
Home » Posts tagged 'Heritability'
Tag Archives: Heritability
Why Heritability Estimates are Flawed: A Conceptual Account
1200 words
Introduction
Heritability estimates have been used as a cornerstone and psychology and genetic research. They are designed to quantify the proportion of phenotypic variance in a population that can be attributed to genetic differences among individuals. We’ve known for a while now that heritability isn’t a measure of genetic strength (Moore and Shenk, 2016), but it’s a population-specific estimate of variance. Here I will provide two a priori arguments (one methodological on twins and the EEA and one theoretical based on Noble’s biological relativity argument). The twin critique shows that the twin researcher’s main assumption (equal environments) does not hold while the biological relativity critique shows that h2 is conceptually invalid. This is why there is a missing heritability problem—it never existed in the first place, and the assumptions twin researchers have are false.
The classical twin method
The CTM compares MZ and DZ twins to attempt to quantify the relative contributions of genes and environment in relation to the origin of trait differences between individuals. Perhaps the biggest assumption of the twin researcher is the equal environments assumption (EEA). The EEA assumes that MZ and DZ twins experience equivalent shared environments.
The EEA seems plausible enough: twins reared together should experience compatible environments, regardless of zygosity. But since DZ twins are more genetically similar than DZ twins so they’ll be more phenotypically similar as well. MZ twins are dressed alike, mistaken for one another, or placed in similar social roles compared to DZ twins which leads to more similar environments. So the shared environmental variance for MZ twins exceeds that for DZ twins, violating the EEA.
Clearly this violation throws a wrench into the logic of the CTM. The formula assumes that the greater similarity in MZ twins stems solely from their genetic identity. But if MZ twins experience more similar environments due to their phenotypic similarity (Fosse, Joseph and Richardson, 2015; Joseph et al, 2015), the difference in correlations between MZs and DZs captures genetic variance and excess environmental similarity. Thus, heritability is overestimated (see eg Bingley, Cappellari, and Tatsiramos, 2023) increasing the effect of genes while masking the effects of the environment—in effect, environment is made to look like genes. Thus, h2 fails to isolate genetic influence as intended. (Note that Grayson 1989 explains this as well, but it seems that it’s just ignored.) Here’s the argument:
(1) The classical twin method assumes that its heritability (h2) estimate (Falconer’s formula) isolates the proportion of phenotypic variance due solely to genetic variance.
(2) For the h2 estimate to isolate genetic variance, the shared environmental variance must be equal for MZ and DZ twins.
(3) MZ twins are more genetically similar than DZ twins.
(4) Genetic similarity between individuals leads to greater similarity in their expressed phenotypic traits, and this phenotypic similarity results in greater similarity in their environmental experiences.
(5) Because MZ twins have greater genetic similarity than DZ twins, and genetic similarity leads to phenotypic similarity, which in turn results in environmental similarity, the shared environmental variance is greater for MZ twins compared to DZ twins.
(6) If the shared environmental variance for MZ twins is greater than that for DZ twins, then the EEA is false because it requires that shared environmental variance be equal for both twin types.
(7) If the EEA is false, then we cannot logically infer genetic conclusions from h2, and thus h2 reflects shared environmental variance (c2), rather than genetic variance.
(8) Any method that relies on an assumption that’s logically inconsistent with the principles governing it’s variables – like the relationship between genetic similarity, environmental similarity and phenotypic similarity – cannot accurately isolate its intended causal component and is therefore conceptually untenable.
(9) Thus, the classical twin method is conceptually and logically untenable since it depends on the EEA which, when false, renders h2 a measure of environmental—not genetic—variance.
The biological relativity critique against h2
This argument is theoretical as opposed to methodological, and it relies on Noble’s (2012) biological relativity argument, where there is no privileged level of causation in biological systems. Genes, cells, tissues, organs, organisms, and the environment form an interdependent network where each level influences and is influenced by the other levels. Phenotypes arise from the interaction between all of these levels, not just due to the independent action of any one of the resources.
Heritability rests on a reductionist assumption—that phenotypic variance can be neatly partitioned into genetic and environmental components with genetic effects isolated as a distinct and quantifiable entity. This framework, clearly, privileges the genetic level treating it as separate from the broader biological and ecological context. But Noble’s argument directly contradicts this view. Genes don’t operate in a vacuum and do nothing on their own.
So by attempting to isolate genetic variance, heritability imposed an artificial simplicity on a complex reality (Rose, 2006). Noble’s principle suggests that separation isn’t just an approximation but a fundamental conceptual flaw. Phenotypic variation emerges from the integrated functioning of all biological levels, which then makes it impossible to assign causation to genes alone.
Thus, h2 is conceptually flawed, since it seeks to measure a genetic contribution that cannot be meaningfully disentangled from the holistic system in which it operates. Obviously the conceptual foundation of h2 contradicts the principle of biological relativity. Since h2 attempts to assign a specific portion of trait variance to genes alone, h2 implicitly privileges the genetic level, suggesting that it can be disentangled from the broader biological system. Noble’s argument denies that this is possible while emphasizing holism and rejecting reductionism. Thus, a priori, h2 estimates are fundamentally flawed because they rest on a reductionist framework which assumes a separability of causes which is incompatible with the holistic, relativistic nature of biological causation. Here’s the argument:
(1) Biological relativity holds that there is no privileged level of causation in biological systems: all levels (genes, cells, tissues, organs, organisms, environments) are interdependent in producing phenotypes.
(2) h2 assumes that genetic variance can be isolated and quantified as a distinct contributor to phenotypic variance.
(C) Since biological relativity rejects the isolation of genetic effects, h2 is conceptually invalid as a measure of genetic influence.
Conclusion
Both of these arguments show the same thing—h2 is a deeply flawed concept. The EEA critique exposes a methodological weakness: since MZ twins experience more similar environments than DZ twins, the excess environmental similarity experienced by MZs masquerades as genetic influence, leaving h2 incapable of isolating genetic variance.
But Noble’s biological relativity argument strikes at a deeper conceptual flaw in this practice, since it challenges the theoretical aspects of h2 itself. Since it highlights the interdependence of biological systems, it dismantles the reductionist notion that genetic effects can be separated from other levels of causation. The gene-centric assumption is at ends with the reality of phenotypes being emergent properties of multi-level interactions, which then renders the concept of h2 conceptually incoherent. Therefore, h2 isn’t only empirically questionable but it is theoretically untenable. The conceptual model is just not sound due to how genes really work (Burt and Simon, 2015)
Thus, again, hereditarianism fails conceptually. Even their main “tool” fails for a modicum of reasons not least the main theoretical killshot for heritability estimates—the principle of biological relativity. The reductionist hereditarian paradigm is conceptually and logically untenable, it’s time to throw it away, it’s time to throw it to the dustbin of history.
The “Interactionism Fallacy”
2350 words
A fallacy is an error in reasoning that makes an argument invalid. The “interactionism fallacy” is the fallacy—coined by Gottfredson (2009)—that since genes and environment interact, that heritability estimates are not useful—especially for humans (they are for nonhuman animals where environments can be fully controlled; see Schonemann, 1997; Moore and Shenk, 2016). There are many reasons why this ‘fallacy’ is anything but a fallacy; it is a simple truism: genes and environment (along with other developmental products) interact to ‘construct’ the organism (what Oyama, 2000 terms ‘constructive interactionism—“whereby each combination of genes and environmental influences simultaneously interacts to produce a unique result“). The causal parity thesis (CPT) is the thesis that genes/DNA play an important role in development, but so do other variables, so there is no reason to privilege genes/DNA above other developmental variables (see Noble, 2012 for a similar approach). Genes are not special developmental resources and so, nor are they more important than other developmental resources. So the thesis is that genes and other developmental resources are developmentally ‘on par’.
Genes need the environment. Without the environment, genes would not be expressed. Behavior geneticists claim to be able to partition genes from environment—nature from nurture—on the basis of heritability estimates, mostly gleaned from twin and adoption studies. However, the method is flawed: since genes interact with the environment and other genes, how would it be possible to neatly partition the effects of genes from the effects of the environment? Behavior geneticists claim that we can partition these two variables. Behavior geneticists—and others—cite the “Interactionism fallacy”, the fallacy that since genes interact with the environment that heritability estimates are useless. This “fallacy”, though, confuses the issue.
Behavior geneticists claim to show how genes and the environment affect the ontogeny of traits in humans with twin and adoption studies (though these methods are highly flawed). The purpose of this “fallacy” is to disregard what developmental systems theorists claim about the interaction of nature and nurture—genes and environment.
Gottfredson (2009) coins the “interactionism fallacy”, which is “an irrelevant truth [which is] that an organism’s development requires genes and environment to act in concert” and the “two forces are … constantly interacting” whereas “Development is their mutual product.” Gottfredson also states that “heritability … refers to the percentage of variation in … the phenotype, which has been traced to genetic variation within a particular population.” (She also makes the false claim that “One’s genome is fixed at birth“; though this is false, see epigenetics/methylation studies.) Heritability estimates, according to Phillip Kitcher are “‘irrelevant’ and the fact that behavior geneticists persist
in using them is ‘an unfortunate tic from which they cannot free themselves’ (Kitcher,
2001: 413)” (quoted in Griffiths, 2002).
Gottfredson is engaging in developmental denialism. Developmental denialism “occurs when heritability is treated as a causal mechanism governing the developmental reoccurrence of traits across generations in individuals.” Gottfredson, with her “interactionism fallacy” is denying organismal development by attempting to partition genes from environment. As Rose (2006) notes, “Heritability estimates are attempts to impose a simplistic and reified dichotomy (nature/nurture) on non-dichotomous processes.” The nature vs nurture argument is over and neither has won—contra Plomin’s take—since they interact.
Gottfredson seems confused, since this point was debated by Plomin and Oyama back in the 80s (Plomin’s review of Oyama’s book The Ontogeny of Information; see Oyama, 1987, 1988; Plomin, 1988a, b). In any case, it is true that development requires genes to interact. But Gottfredson is talking about the concept of heritability—the attempt to partition genes and environment through twin, adoption and family studies (which have a whole slew of problems). For example, Moore and Shenk (2016: 6) write:
Heritability statistics do remain useful in some limited circumstances, including selective breeding programs in which developmental environments can be strictly controlled. But in environments that are not controlled, these statistics do not tell us much.
Susan Oyama writes in The Ontogeny of Information (2000, pg 67):
Heritability coefficients, in any case, because they refer not only to variation in genotype but to everything that varied (was passed on) with it, only beg the question of what is passed on in evolution. All too often heritability estimates obtained in one setting are used to infer something about an evolutionary process that occurred under conditions, and with respect to a gene pool, about which little is known. Nor do such estimates tell us anything about development.
Characters are produced by the interaction of nongenetic and genetic factors. The biological flaw, as Moore and Shenk note, throw a wrench into the claims of Gottfredson and other behavior geneticists. Phenotypes are ALWAYS due to genetic and nongenetic factors interacting. So the two flaws of heritability—the environmental and biological flaw (Moore and Shenk, 2016)—come together to “interact” to refute such simplistic claims that genes and environment—nature and nurture—can be separated.
For instance, as Moore (2016) writes, though “twin study methods are among the most powerful tools available to quantitative behavioral geneticists (i.e., the researchers who took up Galton’s goal of disentangling nature and nurture), they are not satisfactory tools for studying phenotype development because they do not actually explore biological processes.” (See also Richardson, 2012.) This is because twin studies ignore biological/developmental processes that lead to phenotypes.
Gamma and Rosenstock (2017) write that the concept of heritability that behavioral geneticists use is “is a generally useless quantity” while “the behavioral genetic dichotomy of genes vs environment is fundamentally misguided.” This brings us back to the CPT; there is causal parity to all processes/interactants that form the organism and its traits, thus the concept of heritability that behavioral geneticists employ is a useless measure. Oyama, Griffiths, and Gray (2001: 3) write:
These often overlooked similarities form part of the evidence for DST’s claim of causal parity between genes and other factors of development. The “parity thesis” (Griffiths and Knight 1998) does not imply that there is no difference between the particulars of the causal roles of genes and factors such as endosymbionts or imprinting events. It does assert that such differences do not justify building theories of development and evolution around a distinction between what genes do and what every other causal factor does.
Behavior geneticists’ endeavor, though, is futile. Aaron Panofsky (2016: 167) writes that “Heritability estimates do not help identify particular genes or ascertain their functions in development or physiology, and thus, by this way of thinking, they yield no causal information.” (Also see Panofsky, 2014; Misbehaving Science: Controversy and the Development of Behavior Genetics.) So, the behavioral genetic method of partitioning genes and environment does not—and can not—show causation for trait ontogeny.
Now, while people like Gottfredson and others may deny it, they are genetic determinists. Genetic determinism, as defined by Griffiths (2002) is “the idea that many significant human characteristics are rendered inevitable by the presence of certain genes.” Using this definition, many behavior geneticists and their sympathizers have argued that certain traits are “inevitable” due to the presence of certain genes. Genetic determinism is literally the idea that genes “determine” aspects of characters and traits, though it has been known for decades that it is false.
Now we can take a look at Brian Boutwell’s article Not Everything Is An Interaction. Boutwell writes:
Albert Einstein was a brilliant man. Whether his famous equation of E=mc2 means much to you or not, I think we can all concur on the intellectual prowess—and stunning hair—of Einstein. But where did his brilliance come from? Environment? Perhaps his parents fed him lots of fish (it’s supposed to be brain food, after all). Genetics? Surely Albert hit some sort of genetic lottery—oh that we should all be so lucky. Or does the answer reside in some combination of the two? How very enlightened: both genes and environment interact and intertwine to yield everything from the genius of Einstein to the comedic talent of Lewis Black. Surely, you cannot tease their impact apart; DNA and experience are hopelessly interlocked. Except, they’re not. Believing that they are is wrong; it’s a misleading mental shortcut that has largely sown confusion in the public about human development, and thus it needs to be retired.
[…]
Most traits are the product of genetic and environmental influence, but the fact that both genes and environment matter does not mean that they interact with one another. Don’t be lured by the appeal of “interactions.” Important as they might be from time to time, and from trait to trait, not everything is an interaction. In fact, many things likely are not.
I don’t even know where to begin here. Boutwell, like Gottfredson, is confused. The only thing that needs to be retired because it “has largely sown confusion in the public about human development” is, ironically, the concept of heritability (Moore and Shenk, 2016)! I have no idea why Boutwell claimed that it’s false that “DNA and experience [environment] are hopelessly interlocked.” This is because, as Schneider (2007) notes, “the very concept of a gene requires an environment.” Since the concept of the gene requires the environment, how can we disentangle them into neat percentages like behavior geneticists claim to do? That’s right: we can’t. Do be lured by the appeal of interactions; all biological and nonbiological stuff constantly interacts with one another.
Boutwell’s claims are nonsense. It would be worth it to quote Richard Lewontin’s forward in the 2000 2nd edition of Susan Oyama’s The Ontogeny of Information (emphasis Lewontin’s):
Nor can we partition variation quantitatively, ascribing some fraction of variation to genetic differences and the remainder to environmental variation. Every organism is the unique consequence of the reading of its DNA in some temporal sequence of environments and subject to random cellular events that arise because of the very small number of molecules in each cell. While we may calculate statistically an average difference between carriers of one genotype and another, such average differences are abstract constructs and must not be reified with separable concrete effects of genes in isolation from the environment in which the genes are read. In the first edition of The Ontogeny of Information Oyama characterized her construal of the causal relation between genes and environment as interactionist. That is, each unique combination of genes and environment produces a unique and a priori unpredictable outcome of development. The usual interactionist view is that there are separable genetic and environmental causes, but the effects of these causes acting in combination are unique to the particular combination. But this claim of ontogenetically independent status of the causes as causes, aside from their interaction in the effects produced, contradicts Oyama’s central analysis of the ontogeny of information. There are no “gene actions” outside environments, and no “environmental actions” can occur in the absence of genes. The very status of environment as a contributing cause to the nature of an organism depends on the existence of a developing organism. Without organisms there may be a physical world, but there are no environments. In like the manner no organisms exist in the abstract without environments, although there may be naked DNA molecules lying in the dust. Organisms are the nexus of external circumstances and DNA molecules that make these physical circumstances into causes of development in the first place. They become causes only at their nexus, and they cannot exist as causes except in their simultaneous action. That is the essence of Oyama’s claim that information comes into existence only in the process of ontogeny. (Oyama, 2000: 16)
There is an “interactionist consensus” (see Oyama, Griffiths, and Grey, 2001; What is Developmental Systems Theory? pg 1-13): the organism and the suite of traits it has is due to the interaction of genetic/environmental/epigenetic etc. resources at every stage of development. Therefore, for organismal development to be successful, it always requires the interaction of genes, environment, epigenetic processes, and interactions between everything that is used to ‘construct’ the organism and the traits it has. Thus “it makes no sense to ask if a particular trait is genetic or environmental in origin. Understanding how a trait develops is not a matter of finding out whether a particular gene or a particular environment causes the trait; rather, it is a matter of understanding how the various resources available in the production of the trait interact over time” (Kaplan, 2006).
Lastly, I will shortly comment on Sesardic’s (2005: chapter 2) critiques on developmental systems theorists and their critique of heritability and the concept of interactionism. Sesardic argues in the chapter that interaction between genes and environment, nature and nurture, does not undermine heritability estimates (the nature and nurture partition). Philosopher of science Helen Longino argues in her book Studying Human Behavior (2013):
By framing the debate in terms of nature versus nurture and as though one of these must be correct, Sesardic is committed to both downplaying the possible contributions of environmentally oriented research and to relying on a highly dubious (at any rate, nonmethodological) empirical claim.
In sum, the “interactionist fallacy” (coined by Gottfredson) is not a ‘fallacy’ (error in reasoning) at all. For, as Oyama writes in Evolution’s Eye: A Systems View of the Biology-Culture Divide “A not uncommon reaction to DST is, ‘‘That’s completely crazy, and besides, I already knew it” (pg 195). This is exactly what Gottfredson (2009) states, that she “already knew” that there is an interaction between nature and nurture; but she goes on to deny arguments from Oyama, Griffiths, Stotz, Moore, and others on the uselessness of heritability estimates along with the claim that nature and nurture cannot be neatly partitioned into percentages as they are constantly interacting. Causal parity between genes and other developmental resources, too, upends the claim that heritability estimates for any trait make sense (not least for how heritability estimates are gleaned for humans—mostly twin, family, and adoption studies). Developmental denialism—what Gottfredson and others often engage in—runs rampant in the “behavioral genetic” sphere; and Oyama, Griffiths, Stotz, and others show how we should not deny development and we should discard with these estimates for human traits.
Heritability estimates imply that there is a “nature vs nurture” when it is “nature and nurture” which are constantly interacting—and, due to this, we should discard with these estimates due to the interaction of numerous developmental resources; it does not make sense to partition an interacting, self-organizing developmental system. Claims from behavior geneticists—that genes and environment can be separated—are clearly false.
DNA—Blueprint and Fortune Teller?
2500 words
What would you think if you heard about a new fortune-telling device that is touted to predict psychological traits like depression, schizophrenia and school achievement? What’s more, it can tell your fortune from the moment of your birth, it is completely reliable and unbiased — and it only costs £100.
This might sound like yet another pop-psychology claim about gimmicks that will change your life, but this one is in fact based on the best science of our times. The fortune teller is DNA. The ability of DNA to understand who we are, and predict who we will become has emerged in the last three years, thanks to the rise of personal genomics. We will see how the DNA revolution has made DNA personal by giving us the power to predict our psychological strengths and weaknesses from birth. This is a game-changer as it has far-reaching implications for psychology, for society and for each and every one of us.
This DNA fortune teller is the culmination of a century of genetic research investigating what makes us who we are. When psychology emerged as a science in the early twentieth century, it focused on environmental causes of behavior. Environmentalism — the view that we are what we learn — dominated psychology for decades. From Freud onwards, the family environment, or nurture, was assumed to be the key factor in determining who we are. (Plomin, 2018: 6, my emphasis)
The main premise of Plomin’s 2018 book Blueprint is that DNA is a fortune teller while personal genomics is a fortune-telling device. The fortune-telling device Plomin most discusses in the book is polygenic scores (PGS). PGSs are gleaned from GWA studies; SNP genotypes are then added up with scores of 0, 1, and 2. Then, the individual gets their PGS for trait T. Plomin’s claim—that DNA is a fortune teller—though, falls since DNA is not a blueprint—which is where the claim that “DNA is a fortune teller” is derived.
It’s funny that Plomin calls the measure “unbiased”, (he is talking about DNA, which is in effect “unbiased”), but PGS are anything BUT unbiased. For example, most GWAS/PGS are derived from European populations. But, for example, there are “biases and inaccuracies of polygenic risk scores (PRS) when predicting disease risk in individuals from populations other than those used in their derivation” (De La Vega and Bustamante, 2018). (PRSs are derived from statistical gene associations using GWAS; Janssens and Joyner, 2019.) Europeans make up more than 80 percent of GWAS studies. This is why, due to the large amount of GWASs on European populations, that “prediction accuracy [is] reduced by approximately 2- to 5-fold in East Asian and African American populations, respectively” (Martin et al, 2018). See for example Figure 1 from Martin et al (2018):
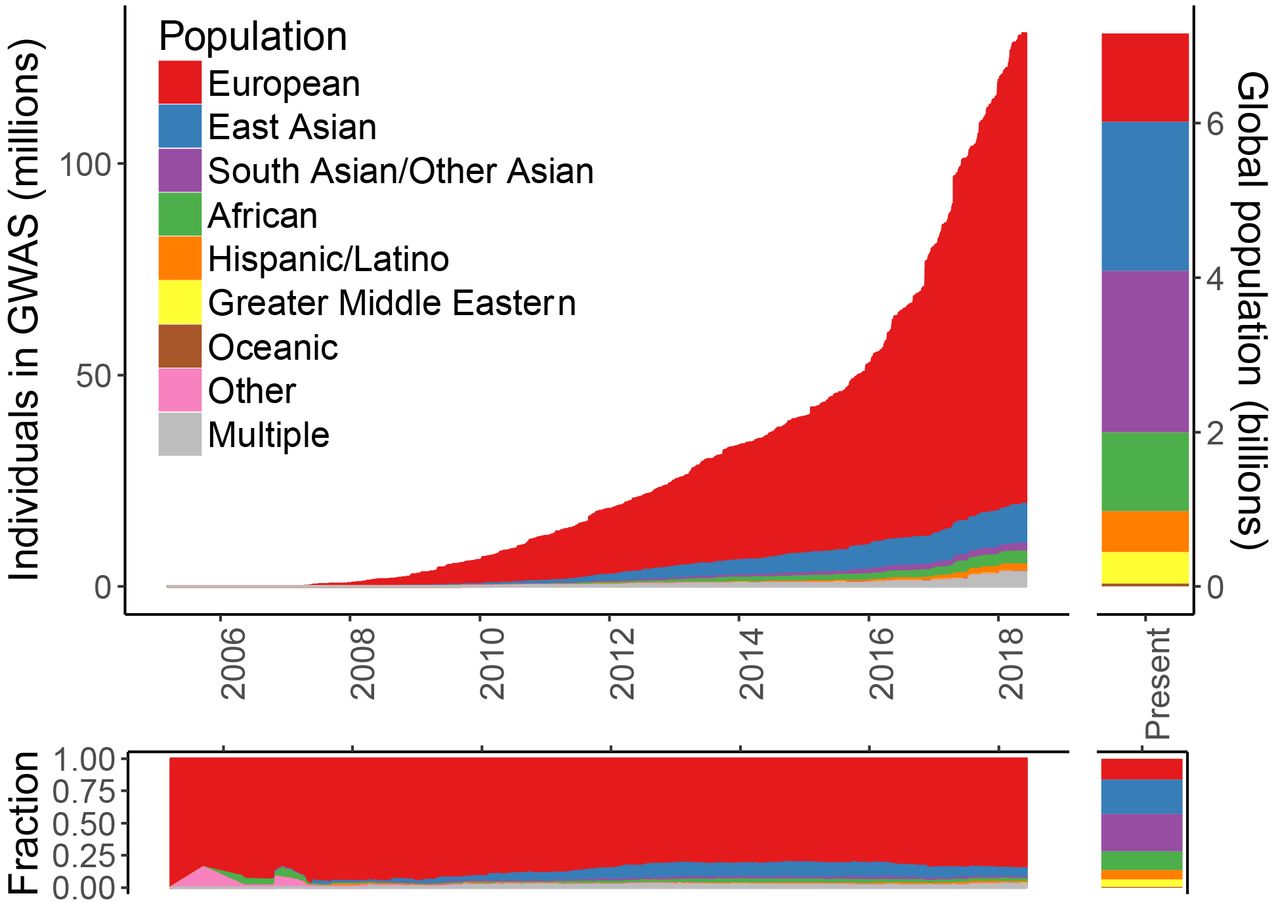
With the huge number of GWAS studies done on European populations, these scores cannot be used on non-European populations for ‘prediction’—even disregarding the other problems with PGS/GWAS.
By studying genetically informative cases like twins and adoptees, behavioural geneticists discovered some of the biggest findings in psychology because, for the first time, nature and nurture could be disentangled.
[…]
… DNA differences inherited from our parents at the moment of conception are the consistent, lifelong source of psychological individuality, the blueprint that makes us who we are. A blueprint is a plan. … A blueprint isn’t all that matters but it matters more than everything else put together in terms of the stable psychological traits that make us who we are. (Plomin, 2018: 6-8, my emphasis)
Nevermind the slew of problems with twin and adoption studies (Joseph, 2014; Joseph et al, 2015; Richardson, 2017a). I also refuted the notion that “A blueprint is a plan” last year, quoting numerous developmental systems theorists. The main thrust of Plomin’s book—that DNA is a blueprint and therefore can be seen as a fortune teller using the fortune-telling device to tell the fortunes of the people’s whose DNA are analyzed—is false, as DNA does not work how it does in Plomin’s mind.
These big findings were based on twin and adoption studies that indirectly assessed genetic impact. Twenty years ago the DNA revolution began with the sequencing of the human genome, which identified each of the 3 billion steps in the double helix of DNA. We are the same as every other human being for more than 99 percent of these DNA steps, which is the blueprint for human nature. The less than 1 per cent of difference of these DNA steps that differ between us is what makes us who we are as individuals — our mental illnesses, our personalities and our mental abilities. These inherited DNA differences are the blueprint for our individuality …
[DNA predictors] are unique in psychology because they do not change during our lives. This means that they can foretell our futures from our birth.
[…]
The applications and implications of DNA predictors will be controversial. Although we will examine some of these concerns, I am unabashedly a cheerleader for these changes. (Plomin, 2018: 8-10, my emphasis)
This quote further shows Plomin’s “blueprint” for the rest of his book—DNA can “foretell our futures from our birth”—and how it affects his conclusions gleaned from his work that he mostly discusses in his book. Yes, all scientists are biased (as Stephen Jay Gould noted), but Plomin outright claimed to be an unabashed cheerleader for his work. Plomin’s self-admission for being an “unabashed cheerleader”, though, does explain some of the conclusions he makes in Blueprint.
However, the problem with the mantra ‘nature and nurture’ is that it runs the risk of sliding back into the mistaken view that the effects of genes and environment cannot be disentangled.
[…]
Our future is DNA. (Plomin, 2018: 11-12)
The problem with the mantra “nature and nurture” is not that it “runs the risk of sliding back into the mistaken view that the effects of genes and environment cannot be disentangled”—though that is one problem. The problem is how Plomin assumes how DNA works. That DNA can be disentangled from the environment presumes that DNA is environment-independent. But as Moore shows in his book The Dependent Gene—and as Schneider (2007) shows—“the very concept of a gene requires the environment“. Moore notes that “The common belief that genes contain context-independent “information”—and so are analogous to “blueprints” or “recipes”—is simply false” (quoted in Schneider, 2007). Moore showed in The Dependent Gene that twin studies are flawed, as have numerous other authors.
Lewkowicz (2012) argues that “genes are embedded within organisms which, in turn, are embedded in external environments. As a result, even though genes are a critical part of developmental systems, they are only one part of such systems where interactions occur at all levels of organization during both ontogeny and phylogeny.” Plomin—although he does not explicitly state it—is a genetic reductionist. This type of thinking can be traced back, most popularly, to Richard Dawkins’ 1976 book The Selfish Gene. The genetic reductionists can, and do, make the claim that organisms can be reduced to their genes, while developmental systems theorists claim that holism, and not reductionism, better explains organismal development.
The main thrust of Plomin’s Blueprint rests on (1) GWA studies and (2) PGSs/PRSs derived from the GWA studies. Ken Richardson (2017b) has shown that “some cryptic but functionally irrelevant genetic stratification in human populations, which, quite likely, will covary with social stratification or social class.” Richardson’s (2017b) argument is simple: Societies are genetically stratified; social stratification maintains genetic stratification; social stratification creates—and maintains—cognitive differentiation; “cognitive” tests reflect prior social stratification. This “cryptic but functionally irrelevant genetic stratification in human populations” is what GWA studies pick up. Richardson and Jones (2019) extend the argument and argue that spurious correlations can arise from genetic population structure that GWA studies cannot account for—even though GWA study authors claim that this population stratification is accounted for, social class is defined solely on the basis of SES (socioeconomic status) and therefore, does not capture all of what “social class” itself captures (Richardson, 2002: 298-299).
Plomin also heavily relies on the results of twin and adoption studies—a lot of it being his own work—to attempt to buttress his arguments. However, as Moore and Shenk (2016) show—and as I have summarized in Behavior Genetics and the Fallacy of Nature vs Nurture—heritability estimates for humans are highly flawed since there cannot be a fully controlled environment. Moore and Shenk (2016: 6) write:
Heritability statistics do remain useful in some limited circumstances, including selective breeding programs in which developmental environments can be strictly controlled. But in environments that are not controlled, these statistics do not tell us much. In light of this, numerous theorists have concluded that ‘the term “heritability,” which carries a strong conviction or connotation of something “[in]heritable” in the everyday sense, is no longer suitable for use in human genetics, and its use should be discontinued.’ 31 Reviewing the evidence, we come to the same conclusion.
Heritability estimates assume that nature (genes) can be separated from nurture (environment), but “the very concept of a gene requires the environment” (Schneider, 2007) so it seems that attempting to partition genetic and environmental causation of any trait T is a fool’s—reductionist—errand. If the concept of gene depends on and requires the environment, then how does it make any sense to attempt to partition one from the other if they need each other?
Let’s face it: Plomin, in this book Blueprint is speaking like a biological reductionist, though he may deny the claim. The claims from those who push PRS and how it can be used for precision medicine are unfounded, as there are numerous problems with the concept. Precision medicine and personalized medicine are similar concepts, though Joyner and Paneth (2015) are skeptical of its use and have seven questions for personalized medicine. Furthermore, Joyner, Boros and Fink (2018) argue that “redundant and degenerate mechanisms operating at the physiological level limit both the general utility of this assumption and the specific utility of the precision medicine narrative.” Joyner (2015: 5) also argues that “Neo-Darwinism has failed clinical medicine. By adopting a broader perspective, systems biology does not have to.”
Janssens and Joyner (2019) write that “Most [SNP] hits have no demonstrated mechanistic linkage to the biological property of interest.” Researchers can show correlations between disease phenotypes and genes, but they cannot show causation—which would be mechanistic relations between the proposed genes and the disease phenotype. Though, as Kampourakis (2017: 19), genes do not cause diseases on their own, they only contribute to its variation.
Edit: Take also this quote from Plomin and Stumm (2018) (quoted by Turkheimer):
GPS are unique predictors in the behavioural sciences. They are an exception to the rule that correlations do not imply causation in the sense that there can be no backward causation when GPS are correlated with traits. That is, nothing in our brains, behaviour or environment changes inherited differences in DNA sequence. A related advantage of GPS as predictors is that they are exceptionally stable throughout the life span because they index inherited differences in DNA sequence. Although mutations can accrue in the cells used to obtain DNA, like any cells in the body these mutations would not be expected to change systematically the thousands of inherited SNPs that contribute to a GPS.
Turkheimer goes on to say that this (false) assumption by Plomin and Stumm (2018) assumes that there is no top-down causation—i.e., that phenotypes don’t cause genes, or there is no causation from the top to the bottom. (See the special issue of Interface Focus for a slew of articles on top-down causation.) Downward causation exists in biological systems (Noble, 2012, 2017), as does top-down. The very claim that “nothing in our brains, behaviour or environment changes inherited differences in DNA sequence” is ridiculous! This is something that, of course, Plomin did not discuss in Blueprint. But in a book that, supposedly, shows “how DNA makes us who we are”, why not discuss epigenetics? Plomin is confused, because DNA methylation impacts behavior and behavior impacts DNA methylation (Lerner and Overton, 2017: 114). Lerner and Overtone (2017: 145) write that:
… it should no longer be possible for any scientist to undertake the procedure of splitting of nature and nurture and, through reductionist procedures, come to conclusions that the one or the other plays a more important role in behavior and development.
Plomin’s reductionist takes, therefore again, fail. Plomin’s “reluctance” to discuss “tangential topics” to “inherited DNA differences” included epigenetics (Plomin, 2018: 12). But it seems that his “reluctance” to discuss epigenetics was a downfall in his book as epigenetic mechanisms can and do make a difference to “inherited DNA differences” (see for example, Baedke, 2018, Above the Gene, Beyond Biology: Toward a Philosophy of Epigenetics and Meloni, 2019, Impressionable Biologies: From the Archaeology of Plasticity to the Sociology of Epigenetics see also Meloni, 2018). The genome can and does “react” to what occurs to the organism in the environment, so it is false that “nothing in our brains, behaviour or environment changes inherited differences in DNA sequence” (Plomin and Stumm, 2018), since our behavior and actions can and do methylate our DNA (Meloni, 2014) which falsifies Plomin’s claim and which is why he should have discussed epigenetics in Blueprint. End Edit
Conclusion
So the main premise of Plomin’s Blueprint is his two claims: (1) that DNA is a fortune teller and (2) that personal genomics is a fortune-telling device. He draws these big claims from PGS/PRS studies. However, over 80 percent of GWA studies have been done on European populations. And, knowing that we cannot use these datasets on other, non-European datasets, greatly hampers the uses of PGS/PRS in other populations—although the PGS/PRS are not that useful in and of itself for European populations. Plomin’s whole book is a reductionist screed—“Sure, other factors matter, but DNA matters more” is one of his main claims. Though, a priori, since there is no privileged level of causation, one cannot privilege DNA over any other developmental variables (Noble, 2012). To understand disease, we must understand the whole system and how when one part of the system becomes dysfunctional how it affects other parts of the system and how it runs. The PGS/PRS hunts are reductionist in nature, and the only answer to these reductionist paradigms are new paradigms from systems biology—one of holism.
Plomin’s assertions in his book are gleaned from highly confounded GWA studies. Plomin also assumes that we can disentangle nature and nurture—like all reductionists. Nature and nurture interact—without genes, there would be an environment, but without an environment, there would be no genes as gene expression is predicated on the environment and what occurs in it. So Plomin’s reductionist claim that “Our future is DNA” is false—our future is studying the interactive developmental system, not reducing it to a sum of its parts. Holistic biology—systems biology—beats reductionist biology—the Neo-Darwinian Modern Synthesis.
DNA is not a blueprint nor is it a fortune teller and personal genomics is not a fortune-telling device. The claim that DNA is a blueprint/fortune teller and personal genomics is a fortune-telling device come from Plomin and are derived from highly flawed GWA studies and, further, PGS/PRS. Therefore Plomin’s claim that DNA is a blueprint/fortune teller and personal genomics is a fortune-telling device are false.
(Also read Erick Turkheimer’s 2019 review of Plomin’s book The Social Science Blues, along with Steve Pitteli’s review Biogenetic Overreach for an overview and critiques of Plomin’s ideas. And read Ken Richardson’s article It’s the End of the Gene As We Know It for a critique of the concept of the gene.)
Twin Studies, Adoption Studies, and Fallacious Reasoning
2350 words
Twin and adoption studies have been used for decades on the basis that genetic and environmental causes of traits and their variation in the population could be easily partitioned by two ways: one way is to adopt twins into separate environments, the other to study reared-together or reared-apart twins. Both methods rest on a large number of (invalid) assumptions. These assumptions are highly flawed and there is no evidential basis to believe these assumptions, since the assumptions have been violated which invalidates said assumptions.
Plomin et al write (2013) write: For nearly a century, twin and adoption studies have yielded substantial estimates of heritability for cognitive abilities.
But the validity of the “substantial estimates of heritability for cognitive abilities” is strongly questioned due to unverified (and false) assumptions that these researchers make.
Adoption studies
The problem with adoption studies are numerous, not least: restricted range of adoptive families; selective placement; late separation; parent-child attachment disturbance; problems with the tests (on personality, ‘IQ’); the non-representativeness of adoptees compared to non-adoptees; and the reliability of the characteristic in question.
In selective placement, the authorities attempt to place children in homes close to their biological parents. They gage how “intelligent” they believe they are (on the basis of parental SES and the child’s parent’s perceived ‘intelligence’), thusly this is a pretty huge confound for adoption studies.
According to adoption researcher Harry Munsinger, a “possible source of bias in adoption studies is selective placement of adopted children in adopting homes that are similar to their biological parents’ social and educational backgrounds.” He recognized that “‘fitting the home to the child’ has been the standard practice in most adoption agencies, and this selective placement can confound genetic endowment with environmental influence to invalidate the basic logic of an adoptive study (Munsinger, 1975, p. 627). Clearly, agency policies of “fitting the home to the child” are a far cry from random placement of adoptees into a wide range of adoptive homes. (Joseph, 2015: 30-1)
Richardson and Norgate (2005) argue that simple additive effects for both genetic and environmental effects are false; that IQ is not a quantitative trait; while other interactive effects could explain the IQ correlation.
1) Assignment is nonrandom. 2) They look for adoptive homes that reflect the social class of the biological mother. 3) This range restriction reduces the correlation estimates between adopted children and adopted parents. 4) Adoptive mothers come from a narrow social class. 5) Their average age at testing will be closet to their biological parents than adopted parents. 6) They experience the womb of their mothers. 7) Stress in the womb can alter gene expression. 8) Adoptive parents are given information about the birth family which may bias their treatment. 9) Biological mothers and adopted children show reduced self-esteem and are more vulnerable to changing environments which means they basically share environment. 10) Conscious or unconscious aspects of family treatment may make adopted children different from other adopted family members. 11) Adopted children also look more like their biological parents than their adoptive parents which means they’ll be treated accordingly.
Twin studies
Personally, my favorite thing to discuss. Twin studies rest on the erroneous assumption that DZ and MZ environments are equal; that they get treated equally the same. This is false, MZ twins get treated more similarly than DZ twins, which twin researchers have conceded decades ago. But in order to save their field, they attempt to use circular argumentation, known as Argument A. Argument A states that MZTs (monozygotic twins reared together) are more genetically similar than DZTs (dizygotic twins reared together) and thusly this causes greater behavioral similarity. But this is based on circular reasoning: the researchers already implicitly assumed that genes played a role in their premise and, not surprisingly, in their conclusion genes are the cause for the similarities of the MZTs. So Argument A is used, twin researchers circularly assume that MZTs greater behavioral similarity is due to genetic similarity, while their argument that genetic factors explain the greater behavioral similarity of MZTs is a premise and conclusion of their argument. “X is true because Y is true; Y is true because X is true.” (Also see Joseph et al, 2015.)
We have seen that circular reasoning is “empty reasoning in which the conclusion rests on an assumption whose validity is dependent on the conclusion” (Reber, 1985, p. 123). … A circular argument consists of “using as evidence a fact which is authenticated by the very conclusion it supports,” which “gives us two unknowns so busy chasing each other’s tails that neither has time to attach itself to reality” (Pirie, 2006, p. 27) (Joseph, 2016: 164).
Even if Argument A is accepted, the causes of behavioral similarities between MZ/DZ twins could still come down to environment. Think of any type of condition that is environmentally caused but is due to people liking what causes the condition. There are no “genes for” that condition, but their liking the thing that caused the condition caused an environmental difference.
Argument B also exists. Those that use Argument B also concede that MZs experience more similar environments, but then argue that in order to show that twin studies, and the EEA, are false, critics must show that MZT and DZT environments differ in the aspects that are relevant to the behavior in question (IQ, schizophrenia, etc).
An example of an Argument B environmental factor relevant to a characteristic or disorder is the relationship between exposure to trauma and post-traumatic stress disorder (PTSD). Because trauma exposure is (by definition) an environmental factor known to contribute to the development of PTSD, a finding that MZT pairs are more similarly exposed to trauma than DZT pairs means that MZT pairs experience more similar “trait-relevant” environments than DZTs. Many twin researchers using Argument B would conclude that the EEA is violated in this case. (Joseph, 2016: 165)
So twin researchers need to rule out and identify “trait-relevant factors” which contribute to the cause of said trait, along with experiencing more similar environments, invalidates genetic interpretations made using Argument B. But Argument A renders Argument B irrelevant because even if critics can show that MZTs experience more similar “trait-relevant environments”, they could still argue that the twin method is valid by stating that (in Argument A fashion) MZTs create and elicit more similar trait-relevant environments.
One more problem with Argument A is that it shows that twins behave accordingly to “inherited environment-creating blueprint” (Joseph, 2016: 164) but at the same time shows that parents and other adults are easily able to change their behaviors to match that of the behaviors that the twins show, which in effect, allows them to “create” or “elicit” their own environments. But the adults’ “environment-creating behavior and personality” should be way more unchangeable than the twins’ since along with the presumed genetic similarity, adults have “experienced decades of behavior-molding peer, family, religious, and other socialization influences” (Joseph, 2016: 165).
Whether or not circular arguments are “useful” or not has been debated in the philosophical literature for some time (Hahn, Oaksford, and Corner, 2005). However, assuming, in your premise, that your conclusion is valid is circular and therefore While circular arguments are deductively valid, “it falls short of the ultimate goal of argumentative discourse: Whatever evaluation is attached to the premise is transmitted to the conclusion, where it remains the same; no increase in degree of belief takes place” (Hahn, 2011: 173).
However, Hahn (2011: 180) concludes that “the existence of benign circularities makes clear that merely labeling something as circular is not enough to dismiss it; an argument for why the thing in question is bad still needs to be made.” This can be simply shown: The premise that twin researchers use (that genes cause similar environments to be constructed) is in their conclusion. They state in their premise that MZT behavioral similarity is due to greater MZT genetic similarity in comparison to DZTs (100 vs. 50 percent). Then, in the conclusion, they re-state that the behavioral similarities of MZTs is due to their genetic similarity compared to DZTs (100 vs. 50 percent). Thus, a convincing argument for conclusion C (that genetic similarity explains MZT behavioral similarity) cannot rest on the assumption that conclusion C is correct. Thus, Argument A is fallacious due to its circularity.
What causes MZT behavioral similarities is their more similar environment: they get treated the same by peers and parents, and have higher rates of identity confusion and had a closer emotional bond compared to DZTs. The twin method is based on the (erroneous) assumption that MZT and DZT pairs experience roughly equal environments, which twin researchers conceded was false decades ago.
Richardson and Norgate (2005: 347) conclude (emphasis mine):
We have shown, first, that the EEA may not hold, and that well-demonstrated treatment effects can, therefore, explain part of the classic MZ–DZ differences. Using published correlations, we have also shown how sociocognitive interactions, in which DZ twins strive for a relative ‘apartness’, could further depress DZ correlations, thereby possibly explaining another part of the differences. We conclude that further conclusions about genetic or environmental sources of variance from MZ–DZ twin data should include thorough attempts to validate the EEA with the hope that these interactions and their implications will be more thoroughly understood.
Of course, even if twin studies were valid and the EEA was true/ the auxiliary arguments used were true, this would still not mean that heritability estimates would be of any use to humans, since we cannot control environments as we do in animal breeding studies (Schonemann, 1997; Moore and Shenk, 2016). I have chronicled how 1) the EEA is false and how flawed twin studies are; 2) how flawed heritability estimates are; 3) how heritability does not (and cannot) show causation; and 4) the genetic reductionist model that behavioral geneticists rely on is flawed (Lerner and Overton, 2017).
So we can (1) accept the EEA, that the greater behavioral resemblance indicates the importance of genetic factors underlying most human behavioral differences and behavioral disorders or we can (2) reject the EEA and state that the greater behavioral resemblance is due to nongenetic (environmental) factors, which means that all genetic interpretations of MZT/DZT studies must be rejected. Thus, using (2), we can infer that all twin studies measure is similarity of the environment of DZTs, and it is, in fact, not measuring genetic factors. Accepting explanation 2 does not mean that “twin studies overestimate heritability, or that researchers should assess the EEA on a study-by-study basis, but instead indicates that the twin method is no more able than a family study to disentangle the potential influences of genes and environment” (Joseph, 2016: 181).
What it does mean, however, is that we can, logically, discard all past, future, and present MZT and DZT comparisons and these genetic interpretations must be outright rejected, due to the falsity of the EEA and the fallaciousness of the auxiliary arguments made in order to save the EEA and the twin method overall.
There are further problems with twin studies and heritability estimates. Epigenetic supersimilarity (ESS) also confounds the relationship. Due to the existence of ESS “human MZ twins clearly cannot be viewed as the epigenetic equivalent of isogenic inbred mice, which originate from separate zygotes. To the extent that epigenetic variation at ESS loci influences human phenotype, as our data indicate, the existence of ESS establishes a link between early embryonic epigenetic development and adult disease and may call into question heritability estimates based on twin studies” (Van Baak et al, 2018). In other words, ESS is an unrecognized phenomenon that contributes to the phenotypic similarity of MZs, which calls into question the usefulness of heritability studies using twins. The uterine environment has been noted to be a confound by numerous authors (Devlin, Daniels, and Roeder, 1997; Charney, 2012; Ho, 2013; Moore and Shenk, 2016).
Conclusion
Adoption studies fall prey to numerous pitfalls, most importantly, that children are adopted into similar homes compared to their birth parents, which restricts the range of environments for adoptees. Adoption placement is also non-random, the children are placed into homes that are similar to their biological parents. Due to these confounds (and a whole slew of other invalidating problems), adoption studies cannot be said to show genetic causation, nor can they separate genetic from environmental factors.
Twin studies suffer from the biggest flaw of all: the falsity of the EEA. Since the EEA is false—which has been recognized by both critics and supporters of the assumption—the supporters of the assumption have attempted to redefine the EEA in two ways: (1) that MZTs experience more similar environments due to genetic similarity (Argument A) and (2) that it is not whether MZTs experience more similar environments, but whether or not they share more similar trait-relevant environments. Thus, unless these twin researchers are able to identify trait-relevant factors that contribute to the trait in question, we must conclude that (along with the admission from twin researchers that the EEA is false; that MZTs experience more similar environments than DZTs) genetic interpretations made using Argument B are thusly invalidated. Fallacious reasoning (“X causes Y; Y causes X) does not help any twin argument. Because their conclusion is already implicitly assumed in their premise.
The existence of ESS (epigenetic supersimilarity) further shows how invalid the twin method truly is, because the confounding starts in the womb. Attempts can be made (however bad) to control for shared environment by adopting different twins into different homes, but they still shared a uterine environment which means they shared an environment, which means it is a confound and it cannot be controlled for (Charney, 2012).
Adoption and twin studies are highly flawed. Like family studies, twin studies are no more able to disentangle genetic from environmental effects than a family study, and thus twin studies cannot separate genes from environment. Last, and surely not least, it is fallacious to assume that genes can be separated so neatly into “heritability estimates” as I have noted in the past. Heritability estimates cannot show genetic causation, nor can it show how malleable a trait is. They’re just (due to how we measure) flawed measures that we cannot fully control so we must make a number of (false) assumptions that then invalidate the whole paradigm. The EEA is false, all auxiliary arguments made to save the EEA are fallacious; adoption studies are hugely confounded; twin studies are confounded due to numerous reasons, most importantly the uterine environment (Van Baak et al, 2018).
Behavior Genetics and the Fallacy of Nature vs Nurture
3250 words
People appeal to moderate to high heritability estimates as evidence that a trait is controlled by genes. They then assume that because something has a high heritability then that it must show something about causation. The fact of the matter is, they do not. Heritability estimates assume a false dichotomy of nature vs nurture; it assumes that we can neatly partition genetic from environmental effects. It assumes that the higher a trait’s heritability the more genes control said trait. These are all false. One of the main ways that heritability is estimated is by the CTM (classic twin method). This method, though, has a ton of assumptions poured into it—most importantly, the assumption that DZ and MZ fraternal twins experience roughly equal environments—the equal environments assumption (EEA). Heritability studies are useless for humans; twin studies bias estimates upwards with a whole host of assumptions.
I will show that i) heritability estimates are highly flawed (due to erroneous assumptions); ii) nature vs nurture cannot be separated (like behavior geneticists claim) and so their main tool (the heritability estimate) should be discontinued; iii) genetic reductionism is not a tenable model due to what we now know about how genes work. All three of these reasons are enough to discontinue heritability estimates. If the nature vs nurture debate rests on a fallacy, and this fallacy is used as a vehicle for heritability estimates, then they should be discontinued for humans and only be used for breeding animals where they can control the environment fully (Schonemann, 1997; Moore and Shenk, 2016).
Heritability, twin studies, and equal environments
Back in 2014-2015, there was a debate in the criminological literature that had implications for heritability studies as a whole. Burt and Simons (2014) stated that it was time to get rid of heritability studies. Barnes et al (2015) responded that this was “a de facto form of censorship” (pg 2). Joseph et al (2015) respond to these accusations, writing, “It was good science and not “censorship” when earlier scientists called for ending studies based on craniometry, phrenology, and physiognomy, and any contemporary criminologist calling for the use of astrological charts to predict whether certain people will commit violent crimes would be justifiably ridiculed.” The main thing here, in my opinion, is that heritability estimates are based on an oversimplified (and wrong) model of the gene. Partitioning variance assumes that you can partition how much a trait is influenced by “nature” or “nurture” which is a false dichotomy (Moore, 2002; Schneider, 2007; Moore and Shenk, 2016).
More importantly, no “genes have been found” (I know that’s everyone’s favorite thing to hear) for traits that supposedly have high heritabilities. On page 179 of his book (nook version), Misbehaving Science, Controversy and the Development of Behavior Genetics Panofsky (2014) writes:
Molecular genetics has been a major dissapointment, if not an outright failure, in behavior genetics. Scientists have made many bold claims about genes for behavioral traits or mental disorders only to later retract them or to have them not be replicated by other scientists. Further, the findings that have been confirmed, or not yet falsified, have been few, far between, and small in magnitude.
There seems to be a huge disconnect between heritability estimates gleaned from twin studies and what the actual molecular genetic evidence says. This is because the EEA—that fraternal MZ twins experience roughly similar environments compared to fraternal DZ twins—is false. Fraternal MZ twins end up experiencing more similar environments when compared with fraternal DZ twins. Though most researchers attempt to save face by stating that MZ twins “seek out” and “elicit” their own environments which then makes them more similar compared to DZ twins. However, this is circular logic. The conclusion (that twins experience more similar environments) is in the premise, and therefore it is an invalid argument due to the logical fallacy. (It should also be noted that identical twins’ genes are not identical.)
Heritability studies assume an outdated model of the gene. The flaw regarding heritability estimates is simple: they imply a false dichotomy of nature vs nurture, while also assuming that genes and environment are independent, while the contribution to complex behaviors can be precisely quantified (Charney, 2013). This is one of the most critical parts of the heritability debate. Prenatal environments of DZ twins “can be significantly more stressful than that of DZ twins, and hence a cause of greater stress-related phenotypic concordance, the equal environment assumption will not hold in relation to behavioral phenotypes potentially associated with prenatal stress” (Charney, 2012: 20). This also is cause for concern regarding studies of twins reared apart. While twins are reared apart to eliminate shared environmental confounds, it cannot eliminate perhaps the most important confound of all—the prenatal environment (Moore and Shenk, 2016).
One of the most-cited studies regarding twins reared apart is Bouchard (1990). Though there are a whole slew of problems with this study.
1) You have the huge confound of similar environments before birth.
2) Full details for the MISTRA have never been published, so we don’t know how ‘separated’ the twins were. Though Bouchard et al do say that they were separated between 0 to 48.7 months (table 1) so some pairs spent at least 4 years together. Some of the twins even had reunions and spent a lot of time together.
3) They’re not representative and twins who do sign up for this research are self-selecting. Ken Richardson says in his book (2017, pg 55): “Twins generally tend to be self-selecting in any twin study. They may have responded to advertisements placed by investigators or have been prompted to do so by friends or family, on the grounds that they are alike. Remember, at least some of them knew each other prior to the study. Jay Joseph has suggested that the twins who elected to participate in all twin studies are likely to be more similar to one another than twins who chose not to participate. This makes it difficult to claim that the results would apply to the general population.”
4) And the results aren’t fully reported. Richardson also states that (2017, pg 55) “… of two IQ tests administered in the MISTRA, results have been published for one but not the other. No explanation was given for that omission. Could it be they produced different results?” He even states that attempts to get the data, by researchers like Jay Joseph, have been denied. Why would you refuse to publish, or give to another researcher, your data when asked?
We don’t know the relevant environments, the children’s average age at testing is closer to the biological mother than adopted mother; the biological mother and child will have reduced self-esteem and be more vulnerable to difficult situations, and in this sense they share environments; and conscious or unconscious bias make adopted children different from other family members. Adoption agencies also attempt to put children into similar homes as the biological mother too.
Charney (2012: 25) brings up an important point: “For phenotypes of any degree of complexity, DNA does not contain a determinate genetic program (analogous to the digital code of a computer) from which we can predict phenotype. If DNA were the sole carrier of information relevant to phenotype formation, and contained a genetic program sufficiently determinate that solely by reading it we could predict phenotype, then humans (and all other organisms) would be largely lacking in phenotypic plasticity.” Moore and Shenk (2016) also state that “we inherit developmental resources, not traits.”
1 For twin studies to be valid DZ twins and MZ fraternal twins would have to experience roughly equal environments. 2 Fraternal MZ twins experience much more similar environments than DZ twins. 3 Therefore the EEA is false and no genetic interpretations can be drawn from the data.
Heritability estimates cannot detangle genes and environment, and therefore they should be discontinued or reinterpreted (Joseph et al, 2015). Burt and Simons (2014: 110) also conclude: “Rejecting heritability studies and the false nature–nurture dichotomy and gene-centric model on which they are grounded is a necessary step forward that will pave the way for a reconceptualization of the link between the biological and the social in shaping criminal propensities in ways that are consistent with postgenomic knowledge“. I disagree with Barnes et al (2015) when they say that ending heritability estimates are “a defacto form of censorship“, because if nature vs nurture is a false dichotomy and the gene-centric model that heritability estimates rely on is wrong, then we need to either discontinue or reinterpret the estimates, not saying that ‘this is how much nature contributes to X and this is how much nurture contributes to Y’. (See also Richardson and Norgate, 2005 for more arguments regarding the EEA.)
Sapolsky (2017: 219) writes:
Oh, that’s right, humans. Of all species, heritability scores in humans plummet the most when shifting from a controlled experimental setting to considering the species’ full range of habitats. Just consider how much the heritability score for wearing earrings, with its gender split, has declined since 1958.
Heritability flaws
High heritability estimates have been used as evidence for causation—that genes control a large part of the trait in question. This reasoning, however, is highly flawed. People confuse “heritable” with “inheritable” (Moore and Shenk, 2016). Heritability does not inform us what causes a trait, how much environment contributes to a trait, nor does it tell us the relative influence of genes on a trait. Moore and Shenk (2016) agree with Joseph et al (2015) and Burt and Simons (2014) that heritability studies need to end, but Moore and Shenk’s reasoning slightly differs: they say we should end estimates because people confuse “heritable” with “inheritable”. Likewise, Guo (2000: 299) concurs, writing “it can be argued that the term ‘heritability’, which carries a strong conviction or connotation of something ‘heritable’ in everyday sense, is no longer suitable for use in human genetics and its use should be discontinued.”
Some may say that if a trait turns out to be mildly heritable then we can say that genes have some effect, but we know that genes affect all traits so it seems kind of redundant to have a useless measure that assumes a false dichotomy and relies on an outdated, additive model of the gene.
Rose (2006), too, agrees that heritability estimates imply a false dichotomy of nature vs nurture onto biological systems:
Biological systems are complex, non-linear, and non-additive. Heritability estimates are attempts to impose a simplistic and reified dichotomy (nature/nurture) on non-dichotomous processes.
Likewise, Lewontin (2006) argues we should be analyzing and studying causes, not variance.
There are numerous hereditarian scientific fallacies which include: 1) trait heritability does not predict what would occur when environments/genes change; 2) they’re inaccurate since they don’t account for gene-environment covariation or interaction while also ignoring nonadditive effects on behavior and cognitive ability; 3) molecular genetics does not show evidence that we can partition environment from genetic factors; 4) it wouldn’t tell us which traits are ‘genetic’ or not; and 5) proposed evolutionary models of human divergence are not supported by these studies (since heritability in the present doesn’t speak to what traits were like thousands of years ago) (Bailey, 1997).
Bailey (1997) brings up important arguments against the use of heritability, and even discusses fallacious writing from Rushton on the matter:
Rushton (1995), for example, thinks that if observed differences among the
racial groups that he defines are higher for traits that have high heritability within the groups, the hypothesis of genetically caused differences among the groups is supported.
Bailey (1997) then goes on to discuss three lakes: Otter lake, Welcome lake, and Bark lake. Otter lake has very high primary production, while Bark lake has very little and Welcome lake is somewhere in between (you can see that ‘Otter’, ‘Bark’ and ‘Welcome’ lakes are analogies for ‘Orientals’, ‘Blacks’, and ‘Whites’ as said by Rushton). But there is variation within the lakes, there are high production pockets of water in Bark lake while there are low production pockets of water in Otter lake. All three lakes are visited and measurements are taken. Bailey (1997) states that his conclusion would be that they differ in how much light each receives. Bailey (1997: 131) writes:
If I substitute three groups of people for my lakes, IQ for primary production, and genes for light levels, the fallacy of the slippery scale, as applied to human behaviour genetics, becomes clear. Even if we are sure that there is a difference among groups of people in IQ, and we are sure that IQ has high heritability within
each of the groups (i.e. variation in IQ is largely caused by genetic variation), we can make no inference about the cause of differences in IQ among the groups. The differences might be caused by genetic differences or they might not, but the heritability studies within the groups can’t help us make that judgment.

(Genes don’t cause IQ scores—or behavior—but that’s for another day.)
Heritability estimates for, say, IQ, are higher than any other trait in the animal kingdom. Heritability estimates for animal traits are low—lower than the stratospheric heritability of IQ. For example, heritability estimates of the bodyweight of farm animals is about 30 percent, which is the same for egg and milk production. Body fat in pigs and wool on sheep has a heritability of about 50 percent. But these estimates pale in comparison to the heritability estimates of IQ: estimates have been as high as 80 percent (but Schonemann, 1997 states it’s 60 percent but it’s as high as 80-90 percent today); this heritability estimate for IQ “surpasses almost anything found in the animal kingdom” (Schonemann, 1997: 104).
This high heritability estimate for IQ, of course, comes to us from the highly flawed twin studies discussed above. The reason why farmers and botanists use heritability estimates is that they can perfectly control the environment, and therefore get accurate—or close enough to it—estimates that will help them in their breeding efforts. Conversely, for humans, environments cannot be perfectly controlled and it is, of course, unethical to rear twins, MZ and DZ, in a controlled environment. Proponents of the twin method may say “It doesn’t matter if it’s flawed, it still shows there is a genetic component to trait X!”. But as discussed by Moore and Shenk (2016), that’s irrelevant because genetic factors influence all of our characteristics.
Heritability and causation
In the final section, I will shortly discuss how people fallaciously assume that high heritability estimates imply that a trait is strongly influenced by genetic factors.
In his essay in the book Postgenomics: Perspectives on Biology After the Genome, sociologist Aaron Panofsky (2016: 167; nook version) writes:
Heritability estimates do not help identify particular genes or ascertain their functions in development or physiology, and thus, by this way of thinking, they yield no causal information.
This is important to note: to those who truly believe that heritability estimates tell us anything about causation, how could they, logically, give us causal information if genes that lead to trait variation are not identified (Richardson, 2012)?
Panofsky (2014: 102-103) writes:
Experimental evidence from plants and animals suggest that shapes of the curves cannot inferred in advance and rarely follow the smooth, nonintersecting pattern like in figure 3.2. Thus true causal interpretations of heritability are hopeless and must be abandoned. Behavior geneticists did not claim direct experimental evidence, but they thought these various indirect lines of evidence provided a reasonable set of assumptions that would enable them to interpret heritability scores causally—provided they offer apporopriate, reasonable qualifications.
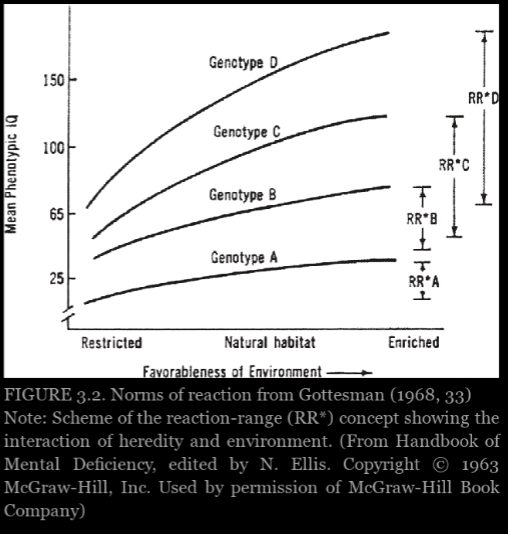
Graph from Panofsky (2014: 103)
Heritability estimates imply nothing about causation. It is about associations with variance, not identity and causes (Richardson, 2017: 69). A heritability of 0 does not mean that genes do not play a role in the development of form and function and phenotypic variation, it just means that, for whatever reason, there is little correlation between the two.
Scheneider (2007) writes (emphasis mine):
Heritability estimates apply only to groups, and are inherently inapplicable to individuals in any sense. And they do not imply causation. As Moore notes, all of these important limitations have been frequently ignored or minimized.
Reductionism
Heritability estimates imply nothing about causation. Behavior geneticists and others assume that heritability estimates will lead to ‘finding the genes’ that ’cause’ or are ‘associated with’ behavior. Their models are also, of course, extremely reductionist. It is then important to note that genes do not determine behavior. To quote Lerner and Overton (2017: 114):
Data presented in a 2016 special section of the journal Child Development indicate
that “some behaviors may be affected by only slight changes in DNA methylation,
while others may require a larger percent change in methylation; of course, the
effects are also likely bidirectional, with behavior impacting changes in methylation” [Lester et al., 2016, p. 31]. This point is key . It underscores the absurdity of genetic reductionist models: Genes do not determine behavior.
Methylation impacts behavior; behavior impacts methylation. It is the relations between methylation and behavior, not the genes acting as the “command center”, the “executive” of human behavior and development, that constitute the basic role of biology across the developmental course. This is the fatal flaw of reductionist models. Lastly, Lerner and Overton (2017: 145) write (emphasis mine):
That is, with the recent advances in understanding the role of epigenetics and recent research findings supporting this role, it should no longer be possible for any scientist to undertake the procedure of splitting of nature and nurture and, through reductionist procedures, come to conclusions that the one or the other plays a more important role in behavior and development.
[Richardson (2017: 129) also writes: “Note that this environmental source of [epigenetic] variation will appear in the behavioral geneticists twin-study as genetic variation: quite probably another way in which heritability estimates are distorted.”]
Reductionism in biology is fatally flawed. Reductionism, of course, has greatly increased our understanding of biology. However, it is time to move past the false dichotomy of nature vs nurture, and with that, move past heritability estimates since they prop up the fallacy of nature vs nurture. There is no way to separate the two since they are intertwined, but behavior geneticists would like you to believe that by studying twins raised apart will tell you anything about how ‘genetic’ or ‘environmental’ variation in a trait is in a population. Since heritability estimates are gleaned from the highly flawed studies of twins reared apart, a whole host of assumptions is poured in and these estimates are highly inflated, showing that genes influence a trait more than they supposedly do.
Twin studies, and along with it, heritability estimates, are useless for figuring out, and describing, trait variation in humans. The developmental system is more complex than the genetic reductionists (behavior geneticists) would like one to believe. The reductionist model has been heavily attacked in recent years (Regenmortal, 2004; Noble, 2008, 2012, 2015, 2016; Joyner, 2011, b; Joyner and Pederson, 2011).
Nature vs nurture has also shown to be a false dichotomy because the system develops in whichever environment it finds itself in (Oyama, 1985, 1999; 2000; Moore, 2002; Schneider, 2007)
Conclusion
Since the genetic reductionist model is wrong, along with heritability estimates (because of the nature/nurture fallacy), both should be discontinued. One of the main vehicles of these two models—twin studies—should also be discontinued. These fatal flaws of the behavior geneticists’ paradigm should be enough to discontinue these techniques in the study of human development and behavior. Heritability estimates give no causal information and they also use an outdated model of the gene; twin studies assume too many things for it to be a viable model in the discovering how traits manifest (most importantly, twin studies keep the nature/nurture fallacy alive and should be discontinued on that note only, in my opinion); and genetic reductionist models have been shown to be fatally flawed in recent years. We now have a better understanding of what a gene is today (Portin and Wilkins, 2017), and due to this, we should discontinue whatever implies the fallacy of nature vs nurture because it is irrelevant and a false dichotomy. That, alone, should be enough to discontinue twin studies and heritability estimates.
