
Home » Posts tagged 'Race Realism' (Page 2)
Tag Archives: Race Realism
Races and Populations: Existence and Reality
3000 words
Over at the blog Anthropology 365 the author—Adam Johnson, biocultural anthropologist—wrote an article titled Populations, Race, and The Sorites Paradox, in which he argues that, since there are no “clear lines” and they are “wuzzy”, we cannot say where one race ends and another begins, therefore race does not exist. His whole argument is largely just the continuum fallacy—that since we cannot show where one race, in this instance, ends and another begins, therefore, race does not exist. This reasoning, however, is very flawed.
The beginning of his article is concerned with laying out the sorites paradox. Imagine zero grains of sand, then continuously add grains of sand, 1, 5, 10, 100, 1000, etc. When does the heap become a pile of sand? Johnson attempts to use this logic regarding races and populations: where does one population end and another begin? (You already know where this is headed; it seems that this is the ‘argument’ that gets the most play nowadays when it comes to race-denialism and racial eliminativism when there are better, non-fallacious, arguments out there to attack the concept of race in our ontology. Using the old and tired “continuum fallacy” no longer makes sense because the objection that “Race does not exist because we cannot tell where one race ends and another begins” has been responded to numerous times, most recently (and forcefully) by philosophers of race Michael Hardimon and Quayshawn Spencer.)
He defines “population”, stating that—in biocultural anthropology—that a population is simply a group of like kinds that interbreed with each other which are separated by geographic barriers. Nothing wrong with that—it’s true. He then makes the huge leap in logic to a within-country comparison (America), showing two arbitrarily circled “populations” on the east and west coasts of America. He admits the circles are “arbitrary”, then adds another purple circle in the middle, and finally a green and purple circle in between the original circles, signifying five populations (the image can be seen below).

He says that “It is often impossible to draw neat boundaries around a group”, but I am aware of no author making any claim that it IS possible (and easy) to draw neat boundaries around groups. To do so, you only need simple conditions; and if there is any deviation out of those conditions, then the population in question do not fit the definition of what you were constructing and they can thus be removed. Johnson says “where does yellow end and purple begin?” since there is so much overlap between all five colors in this image. He says that this reasoning shows how “crude” the concept of population is regarding the accepted definition: a group of like kinds that can interbreed but are geographically separated.
One who denies Hardimon’s (2017) 3 conditions for to establish that populations are minimalist races (C1. visible patterns of distinct physical features which correspond to geographic ancestry; C2. that the members in this group are linked by a common ancestry; and C3. they must originate from a distinct geographic location) may then take to this idea that these arbitrarily drawn circles which are supposed to be “populations” (to Johnson) are then races; but Johnson never left any conditions, only a vague definition. One could argue that two of those clusters satisfy C1-C3 (that the cluster in question shares visible patterns of distinct physical features which correspond to geographic ancestry [the people who, say, make up one town in one of the arbitrarily drawn circles may have different visible patterns of distinct physical features which correspond with their ‘geographic ancestry’], that the members are linked by a common ancestry [the town they now live in, say], and they derive from a distinct geographic location [the arbitrarily drawn circle is a distinct geographic location].
However, for one to say that C1 holds for these arbitrarily drawn circles, they have to stretch the definition in order to accept random populations within a country. They then need to say that C2 refers to any type of “common ancestry” of a certain town; and that C3 then shows that they derive from a distinct geographic location. However, in regard to C2 and C3, one who would attempt such an argument would be equivocating on “geographic ancestry” and “distinct geographic location”, thusly claiming that an infinitude of races exist because the conditions are vague. While I do admit that minimalist concept is vague, in my view, it does not allow for one to equivocate on certain words used in the argument to show that any and all arbitrary populations can be called “races”; it does not work like that because there are distinctive conditions that must be met before further thinking on whether or not a population in question is a “race” or not.
Johnson then quotes Scientific American writer John Terrel who writes in his article “Plug and Play” Genetics, Racial Migrations and Human History:
“Distinguishing between races and populations is effectively making a distinction without a difference. If this comes across as sounding crazy to you, then tell me this. What is a population? How can you tell whether you are “inside” a population or “outside” it? How many of them are there “out there” in the real world? How many did there used to be? More than today, or fewer? (Now substitute in these simple questions the word “race.” Doesn’t make much difference, right?)”
What is a population? Good question. The definition left by Johnson above is alright, but we can refine it. I can simply cite Michael Hardimon’s definition of “populationist race” (Hardimon, 2017: 99; my emphasis):
“A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.”
Using this definition of race, a race is a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the groups’ geographic ancestry. Thus, with “population” having a much more non-vague definition, we can then begin to look for populations that exist in reality (not arbitrarily demarcated “populations” like Johnson did—using arbitrary circles as population groups in America).
Now that population is defined, what about the next question: “How can you tell whether you are “inside” a population or “outside” it?” Since we now have a better grasp of what “population” means in this context, then this question is simple to answer. You can tell whether you are “inside”‘ or “outside” a population by looking in a mirror and then thinking about any “population” as defined above. It really is that simple. However, it is hard when “population” is defined so vaguely, and so you get flaws in reasoning like the one from Johnson.
Now that we know that we can tell whether or not we are “inside” or “outside” a population, his next question is: “How many of them are there “out there” in the real world?” According to the definition presented by Hardimon above, there are 5 current races in the human subspecies. That’s the number of races that are ““out there” in the real world” (as opposed to a possible world we can imagine—which is not the topic of contention).
Now that we know how many of “them” [races] exist, the next questions are: “How many did there used to be? More than today, or fewer?” I won’t pretend to know the answer to this question, but I will say one thing: the number of races that used to exist in the past comes down to the number of populations that exhibit a distinctive pattern of visible physical features which are genetically transmitted by geographically and reproductively isolated founding populations. Though, the number of races that “used to” exist is irrelevant to the fact that races exist today and the number of races that do exist today.
Johnson then claims that we, in the West, have a “long history” of constructing different races. And while this is true, this does not go against the claim that biological racial realism is true. Johnson says that “We homogenized entire continents of people into essential “types” and used the assumptions intrinsic to those types to make grand statements about the “natural” divisions in the human species and the value and meaning associated.” Well, these “entire homogenized continents of people” DO fit into “types”—though they are not “essential”; there are “natural” divisions within the human species BUT one does not have to put value and meaning onto the existence of these populations that we call ‘races’, since they are based solely on distinct pattern of genetically transmitted characters which then correspond with the group’s geographic ancestry.
“Anthropology has since moved on from it’s [sic] assumption that the human species is divided up into natural kinds“, Johnson writes. It seems that Johnson is ignorant to the work of Hardimon (2017) and his racial typology using the minimalist concept of race along with its “scientific equivalent” the populationist race concept. Minimalist races are a biological kind “if only a modest one” (Hardimon, 2017: 91), and so, just because “Anthropology has since moved on from it’s [sic] assumption that the human species is divided up into natural kinds” DOES NOT MEAN THAT there are no “kinds” within the human species. The argument for the existence of minimalist races establishes the claim that the human species is, in fact, divided up into kinds:
P1) There are differences in patterns of visible physical features which correspond to geographic ancestry
P2) These patterns are exhibited between real groups, existing groups (i.e., individuals who share common ancestry)
P3) These real, existing groups that exhibit these physical patterns by geographic ancestry satisfy conditions of minimalist race
C) Therefore race exists and is a biological reality
Minimalist races exist and are biologically real; if minimalist races exist, then populationist races exist; populationist race is the “scientization” of minimalist race; minimalist races entail kinds, and so since minimalist races entail kinds then so do populationist races; therefore both concepts speak to kinds within the human species and their biological reality.
Either way, we can also accept that anthropology has moved away from the assumption that the human race is divided into kinds and not have to give up the argument for the existence of race. Instead of arguing that human races are “kinds” as Hardimon (2017) does, Spencer (2014) argues that since Americans defer to the US Census Bureau regarding race, the must be referring to biologically real groups. The US Census Bureau defers to the Office of Management and Budget. The OMB discusses “sets of” populations. K= 5 delineates populations that Americans refer to when referring to race. So since Americans defer to the Census Bureau and the Census Bureau defers to the OMB, when we Americans talk about race, we talk about proper names for population groups as denoted by the OMB—even though ‘race’ looks like a ‘kind’ term, according to Spencer (2014: 1028) “its current use in US racial discourse is that of a proper name. It is a term that rigidly designates a particular set of “population groups.” This means that race is a particular, not a kind.”
So, there are two sound arguments for the existence of race (the argument for the existence of populationist races from Hardimon and the argument for the existence of Blumenbachian partitions—which both use the same population genetics paper (Rosenberg et al, 2002) to buttress their claims that their “kinds” (Hardimon, 2017) and “partitions” (Spencer, 2014) exist in reality.
Lastly, Johnson cites Galanter et al (2012) who genotyped “populations” throughout South America:
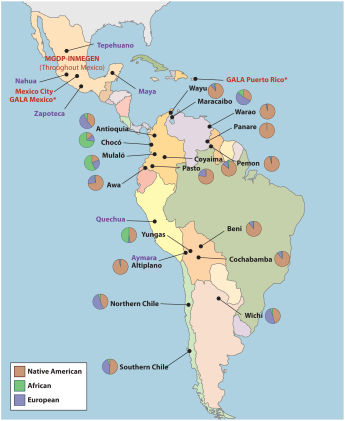
He then states that we have a bunch of South American populations here, all with differing amounts of admixture (which, of course, coincide with three of the five populationist races). He pretty much says, “How can we draw neat circles around these populations to call them “populations”, and what about those other populations not sampled in the analysis?” It makes no sense; when you’re just drawing circles anywhere on any map and then claiming that they are “populations” that satisfy a vague criteria/definition, then you don’t understand any of the newer arguments put forth by philosophers on the existence and reality of racial population groups.
He concludes the article simply:
To conclude, it’s always important to parse in our assumptions and take into account that our levels of analysis (the unit we are studying) may not represent reality. When we equivocate levels of analysis with levels of reality when examining human diversity, as Terrell says, we end up making a distinction between race and populations with no real difference. However, if we understand that the “population(s)” of interest are not reflections of reality, but merely constructed entities that represents an amalgamated web of kinship, political, biological, economic, and random histories at a particular time and place, we can avoid the trap of racial thinking (without using ‘race’) that some scholars fall in to.
He seems to be conflating two concepts here: how we view these visible physical features which correspond to geographic ancestry (our socialview of these populations) and their actual existence completely removed from our social conventions. Yes, socialraces are groups that are taken to be racialist races (that is to say, they are taken to have a specific essence particular to that race and only that race); but the concept of socialrace—the types of social values we give to these populations (think that the minimalist concept of race denotes certain social groups on the basis of distinct visible patterns which correspond to geographic ancestry; the socialrace concept is a good concept since it presents a way of thinking about (1) social groups that are taken to be races (such as ‘Latinos’/’Hispanics’); (2) the social positions that the social groups occupy; and (3) the systems of social structure of which those positions are parts (Hardimon, 2017: 139).
The “populations of interest”, are, indeed, of interest because they pick out what ‘we already know to be’ races.
Races, then, are both socially and biologically constructed. The minimalist concept of race shows the phenotypes that the socialrace concept chooses out when denoting a population its socialrace status in a given society. It shows that there are both biological and social underpinnings to racial categories—that is, there is both a “biological” and “social” realm to race in our ontology, and if we want to understand both ontologies, then we must first think of the consequences of thinking of “race” as only a biological concept and only a social concept and then—after we have thought of “race” as a biological and social concept on its own—we can think of “race” as both a social and biological phenomenon because that’s the best way to describe race in out ontology.
I find it funny how Johnson brings up “population thinking”; but I am probably thinking of it in a different way then he was in his article. When he brings up “population thinking” he wants you to think in terms of his definition of “population”, which pretty much means any group he circles is deemed a population, and thus, since there is no easy way to delineate populations from each other, therefore race does not exist (we must be eliminativist about race). Though when I think of the term “population thinking”, I think of Ernst May’s use of the phrase populationist thinking is more apt: “populationist thinking” is directly opposed to “typological thinking”: “populationist thinking” holds that there are no intrinsic “biological essences”, nor any property—or set of properties—that all, and only all, members of a population share.
For the populationist “all organisms and organic phenomena are composed of unique features and can be described in collectively only in statistical terms. Individuals, or any kind of biological entities, form populations of which we can determine the artihmetic mean and the statistics of variation. Averages are merely statistical abstractions. . . . For the typologist the type (eidos) is real and the variation is an illusion, while for the populationist the type (average is an abstraction and only the variation is real (Mayr, 1976; quoted in Hardimon, 2017: 20).
For example, “Caucasian” is a valid taxonomic category when discussing populationist races. One classified as “Caucasian” might have absolutely none of the genotypic or phenotypic markers associated with “Caucasian-ness”; that is, population thinking does not assume that any one genotype or phenotype is essential to any one population. Thus, there are no intrinsic properties that all members of a race—and only members of that race—share.
To conclude, contrary to the claims of Johnson and Terrel, race does exist and there are reasons why we should accept the existence of these population groups we call races. Johnson largely uses the old and tired continuum fallacy—the fallacy of the beard, whichever name you like—to attempt to argue that “race” does not exist. But he did not even state any conditions on what “population” entails; he just drew random, overlapping circles proclaiming “Ha! Where does X color end and Y color begin!!??” This type of thinking, though, is fallacious, as can be seen. It is completely possible to delinate races on the basis of visible physical features which correspond to geographic ancestry.
Articles like Johnson’s and Terrel’s are easy to come by: they just adopt a racial eliminativist stance on race (that it should be removed from our ontology entirely). They use fallacies like the continuum fallacy to show that since there is no clear ‘genetic line’ (see my article You Don’t Need Genes to Delineate Race) separating so-called races, therefore races do not exist (we must then take an eliminativist approach to race). I’m of the belief that the answer to the question “Does race exist?” will be—and only can be—answered by philosophers of race. We know that geographic variation exists—however small it may be. We know that we can distinguish continental populations on the basis of visible physical features. From there, it’s only a short bit of reasoning to reason, correctly, that race exists and is a biological reality (as the arguments in Spencer, 2014 and Hardimon, 2017 attest to).
The Existence and Reality of Populationist Races
2300 words
Michael Hardimon has some of the best defenses of the reality of race that I am aware of. His 4 concepts are: the racialist concept (he says racialist races do not exist, which I will cover in the future), the minimalist race concept, the socialrace concept (which also will be covered more in depth in the future) and the populationist race concept. Racialist races do not exist, according to Hardimon. However, that does not mean that race does not exist nor does it mean that race isn’t real. On the contrary, race exists and is a biological reality. Simple arguments for the existence of race do indeed exist and see where mixed-race individuals, ‘Latinos’, and Brazilians fall. (Author of the book A Theory of Race Joshua Glasgow also reviewed Hardimon’s book (Glasgow, 2018), and I also left my thoughts on his review.)
Now, minimalist races exist and are biologically real. The concept, though, is vague. It doesn’t state which populations are races, but the populationist race concept, however, does. Hardimon (2017: 99) defines populationist races:
“A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.”
Are there groups that exhibit patterns of a distinctive pattern of visible physical features which are genetically transmitted and correspond to the group’s geographic ancestry? Are there groups that belong to a biological line of descent which was initiated by geographically and reproductively isolated founding populations? The answer is, obviously, yes. Which groups satisfy the definition of populationist races? I will discuss this below.
An important question to answer is: are races subspecies? The two terms are similar. Merriam Webster defines subspecies as: “a category in biological classification that ranks immediately below a species and designates a population of a particular geographic region genetically distinguishable from other such populations of the same species and capable of interbreeding successfully with them where its range overlaps theirs.” While “race” is similarly defined. So, are races subspecies?
The fixation index (Fst) is a measure of population differentiation due to genetic structure which is estimated from SNPs or microsattelites. Generally, the accepted criterion for subspeciation is between .25 and .30. Human groups have an Fst between .05 and .15, so human groups fall way short of subspeciation. Fst estimates for humans fall between .05 and .15, which is far and away what the consensus is on the delineation of subspecies within a group of like kinds. Further, Fst does not support the existence of distinct clusters in humans (Maglo, Mersha, and Martin, 2016; it should be noted that they believe that for human races to exist, human races must be subspecies—similar views are held by philosopher of science Adam Hochman—but their contentions were addressed by Spencer, 2015). Human populations are not subspecies, and the fact that they are not subspecies does not rail against the existence of populationist races.
Hochman (2013) makes the case that in order to claim that clusters represent subspecies, four conditions have to be met: “(i) the range of allele frequency differences between genetic Fstclusters corresponding to race must be relatively uniform, (ii) there must be a determinate number of such clusters, (iii) the allelic frequencies within such clusters must be relatively homogeneous, and (iv) there must be a large jump in genetic differences between such clusters” (Hardimon, 2017: 108).
Thus, the human species does not contain subspecies in the technical sense of the word, as humans Fst estimates range between .05 to .15. This further attests to the fact that the clusters—identified by Rosenberg et al (2002)—are not subspecies. “There is no need for US racial groups to be subspecies or clades, have high genetic variation among them, or be fundamental categories in human population genetics just in order to be biologically real races. Rather, in order for US racial groups to be biologically real races, they just need to be races and biologically real (Spencer, 2015: 6).
The populationist race concept, however, does not require that a division in a species be represented by a particular Fst estimated. It further doesn’t say that Hochman’s (2013) conditions must be met in order for the clusters to be races. Therefore the populationist race concept is not a subspecies concept; there are no subspecies in our genus. Though, if we were forced to accept Hochman’s (2013) conditions (which we do not have to), human races do not exist.
Next is the concept of phylogeny. If phylogenetic is taken to in the normal biological terminology, then the question is whether or not racial lines of descent capture evolutionary significant relationships. And if “evolutionary significant relationships” are taken in the normal biological context then the answer to the question is “no.” This is because the term “evolutionary significance”, taken in the general biological terminology, is understood in a way that for a relationship between populations to be “evolutionarily significant”, then the differences between these populations must be blocked by extensive gene flow.
However, regarding the populations that we take to be populationist races, if the features of these races have adaptive significance, such as skin color for differing climates, then the populationist race concept is of interest to evolutionary biologists since biological raciation makes it possible for divisions of Homo sapiens to survive in different climates. Thus, when discussing how and why divisions of our species adapted to different climates—physically speaking—then this concept is of use to evolutionary biologists since it can explain the adaptive physical features of divisions of Homo sapiens. We then have two choices. We can then further take the idea that to be “phylogenetic”, populations must block extensive gene flow, though we can grant that populationist races may well be of interest to evolutionary biologists (due to their adaptive features that arose due to climatic adaption), despite the fact that populationist races are nonphylogenetic (Hardimon, 2017: 111).
The populationist race concept is a candidate scientific concept. This is because the concept uses biological terminology such as “reproductive isolation”, “transmitted phenotypic characteristics”, “founding population”, and “geographic ancestry.” Hardimon then discusses how and why the concept can form a scientific concept:
“… concept C has the “form” of a scientific concept in biology if
(i) it is formulated in a “biological vocabulary”,
(ii) it is framed in terms of an accepted biological outlook,
(iii) it is suitable for deployment in an accepted branch of biological inquiry, and
(iv) it presents the scientific ground of the phenomenon it represents” (Hardimon, 2017: 112).
This concept satisfies all four conditions. It satisfies (i) since it uses biological vocabulary (e.g., phenotype, reproductive isolation). It satisfies (ii) since it’s framed in what Mayr terms “population thinking” (which is the rejection of essentialism—“the view that some properties of objects are essential to them.”. It satisfies (iii) since it is suitable for deployment in ecology, ethology and evolutionary biology. Areas of study, for example, can focus on how and why differing populationist races have differing patterns of visible physical features (i.e., how and why phenotypes changed as migration occurred out of Africa into Eurasia, the Pacific Islands and the Americas). And it satisfies (iv) in that representing populationist races as having arisen from reproducively isolated founding populations.
Now which groups are candidates for populationist races? There are two conditions: (1) they exhibit distinctive patterns of phenotypic characters which correspond to that population’s geographic ancestry and (2) belong to biological lines of descent which then trace back to geographically separated and reproductively isolated founding populations.
There are populations which exhibit distinctive patterns of visible physical features which correspond to geographic ancestry, and they are Sub-Saharan Africans, Caucasians, East Asians, Native Americans and Pacific Islanders. The distinctive patterns of visible physical features are genetically transmitted, and they correspond to geographic ancestry. These populations belong to biological lines of descent which can then be traced back to geographically separated and reproductively isolated founding populations. Thus, conditions (1) and (2) are satisfied, therefore populationist races exist.
Further support for (iii) (that the populationist race concept can be deployed in the biological sciences) can be found in my article You Don’t Need Genes to Delineate Race. I discussed differences in gross morphology between the races; I discussed differences in physiognomy between the races; and, of course, the differences in geographic ancestry that caused the differences in morphology and physiognomy (see here for discussions on skin color). Differences in climate that Homo sapiens encountered after trekking out of Africa then caused the distinctive differences in visible physical features which correspond with geographic ancestry which then make up populationist races. Thus, the study of populationist races will elucidate the caused of phenotypic differences between populationist races since they exist and are a biological reality.
There is a relationship between populationist and minimalist races, though they’re defined by different concepts. However if minimalist races are populationist races, then the kind minimalist race=populationist race. “The claim that minimalist race=populationist race is analogous to the claim that water=H2O. The latter claim, since true, provides scientific insight into the nature of minimalist race” (Hardimon, 2017: 120).
Furthermore, we can assume that the populations identified by Lewontin (1972) as races can be interpreted as lending support to the biological reality of populationist races exist. We can also assume that African, Caucasians, East Asians, Oceanians, and Native Americans constitute populationist races, then Rosenberg et al (2002) show support for the biological reality of populationist races, even though the fraction of diversity separating the clusters is between 3-5 percent, this still shows that populationist races capture a portion of biological human variation, no matter how small it is.
“If it is assumed that Africans, Eurasians, East Asians, Oceanians, and Americans constitute continental-level populationist races, Rosenberg and colleagues’ 2002 study can be interpreted as providing support for the biological reality of populationist race inasmuch as it shows that a very small fraction (3-5 percent) of human genetic variation is due to difference among continental-level populationist races. Modulo our assumption, the study results indicate that populationist race is a minor principle of human genetic structure and that populationist race is a minor principle of human variation.” (Hardimon, 2017: 124)
The same points made that minimalist races are human population partitions, that races can be distinguished at the level of the gene, and that the continental-level minimalist races differ in a small number of coding genes, also carry over to the populationist race concept since minimalist race=populationist race, so the biological reality of minimalist race carry over to populationist race. So if the five populations are populationist races, then populationist race correspond to a partition of genetic variation found between the races in the human species, which is then evidence for the existence of populationist races.
The five populations that make up populationist races are Native Americans, Caucasians, East Asians, Pacific Islanders, and Sub-Saharan Africans. These populations are biologically real, and they exist. They generically transmit phenotypic characteristics across the generations; these phenotypic characteristics differ due to geographic ancestry. These populations are identified in numerous K = 5 runs. So if we assume that the five populations are populationist races then K = 5 shows the real, but small, human genetic variation found within continental-level populationist races which is how the visible patterns of visible physical features which correspond to geographic ancestry are genetically transmitted.
The populationist race concept is a candidate scientific concept. This is a way to study the small genetic variation between the continental-level clusters. Human phenotypic (and physiologic) differences arose due to adaption to different climates. Thus, since populationist race is a biological reality then studying populationist races will better elucidate how and why differences in phenotype arose.
Both the populationist and minimalist race concepts are vague, I admit. However, they’re not so vague that one could argue that villages, countrys, social classes etc are populationist races. It should be noted, though, that it is implicitly stated in the definition for populationist race, that a morphological component exists. Therefore, groups like the Amish, social classes etc. Thus, the populationist race concept gaurentees that races will be races in the ordinary sense of the word (see Hardimon, 2003). So we can take two groups—G1 and G2—and if G1 does not have any pattern of visible physical features which distinguish it from another group, G2, then G1 is not a race. These visible physical differences that distinguish races from one another are biological in nature—hair color/type, skin color, eye type, morphology etc. This gaurentees that different villages, countries, economic classes and ethnies within a race are not counted as “races”, so defined.
The thing about the populationist race concept is that it directly relates to the minimalist race concept. Once we acknowledge that races exist and are real (since minimalist races exist and are real), then we start thinking “Which populations sastisfy the conditions of populationist races?” The populationist race concept—in tandem with the minimalist race concept—shows us that the patterns in visible physical features are genetically transmitted characters which which correspond to the population’s geoprahic ancestry who belong to biological lines of descent which were initiated by geographically separated and genetically isolated founding populations. The populationist race concept supports the claim that the minimalist race concept is a biological concept and secures the existence of minimalist races since minimalist race=populationist race.
P1) The five populations demarcated by Rosenberg et al (2002) are populationist races; K = 5 demarcates populationist races.
P2) Populationist race=minimalist race.
P3) If populationist race=minimalist race, then everything from showing that minimalist races are a biological reality carrys over to populationist races.
P4) Populationist races capture differences in genetic variation between continents and this genetic variation is responsible for the distinctive patterns of visible physical features which correspond to geographic ancestry who belong to biological lines of descent which were initiated by geographically isolated founding populations.
C) Therefore, since populationist races=minmalist races, and visible physical features which correspond to geographic ancestry are genetically transmitted by populations who belong to biological lines of descent, initiated by reproductively isolated founding populations, then populationist races exist and are biologically real.
The Hierarchical Nature of Living Systems, Species, and Race
2100 words
Biology is one of the most interesting sciences since, at its core, it is the study of life and living systems. The biological organization of living systems and the ecosystems these living systems find themselves in are interesting to learn about, since we can then discern different species and learn how and when to delineate separate species based on a set of pre-conceived measures. The classification of human races in these systems will be discussed, along with why human races are not different species.
The organization of living systems
Living systems show hierarchical organization, each system—from the physiological to the physical—interacting with each other. However, a key factor in the organization of these interactions is the degree of the complexity of the interactions in question. We can look at the organization of the biological world as hierarchical—that is, each level builds on the preceding level, so we get from atoms to the biosphere and everything in between is what we call “life” and also show how these complex, living biological systems live and exist due to the hierarchical organization of living systems. The point is, life does not have a simple definition, but all living systems share similar characteristics that can describe life. Biologists organize living systems hierarchically, from the subcellular level to the entire biosphere, and then study the interactions that occur which cannot be predicted from just studying the sum of its parts. This is why a holistic—and not reductionistic—approach needs to be taken when studying and describing living systems.
The hierarchy is:
The cellular level, which includes: atoms, molecules, macromolecules, and organelles; the organismal level which include: tissue, organs, the organ system, and the organism; the populational level which includes: the population, species, and the community; and finally the highest level, the ecosystem level which includes the ecosystem and the biosphere.
At the cellular level, we have atoms which are the fundamental elements of matter and are joined together by chemical bonds called molecules. large and complex molecules are called macromolecules, DNA—which stores hereditary information—is a type of macromolecule. Complex biological molecules are then assembled into organelles, where cellular activities are organized. A mitochondrion is, for example, an organelle with a cell that extracted energy from consumed food molecules. And finally, we have cells, which are the basic unit of life.
Next, we have the organismal level, and cells of multicellular organisms make up three levels of organization. Tissues, which are groups of similar cells which function together as a unit. Tissues then are grouped into organs which are structures of the body which are composed of many different kinds of tissues which act in a structural manner and as a unit. Then we have organ systems, such as the nervous system which is the sensory organs, brain and spinal cord, and the network of neurons that convey signals to different parts of the body.
Then we have the populational level. This includes the individual organisms which occupy various hierarchical levels in the biological world. A population is a group of organisms all living in the same place. Together, all populations of a particular kind form a species—members of a species must look similar and be able to interbreed. Then finally, we have the biological community which consists of all of the populations coexisting together in one place.
Lastly, we have the ecosystem level. This is the highest tier of biological organization (the lowest being the cellular level). A biological community and its physical habitat (such as soil composition, available water etc) in which it finds itself in and lives and competes with other organisms constitute an ecosystem while the entire planet is the highest of all levels of biological organization—the biosphere. All of these systems together can be seen as the hierarchical organization of living systems.
(See Mason et al, 2018 for more discussion of the above points.)
Organismal classification
Now, in these differing biological hierarchies, we find differing Eukarya, Prokarya, and Bacteria. The in-use classification system is the Linnean hierarchy. Differences exist between organisms, this is obvious. But it is a bit more tricky to classify these organisms and place them into like groups. Then, in the 1750s, Carolus Linnaeus came along and instituted a binomial classification system for organisms—the most commonly-known binomial being Homo sapiens—which was much simpler than the polynomial names
The hierarchy is as follows:
1. Species;
2. Genus;
3. Family;
4. Order;
5. Class;
6. Phylum;
7. Kingdom; and
8. Domain. Domains can then be split into Archaea, Bacteria, and Eukarya. Domains are the largest taxons, being that they comprise every organism that we know of.
For example, our species is sapiens, our genus is Homo, our family is Hominidae, our order is primates, our class is Mammalia, our phylum is Chordata (with a subphylum Craniata), our kingdom is Animalia and our domain is Eukarya. This is our species’ taxonomic classification.
The traditional classification system—the Linnean system—groups species into genera, families, orders, classes, phyla, and kingdoms. Thus, these systems classify different organisms on the basis of similar traits, and since they consist of a mix of derived and ancestral traits, they do not necessarily take into account different evolutionary relationships.
There are of course limitations to the Linnean hierarchy:
1) Many “higher” taxonomic ranks are not monophyletic and so do not represent real groups (like Reptilia). For something to be a “natural group”, a common ancestor and its descendants must all derive from descent from a common ancestor, so any other type of taxonomic ranks are created by taxonomists, such as paraphyletic and polyphyletic.
2) Linnean ranks are not equivalent. Two families may not represent clades that arose at the same time, because one family may have diverged millions of years before the other family and so the two families had differing amounts of time to diverge and acquire new traits. So comparisons in the Linnean sense may be misleading and we should then use hypotheses of phylogenetic relationships.
What is a species?
It should first be noted that species are, indeed, real. New species arise when isolated organisms of one population become genetically/geographically isolated for a period of time. Over time, as the split population spends time geographically and genetically isolated, they cannot interbreed with the parent population and thusly attain separate species status. This is the received view, the biological species concept.
There are a wide range of species concepts and they all capture the differences that different theorists believe we should emphasize in our classification of organisms.
The phenetic species which appeal to the intrinsic similarities of organisms. The biological species concept which appeals to reproductive isolation (one version of the biological species concept is the recognition concept, which defines species as a system of mating recognition. The cohesion species concept which generalizes the biological species concept and it recognizes that gene flow isn’t the only factor that holds a population together and makes it different from other populations. The ecological species concept which defines species by appealing to the fact that members of a species are in competition with one another because of the need the same resources. And the phylogenetic and evolutionary species concept which define species as segments on the tree of life (the phylogenetic species concept, for instance, holds the term ‘species’ should be reserved for groups of populations that have been evolving independently of other populations.
Sterelny and Griffiths (1999) tackled this in their book Sex and Death: An Introduction to Philosophy of Biology:
While we think cladism presents the best view of systematics, biological classification nevertheless poses an unsolved problem. If we were to accept either evolutionary taxonomy, which builds disparity into its classification system, or phenetic taxonomy, which is based on the idea of nested levels of similarity, traditonal taxonomic levels would be quite defensible. Within those taxonomic pictures, the idea of genus, family, order, and so on makes quite good sense. If cladism is the only defensible picture of systematics, the situation is more troubling. From that perspective, these taxonomic ranks make little sense. Cladists do not think there is a well-defined objective notion of the amount of evolutionary divergence. That, in part, is why they are cladists. Hence, they do not think there will be any robust answer to the questions, when should we call a monophyletic group of species a genus? a family? an order? Only monophyletic groups should be called anything, for they are well-defined chucnks of the tree. But only science greets the question, are the chimps plus humans a genus? It has long been receieved wisdom in taxonomy that there is something arbitrary about taxonomic classification above species. These decisions are judgement calls. So cladists only show a somewhat more extreme version of a skepticism that has long existed. The problem of high taxonomic ranks would not matter except for the importance of the information expressed using them. Hence cladism reinforces the worry that when, for example, we consider divergent extinction and survival patterns, our data may not be tobust, for our units may not be commensurable. Unfortunately, it does this without suggesting much of a cure.
Where does race fit in?
Racehood is simple: A race is a group of humans that: Condition 1; is distinguished from other groups of humans by patterns of visible physical features; Condition 2: is linked by common geographic ancestry which is peculiar to members of this group; and Condition 3: originates from a distinctive geographic location.
So now all we need to do is go through four steps: 1) recognize that there are patterns of visible physical features which correspond to geographic ancestry; 2) observe that these patterns of visible physical features which correspond to geographic ancestry are exhibited between real, existing groups; 3) note that these real existing groups that exhibit these patterns by geographic ancestry satisfy C1-C3; and 4) infer that race exists.
Some may argue that the races are different species, citing the same patterns of visible physical features discussed above. However, if we are referring to the biological species concept, then the human races are not different species at all since all human races can produce fertile offspring with one another. Our genus, of course, is Homo, all of the human races are the same genus; though some may attempt to use the previously-discussed conditions for racehood as conditions for specieshood for humans, the most preferred method for delineating species currently is the biological species concept, and since all of the human races can produce fertile offspring then the human races are not different species.
In keeping with the classification system that is currently used today (see above), where would human races fit into our taxonomy? Falling within our species sapiens seems like a good start, and since the races can interbreed and have fertile offspring, then they are not different species but are the same species, despite phenotypic differences. Thus, human races would be within species but under subspecies. Using this line of logic, human races cannot be different species, despite claims to the contrary that human races are different species based on patterns of visible physical features which correspond to geographic ancestry. That’s enough to denote racehood, not specieshood.
Conclusion
The study of life—in all of its forms and in all of its environments—is one of the most important things we, as humans, can do. From it, we can learn where we came from and even—possibly—where we may be going. Once we understood the biological hierarchy and how upper levels are built from lower levels working together, then we were better able to understand how living systems act on the inside—cellularly and physiologically—to the outside—organismal and environmental interaction. From organismal and environmental interaction, speciation may occur. The highest level of the organization of living systems is the biosphere—and it is so because the living systems that are driven by the smallest cellular interactions interact with other species, the ecosystem and the biosphere.
Species do exist, but there are numerous species concepts—over twenty. One of the more popular species concepts in use is the cladistic species concept. In this species concept, a species is a lineage of populations between two specific branch points. The cladistic concept thusly recognizes differing species by differing branch points and how much change occurs between them (see Ridley, 1989).
The classification of different organisms into different species is pretty straightforward, though it falls prey to oversimplification since it only focuses on similar traits. Species exist, this is established. But races are not species, contrary to some beliefs. Different races can interbreed and, I would argue, that for there to be separate species, human races would not be able to interbreed. Yes, there are physical and morphological differences between races, but, as argued, this is not enough to denote speciation, but it is enough to denote raciation.
Blumenbachian Partitions and Mimimalist Races
2100 words
Race in the US is tricky. On one hand, we socially construct races. On the other, these socially constructed races have biological underpinnings. Racial constructivists, though, argue that even though biological races are false, races have come into existence—and continue to exist—due to human culture and human decisions (see the SEP). Sound arguments exist for the existence of biological races. Biological races exist, and they are real. One extremely strong view is from philosopher of science Quayshawn Spencer. In his paper A Radical Solution to the Race Problem, Spencer (2014) argues that biological races are real; that the term “race” directly refers; that race denotes proper names, not kinds; and these sets of human populations denoted by Americans can be denoted as a partition of human populations which Spencer (2014) calls “the Blumenbach partition”.
To begin, Spencer (2014) defines “referent”: “If, by using appropriate evidential methods (e.g., controlled experiments), one finds that a term t has a logically inconsistent set of identifying conditions but a robust extension, then it is appropriate to identify the meaning
of t as just its referent.” What he means is that the word “race” is just a referent, which means that the term “race” lies in what points out in the world. So, what “race” points out in the world becomes clear if we look at how Americans define “race”.
Spencer (2014) assumes that “race” in America is the “national meaning” of race. That is, the US meaning of race is just the referent to the Census definitions of race, since race-talk in America is tied to the US Census. But the US Census Bureau defers to the Office of Management and Budget (OMB). Therefore, since the US Census Bureau defers to the OMB on matters of race, and since Americans defer to the US Census Bureau, then Americans use the OMB definitions of race.
The OMB describes a “comprehensive set” of categories (according to the OMB) which lead Spencer (2014) to believe that the OMB statements on race are pinpointing Caucasians, Africans, Pacific Islanders, East Asians, and Amerindians. Spencer (2014: 1028-29) thusly claims that race in America “is a term that rigidly designates a particular set of “population groups.” Now, of course, the question is this: are these population groups socially constructed? Do they really exist? Are the populations identified arbitrary? Of course, the answer is that they identify a biologically real set of population groups.
To prove the existence of his Blumenbachian populations, Spencer (2014) invokes populational genetic analyses. Population geneticists first must make the assumption at how many local populations exist in the target species. According to Spencer, “The current estimate for humans is 7,105 ethnic groups, half of which are in Africa and New Guinea.” After the assumptions are made, the next step is to sample the species’ estimated local populations. Then they must test noncoding DNA sequences. Finally, they must attempt to partition the sample so that each partition at each level is unique which then minimizes genetic differences in parts and maximizes genetic differences among parts. There are two ways of doing this: using structure and PCA. For the purposes of this argument, Spencer (2014) chooses structure, invoking a 5-population racial model, (see e.g., Rosenberg et al, 2002).
K = 5 corresponds to 5 populational clusters which denote Africans, Oceanians, East Asians, Amerindians, and Caucasians (Spencer, 2014; Hardimon, 2017b). K = 5 shows that the populations in question are genetically structured—that is, meaningfully demarcated on the basis of genetic markers and only genetic markers. Thus, that the populations in question are meaningfully demarcated on the basis of genetic markers, this is evidence that Hardimon’s (2017b) minimalist races are a biological reality. Furthermore, since Rosenberg et al (2002) used microsatellite markers in their analysis, this is a nonarbitrary way of constructing genetic clusters which then demarcate the continental-level minimalist races (Hardimon, 2017b: 90).
Thus, Spencer (2014) argues to call the partition identified in K = 5 “the Blumenbachian partition” in honor of Johann Blumenbach, anthropologist, physician, physiologist, and naturalist. (Though it should be noted that one of his races “Malays” was not a race, but Oceaninans are, so he “roughly discovered” the population partition.) So we can say that “the Blumenbach partition” is just the US meaning of “race”, the partitions identified by K = 5 (Rosenberg et al, 2002).
Furthermore, like Lewontin (1972), Rosenberg et al (2002) found that a majority of human genetic variation is between individuals, not races. That is, Rosenberg et al (2002) found that only 4.3 percent of human genetic variation was found to lie between the continental-level minimalist races. Thus, minimalist races are a biological kind, “if only a modest one” (Hardimon, 2017b: 91). Thus, Rosenberg et al (2002) support the contention that minimalist races exist and are a biological reality since a fraction of human population variation is due to differences among continental-level minimalist races (Africans, Caucasians, East Asians, Oceanians, and Amerindians). The old canard is true, there really is more genetic variation within races than between them, but, as can be seen, that does not rail against the reality of race, since that small amount of genetic variation shows that humanity is meaningfully clustered in a genetic sense.
Spencer (2014: 1032) then argues why Blumenbachian populations are “race” in the American sense:
It is not hard to generate accessible possible worlds that support the claim that US race terms are just aliases for Blumenbachian populations. For example, imagine a possible world τ where human history unfolded exactly how it did in our world except that every Caucasian in τ was killed by an infectious disease in the year 2013. Presumably, we have access to τ, since it violates no logical, metaphysical, or scientific principles. Then, given that we use ‘white’ in its national American meaning in our world, and given that we use ‘Caucasian’ in its Blumenbachian meaning in our world, it is fair to say that both ‘Caucasian’ and ‘white’ are empty terms in τ in 2014—which makes perfect sense if ‘white’ is just an alias for Caucasians. It is counterfactual evidence like this that strongly suggests that the US meaning of ‘race’ is just the Blumenbach partition.
Contrary to critics, this partition is biologically real and demarcates the five genetically structured populations of the human race. Rosenberg et al (2005) found that if sufficient data are used, “the geographic distribution of the sampled individuals has little effect on the analysis“, while their results verify that genetic clusters “arise from genuine features of the underlying pattern of human genetic variation, rather than as artifacts of uneven sampling along continuous gradients of allele frequencies.”
Some may claim that K = 5 is “arbitrary”, however, constructing genetic clusters using microsatellites is nonarbitrary (Hardimon, 2017b: 90):
Constructing genetic clusters using microsatellites constitutes a nonarbitrary way of demarcating the boundaries of continental-level minimalist races. And the fact that it is possible to construct genetic clusters corresponding to continental-level minimalist races in a nonarbitrary way is itself a reason for thinking that minimalist race is biologically real 62.
It should also be noted that Hardimon writes in note 62 (2017b: 197):
Just to be perfectly clear, I don’t think that the results of the 2002 Rosenberg article bear on the question: Do minimalist races exist? That’s a question that has to be answered separately. In my view, the fundamental question in the philosophy of race on which the results of this study bear is whether minimalist race is biologically real. My contention is that they indicate that minimalist race (or more precisely, continental-level minimalist race) is biologically real if sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races.
Sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races, therefore race is a biological reality. We can pinpoint them on the basis of patterns of visible physical features; these visible physical features correspond to geographic ancestry; this satisfies the criteria for minimalist races; therefore race exists. Race exists as a biological kind.
Furthermore, if these five populations that Rosenberg et al (2002) identified (the Blumenbachian populations) are minimalist races, then minimalist race is “a minor principle of human genetic structure” (Hardimon, 2017b: 92). Since minimalist races constitute a dimension within the small amount of human genetic variation that is captured between the continental-level minimalist races (4.3 percent), then it is completely possible to talk meaningfully about the racial structure of human genetic variation which consists of the human genetic variation which corresponds to continental-level minimalist races.
Thus, the US meaning of race is just a referent; the US meaning of race refers to a particular set of human populations; races in the US are classically-defined races (Amerindian, Caucasian, African, East Asian, and Oceanians; the Blumenbach partition); and race is both a biological reality as well as socially constructed. These populations are biologically real; if these populations are biologically real, then it stands to reason that biological racial realism is true (Hardimon, 2012 2013, 2017a; 2017b; Spencer, 2014, 2015).
Human races exist, in a minimalist biological sense, and there are 5 human races. Defenders of Rushton’s work—who believed there are only 3 primary races: Caucasoids, Mongoloids, and Negroids (while Amerindians and others were thrown into the “Mongoloid race” and Pacific Islanders being grouped with the “Negroid race” (Rushton, 1988, 1997; see also Liberman, 2001 for a critique of Rushton’s tri-racial views)—are forced into a tri-racial theory, since he used this tri-racial theory as the basis for his, now defunct, r/K selection theory. The tri-racial theory, that there are three primary races of man—Caucasoid, Mongoloid, and Negroid—has fallen out of favor with anthropologists for decades. But what we can see from new findings in population genetics since the sequencing of the human genome, however, is that human populations cluster into five populations and these five populations are races, therefore biological racial realism is true.
Biological racial realism (the fact that race exists as a biological reality) is true, however, just like with Hardimon’s minimalist races, they do not denote “superiority”, “inferiority” for one race over another. Most importantly, Blumenbachian populations do not denote those terms because the genetic evidence that is used to support the Blumenbachian partition use noncoding DNA. (It should also be noted that the terms “superior” and “inferior” are nonsensical, when used outside of their anatomic contexts. The head is the most superior part of the human body, the feet are the most inferior part of the human body. This is the only time these terms make sense, thus, using the terms outside of this context makes no sense.)
It is worth noting that, while Hardimon’s and Spencer’s views on race are similar, there are some differences between their views. Spencer sees “race” as a referent, while Hardimon argues that race has a set descriptive meaning on the basis of C (1)-(3); (C1) that, as a group, is distinguished from other groups of human beings by patterns of visible physical features, (C2) whose members are linked be a common ancestry peculiar to members of that group, and (C3) that originates from a distinctive geographic location” (Hardimon, 2017b: 31). Whether or not one prefers Blumenbachian partitions or minimalist races depends on whether or not one prefers race in a descriptive sense (i.e., Hardimon’s minimalist races) or if the term race in America is a referent to the US Census discourse, which means that “race” refers to the OMB definitions which then denote Blumenbachian partitions.
Hardimon also takes minimalist races to be a biological kind, while Spencer takes them to be a proper name for a set of population groups. Both of these differing viewpoints regarding race, while similar, are different in that one is describing a kind, while the other describes a proper name for a population group; these two views regarding population genetics from these two philosophers are similar, they are talking about the same things and hold the same deflationary views regarding race. They are talking about how race is seen in everyday life and where people get their definitions of “race” from and how they then integrate it into their everyday lives.
“Race” in America is a proper name for a set of human population groups, the five population groups identified by K = 5. Americans defer to the US Census Bureau on race, who defers to the Office of Management and Budget to define race. They hold that races are a “set”, and these “sets” are Oceanians, Caucasians, East Asians, Amerindians, and Africans. Race, thusly, refers to a set of population groups; “race” is not a “kind”, but a proper name for known populational groups. K = 5 then shows us that the demarcated clusters correspond to continental-level minimalist races, what is termed “the Blumenbach partition.” This partition is “race” in the US sense of the term, and it is a biological reality, therefore, like Hardimon’s minimalist races, the Blumenbach partition identifies what we in America know to be race. (It’s worth noting that, obviously, the Blumenbach partition/minimalist races are one in the same, Spencer is a deflationary realist regarding race, just like Hardimon.)
Cold Winter Theory, the Vitamin D Hypothesis and the Prediction of Novel Facts
2400 words
HBDers purport that as one moves further north from Africa that IQ raises as a function of how the population in question needed to survive. The explanation is that as our species migrated out of Africa, more “intelligence” was needed and this is what explains the current IQ disparities across the world: the ancestors of populations evolving in different areas with different demands then changed their “IQs” and this then is responsible for differential national development between nations. Cold winter theory (CWT) explains these disparities.
On the other hand is the vitamin D hypothesis (VDH). The VDH purports to explain why populations have light skin at northern latitudes. As the migration north out of Africa occurred, peoples needed to get progressively lighter in order to synthesize vitamin D. The observation here is that as light skin is selected for in locations where UVB is absent, seasonal or more variable whereas dark skin is selected for where UVB is stronger. So we have two hypotheses: but there is a problem. Only one of these hypotheses makes novel predictions. Predictions of novel predictions are what science truly is. A predicted fact is a novel fact for a hypothesis if it wasn’t used in the construction of the hypothesis (Musgrave, 1988). In this article, I will cover both the CWT and VDH, predictions of facts that each made (or didn’t make) and which can be called “science”.
Cold winter theory
The cold winter theory, formulated by Lynn and Rushton, purports to give an evolutionary explanation for differences in national IQs: certain populations evolved in areas with deathly cold winters in the north, while those who lived in tropical climes had, in comparison to those who evolved in the north, an “easier time to live”. Over time as populations adapted to their environments, differences in ‘intelligence’ (whatever that is) evolved due to the different demands of each environment, or so the HBDers say.
Put simply, the CWT states that IQ differences exist due to different evolutionary pressures. Since our species migrated into cold, novel environments, this was the selective pressure needed for higher levels of ‘intelligence’. On the other hand, humans who remained in Africa and other tropical locations did experience these novel, cold environments and so their ‘intelligence’ stayed at around the same level as it was 70,000 years ago. Many authors hold this theory, including Rushton (1997), Lynn (2006), Hart, (2007) Kanazawa (2008), Rushton and Templer (2012; see my thoughts on their hypothesis here) and Wade (2014). Lynn (2013) even spoke of a “widespreadonsensus” on the CWT, writing:
“There is widespread consensus on this thesis, e.g. Kanazawa (2008), Lynn (1991, 2006), and Templer and Arikawa (2006).”
So this “consensus” seems to be a group of his friends and his own publications. We can change this sentence to ““There is widespread consensus on this thesis, including two of my publications, a paper where the author assumes that the earth is flat: “First, Kanazawa’s (2008) computations of geographic distance used Pythagoras’ theorem and so the paper assumed that the earth is flat (Gelade, 2008).” (Wicherts et al, 2012) and another publication where the authors assume hot weather leads to lower intelligence. Oh yea, they’re all PF members. Weird.” That Lynn (2013) calls this “consensus” is a joke.
What caused higher levels of ‘intelligence’ in those that migrated out of Africa? Well, according to those who push the CWT, finding food and shelter. Kanazawa, Lynn, and Rushton all argue that finding food, making shelter and hunting animals were all harder in Eurasia than in Africa.
One explanation for high IQs of people who evolved recently in northern climes is their brain size. Lynn (2006: 139) cites data showing the average brain sizes of populations, along with the temperatures in that location:
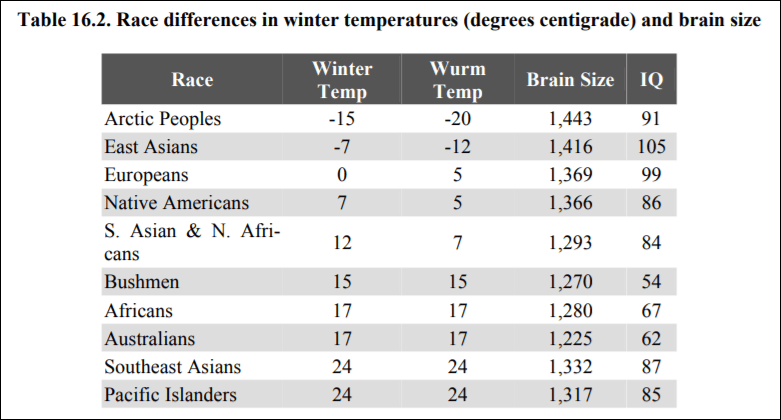
Do note the anomaly with the Arctic peoples. To explain this away in an ad-hoc manner, Lynn (2006: 156-7) writes:
These severe winters would be expected to have acted as a strong selection for increased intelligence, but this evidently failed to occur because their IQ is only 91. The explanation for this must lie in the small numbers of the Arctic Peoples whose population at the end of the twentieth century was only approximately 56,000 as compared with approximately 1.4 billion East Asians.
This is completely ad-hoc. There is no independent verifier for the claim. That the Arcitic don’t have the highest IQs but experienced the harshest temperatures and therefore have the biggest brain size is a huge anomaly, which Lynn (2006) attempts to explain away by population size.
He does not explain why natural selection among Arctic peoples would result in larger brain sizes or enhanced visual memory yet the same evolutionary pressures associated with a cold environment would not also produce higher intelligence. Arctic peoples have clear physical adaptations to the cold, such as short, stocky bodies well-suited to conserving heat.
Furthermore, the argument that Lynn attempts is on the mutations/population size is special pleading—he is ignoring anomalies in his theory that don’t fit it. However, “evolution is not necessary for temperature and IQ to co-vary across geographic space” (Pesta and Poznanski, 2014).
If high ‘intelligence’ is supposedly an adaptation to cold temperatures, then what is the observation that disconfirms a byproduct hypothesis? On the other hand, if ‘intelligence’ is a byproduct, which observation would disconfirm an adaptationist hypothesis? No possible observation can confirm or disconfirm either hypothesis, therefore they are just-so stories. Since a byproduct explanation would explain the same phenomena since byproducts are also inherited, then just saying that ‘intelligence’ is a byproduct of, say, needing larger heads to dissipate heat (Lieberman, 2015). One can make any story they want to fit the data, but if there is no prediction of novel facts then how useful is the hypothesis if it explains the data it purports to explain and only the data it purports to explain?
It is indeed possible to argue that hotter climates need higher levels of intelligence than colder climates, which has been argued in the past (see Anderson, 1991; Graves, 2002; Sternberg, Grigorenko, and Kidd, 2005). Indeed, Sternberg, Grigorenko, and Kidd (2005: 50) write: “post hoc evolutionary arguments … can have the character of ad hoc “just so” stories designed to support, in retrospect, whatever point the author wishes to make about present-day people.” One can think up any “just-so” story to explain any data. But if the “just-so” story doesn’t make any risky predictions of novel facts, then it’s not science, but pseudoscience.
Vitamin D hypothesis
The VDH is simple: those populations that evolved in areas with seasonal, absent, or more variable levels of UVB have lighter skin than populations that evolved in areas with strong UVB levels year-round (Chaplan and Jablonksi, 2009: 458). Robins (2009) is a huge critic of the VDH, though her objections to the VDH have been answered (and will be discussed below).
The VDH is similar to the CWT in that it postulates that the adaptations in question only arose due to migrations out of our ancestral lands. We can see a very strong relationship between high UVB rays and dark skin and conversely with low UVB rays and light skin. Like with the CWT, the VDH has an anomaly and, coincidentally, the anomaly has to do with the same population involved in the CWT anomaly.
Arctic people have dark-ish skin for living in the climate that they do. But since they live in very cold climates then we have a strange anomaly here that needs explaining. We only need to look at the environment around them. They are surrounded by ice. Ice reflects UVB rays. UVB rays hit the skin. Arctic people consume a diet high in vitamin D (from fish). Therefore what explains Arctic skin color is UVB rays bouncing off the ice along with their high vitamin D diet. The sun’s rays are, actually, more dangerous in the snow than on the beach, with UVB rays being 2.5 more times dangerous in the snow than beach.
Evolution in different geographic locations over tens of thousands of years caused skin color differences. Thus, we can expect that, if peoples are out of the conditions where their ancestors evolved their skin color, that there would then be expected complications. For example, if human skin pigmentation is an adaptation to UV rays (Jablonski and Chaplan, 2010), we should expect that, when populations are removed from their ancestral lands and are in new locations with differing levels of UV rays, that there would be a subsequent uptick in diseases caused by vitamin D deficiencies.
This is what we find. We find significant differences in circulating serum vitamin D levels, and these circulating serum vitamin D levels then predict health outcomes in certain populations. This would only be true if sunlight influenced vitamin D production and that skin progressively gets lighter as one moves away from Africa and other tropical locations.
Skin pigmentation regulates vitamin D production (Neer, 1975). This is due to the fact that when UVB rays strike the skin, we synthesize vitamin D, and the lighter one’s skin is, the more vitamin D can be synthesized in areas with fewer UVB rays. (Also see Daraghmeh et al, 2016 for more evidence for the vitamin D hypothesis.)
P1) UV rays generate vitamin D in human skin
P2) Human populations that migrate to climates with less sunlight get fewer UV rays
P3) To produce more vitamin D, the skin needs to get progressively lighter
C) Therefore, what explains human skin variation is climate and UV rays linked to vitamin D production in the skin.
Novel predictions
Science is the generation of novel facts from risky predictions (Musgrave, 1988; Winther, 2009). And so, hypotheses that predict novel facts from risky predictions are scientific hypotheses, whereas those hypotheses that need to continuously backtrack and think up ad-hoc hypotheses are then pseudoscientific. Pseudoscience is simple enough to define. The Stanford Encyclopedia of Philosophy defines it as:
“A pretended or spurious science; a collection of related beliefs about the world mistakenly regarded as being based on scientific method or as having the status that scientific truths now have.”
All theories have a protective belt of ad hoc hypotheses. Theories become pseudoscientific when they fail to make new predictions and must take on more and more ad-hoc hypotheses that have no predictive value. If the ad-hoc hypotheses that are added to the main hypothesis have no predictive value then the new explanations for whichever hypothesis that is in danger of being falsified are just used to save the hypothesis from being refuted and it thus becomes pseudoscience.
In the case of CWT, it makes no prediction of novel facts; it only explains the data that it purports to explain. What is so great about the CWT if it makes no predictions of novel facts and only explains what it purports to explain? One may attempt to argue that it has made some ‘novel’ predictions but the ‘predictions’ that are proposed are not risky at all.
For example, Hart (2007: 417) makes a few “predictions”, but whether or not they’re “risky” or “novel” I’ll let you decide (I think they’re neither, of course). He writes that very few accomplishments will be made by Africans, or Australian or New Guinean Aborigines; members of those groups will not be highly represented in chess; and that major advances in scientific fields will come from those of European ancestry or the “Monglids”, Koreans, Chinese or Japanese.
On the other hand, Hart (2007: 417) makes two more “predictions”: he says that IQ data for Congoid Pygmies, Andaman Islanders, and Bantu-speaking people are few and far between and he believes that when enough IQ testing is undertaken there he expects IQ values between 60 and 85. Conversely, for the Lapps, Siberians, Eskimoes, Mongols and Tibetans, he predicts that IQ values should be between 85-105. He then states that if these “predictions” turn out to be wrong then he would have to admit that his hypothesis is wrong. But the thing is, he chose “predictions” that he knew would come to pass and therefore these are not novel, risky predictions but are predictions that Hart (2007) knows would come to pass.
What novel predictions has the VDH made? This is very simple. The convergent evolution of light skin was predicted in all hominids that trekked out of Africa and into colder lands. This occurred “because of the importance of maintaining the potential for producing pre-vitamin D3 in the skin under conditions of low annual UVB (Jablonski and Chaplin, 2000; Jablonski, 2004)” while these predictions “have been borne out by recent genetic studies, which have demonstrated that depigmented skin evolved independently by different molecular mechanisms multiple times in the history of the human lineage” (Chaplan and Jablonksi, 2009: 452). This was successfully predicted by Chaplan and Jablonski (2000).
The VDH still holds explanatory scope and predictive success; no other agent other than vitamin D can explain the observation that light skin is selected for in areas where there is low, absent or seasonal UVB. Conversely, in areas where there is a strong, year-round presence of UVB rays, dark skin is selected for.
Conclusion
Scientific hypotheses predict novel facts not known before the formulation of the hypothesis. The VDT has successfully predicted novel facts, whereas I am at a loss thinking of a novel fact that the CWT predicted.
In order to push an adaptationist hypothesis for CWT and ‘intelligence’, one must propose an observation that would confirm the adaptationist hypothesis while at the same time disconfirming the byproduct hypothesis. Since byproducts are inherited to, the byproduct hypothesis would predict the same things that an adaptationist hypothesis would. Thus, the CWT is a just-so story since no observation would confirm or disconfirm either hypothesis. On the other hand, the CWT doesn’t make predictions of novel facts, it makes “predictions” that are already known and would not undermine the hypothesis if disproved (but there would always be a proponent of the CWT waiting in the wings to propose an ad-hoc hypothesis in order to save the CWT, but I have already established that it isn’t science).
On the other hand, the VDT has successfully predicted that hominins that trekked out of Africa would have light skin which was then subsequently confirmed by genomic evidence. The fact that strong UVB rays year-round predict dark skin whereas seasonal, absent, or low levels of UVB predict light skin has been proved to be true. With the advent of genomic testing, it has been shown that hominids that migrated out of Africa did indeed have lighter skin. This is independent verification for the VDH; the VDH has predicted a novel fact whereas the CWT has not.
“Latinos”, Brazilians, Mixed-Race Individuals and Race Concepts
2050 words
How do “Latinos”, Brazilians, and mixed-race individuals fit into Hardimon’s (2017) differing race concepts (racialist, minimalist, populationist, and socialrace)? It’s easier explaining how “Latinos” fit into this, but mixed-race individuals are a bit trickier (for instance, the minimalist concept of race does not say anything about it and is therefore vague in that respect). This article will discuss these two populations and see where they fit into these categories.
Mixed-race individuals
Mixed-race individuals are tricky to place in these conceptions of race that Hardimon (2017) lays out and defends. For example, minimalist race itself is vague; it does not say which populations/individuals with populational characteristics would be placed, the argument just establishes the biological reality and significance of race. The concept of the “one drop rule” (was a legal) is a social standard in that anyone with “one drop” of “black blood” was deemed black (which, it seems, did well for so-called conceptions of “racial purity” since most white Americans have low amounts of black ancestry; Bryc et al, 2015).
The one drop rule was an attempt to limit racial miscegenation (racial mixing), and it seems to have, for the most part, worked since many white Americans have low to no African ancestry (since 95 percent of white Americans have no African ancestry; Bryc et al, 2015).
Though, as Hardimon (2017: 49) writes, the fact that individuals must have a race is a holdover from the racialist concept; minimalist race, as I’ve covered, is not defined by the features of an individual but is defined by the features of the group said individual belongs to. It is defined in terms of group—not individual—characteristics. So just because individual I doesn’t look like their R but instead looks like an R2, for example, doesn’t make individual I an R2; individual I is still an R even though they look like an R2 since the concept is based on shared group characteristics.
Hardimon doesn’t really discuss mixed-race individuals in his book; there’s only really one note on the subject (and it’s about social race, pg. 209, note 54):
People who are members of more than one socialrace in a socialrace regime that does not recognize mixed race as a racial position will be in the anomalous situation of not having an established socialrace position in society. Having such a position is one way of being a “normal” member of society organized around the institution of socialrace. Not to have such a position is to have no place in the social world along the dimension of socialrace. Hence, perhaps, the pathos of mixed-race individuals seeking social recognition for their distinctive mixed race identity.
Though, in certain (racialist/socialrace) societies, a mixed-race individual would become what the “lower” race is in that society. For instance, if an individual were half white and half black—like the former President of the United States Barack Obama—he would be designated as “black”, as we all know (since he’s the first black President of the United States). This is known as the concept of “hypodescent” and has its basis in Hardimon’s socialrace concept since racial status of the offspring is designated to the parent who has lower “standing” than the majority in said country. So, therefore, the concept of hypodescent is the concept of socialrace in action.
Mixed-race individuals are seen as members of their “lower-status parent group“, which shows how racial constructivism is alive today. One is designated the lower-status of their parent in a hierarchical manner—one way in which the socialrace concept borrows from the racialist race concept in that it is hierarchical.
The researchers found, for example, that one-quarter-Asian individuals are consistently considered more white than one-quarter-black individuals, despite the fact that African Americans and European Americans share a substantial degree of genetic heritage. One Drop Rule Persists—Harvard
Brazil
How does this work in Brazil? Surely this makes problems for racial concepts, right?
Brazilians don’t use the term “race” (raca), but the term “color” (cor). “The reason the word Color (capitalized to call attention to this particular meaning) is preferred to race in Brazil is probably because it captures the continuous aspects of phenotypes” (Parra et al, 2003). Clearly, the conception of “race” (raca) in Brazil comes down to what “color” (cor) one is; and so we should state that Brazilian society is stratified into “colors” and not true “races”. The 1872 Brazilian census created four “color” groups “white, caboclo, black and brown (branco, caboclo (mixed indigenous-European), negro and pardo). These groups were always defined by the same formula: Colour group = members of a pure race + phenotypes of this race in the process of reversion (Guimaraes 1999)” (Guimaraes, 2012: 1157).
Though, distinct from Hardimon (2017), Guimaraes (2012: 1160) argues that the Brazilian color system is, in fact, a racial system:
What makes colour in Brazil a racial term is precisely the fact that the physiognomic traits used by racialists to distinguish different human races became convoluted with the original European system of classification based on shades of skin colour.
Hardimon (2017: 49) writes:
On the other hand, it is not clear that the Brazilian concept of COR is altogether independent of the phenomenon we Americans designate using ‘race.’ The color that ‘COR’ picks out is racial skin color. The well-known, widespread preference for lighter (whiter) skin in Brazil is at least arguably a racial preference. It seems likely that white skin color is preferred because of its association with the white race. This provides a reason for thinking that the minimalist concept of race may be lurking in the background of Brazilian thinking about race.
Someone who is “white” (identifies as white) in Brazil will (most likely) not be white in America; this is due to their differing classification system based on color, which is loosely tied with the minimalist concept, but is distinct from it in that it’s just based largely on the color of one’s skin (one of many requisites for minimalist racehood).
The role of race and color in regard to Brazilian society is complex, biologically, sociologically, and psychologically; but it’s clear that some concept of what we could call a “classical” race concept—however crude—could be said to be in use in Brazil today.
Latinos/Hispanics
The US Census Bureau states that “People who identify their origin as Hispanic, Latino, or Spanish may be of any race.” This is true; just because one derives from a Latin American country does not mean that they are some kind of “Latino” or “Hispanic” race.
Take, as an example, the case of Alberto Fujimori. Alberto Fujimori is the son of Japanese immigrants to Peru, and he eventually ended up becoming the President of Peru. His parents emigrated to Peru and he was born in Lima. Now here’s where things get tricky: is he all of a sudden some new type of “race” called “Hispanic” or “Latino”, all because he was born across the ocean? Is Pope Francis all of a sudden not Italian by ancestry since he was born in Argentina? Are people who are born in Argentina, Chile, and Paraguay with direct ancestry to Germany and Italy all of a sudden no longer German or Italian but some new “Hispanic” or “Latino” race? No! Just because you’re born not in your ancestral home does not mean you “become” whatever society designates that part of the world (in this case “Latin America”). Their race does not change on the basis of where they are born; their individual ancestries can still be traced back to their countries of origin, therefore attempting to “racialize” the terms “Hispanic” or “Latino” do not make any sense.
“Hispanics” do not—and cannot—count as a minimalist race on the basis of one condition: they do not share a single pattern of visible physical features. No one pattern of hair, skin color, lip shape, eye shape etc. They do not share a single geographic origin; they have a mixture of Ancestry from Africa, Europe, and Asia. “Hispanics” can be seen, obviously, as a mixture of minimalist races. “Hispanics” are denied minimalist racehood since they do not exhibit characteristics of minimalist races, which is even echoed, as shown above, by the US Census Bureau.
Hardimon (2017: 39) writes:
To deny that Latinos constitute a race is not to deny that individual Latinos or Latinos as a group can be the targets of racism (for example, owing to skin color). Nor is it to deny that Latinos are often regarded as “racially other” (as differing in some essential humanly important way corresponding to skin color) by members of other racialized groups (for example, Anglos). … Nor is it to deny that they constitute a socialrace in my sense of the term. Still less does it imply that Latinos ought not to aspire to a degree of solidarity connoted by the Spanish word raza.
So “Latinos” can be designated as a socialrace (though socialraces do not always have a mirrored minimalist race), but not as a minimalist race since they do not fit the criteria for minimalist racehood. Many “Latinos” can be said to be mestizos, which are half European (normally of Spanish descent) and half Indian. Still, further, they can be castizos, about three-quarters European and one-quarter Indian. Then you have the “Latino” Carribean countries (Dominican Republic, Cuba, Puerto Rico) with differing amounts of admixture from all over, from different minimalist races. Though, in America, most Dominicans would be counted as “black” under the concept of “hypodescent”—the one drop rule. Many Cubans can be seen to have majority Spanish ancestry, and its the same for Puerto Ricans.
“Latinos/Hispanics” do not constitute a major race (in the minimalist and populationist sense) because they are a mixture of different minimalist races. This does not mean, however, that one designated as “Latino” in America does not have full ancestry to a European country; this is how the concept of “Latino” in America uses the concept of socialrace—it’s only based on the perceived race of the individual (that they derive from a “Latino” country) and that therefore makes them “Latino”, all the while ignoring their actual racial ancestry.
There is even a phrase in Latin America “Mejorar la raza” or “improve the race” by having children with lighter-skinned people since light skin is seen as beautiful in Latin America. They want to better “their race” (even though they—as “Latinos”—don’t have a race), and so they will attempt to have children with lighter and lighter people (i.e., people who have more and more European ancestry) to “improve their race” (i.e., their socialrace) since European features are seen as more beautiful in Latin America.
Most of the ruling class in Latin America is of European descent, while the lower classes are of admixed/unadmixed Indians (coming from differing tribes). This, one can say, is one way that socialrace is used in Latin America.
Conclusion
Brazilians and “Latinos/Hispanics” clearly could have been grouped in the mixed-race category, but each of these subjects has distinct concepts it needed to discuss. The Brazilian concept of “cor” and “raca” are loosely intertwined; they can be said to use aspects of mimimalist races. On the other hand, “Latinos/Hispanics” are not designated minimalist racehood on the basis that they do not share a single pattern of physical features, nor do they share a geographic origin, since the groups that make up “Latinos” (which are minimalist races) are the geographic locations in question. “Latinos/Hispanics” are not minimalist races because they do not exhibit the features of minimalist races.
Mixed-race individuals, regarding the socialrace concept, can be seen to be the “lower” of the races they are admixed with on the social ladder—which is how it is in America (the concept of hypodescent). The existence of mixed-race individuals does not invalidate the concept of race; minimalist racehood is not defined on the basis of individual characters, but on the basis of the characters of the group. Therefore this does not go against the concept of minimalist race.
The concepts of race can definitely survive these anomalies when describing the biological realities of race; some of them can be said to be socialraces for one respect, whereas in reality, they are mixtures of minimalist races. Races exist and the existence of Brazilians (even with their own categorization of races/colors) and “Latinos/Hispanics” and other mixed-race groups/individuals do not rail against any concepts that purport to argue for the existence and biological reality of race.
Is Racial Superiority in Sports a Myth? A Response to Kerr (2010)
2750 words
Racial differences in sporting success are undeniable. The races are somewhat stratified in different sports and we can trace the cause of this to differences in genes and where one’s ancestors were born. We can then say that there is a relationship between them since, they have certain traits which their ancestors also had, which then correlate with geographic ancestry, and we can explain how and why certain populations dominate (or would have the capacity to based on body type and physiology) certain sporting events. Critiques of Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It are few and far between, and the few that I am aware of are alright, but this one I will discuss today is not particularly good, because the author makes a lot of claims he could have easily verified himself.
In 2010, Ian Kerr published The Myth of Racial Superiority in Sports, who states that there is a “dark side” to sports, and specifically sets his sights on Jon Entine’s (2000) book Taboo. In this article, Kerr (2010) makes a lot of, in my opinion, giant claims which provide a lot of evidence and arguments in order to show their validity. I will discuss Kerr’s views on race, biology, the “environment”, “genetic determinism”, and racial dominance in sports (which will have a focus on sprinting/distance running in this article).
Race
Since establishing the reality and validity of the concept of race is central to proving Entine’s (2002) argument on racial differences in sports, then I must prove the reality of race (and rebut what Kerr 2010 writes about race). Kerr (2010: 20) writes:
First, it is important to note that Entine is not working in a vacuum; his assertions about race and sports are part of a larger ongoing argument about folk notions of race. Folk notions of race founded on the idea that deep, mutually exclusive biological categories dividing groups of people have scientific and cultural merit. This type of thinking is rooted in the notion that there are underlying, essential differences among people and that those observable physical differences among people are rooted in biology, in genetics (Ossorio, Duster, 2005: 2).
Dividing groups of people does have scientific, cultural and philosophical merit. The concept of “essences” has long been discarded by philosophers. Though there are differences in both anatomy and physiology in people that differ by geographic location, and this then, at the extreme end, would be enough to cause the differences in elite sporting competition that is seen.
Either way, the argument for the existence of race is simple: 1) populations differ in physical attributes (facial, morphological) which then 2) correlate with geographic ancestry. Therefore, race has a biological basis since the physical differences between these populations are biological in nature. Now that we have established that race exists using only physical features, it should be extremely simple to show how Kerr (2010) is in error with his strong claims regarding race and the so-called “mythology” of racial superiority in sports. Race is biological; the biological argument for race is sound (read here and here, and also see Hardimon, 2017).
Genetic determinism
True genetic determinism—as is commonly thought—does not have any sound, logical basis (Resnick and Vorhaus, 2006). So Kerr’s (2010) claims in this section need to be dissected here. This next quote, though, is pretty much imperative to the soundness and validity of his whole article, and let’s just say that it’s easy to rebut and invalidates his whole entire argument:
Vinay Harpalani is one of the most outspoken critics of using genetic determinism to validate notions of inferiority or the superiority of certain groups (in this case Black athletes). He argues that in order for any of Entine’s claims to be valid he must prove that: 1) there is a systematic way to define Black and White populations; 2) consistent and plausible genetic differences between the populations can be demonstrated; 3) a link between those genetic differences and athletic performance can be clearly shown (2004).
This is too easy to prove.
1) While I do agree that the terminology of ‘white’ and ‘black’ are extremely broad, as can be seen by looking at Rosenberg et al (2002), population clusters that cluster with what we call ‘white’ and ‘black’ exist (and are a part of continental-level minimalist races). So is there a systematic way to define ‘Black’ and ‘White’ populations? Yes, there is; genetic testing will show where one’s ancestors came from recently, thereby proving point 1.
2) Consistent and plausible genetic differences between populations can be demonstrated. Sure, there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016). Even these small between-continent/group differences would have huge effects on the tail end of said distribution.
3) I have compiled numerous data on genetic differences between African ethnies and European ethnies and how these genetic differences then cause differences in elite athletic performance. I have shown that Jamaicans, West Africans, Kenyans and Ethiopians (certain subgroups of the two aforementioned countries) have genetic/somatypic differences that then lead to differences in these sporting competitions. So we can say that race can predict traits important for certain athletic competitions.
1) The terminology of ‘White’ and ‘Black’ are broad; but we can still classify individuals along these lines; 2) consistent and plausible genetic differences between races and ethnies do exist; 3) a link between these genetic differences between genes/athletic differences between groups can be found. Therefore Entine’s (2002) arguments—and the validity thereof—are sound.
Kerr (2010) then makes a few comments on the West’s “obsession with superficial physical features such as skin color”, but using Hardimon’s minimalist race concept, skin color is a part of the argument to prove the existence and biological reality of race, therefore skin color is not ‘superficial’, since it is also a tell of where one’s ancestors evolved in the recent past. Kerr (2010: 21) then writes:
Marks writes that Entine is saying one of three things: that the very best Black athletes have an inherent genetic advantage over the very best White athletes; that the average Black athlete has a genetic advantage over the average White athlete; that all Blacks have the genetic potential to be better athletes than all Whites. Clearly these three propositions are both unknowable and scientifically untenable. Marks writes that “the first statement is trivial, the secondly statistically intractable, and the third ridiculous for its racial essentialism” (Marks, 2000: 1077).
The first two, in my opinion (the very best black athletes have an inherent genetic advantage over the very best white athletes and the average black athlete has a genetic advantage over the average white athlete), are true, and I don’t know how you can deny this; especially if you’re talking about AVERAGES. The third statement is ridiculous, because it doesn’t work like that. Kerr (2010), of course, states that race is not a biological reality, but I’ve proven that it is so that statement is a non-factor.
Kerr (2010) then states that “ demonstrating across the board genetic variations between
populations — has in recent years been roundly debunked“, and also says “ Differences in height, skin color, and hair texture are simply the result of climate-related variation.” This is one of the craziest things I’ve read all year! Differences in height would cause differences in elite sporting competition; differences in skin color can be conceptualized as one’s ancestors’ multi-generational adaptation to the climate they evolved in as can hair texture. If only Kerr (2010) knew that this statement here was the beginning of the end of his shitty argument on Entine’s book. Race is a social construct of a biological reality, and there are genetic differences between races—however small (Risch et al, 2002; Tang et al, 2005) but these small differences can mean big differences at the elite level.
The “environment” and biological variability
Kerr (2010) then shifts his focus over to, not genetic differences, but biological differences. He specifically discusses the Kenyans—Kalenjin—stating that “height or weight, which play an instrumental role in helping define an individual’s athletic prowess, have not been proven to be exclusively rooted in biology or genetics.” While estimates of BMI and height are high (both around .8), I think we can disregard the numbers since they came from highly flawed twin studies, since molecular genetic evidence shows lower heritabilities. Either way, surely height is strongly influenced by ‘genes’. Another important caveat is that Kenya has one of the lowest BMIs in the world, 20.7 for Kenyan men, which also is part of the cause of why certain African ethnies dominate running competitions.
I don’t disagree with Kerr (2010) here too much; many papers show that SES/cultural/social factors are very important to Kenyan runners (Onywera et al, 2006; Wilbur and Pistiladis, 2012; Tucker, Onywera, and Santos-Concejero, 2015). You can have all of the ‘physical gifts’ in the world, if it’s not combined with the will to want to do your best, along with cultural and social factors you won’t succeed. But having an advantageous genotype and physique are useless without a strong mind (Lippi, Favaloro, and Guidi, 2008):
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.”
Dissecting this, though, is tougher. Because being born at certain altitudes will cause certain advantageous traits, such as a larger lung capacity (and you will have an advantage in lung capacity when competing at lower altitudes), but certain subpopulations live in these high-altitude areas, so what is it? Genetic? Cultural? Environmental? All three? Nature vs nurture is a false dichotomy; so it is a mixture of the three.
How does one explain, then, the athlete who trains countless hours a day fine-tuning a jump shot, like LeBron James or shaving seconds off sub-four minute miles like Robert Kipkoech Cheruiyot, a four time Boston Marathon winner?
Literally no one denies that elite athletes put in insane amounts of practice; but if everyone has the same amount of practice they won’t have similar abilities.
He also briefly brings up muscle fibers, stating:
These include studies on African fast twitch muscle fibers and development of motor skills. Entine includes these studies to demonstrate irrevocable proof of embedded genetic differences between populations but refuses to accept the fact that any differences may be due to environmental factors or training.
This, again, shows ignorance of the literature. An individual’s muscle fibers are formed during development from the fusion of several myoblasts, with differentiation being completed before birth. Muscle fiber typoing is also set at age 6, no difference in skeletal muscle tissue was found when comparing 6-year-olds and adults, therefore we can state that muscle fiber typing is set by age 6 (Bell et al, 1980). You can, of course, train type II fibers to have similar aerobic capacity to type I fibers, but they’ll never be fully similar. This is something that Kerr (2010) obviously is ignorant to because he’s not well-read on the literature which causes him to make dumb statements like “any differences [in muscle fiber typing] may be due to environmental factors or training“.
Black domination in sports
Finally, Kerr (2010) discusses the fact that whites dominated certain running competitions in the Olympics and that before the 1960s, a majority of distance-running gold medals went to white athletes. He then states that the 2008 Boston Marathon winner was Kenyan; but the next 4 behind him were not. Now, let’s check out the 2017 Marathon winners: Kenya, USA, Japan for the top 3; while 5 Kenyans/Ethiopians are in the top 15 while the same is also true of women; a Kenyan winner, with Kenyans/Ethiopians taking 5 of the top 15 spots. The fact that whites used to do well in running sports is a non-factor; Jesse Owens blew away the competition in the Games in Germany, which showed how blacks would begin to dominate in the US decades later.
Kerr (2010) then ends the article with a ton of wild claims; the wildest one, in my opinion, being that “Kenyans are no more genetically different from any other African or European population on average“, does anyone believe this? Because I have data to the contrary. They have a higher Vo2 max, which of course is trainable but with a ‘genetic’ component (Larsen, 2003), while other authors argue that genetic differences between populations account for differences in success in running competition between populations (Vancini et al, 2014), while male and female Kenyan and Ethiopian runners are the fastest in the half and full marathon (Knechtle et al, 2016). There is a large amount of data out there that speaks about Kenyan/Ethiopian and others’ dominance in running; it seems Kerr (2010) just ignored the data. I agree with Kerr that Kenyanholos show that humans can adapt to their environment; but his conclusion here:
The fact that runners coming from Kenya do so well in running events attests to the fact the combination of intense high altitude training, consumption of a low-fat, high protein diet, and a social and cultural expectation to succeed have created in recent decades an environment which is highly conducive to producing excellent long-distance runners.
is very strong, and while I don’t disagree at all with anything here, he’s disregarding how somatype and genes differ between Kenyans and other populations that compete in these sports that then lead to differences in elite sporting competitions.
Elite sporting performance is influenced by myriad factors, including psychology, ‘environment’, and genetic factors. Something that Kerr (2010) doesn’t understand—because he’s not well-read on this literature—is that many genetic factors that influence sporting performance are known. The ability to become elite depends on one’s capacity for endurance, muscle performance, the ability of the tendons and ligaments to withstand stress and injury, and the attitude to train and push above and beyond what normal people can do (Lippi, Longo, and Maffulli, 2010). We can then extend this to human races; some are better-equipped to excel in running competitions than others.
On its face, Kerr’s (2010) claim that there are no inherent differences between races is wrong. Races differ in somatype, which is due to evolution in different geographic locations for tens of thousands of years. The human body is perfectly adapted to for long distance running (Murray and Costa, 2012), and since our capabilities for endurance running evolved in Africa and they, theoretically, have a musculoskeletal structure similar to the Homo sapiens that left Africa around 70 kya, then it’s only logical to state that African’s, on average, have an inherent ability in running competitions (West and East Africans, while North Africans fare very well in middle distance running, which, again, comes down to living in higher altitudes like Kenyans and Ethiopians).
Wagner and Heyward (2000) reviewed many studies on the physiological differences between blacks and whites. Blacks skew towards mesomorphy; black youths had smaller billiac and bitrochanteric width (the widest measure of the pelvis at the outer edges and the flat process on the femur, respectively), and black infants had longer extremities than white infants (Wagner and Heyward, 2000). We have anatomic evidence that blacks are superior runners (in an American context). Mesomorphic athletes are more likely to be sprinters (Sands et al, 2005; which is also seen in prepubescent children: Marta et al, 2013) Kenyans are ecto-dominant (Vernillo et al, 2013) which helps to explain their success at long-distance running. So just on only looking at the phenotype (a marker for race with geographic ancestry, proving the biological existence of race) we can confidently state, on average just by looking at an individual or a population, how they will fare in certain competitions.
Conclusion
Kerr’s (2010) arguments leave a ton to be desired. Race exists and is a biological reality. I don’t know why this paper got published since it was so full of errors; his arguments were not sound and much of the literature contradicts his claims. What he states at the end about Kenyans is not wrong at all, but to not even bring up genetic/biologic differences as a factor influencing their performance is dishonest.
Of course, a whole slew of factors, be they biological, cultural, psychological, genetic, socioeconomic, anatomic, physiologic etc influence sporting performance, but certain traits are more likely to be found in certain populations, and in the year 2018 we have a good idea of what influences elite sporting performance and what does not. It just so happens that these traits are unevenly distributed between populations, and the cause is evolution in differing climates in differing geographic locations.
Race exists and is a biological reality. Biological anatomic/physiological differences between these races then manifest themselves in elite sporting competition. The races differ, on average, in traits important for success in certain competitions. Therefore, race explains some of the variance in elite sporting competition.
Minimalist Races Exist and are Biologically Real
3050 words
People look different depending on where their ancestors derived from; this is not a controversial statement, and any reasonable person would agree with that assertion. Though what most don’t realize, is that even if you assert that biological races do not exist, but allow for patterns of distinct visible physical features between human populations that then correspond with geographic ancestry, then race—as a biological reality—exists because what denotes the physical characters are biological in nature, and the geographic ancestry corresponds to physical differences between continental groups. These populations, then, can be shown to be real in genetic analyses, and that they correspond to traditional racial groups. So we can then say that Eurasian, East Asian, Oceanian, black African, and East Asians are continental-level minimalist races since they hold all of the criteria needed to be called minimalist races: (1) distinct facial characters; (2) distinct morphologic differences; and (3) they come from a unique geographic location. Therefore minimalist races exist and are a biological reality. (Note: There is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016), but this does not mean that the minimalist biological concept of race has no grounding in biology.)
Minimalist race exists
The concept of minimalist race is simple: people share a peculiar geographic ancestry unique to them, they have peculiar physiognomy (facial features like lips, facial structure, eyes, nose etc), other physical traits (hair/hair color), and a peculiar morphology. Minimalist races exist, and are biologically real since minimalist races can survive findings from population genetics. Hardimon (2017) asks, “Is the minimalist concept of race a social concept?” on page 62. He writes that social concepts are socially constructed in a pernicious sense if and only if it “(i) fails to represent any fact of the matter and (ii) supports and legitimizes domination.” Of course, populations who derive from Africa, Europe, and East Asia have peculiar facial morphology/morphology unique to that isolated population. Therefore we can say that minimalist race does not conform to criteria (i). Hardimon (2017: 63) then writes:
Because it lacks the nasty features that make the racialist concept of race well suited to support and legalize domination, the minimalist race concept fails to satisfy condition (ii). The racialist concept, on the other hand, is socially constructed in the pernicious sense. Since there are no racialist races, there are no facts of the matter it represents. So it satisfies (i). To elaborate, the racialist race concept legtizamizes racial domination by representing the social hierarchy of race as “natural” (in a value-conferring sense): as the “natural” (socially unmediated and inevitable) expression of the talent and efforts of the inidividuals who stand on its rungs. It supports racial domination by conveying the idea that no alternative arrangment of social institutions could possibly result in racial equality and hence that attempts to engage in collective action in the hopes of ending the social hierarchy of race are futile. For these reasons the racialist race concept is also idealogical in the prejorative sense.
Knowing what we know about minimalist races (they have distinct physiognomy, distinct morphology and geographic ancestry unique to that population), we can say that this is a biological phenomenon, since what makes minimalist races distinct from one another (skin color, hair color etc) are based on biological factors. We can say that brown skin, kinky hair and full lips, with sub-Saharan African ancestry, is African, while pale/light skin, straight/wavy/curly hair with thin lips, a narrow nose, and European ancestry makes the individual European.
These physical features between the races correspond to differences in geographic ancestry, and since they differ between the races on average, they are biological in nature and therefore it can be said that race is a biological phenomenon. Skin color, nose shape, hair type, morphology etc are all biological. So knowing that there is a biological basis to these physical differences between populations, we can say that minimalist races are biological, therefore we can use the term minimalist biological phenomenon of race, and it exists because there are differences in the patterns of visible physical features between human populations that correspond to geographic ancestry.
Hardimon then talks about how eliminativist philosophers and others don’t deny that above premises above the minimalist biological phenomenon of race, but they allow these to exist. Hardimon (2017: 68-69) then quotes a few prominent people who profess that there are, of course, differences in physical features between human populations:
… Lewontin … who denies that biological races exist, freely grants that “peoples who have occupied major geographic areas for much of the recent past look different from one another. Sub-Saharan Africans have dark skin and people who have lived in East Asia tend to have a light tan skin and an eye color and eye shape that is difference from Europeans.” Similarly, population geneticist Marcus W. Feldman (final author of Rosenberg et al., “Genetic Stucture of Human Populations” [2002]), who also denies the existence of biological races, acknowledges that “it has been known for centuries that certain physical features of humans are concentrated within families: hair, eye, and skin color, height, inability to digest milk, curliness of hair, and so on. These phenotypes also show obvious variation among people from different continents. Indeed, skin color, facial shape, and hair are examples of phenotypes whose variation among populations from different regions is noticeable.” In the same vein, eliminative anthropologist C. Loring Brace concedes, “It is perfectly true that long term residents of various parts of the world have patterns of features that we can identify as characteristic of they area from which they come.”
So even these people who claim to not believe in “biological races”, do indeed believe in biological races because what they are describing is biological in nature and they, of course, do not deny that people look different while their ancestors came from different places so therefore they believe in biological races. We can then use the minimalist biological phenomenon of race to get to the existence of minimalist races.
Hardimon (2017: 69) writes:
Step 1. Recognize that there are differences in patterns of visible physical features of human beings that correspond to their differences in geographic ancestry.
Step 2. Observe that these patterns are exhibited by groups (that is, real existing groups).
Step 3. Note that the groups that exhibit these patterns of visible physical features correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race.
Step 4. Infer that minimalist race exists.
Those individuals mentioned previously who deny biological races but allow that people with ancestors from differing geographic locales look differently do not disagree with step 1, nor does anyone really disagree with step 2. Step 4’s inference immediately flows from the premise in step 3. “Groups that exhibit patterns or visible physical features that correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race. Call (1)-(4) the argument from the minimalist biological phenomenon of race” (Hardimon, 2017: 70). Of course, the argument does not identify which populations may be called races (see further below), it just shows that race is a biological reality. Because if minimalist races exist, then races exist because minimalist races are races. Minimalist races exist, therefore biological races exist. Of course, no one doubts that people come from Europe, sub-Saharan Africa, East Asia, the Americas, and the Pacific Islands, even though the boundaries between them are ‘blurry’. They exhibit patterns of visible physical characters that correspond to their differing geographic ancestry, they are minimalist races therefore minimalist races exist.
Pretty much, the minimalist concept of race is just laying out what everyone knows and arguing for its existence. Minimalist races exist, but are they biologically real?
Minimalist races are biologically real
Of course, some who would assert that minimalist races do not exist would say that there are no ‘genes’ that are exclusive to one certain population—call them ‘race genes’. Of course, these types of genes do not exist. Whether or not one individual is a part of one race or not does not rest on the basis of his physical characters, but is determined by who his parents are, because one of the three premises for the minimalist race argument is ‘must have a peculiar geographic ancestry’. So it’s not that members of races share sets of genes that other races do not, it’s based on the fact that they share a distinctive set of visible physical features that then correspond with geographic ancestry. So of course if the minimalist concept of race is a biological concept then it entails more than ‘genes for’ races.
Of course, there is a biological significance to the existence of minimalist biological races. Consider that one of the physical characters that differ between populations is skin color. Skin color is controlled by genes (about half a dozen within and a dozen between populations). Lack of UV rays for individuals with dark skin will lead to diseases like prostate cancer, while darker skin is a protectant against UV damage to human skin (Brenner and Hearing, 2008; Jablonksi and Chaplin, 2010). Since minimalist race is biologically significant and minimalist races are partly defined by differences in skin color between populations then skin color has both medical and ecological significance.
(1) Consider light skin. People with light skin are more susceptible to skin cancer since they evolved in locations with poor UVR radiation (D’Orazio et al, 2013). The body needs vitamin D to absorb and use calcium for maintaining proper cell functioning. People who evolved near the equator don’t have to worry about this because the doses of UVB they absorb are sufficient for the production of enough previtamin D. While East Asians and Europeans on the other hand, became adapted to low-sunlight locations and therefore over time evolved lighter skin. This loss of pigmentation allowed for better UVB absorption in these new environments. (Also read my article on the evolution of human skin variation and also how skin color is not a ‘tell’ of aggression in humans.)
(2) While darker-skinned people have a lower rate of skin cancer “primarily a result of photo-protection provided by increased epidermal melanin, which filters twice as much ultraviolet (UV) radiation as does that in the epidermis of Caucasians” (Bradford, 2009). Dark skin is thought to have evolved to protect against skin cancer (Greaves, 2014a) but this has been contested (Jablonski and Chaplin, 2014) and defended (Greaves, 2014b). So therefore, using (1) and (2), skin color has evolutionary signifigance.
So as humans began becoming physically adapted to their new niches they found themselves in, they developed new features distinct from the location they previously came from to better cope with the new lifestyle due to their new environments. For instance “Northern Europeans tend to have light skin because they belong to a morphologically marked ancestral group—a minimalist race—that was subject to one set of environmental conditions (low UVR) in Europe” (Hardimon, 2017: 81). Of course explaining how human beings survived in new locations falls into the realm of biology, while minimalist races can explain why this happened.
Minimalist races clearly exist since minimalist races constitute complex biological patterns between populations. Hardimon (2017: 83) writes:
It [minimalist race] also enjoys intrinsic scientific interest because it represents distinctive salient systematic dimension of human biological diversity. To clarify: Minimalist race counts as (i) salient because human differences of color and shape are striking. Racial differences in color and shape are (ii) systematic in that they correspond to differences in geographic ancestry. They are not random. Racial differences are (iii) distinctive in that they are different from the sort of biological differences associated with the other two salient systematic dimensions of human diversity: sex and age.
[…]
An additional consideration: Like sex and age, minimalist race constitutes one member of what might be called “the triumverate of human biodiversity.” An account of human biodiversity that failed to include any one of these three elements would be obviously incomplete. Minimalist race’s claim to be biologically real is as good as the claim of the other members of the triumverate. Sex is biologically real. Age is biologically real. Minimalist race is biologically real.
Real does not mean deep. Compared to the biological associated with sex (sex as contrasted with gender), the biological differences associated with minimalist race are superficial.
Of course, the five ‘clusters’ and ‘populations’ identified by Rosenberg et al’s (2002) K=5 graph, which told structure to produce 5 genetic clusters, corresponds to Eurasia, Africa, East Asia, Oceania, and the Americas, are great candidates for minimalist biological races since they correspond to geographic locations, and even corroborates what Fredrich Blumenbach said about human races back in the 17th century. Hardimon further writes (pg 85-86):
If the five populations corresponding to the major areas are continental-level minimalist races, the clusters represent continental-level minimalist races: The cluster in the mostly orange segment represents the sub-Saharan African continental-level minimalist race. The cluster in the mostly blue segment represents the Eurasian continental-level minimal race. The cluster in the mostly pink segment represents the East Asian continental-level minimalist race. The cluster in the mostly green segment represents the Pacific Islander continental-level minimalist race. And the cluster in the mostly purple segment represents the American continental-level minimalist race.
[…]
The assumption that the five populations are continental-level minimalist races entitles us to interpret structure as having the capacity to assign individuals to continental-level minimalist races on the basis of markers that track ancestry. In constructing clusters corresponding to the five continental-level minimalist races on the basis of objective, race-neutral genetic markers, structure essentially “reconstructs” those races on the basis of a race-blind procedure. Modulo our assumption, the article shows that it is possible to assign individuals to continental-level races without knowing anything about the race or ancestry of the individuals from whose genotypes the microsattelites are drawn. The populations studied were “defined by geography, language, and culture,” not skin color or “race.”
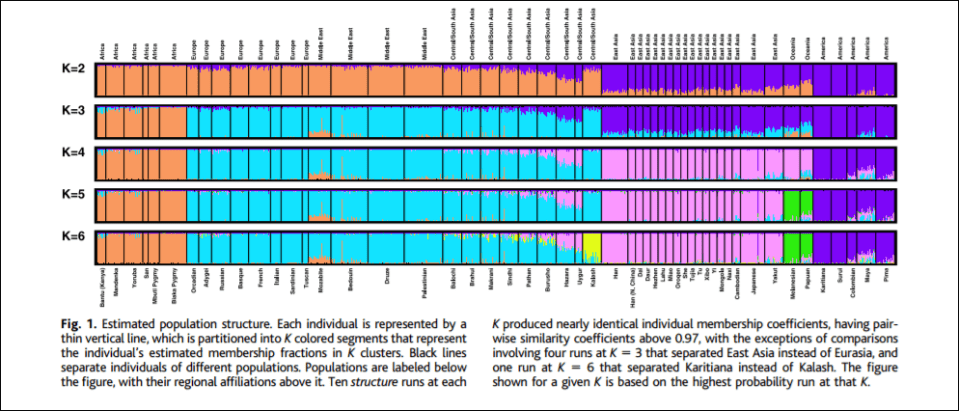
Of course, as critics note, the researchers predetermine how many populations that structure demarcates, for instance, K=5 indicates that the researchers told the program to delineate 5 clusters. Though, these objections do not matter. For the 5 populations that come out in K=5 “are genetically structured … which is to say, meaningfully demarcated solely on the basis of genetic markers” (Hardimon, 2017: 88). K=6 brings one more population, the Kalash, a group from northern Pakistan who speak an Indo-European language. Though “The fact that structure represents a population as genetically distinct does not entail that the population is a race. Nor is the idea that populations corresponding to the five major geographic areas are minimalist races undercut by the fact that structure picks out the Kalash as a genetically distinct group. Like the K=5 graph, the K=6 graph shows that modulo our assumption, continental-level races are genetically structured” (Hardimon, 2017: 88).
Though of course there are naysayers. Svante Paabo and David Serre, Hardimon writes, state that when individuals are sampled from homogeneous populations from around the world, the gradients of the allele frequencies that are found are distributed randomly across the world rather than clustering discretely. Though Rosenberg et al responded by verifying that the clusters they found are not artifacts of sampling as Paabo and Serre imply, but reflect features of underlying human variation. Though Rosenberg et al agree with Paabo and Serre in that that human genetic diversity consists of clines in variation in allele frequencies (Hardimon, 2017: 89). Other naysayers also state that all Rosenberg et al show is what we can “see with our eyes”. Though a computer does not partition individuals into different populations based on something that can be done with eyes, it’s based on an algorithm.
Hardimon also accepts that black Africans, Caucasians, East Asians, American Indians and Oceanians can be said to be races in the basic sense because “they constitute a partition of the human species“, and that they are distinguishable “at the level of the gene” (Hardimon, 2017: 93). And of course, K=5 shows that the 5 races are genetically distinguishable.
Hardimon finally discusses some medical significance for minimalist races. He states that if you are Caucasian that it is more likely that you have a polymorphism that protects against HIV compared to a member of another race. Meanwhile, East Asians are more likely to carry alleles that make them more susceptible to Steven-Johnson syndrome or another syndrome where their skin falls off. Though of course, the instances where this would matter in a biomedical context are rare, but still should be at the back of everyone’s mind (as I have argued), even though instances where medical differences between minimalist races are rare, there are times where one’s race can be medically significant.
Hardimon finally states that this type of “metaphysics of biological race” can be called “deflationary realism.” Deflationary because it “consists in the repudiation of the ideas that racialist races exist and that race enjoys the kind of biological reality that racialist race was supposed to have” and realism which “consists in its acknowledgement of the existence of minimalist races and the genetically grounded, relatively superficial, but still significant biological reality of minimalist race” (Hardimon, 2017: 95-96).
Conclusion
Minimalist races exist. Minimalist races are a biological reality because distinct visible patterns show differences between geographically isolated populations. This is enough for the classification of the five classic races we know of to be called race, be biologically real, and have a medical significance—however small—because certain biological/physical traits are tied to different geographic populations—minimalist races.
Hardimon (2017: 97) shows an alternative to racialism:
Deflationary realism provides a worked-out alternative to racialism—it it a theory that represents race as a genetically grounded, relatively superficial biological reality that is not normatively important in itself. Deflationary realism makes it possible to rethink race. It offers the promise of freeing ourselves, if only imperfectly, from the racialist background conception of race.
It is clear that minimalist races exist and are biologically real. You do not need to speak about supposed mental traits between these minimalist races, they are irrelevant to the existence of these minimalist biological races. As Hardimon (2017: 67) writes: “No reference is made to normatively important features such as intelligence, sexuality, or morality. No reference is made to essences. The idea of sharp boundaries between patterns of visible physical features or corresponding geographical regions is not invoked. Nor again is reference made to the idea of significant genetic differences. No reference is made to groups that exhibit patterns of visible physical features that correspond to geographic ancestry.”
The minimalist biological concept of race stands up to numerous lines of argumentation, therefore we can say without a shadow of a doubt that minimalist biological race exists and is real.
Stress and Race Redux: A ‘Hispanic’ Paradox?
1500 words
American Renaissance published an article the other day titled “Is ‘Racism’ Killing Black People?” and, for the most part, I largely agree with it. However, there are a few faults in it that I need to address.
First, off, as the article rightly noted, it’s not only perceived ‘racism’ that is the cause for these health disparities, but stress from other blacks as well. Gregory Hood (the author of the AmRen article) cites a new study showing that blacks who move out of the ‘hood’ see a subsequent decrease in BP (Kershaw et al, 2017). They followed 2,290 people 974 were men and 1,306 were women. This is data collected from the CARDIA study which has helped us to understand racial disparities in all types of different health outcomes. Blacks who lived in high segregation neighborhoods had higher levels of SBP (systolic blood pressure), and saw a decrease in their SBP when they moved to less segregated neighborhoods. The authors conclude that “policies that reduce segregation may have meaningful health benefits.” What kind of policies will ‘reduce segregation’? Most races/ethnic groups group together in an area, so I don’t see how this would happen.
In regards to the argument on black maternal mortality and ‘racism’, I think it’s much more nuanced. Black women are 2 to 6 times more likely to die giving birth than white women; while the leading causes of maternal death in black women is pregnancy-induced hypertension, and embolism (Chang et al, 2003), though reasons for the mortality rate are not explainable at present (Flanders-Stepans, 2002). Further, in regards to preeclampasia, women who get pregnant at younger ages are more likely to acquire the disease while pregnant, and blacks and other non-whites are more likely to get pregnant at younger ages than whites (Main et al, 2015).
However, there are ways to reduce maternal mortality in black women. In a RCT undertaken between the years of 1990-2011 in Memphis, Tennesee, black women were followed with their live-in children and placed into one of four groups: “treatment 1 (transportation for prenatal care [n = 166]), treatment 2 (transportation plus developmental screening for infants and toddlers [n = 514]), treatment 3 (transportation plus prenatal/postpartum home visiting [n = 230]), and treatment 4 (transportation, screening, and prenatal, postpartum, and infant/toddler home visiting [n = 228])” (Olds et al, 2014). They conclude that pre- and post-natal care greatly reduces maternal/infant mortality in black women, “living in highly disadvantaged settings.”
Further, the racial disparity in post-term neonates is largely driven by “CHD among term infants with US-born mothers is driven predominately by the postneonatal survival disadvantage of African-American infants” (Collins et al, 2017). Though, as can be seen in the study by Olds et al (2014), pre- and post-natal care can greatly reduce both infant and maternal mortality.
Stress can also be measured in pregnant women by measuring the level of blood cortisol (Gillespie et al, 2017). They show that, independent of adulthood stress, stress during childhood may shape birth timing, with cortisol being the biological mediator. This may be an explanation for what Gregory Hood notes. He states in his article that there has to be an explanation for why black women birth earlier, and while I am sympathetic to biological models ala Rushton (1997), Gillespie et al (2017) drive a hard argument that stress during childhood using cortisol as a biological mediator makes a lot of sense.
There are a few studies that attest to pre- and post-natal care having a large effect on the morality of black women, and that having the carers being black women seems to have a positive effect (Guerra-Reyes and Hamilton, 2017). They conclude that “Recognition, support, and increasing the number of African-American midwives and birth assistants is vital in tackling health inequalities.” In regards to infant mortality rate (IMR), 18 states will achieve racial equality by 2050 if current trends from 1999-2013 hold (Joedrecka et al, 2017).
Now for the main reason I decided to write this: the ‘Hispanic’ paradox. This paradox is that for the past twenty years, ‘Hispanics’ with low SES have similar or better health outcomes than whites (Franzini, Ribble, and Keddie, 2001). However, more recent analyses show that the ‘Hispanic’ paradox does not exist, mostly due to methodological problems and migrant selectivity (Crimmins et al, 2007; Teruya and Bazargan-Hezeji, 2013) and was not noticed in Chile either (Cabiesies, Tunstall, and Pickett, 2013). There is no migrant selectivity in regards to smoking, however (Fenelon, 2013, 2016).
Teruya and Bazargan-Hezejie (2013) write:
Studies which advocate the validity of the Immigrant Paradox are countered by those which report specific, negative physical and mental health outcomes, and higher rates of substance use, especially among immigrant adolescents. Findings may also be compromised by fundamental methodological concerns such as migrant health selectivity, and approaches that consider only selectively healthy groups. Moreover, the Immigrant Paradox’s benefits do not appear to extend evenly and consistently to all races, ethnicities and subgroups. Similarly, the Hispanic Paradox does not protect consistently across all Latino ethnicities, age groups and genders, with Puerto Ricans and Cubans in particular found to enjoy fewer health advantages.
This is good evidence that the people who migrate to America are healthier, and that the symptoms of low SES show in their children, but not in them because they are a self-selected population. There is no ‘Hispanic’ paradox (Smith and Bradshaw, 2006; Schoenthaler, 2016). Even a new meta-analysis on this ‘paradox’ states “Immigrant children and youth suffer from an immigrant mortality disadvantage” (Shor, Roelfs, and Vamg, 2017).
Lastly, Gregory Hood brings up stress and suicide, stating that if blacks were really more stressed than whites then blacks would have higher rates of suicide, but some studies show that whites have a higher rate of suicidal ideation, while others do not show this (Perez-Rodriguez et al, 2010). Though, as Balis and Postolache (2010) show, studies show that while there is conflicting evidence in regards to racial/ethnic differences in suicide, whites still attempt it the most. However, suicide for young black Americans is on the rise. Ahmedani et al (2016) show that “Nearly 27% of White individuals made a mental health visit versus less than 20% of Asian, Hawaiian/Pacific Islander, and Black individuals in this period. Within 4 weeks, all visits and mental health visits remained most common for White individuals (67.3% and 47.4%, respectively) and least common among Asian individuals (52.8% and 31.9%, respectively). Within 52-weeks, more than 90% made any visit. Alaskan Native/Native American (81.5%) and White individuals (79.5%) made mental health visits 10–25% more often than other groups.” However, at least in Fulton County, Georgia, black suicide decedents were less likely to report depression than white suicide decedents (Abe et al, 2008).
However, for whites, as noted in this 1982 New York Times article, suicidal feelings “reflects feelings of loneliness and hopelessness, which can be greater factors as one grows older; for instance, after loved ones have died” whereas for older white men, loss of status may be a cause, which would not be that prevalent in lower SES ethnicities. The article seems to implicate loss of status as a main cause for higher rates of suicide in white Americans, and states that as other, lower SES ethnies attain higher status, that suicide rates would rise for them as well.
Another cause could be prescription drugs, for instance in the Northeast which has been hit hard by the opiate/heroin crisis which leads to more white deaths. Robert Putnam puts this on “the links between poverty, hopelessness and health” and states that the suicide rate has declined for two groups, black males and males over the age of 75. Further, “divorce, economic strain, or political repression are often characterized as suicide risks.” Cheng et al (2010) show that “A high level of identification with one’s ethnic group was associated with lower rates of suicide attempts.” So, it seems that if one keeps their status, and has a high level of identification with their ethnic group, whites would then be protected against suicidal ideation. Nonmetropolitan counties also have higher rates of suicide than metropolitan counties (Ivey-Stephenson et al, 2017). People who livee in rural counties are less likely to seek help for mental problems (Carpenter-Song and Snell-Rood, 2016). Whites are also more likely to live in rural areas. This could explain higher rates of suicide in whites, along with loss of status, depression and drug use.
In conclusion, the ‘Hispanic’ paradox doesn’t exist; whites attempt/commit suicide more due to loss of status and since most whites live in rural areas, they do not seek help for their mental health problems which then leads to suicide. In regards to black maternal mortality/infant mortality rates, if they have midwives present during and after the birth, mortality rates have decreased. If these trends continue, there will be racial equality in terms of maternal/infant mortality in 18 states. The AmRen article was good and well written, but there were a few huge flaws. The author assumed that since the ‘Hispanic paradox’ exists, that this should have one disregard the effects of, say, stress on blood pressure in black Americans, as I have discussed in the past. But since the ‘Hispanic’ paradox does not exist, then you cannot say that (perceived) discrimination and ‘racism’ is not a cause for higher rates of mortality in blacks compared to whites.
Blood Pressure, Stress, and the Social Environment: On Black-White Differences in Blood Pressure
1800 words
Blood pressure (BP) is a physiological variable. Therefore since it is a physiological variable then it can be affected by environmental and social changes. How do racial differences come into play here, for instance? Since blacks face more (perceived) discrimination, then they should, on average, have higher BP levels than whites. This is what we find—but the effect is mostly seen in low-income blacks. How do psychosocial factors come into play here in the black-white BP gap?
BP is regulated by cardiac output, vascular resistance of blood flow, blood volume, arterial stiffness, and, of course, the individual’s emotional state which can decrease or increase BP. Neural mechanisms also exist which regulate BP (Chopra, Baby, and Jacob, 2011). Knowing how and why BP increases or decreases will have us better understand the social contexts of increased BP in low SES blacks.
BP is a complex physiological trait. It can go up and down due to what occurs in the immediate environment. Values of 120/80 mmHg are cited as ‘average’ values, but we have no idea what an ‘average’ BP is. Nevertheless—like most/all physiological variables—there is a wide range of what is considered ‘normal’. Due to the variance in human physiological systems, what is ‘normal’ for one individual is not ‘normal’ for another. Variation in BP (like, say, 120 SBP (systolic blood pressure) to 140 SBP) is ‘normal’. I believe even around 110 for SBP is within that range. For DPB (diastolic blood pressure) between 75 and 90 is within normal diurnal fluctuations due to activity/eating/etc (Taylor, Wilt, and Welch, 2011). BP, like testosterone, is one of those tricky variables to measure and so must be measured upon waking to see if there are any problems. So even for a trait like BP, there seems to be a ‘normal range’.
About 33 percent of blacks have hypertension (HTN) (Peters, Arojan, and Flack, 2006). Urban blacks are more likely to have higher BP levels than whites, but “At present, there is no complete explanation for these differences and further research is required” (Lindhorst et al, 2007). Low SES is correlated with higher levels of BP in black Americans—especially those with darker skin—but not Africans in Africa (Fuchs, 2011), suggesting that this is an American phenomenon that needs to be addressed. One good explanation, in my view, is the social environment. Physiological traits are extremely malleable due to the need to be able to ‘change gears’ in an instant, for instance to either fight or flight. Though, in our modernized world, these responses—mostly—have no need and so (due to our supposed civilized behavior), one’s BP rises due to social stress and other environmental factors and it is due to the urban environment.
What is the cause of high BP in blacks?
One explanation that has been given to explain higher rates of BP in blacks when compared to whites is discrimination. However, studies show mixed evidence on whether or not so-called discrimination raises BP (Couto, Goto, and Bastos, 2012). The same American effect (American blacks having higher BP than American whites) is seen even in the UK London area (Agyemang and Bhopal, 2003). This, yet again, is more evidence that the social environment drives these differences—again, regardless of whether or not any of the discrimination is real or imagined. Say most of it were imagined: it’d be irrelevant because the imagined discrimination leads to very real physiological outcomes in BP.
The country of birth also has an effect on BP. In one study, it was noted that Africans had significantly higher BP when compared to Asians (which is identical/lower) and native French living in France (Bahous et al, 2015). Ethnic differences in BP increase due to similar sodium intake is lower than what is usually cited (Graudal and Jurgens, 2015). However, other authors have pointed out that basing conclusions off of observational studies have problems, like the estimation of sodium intake being inaccurate since it’s a one-time measure; (Gunn et al, 2013; Cobb et al, 2014)
There is also evidence—along with pathways—that show how certain social activities work to lower stress and BP, including participation at church (Livingstone, Devine, and Moore, 1991). Black Americans can make other lifestyle changes in order to decrease BP, such as exercise and other lifestyle interventions. Redman, Baer, and Hicks state that “gene-environment interactions, job-related stress, racism, and other psychosocial factors to racial/ethnic disparities” need to be explored as causes for higher rates of HTN in blacks compared to whites. And with the knowledge of how all physiological systems work in terms of stress and other factors, should be explored as causes for this disparity.
Grim et al (1990) state that factors that influence high BP in blacks compared to whites are inherited and that is the major source of variation between these populations. However, the other mounting social/physiological evidence deserves an explanation; that is not inherited, and what we know about how our physiology responds to stress and discrimination—whether real or imagined—are extremely important and lead to extremely real, and important, outcomes in these populations. It is also argued that since blacks en route to America during the slave trade died from salt-depletive diseases, that blacks with a higher genetic propensity to absorb salt survived and this is why blacks have a higher propensity to absorb salt and are more ‘salt-sensitive’, which also could explain higher rates of HTN in American blacks compared to their cousins in Africa (Wilson and Grim, 1991). However, Curtin (1992) disputes this because “There is no evidence that diet or the resulting patterns of disease and demography among slaves in the American South were significantly different from those of other poor southerners”.
However, in regards to the social environment, Williams (1992) drives one of the best arguments I have encountered in this literature so far, stating that while genetic factors play a small part in regards to the BP gap between blacks and whites, social factors are arguably more important than genetic ones (and with our homeodynamic physiology, this does make sense). Dressler (1990) for instance, argues that skin color is a proxy for both social class and discrimination and these factors explain a large amount of the variation. Psychosocial variables can also explain heightened BP (Marmot, 1985; Cuffee et al, 2014). Yan et al (2003) also note how “time urgency/impatience” and “hostility” “were associated with a dose-response increase in the long-term risk of hypertension.” Henry (1988) also argues that calcium, obesity and genetic factors cannot be the aetiology of HTN in blacks, while also proposing that high sodium intakes are due to psychosocial stress (Williams, 1992: 136).
Obesity also leads to hypertension (Re, 2009) while blacks are more likely to be obese than whites, however, black American men with more African ancestry are less likely to be obese (Klimentidis et al, 2016). This would imply that the greater amount of African ancestry in American blacks both protects against obesity and along with it HTN. Williams (1992) makes a convincing argument that environmental and social factors are the cause for the black-white BP gap. And while genetic factors are important, no doubt, environmental and social factors are arguably more important to this debate.
Kulkarni et al (1998) show that increased stress leads to subsequent BP elevations which, over time, will lead to HTN. In a 2009 meta-analysis, Gasparin et al show how “individuals who had stronger responses to stressor tasks were 21% more likely to develop blood pressure increase when compared to those with less strong responses.”
Further, in support for the ‘perceived stress’ hypothesis in regards to blacks ‘perceiving’ stress and discrimination, “stress denial in combination with abdominal obesity, alcohol consumption, and smoking may be proxy for a high stress level” (Suter et al, 1997). Carroll et al (2001) also show how there are is “modest support for the hypothesis that heightened blood pressure reactions to mental stress contribute to the development of high blood pressure.” Sparrenberger et al (2009) also did a systematic review of observational studies, finding that “Acute stress is probably not a risk factor for hypertension. Chronic stress and particularly the non-adaptive response to stress are more likely causes of sustained elevation of blood pressure.”
Lastly, Langford (1981) shows that when SES is controlled for, the black-white BP disparity vanishes, implying that social and environmental—not genetic—factors are the cause for elevated HTN levels in black Americans. Sweet et al (2007) showed that for lighter-skinned blacks, as SES rose BP decreased while for darker-skinned blacks BP increased as SES did while implicating factors like ‘racism’ as the ultimate causes. This is solid evidence that both skin color and SES are predictors of higher prevalence of BP in black populations, and since other studies show that this is not noticed in higher class blacks, nor is this noticed in blacks in Africa, then the main causes of this disparity are social and environmental in nature.
(Non, Gravlee, and Mulligan, 2012). Their study suggests that educating black Americans on the dangers and preventative measures of high BP will reduce BP disparities between the races. This is in-line with Williams (1992) in that the social environment is the cause for the higher rates of BP. One hypothesis explored to explain why this effect with education was greater in blacks than whites was that BP-related factors, such as stress, poverty and racial discrimination (remember, even if no racial discrimination occurs, any so-called discrimination is in the eye of the beholder so that will contribute to a rise in physiologic variables) and maybe social isolation may be causes for this phenomenon. Future studies also must show how higher education causes lower BP, or if it only serves as other markers for the social environment. Nevertheless, this is an important study in our understanding of how and why the races differ in BP and it will go far to increase our understanding of this malady. This is a very convincing argument that education and not genetic ancestry cause disparities in BP between blacks and whites.
WebMD states that, of course, both environmental and genetic factors are at play in regards to black’s increased propensity for acquiring HTN. Fuchs (2011) also states that “They [environmental and behavioral factors] could act directly or by triggering mechanisms of blood pressure increase that are dormant in blacks living in Africa” and explain why black Americans have higher rates of BP than Africans in Africa. Further, race and ethnicity are independent predictors of HTN (Holmes et al, 2013).
Conclusion
Blacks and whites do differ in BP, and its aetiology is both complex and hard to untangle Genetic factors probably don’t account for a lot of this variance since Africans in Africa have low levels of BP compared to their black American cousins. Numerous lines of evidence shows that social and environmental factors are the cause, and so to change this, all people—especially blacks—should be educated on how to change these problems in our society. Whether discrimination is real or imagined, the effects of it lead to real physiological outcomes that then lead to increased health disparities between these populations.
