although I can find no fault in the argument, I would never become a vegan/vegetarian as I love meat too much. There is evidence that vegan/vegetarian diets are not good for growing children and pregnant mothers, and although the same can be said for any type of diet that leads to nutrient deficiencies, the risk is much higher in these types of plant-based diets.
The evidence that we were meat-eaters in our evolutionary history is there, but we evolved as eclectic feeders. There was great variability in the Neanderthal diet depending on where they lived, and so the claim that they were “full-on carnivore” is false. The literature attests to great dietary flexibility and variability in both Homo sapiens and Neanderthals, so the claim that they ate meat and only meat is false.
My conclusion in my look into our diet over evolutionary time was:
It is clear that both claims from vegans/vegetarians and carnivores are false: there is no one “human diet” that we “should” be eating. Individual variation in different physiologic processes implies that there is no one “human diet”, no matter what type of food is being pushed as “what we should be” eating. Humans are eclectic feeders; we will eat anything since “Humans show remarkable dietary flexibility and adaptability“. Furthermore, we also “have a relatively unspecialized gut, with a colon that is shorter relative to overall size than in other apes; this is often attributed to the greater reliance on faunivory in humans (Chivers and Langer 1994)” (Ulijaszek, Mann, and Elton, 2013: 58). Our dietary eclectism can be traced back to our Australopithecine ancestors. The claim that we are either “vegetarian/vegan or carnivore” throughout our evolution is false.
There is no evidence for both of these claims from both of these extreme camps; humans are eclectic feeders. We are omnivorous, not vegan/vegetarian or carnivores. Although we did evolve from plant-eating primates and then added meat into our diets over time, there is no evidence for the claim that we ate only meat. Our dietary flexibility attests to that.
Race and Menarche
1100 words
Back in 2016 I wrote about racial differences in menarche and how there is good evidence that leptin is a strong candidate for the cause in my article Leptin and its Role in the Sexual Maturity of Black Girls (disregard the just-so stories). Black girls are more likely to hit puberty at a younger age than white girls. Why? One reason may be that leptin may play a role in the accelerated growth and maturation of black girls, since there was a positive relationship between leptin concentration and obesity in black girls (Wagner and Heyward, 2000). When girls start to develop at younger and younger ages, a phrase you hear often is “It’s the chemicals in the food” in regard to, for example, early breast development on a young, pre-teen girl.
Black girls are more likely to be obese than white girls (Freedman et al, 2000) and it is thought that body fat permits the effects of earlier menarche due to leptin being released from the adipocyte (fat cell) (Salsberry, Reagen, and Pajer, 2010). Freedman et al (2000) showed that black girls experienced menarche 3 months earlier than white girls on average, while over a 20 year period the median age decreased by 9.5 months. There is also evidence of earlier thelarche (breast development) in black girls, which was mediated by gonadotropin (Cabrera et al, 2014). Wong et al (1998) found that circulating serum leptin levels were correlated with earlier menarche in black girls which was related to body fatness and age but lessened after fat mass, maturation and physical fitness. There is a ton of evidence that exists that body fatness is related to obesity and, as I said above, the mechanism is probably fat cells releasing leptin, permitting earlier menarche (see Kaplowitz, 2008). Higher levels of body fat cause earlier menarche; earlier menarche does not cause higher levels of body fat. The evidence is there that leptin indeed plays the permissive role to allow a girl to enter into puberty earlier, and that this is how and why black girls enter menarche earlier than white girls.
So when fat mass increases, so does leptin; when leptin increases, girls have puberty at an earlier age (Apter, 2009). Black girls have higher levels of circulating leptin than white girls (Ambrosious et al, 1998). So knowing the relationship between leptin and obesity and how fat cells release leptin into the body permissing earlier puberty, we can confidently say that leptin is a major cause of earlier pubertal development in black girls. Total body fat correlates with fasted leptin (Ebenibo et al, 2018).
Siervogel et al (2003) write:
A negative relationship between age at menarche, BMI and body fatness in girls has been shown [63, 64, 65]. It is still unclear, however, whether increased early childhood adiposity induces an earlier onset of puberty, if rapid maturation and early puberty induce an increase in body fat later in life, or whether both of these phenomena occur.
The average age of menarche in black girls was 12 years of age whereas for white girls it was 12.5 (Regan et al, 2013). Since we now know the causes of earlier menarche, we can talk about ultimate causation.
Since menarche is correlated with obesity and circulating leptin levels, then food quality would be an easy culprit to look at.
Deardorff et al (2014) write:
Three sets of findings stood out. One, grandmother’s lower education was related to later menarche for Black girls. Two, mother’s unmarried status (at birth and age 7) was associated with earlier menarche for Hispanics and Whites, but not for Blacks. Three, family income at child’s birth was related to earlier menarche for Blacks and Hispanics, but not Whites.
Lower family income at birth was associated with earlier menarche for Blacks (and to some extent for Hispanics), but not for white girls.
Deardorff et al’s (2014) study suggests that social determinants of health can be intervened upon and differences in pubertal timing can be ameliorated since they are driven by social factors.
Black children have higher levels of insulino-glucose ratios than white children, even after adjusting for confounds (Wong et al, 1999). There were similar findings when comparing normal-weight black and white girls matched for age, bone age, weight, and BMI. Black girls also had higher levels of insulin than white girls. Black girls grow faster than white girls beginning at 2 years of age, with the cause hypothesized to be “higher serum insulin concentrations in healthy African American girls suppress the hepatic production of IGFBP-1, which results in higher circulating concentrations of free IGF-I, contributing to the accelerated growth observed in African American girls compared with their white counterparts” (Wong et al, 1999: 297).
Wong et al (1999) found that black girls were older and more sexually mature than white girls according to the Tanner stages of development (using pubic hair as a measure). Even after adjusting for age, black girls were still more sexually mature, heavier, and had higher levels of fat-free mass. Both serum insulin levels and serum concentrations of IGF-1 were significantly higher in black girls compared to white girls. Even after they controlled for differences in pubic hair development and fat mass, the differences in IGF-1 and IGFBP-1 remained significant. So black girls in this study had higher levels of IGF-1 and insulin than white girls.
This is significant because higher levels of IGF-1 at age 8 are associated with earlier menarche in girls (Thankamoney et al, 2012). Earlier menarche is related to higher levels of adiposity, as noted above, and the mechanism for earlier menarche is fat cells releasing insulin which has a permissive effect on menarche. Further, there is evidence that lower levels of circulating growth hormones are related to delayed menarche (Circo, 2014).
IGF-1 is a protein “coded for” by the IGF-1 gene (Hoppener et al, 1985). IGF-1 is molecularly similar to insulin (Laron, 2001). This makes sense. Body fat cannot be stored unless insulin is high; since they are molecularly similar, they both cause growth, albeit in different ways.
In conclusion, there is strong evidence that body fatness is related to earlier menarche, with the mechanism being fat cells releasing leptin which has a permissive effect on pubertal development. Further evidence indicates that circulating hormones like insulin and IGF-1 also have permissive effects on pubertal development. Black girls are more likely to be obese than white girls. Black girls are more likely to have higher levels of circulating hormones like IGF-1 and insulin than white girls. Thus, a cause—a major cause in my opinion—for earlier menarche in black girls compared to white girls is higher levels of body fat and higher levels of circulating hormones like IGF-1 and insulin which have a permissive effect on menarche.
Racial Differences in the Palmaris Longus
1050 words
There are many physical differences between racial/ethnic groups. Some of these differences are obvious to the naked eye, others much less so. One racial difference that exists is variation in having a tendon called the palmaris longus (PL). This muscle rests between the flexor carpi radialis and the flexor carpi urinalis. The radius and ulna are bones in the forearm. The radius supports the lateral (thumb) side of the hand whereas the ulna supports the medial (pinky) side; they enable wrist rotation. To see if you have the tendon, take your pinky and touch it to your thumb. If you have the tendon you should see it poking out of your wrist. Fourteen percent of the population lacks the PL, but there is considerable variation by race. These differences, of course, have should be taken into account when doing a tendon graph operation.
The action that the PL performs is flexing the wrist; the origin is the medial epicondyle of the humerus; it inserts in the palmar aponeurosis and flexor rentinaculum of the hand. The antagonist muscles are extensor carpi radialis brevis, extensor carpi radialis longus, and extensor carpi urinalis.
There are unilateral (affecting one side of the body) differences in the variation of this tendon along with bilateral (affecting both sides of the body) differences. There is differential absence of the tendon depending on which hand is dominant (Eric et al, 2011). The tendon also has been found to contribute to the strength of thumb abuction (Gangata, Ndou, and Louw, 2013). However, it has been shown that whether or not one has the PL or not does not contribute to grip/pinch strength (Sebastin et al, 2005).
Soltani et al (2012) followed patients at hand surgery clinics LA county +University of Southern California Medical Center and Keck Medical Center of the University of Southern California. Their objective was to observe the variation in the PL in regard to race, sex and ethnicity (indeed, they placed people into races based on the US Census designations; see Spencer, 2014, they excluded mixed-race people from the ethnicity part of the analysis). They wanted to see the extent that the PL was missing and whether or not it was bilateral or unilateral. They evaluated a group of 516 multi-ethnic individuals while age, race, ethnicity and sex was accounted for. They then administered the Schaeffer test:
![]()
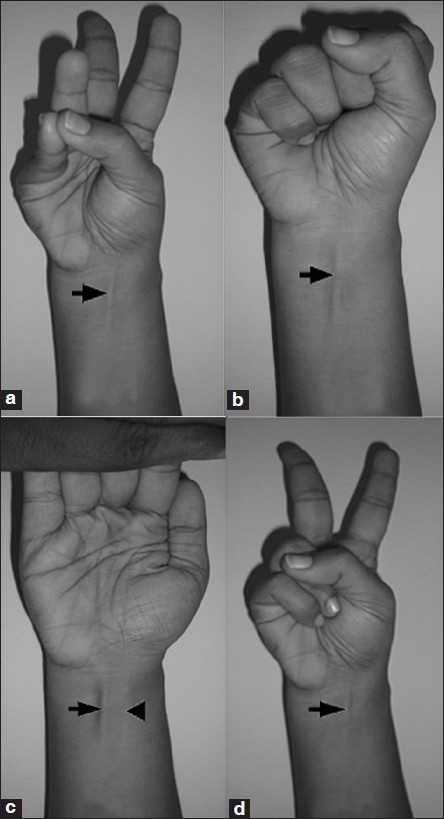
Obviously, this only works with lean individuals. So obese patients needed to get an ultrasound to ascertain whether or not they had the tendon.
In their sample of 516, 415 were Caucasian, 55 were African American, 35 were Asian and 11 were mixed race. The age range was 12-94, while an even number of men and women were tested (288, 288). Soltani et al (2012) write:
There were no differences in the absence of the PL based on laterality. The right side was absent in 11.8% and left 12.0% of the time (see Table 1). Further, there were no differences in the absence of the PL based on gender, value 0.369 (see Table 2). Ethnically, there was no difference in the absence of the PL between White (non-Hispanic) and White (Hispanic) patients, with prevalence of 14.9% and 13.1%, respectively. However, African American (4.5%) and Asian (2.9%) patients had significantly fewer absences of the PL than the Hispanic reference group.
They then write in the discussion that this has implications for plastic surgery—this anatomic variation between the races has implications for surgery:
This is information that should be taken into account preoperatively when planning surgical algorithms in treating tendon injuries or palsy. The PL is one such option as a tendon transfer for opponensplasty in restoring intrinsic function in cases of recurrent median nerve injury. If the PL is absent on the affected side, it is important to know preoperatively to plan using another donor muscle such as the extensor indicis proprius. In our study, the African American population had a statistically significantly lower rate of absent PL (4.5%), which is radically different than previously published reports from Nigeria, where the absence rates were much higher (31%). This could be due to the ethnic heterogeneity of the African American population of the United States compared to the Nigerian population. Nevertheless, the PL is present in high likelihood in this particular ethnic group which bodes well for using the PL in a surgical scenario. The PL is used quite frequently in cases of secondary tendon reconstruction, and it is useful for the surgeon to be aware of that issue preoperatively for surgical safety and efficiency in harvesting the tendon graft. The patient needs to be aware of the location of possible surgical incisions for tendon harvesting. Further, the surgeon should examine all possible tendon donors preoperatively, and one’s suspicion might be heightened by knowing the patient’s ethnicity. This is particularly important for the White population which in our study had the highest rates of absence, in both the Hispanic and non-Hispanic subsets. The surgeon must be aware in these patients that it is more likely that the PL might be absent. Thus, in Caucasian patients, it is particularly important to have a thorough examination of possible tendon donor sites.
Touching on the point of there being no difference between having or not having a PL and grip/pinch strength: we should not expect that, since whites dominate strength competitions (and the Chinese powerlifters are no slouches either, look at some videos of them). So, just because whites have a higher prevalence of not having the tendon does not mean that they do not have a strong hand grip. Indeed, senior blacks did have a stronger hand grip than whites (Araujo et al, 2010), but the PL is not the cause of the slightly higher hand grip strength (this was seen in a Turkish study; Setin et al, 2013). However, tennis players need a strong grip and the PL is more likely to be found in elite tennis players over the recreational athletes (Vercruyssen, Scafoglieri, and Cattrysse, 2016). But Nekkanti et al (2018) conclude that “There was no statistically significant correlation between agenesis of PL and the mean hand grip. Right-handed dominant individuals had a higher incidence of PL.”
In sum, this is tendon varies by race/ethnicity/sex/handedness like many other traits. understanding these differences will lead to a better understanding of plastic surgery in regard to the tendon. There is a racial difference, but it is of no functional relevance in regard to hand grip/power/strength sports.
Is Diet and Exercise Eugenic?
1550 words
Eugenics can be defined as “the science of improving a human population by controlled breeding to increase the occurrence of desirable heritable characteristics.” Though, in his book Genetic Ethics: An Introduction, Farrelly (2018: 30) cites Bertrand Russel’s definition of eugenics. Eugenics is “the attempt to improve the biological character of a breed by deliberate methods adopted to that end.” So, eugenics does not have to be ‘bad’, if it is morally justifiable and defensible, if they treat all people as free and equal (which most eugenic movements in the past have not done. See my article Eugenics and the State for a history of eugenics and the policies that arose from it.)
Dieting (defined as “what one eats”, not the other commonly-used definition “when one is in caloric restriction under TDEE”) and exercise change the expression of genes in the genome. For example, mature skeletal muscle can adapt to numerous stressors—indeed, if it could not, then we would not be able to choose to gain (or lose) muscle mass. Exercise induces the activity of certain genes (Vissing, Anderson, and Schjerling, 2005). One study on endurance athletes showed that there are pronounced effects of gene expression on exercised and non-exercised muscles (Catoire et al, 2012). One study showed that the ingestion of glucose during exercise decreases the gene expression of genes associated with fatty acid metabolism (Civitarese et al, 2005). Numerous epigenetic changes are also induced by exercise (Ntanasis-Stathopoulos, Tzanninis, and Koutsilieris, 2013). Miyamoto-Mikami et al (2018) showed that young men who participated in high-intensity intermittent exercise training showed that 79 genes had an elevated expression whereas 73 genes were significantly reduced.
Williams and Neufer (1996) show that long-term responses in regard to adaptations in regard to a specific exercise “require changes in gene expression, mediated by changes in the rate of transcription of specific genes and in the rate of synthesis of
specific proteins.” Further, diet and exercise can change the transcriptional properties of skeletal muscle, which induce further physiological changes (Hargreaves and Cameron-Smith, 2002). There is even preliminary evidence that diet and exercise affects the epigenome over several generations (Barres and Zierath, 2016). Since changes occur to the epigenome due to environmental stressors, and exercise is an environmental stressor, it follows that exercise, too, can change the epigenome.
People visit the gym to change their biology. Since people visit the gym to change their biology, are they involved in “self-eugenics”? I would say yes, going with the definition from Russell quoted above. That one wants to change their biology means that they are most likely not currently happy with the way their biological phenotype currently is. So they visit the gym, begin a diet (defined here as “caloric restriction”) in order to change their biological phenotype since they are not happy with it. Methods like diet and exercise seek to improve the biological character of a breed through deliberate methods, and so, fall under the umbrella of “eugenics.”
But there is a difference between this type of “eugenics” and methods commonly thought of when “eugenics” are discussed. “When eugenic measures exemplify moral and epistemic virtue rather than vice, they are morally obligatory rather than simply morally permissible” (Farrelly, 2018: 42). Prescribing (a sensible) diet and exercise to a populace can and will improve their health; further, educating people on the right and wrong things to eat (“right and wrong things to eat” in regard to their current goals) and how these things we eat affect our physiology is not morally objectionable nor is it coercive.
Think of the eugenic policies I discussed in my article on eugenics and the State. The policies discussed (such as forced sterilization, infanticide, forced contraceptive measures, and selective breeding) are immoral: the State is attempting to force its ideals on the populace, and so it can be argued that it is immoral since individual autonomy is taken away (or attempted to be taken away). On the other hand, prescribing diet and exercise is not eugenic in this manner: it’s just a prescription, what one should do if they would like to live their life to the highest quality. This includes staying away from highly processed and refined foods (carbs) and other, “non-natural” foodstuffs. This is only a suggestion based on the current state of nutritional knowledge; if one wants to live the best-possible life then they should diet and exercise.
On the other hand, we can take a State-measure and, using the definition in this article from Russell, can say that this measure is eugenic, but its similarities to what is being argued here is irrelevant, since I am arguing for education, not forcing people to do something (though I will state my views on this matter at the end, which I still honestly think about since it conflicts with some of my views.)
Back in the beginning of this decade, then-mayor of NYC Michael Bloomberg “proposed [a] regulation that would bar food service establishments from selling certain sugary drinks in containers larger than sixteen ounces” in an effort to “reduce the city’s obesity rate.” When this law was proposed, I was all for it. People cannot make decisions for themselves, because when they do, they make the “wrong” (in regard to, what I would assume to be what people try to achieve—a healthy lifestyle) decisions, and so, I thought that a policy like this was a good idea, because who the hell needs a 64 oz. Big Gulp soda (which could have up to 700 kcal in the cup) from 7-11? Why would someone need to down almost 2 liters of soda in one go? Note that, I would assume, the individual would not be caught dead drinking out of a 2-liter bottle of soda (though I have seen quite a lot of people do so, even early in the morning). But there is no problem using the cup since its size is kind of deceptive—deceptive in the manner that it does not look like the 2-liter bottle of (family-sized) soda.
I loved this proposal when it was announced. It would, I thought, attempt to address one aspect of our obesity problem (since nearly 40 percent of all of the added sugars we consume come from sugary drinks). Back then, I was more libertarian in my politics, but I thought that the policy was a good idea, even though it conflicted with my views on politics. I now do not believe we should take these types of measures—I believe that education is sufficient, along with getting rid of food deserts which hamper the ability of those in those deserts to get access to good, high-quality food (which affects certain races over others; National Research Council, 2009).
The view held by Bloomberg, and now current NYC mayor Bill Deblasio, is an example of a policy that would take away one’s choice to drink what they would like. Bloomberg’s rationale was that, if people wanted more of the drink, then they can go over and refill their cup so that they can see what and how much they are actually drinking. This, on its face, is sensible. If one wants to drink the same amount they would have drank in, say, a 64 oz. Big Gulp, they can keep refilling their cup in order to get the same amount of liquid they would have gotten in the bigger cup. But, what if someone wants a 64 oz. Big Gulp? What about a 128 oz. Ultra Big Gulp? A 256 oz. Super Ultra Big Gulp? Is there anywhere we should draw this line? Should we?
In any case, I have shown that exercise and diet is “eugenic” in the sense of Russel’s definition. But it is not “eugenic” in the sense of, in my opinion, what most people mean when they discuss “eugenics”: taking away one’s individual autonomy to do what they want, forcing them to do something. (Though, they are not being told they cannot drink sugary drinks, they are being told that there is a size limit on how big their cup is; they would be forced to drink a sugary drink in a small cup.)
Bloomberg’s proposed measure is quite obviously eugenic since it “attempt[s] to improve the biological character of a breed by deliberate methods adopted to that end.” Is the health of the populace more important than individual autonomy to be able to buy their 256 oz. Super Ultra Big Gulp? Or is one’s ability to freely drink their 256 oz. Ultra Big Gulp more important? If it can be shown that this policy would reduce the number of obese people in the City, should it be attempted?
These are important (moral) questions to answer. I am honestly undecided here; this issue is incredibly complex. Though, we do know one thing: exercise and diet is “eugenic” in Russel’s sense, and the measures exemplify moral and epistemic virtue so it is not like “State-forced” eugenics of old. Whether or not there is a negative connotation to “eugenics” depends on whether or not it pursues sound and morally justifiable aims. Therefore, though prescribing diet and exercise is eugenic since they both influence the epigenome and gene expression (along with it falling under Russel’s definition of eugenics), it is not eugenic in a negative sense, since there are sound and morally justifiable aims in prescribing diet and exercise to the population at large.
Race as a Genuine Kind
1600 words
Race realism can be defined as the belief that our racial categories pick out real kinds in nature. Picking out real kinds in nature is a necessary condition for race realism to be true. I’ve extensively covered Hardimon’s (2017) and Spencer’s (2014) arguments on the existence of race. They definitively prove that, all though there is a social/cultural dimension to race, biological racial realism is true, since our racial categories truly do pick out real kinds in nature.
In the philosophy of race, questions such as “What is race?” and “Is race biological?” are asked. Philosophers of race use many tools at their disposal to attempt to answer these and other questions regarding the metaphysics of race. Four main views on race exist: racial anti-realism, the belief that race does not exist; race realism, the belief that race is real; social constructivism, the belief that race is a social construct; and biological racial realism, the belief that race is biologically real (Spencer, 2011).
Note that social constructivists about race are race realists; they believe that race is real but that it is strictly a social reality. Racial anti-realists take the belief that race does not exist at all; so those who make the claim that social constructivists about race say that race is not real, they really mean to say that racial anti-realists claim that race is not real. People hear the phrase “social construct” and automatically assume that that individual is attempting to argue that race is not real. But social constructivists about race do believe that race is real—that is, they are race realists, but only in a social, not biological, manner.
Spencer (2011: 9) argues that genuine kinds are kinds that contribute to long-term scientific progress, meaning that genuine kinds are “a valid kind in a well-ordered SRP [scientific research program].” Spencer (2011: 9) states that valid kinds in an SRP are “useful for playing an epistemic role of scientific kinds in that SRP, as well as a kind that is adequately epistemically justified in its SRP” while also stating that “ a valid kind in an SRP must be useful for playing an epistemic role of scientific kinds in its SRP.” Kinds are valid in SRPs when they can help scientists generate observations for understanding the natural world. Genuine kinds can be epistemically useful without being inductively useful.
Genuine kinds can be epistemically real or just a human (social) construct. Genuine kinds are real enough to use in scientific research—because they tell us just enough about the world to be epistemically useful in a SRP. That a kind is genuine means that it is “real enough” to be used in scientific research.
Now let’s look at Spencer’s argument that biological racial realism should mean “race is a genuine kind in biology”:
(1) The meaning of ‘biological racial realism’ in the race debate should be a metaphysically minimal interpretation of important scientific kindhood that also does the most justice to what counts as an important scientific kind.
(2) A “metaphysically minimal” interpretation of important scientific kindhood is one that does not adopt unnecessary and contentious metaphysical assumptions.
(3) The interpretation of important scientific kindhood that does the most justice to what counts as an important scientific kind is the one that best captures epistemically important scientific kinds—or ‘EIS kinds’ for short.
(4) The candidates for important scientific kindhood in the race debate are naturalo kinds, naturali kinds, naturalu kinds, naturalp kinds, realp biological kinds, reali biological kinds, and genuine kinds.
(5) No kind of kind in the race debate is both metaphysically minimal and does a better job of capturing EIS kindhood than genuine kinds.
(6) Therefore, the meaning of ‘biological racial realism’ in the race debate should be ‘race is a genuine kind in biology’.
This argument provided by Spencer establishes the fact that biological racial realism in the race debate should be ‘race is a genuine kind in biology.’ Now that I have laid out that an entity being biologically real in the race debate is not whether or not it is objective but whether it is epistemically justified in a biological research program, I will now turn to other author’s views on biological racial realism.
The topic on “Race as a Biological Kind” on PhilPapers states that:
One option is to say that one’s ancestor is a member of race X in virtue of sharing similar phenotypic, or observable, properties specific to other members of one’ s reproductively isolated breeding population. A second option is to say that one’s ancestor is a member of race X in virtue of sharing similar genotypic, or genetic, properties specific to other members of one’s reproductively isolated breeding population.
While “… ancestral relations among reproductively isolated breeding populations and either genotypic or phenotypic properties is one way to develop a naturalistic account of race.” Note how this is almost, to the tee, Hardimon’s (2017) populationist race concept.
Andreason (2000) writes that “Most constructivists assume that biological realism and social constructivism are incompatible views about race; I argue that the two conceptions can be compatible.” Indeed, the claim that race is a social construct of a biological reality is a tempting view to take (and one I take to myself). That we socially construct groups does not mean that there is no biological reality to them, as Spencer (2014) shows with his Blumenbachian partitions. The claim that race is a social construct of a biological reality is completely at ends with the claim that race is purely social/political (as Dorothy Roberts argues) and the claim that race is purely biological. Though, we cannot separate ‘social/political’ and ‘biological’ terminology from our ontology. We must use both in conjunction in order to tease out what ‘race’ truly means. Stating that race is a social construct and only a social construct betrays the biology behind race. Stating that race is only biological betrays the social aspects of race. (Though, Hardimon (2017) has one race concept—the minimalist race concept—in which there is nothing ‘social’ about his proposed concept of race.)
Hardimon has four concepts of race: (1) the racialist concept of race—the claim that significant intellectual and moral differences exist between races (which he dispatched in his book); (2) the minimalist concept of race—the claim that groups that exhibit patterns of visible physical features which correspond to geographic ancestry satisfy the conditions of minimalist race; (3) the populationist race concept of race (the scientization of the minimalist race concept)—the claim that “race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population” (Hardimon, 2017: 99); and (4) the socialrace concept of race—the claim that race is a social, not biological, reality, and that society constructs what ‘race’ is.
Our racial categories pick out real kinds in nature, therefore race realism is true. This is established by both Hardimon (2017) and Spencer (2014), who discuss Rosenberg et al’s (2002) paper on the existence of population clusters deemed continental-level minimalist races by Hardimon (2017) and Blumenbachian partitions by Spencer (2014). Both arguments provided by Hardimon and Spencer are sound—they answer numerous objections that critics bring up to argue against Rosenberg et al’s (2002) study and other’s interpretation of what they say.
This article discusses Rosenberg et al (2002), and has this to say about the Kalash (the sixth cluster in that Rosenberg et al (2002) found):
The sixth population which is composed entirely of members of the Kalash, an isolated population in central Pakistan, is omitted by Wade on the grounds it “makes no genetic or geographic sense”. But the Kalash have a significant degree of genetic isolation from others, comparable in magnitude to that of other groups such as Native Americans. Moreover, they all live in the same place.
This is nonsense. That Structure labels a population as genetically distinct does not entail that that population is a continental-level minimalist race. That Structure picks out the Kalash as a genetically distinct group does not undercut Hardimon’s (2017) arguments on the existence of continental-level minimalist races. So, both K=6 and K=5 show that continental-level minimalist races are genetically structured (Hardimon, 2017: 88). (Further responses to critiques of Rosenberg et al can be found here and in Hardimon, 2017, chapter 5 and Spencer, 2014.)
Lastly, I’ll take some time to respond to commenter Oliver D. Smith’s objections to cranial measures and geographic ancestry.
Smith cited Sierp and Henneberg’s (2015) paper Can ancestry be consistently determined from the skeleton? in which the authors show that “no one individual was identified as belonging to only one ‘racial class’” (Sierp and Henneberg, 2015: 23). I don’t find this to be a problem. If we take Hardimon’s minimalist and populationist race concepts, the three conditions that need to be satisfied to delineate races are differences in phenotype which correspond to geographic ancestry, geographic location, and geographic ancestry. The critique from Sierp and Henneberg (2015) only counts against one parameter—if that—which are skulls. When delineating races, we don’t only look at skulls (one visible physical feature), we look at the whole suite of traits that make a “race” “racial.” Therefore, Sierp and Henneberg’s (2015) critique has no bearing on Hardimon’s (2017) or Spencer’s (2014) arguments.
Is race a genuine kind in biology? Do our racial categories pick out real kinds in nature? The answer to both questions is “yes.” Race is a genuine kind in biology since it captures EIS kindhood; our racial categories do pick out real kinds in nature, as shown by Spencer (2014) and his Blumenbachian partitions. Hardimon’s and Spencer’s arguments are definitive: biological racial realism is true.
(i) If biological racial realism is true, then our racial categories would have to pick out real kinds in nature.
(ii) Our racial categories pick out real kinds in nature.
(iii) Therefore, biological racial realism is true.
