
Home » Posts tagged 'Race Realism'
Tag Archives: Race Realism
Reductionism, Natural Selection, and Hereditarianism
3400 words
Assertions derived from genetic reductionist ideas also ignore the abundant and burgeoning evidence that genes are outcomes of evolutionary processes and not bases of them. (Lerner, 2021: 449)
Genetic reductionism places (social) problems “in the genes” and so if these problems are “in the genes” then we can either (1) use gene therapy, (2) reduce the frequency of “the bad genes” in the population (eugenics) or (3) just live with these genetically caused problems. Social groups differ materially and they also differ genetically. To the gene-determinists, social positioning is genetically determined and it is due to a genetically determined intelligence. (See here for arguments against the claim.)
On the basis of heritability estimates derived from flawed methodologies like twin and adoption studies (Richardson and Norgate, 2005; Joseph, 2014; Burt and Simons, 2015; Moore and Shenk, 2016), hereditarians claim that traits like “IQ” (“intelligence”) are strongly genetically determined and if a trait is strongly genetically determined, then environmental interventions are doomed to fail (Jensen, 1969). Since IQ is said to have a heritability of .8, it is then claimed by the reductionist that environmental interventions are useless or near useless. Indeed, this was the conclusion of Jensen’s (1969) (infamous) paper—compensatory education has failed (an environmental intervention) and so the differences are genetic in nature.
Arguments like those have been forwarded for the better part of 100 years—and the arguments are false because they rely on false assumptions. The false assumptions are (1) that natural selection has caused trait differences between populations and (2) that genes are active—not passive—causes. (1) and (2) here can be combined for (3): genes that cause differences between groups were naturally selected and eventually fixed in the populations. This article will review some hereditarian thinking on natural selection and human variation, show how the theorizing is false, show how the theory of natural selection itself cannot possibly be true (Fodor and Piatteli-Palmarini, 2010) and finally will show that by accepting genetic reductionism we cannot achieve social justice since the causes of the social problems reduce to genes.
The ultimate claim from hereditarians is that human behavior, social life, and development can be reduced to—and explained by—genes. Social inequities are the target for social justice. Inequities refer to differences between groups that are avoidable and unjust. So the hereditarian attempts to reduce social ills to genes, thereby getting around what social justice activists want. They just reduce it to genes leading to possibilities (1)-(3) above. This has the possibility of being disastrous, for if we can fix the problems the hereditarians deem as “genetic”, then countless lives will not be made better.
Hereditarianism and natural selection
The crucial selection pressure responsible for the evolution of race differences in intelligence is identified as the temperate and cold environments of the northern hemisphere, imposing greater cognitive demands for survival and acting as selection pressures for greater intelligence. (Lynn, 2006: 135)
Hereditarians are neo-Darwininans and since they are neo-Darwinians, they hold that natural selection is the most powerful “mechanism” of evolution, causing trait changes by culling organisms with “bad” traits which then decreaes the frequency of the genes that supposedly cause the trait. But (1) natural selection cannot possibly be a mechanism as there is no agent of selection (that is, no mind selecting organisms with fitness-enhancing traits for a certain environment), nor are there laws of selection for trait fixation that hold across all ecologies (Fodor and Piattelli-Palmarini, 2010); and (2) genes aren’t causes of traits on their own—they are caused to give the information in them by and for the physiological system (Noble, 2011).
In his article Epistemological Objections to Materialism, in The Waning of Materialism, Koons (2010: 338) has an argument against natural selection with the same force as Fodor and Piattelli-Palmarini (2010):
The materialist must suppose that natural selection and operant conditioning work on a purely physical basis (without presupposing any prior designer or any prior intentionality of any kind). According to anti-Humean materialism, only microphysical properties can be causally efficacious. Nature cannot select a property unless that property is causally efficacious (in particular, it must causally contribute to survival and reproduction). However, few, if any, of the biological features that we all suppose to have functions (wings for flying, hearts for pumping bloods) constitute microphysical properties in a strict sense. All biological features (at least, all features above the molecular level) are physically realized in multiple ways (they consist of extensive disjunctions of exact physical properties). Such biological features, in the world of the anti-Humean materialist, don’t have effects—only their physical realizations do. Hence, the biological features can’t be selected. Since the exact physical realizations are rarely, if ever repeated in nature, they too cannot be selected. If the materialist responds by insisting that macrophysical properties can, in some loose and pragmatically useful way of speaking, be said to have real effects, the materialist has thereby returned to the Humean account, with the attendant difficulties described in the last sub-section. Hence, the materialist is caught in the dilemma.
We can grant that “nature” cannot select a trait if it isn’t causally efficacious. But combining Fodor’s argument with Koons’, if traits are linked then the fitness-enhancing trait cannot be directly selected-for since when you have one, you have the other. In any case, “natural selection” is part of the bedrock of hereditarian theorizing. It was natural selection—according to the hereditarian—that caused racial differences in behavior and “intelligence.” And so, if the hereditarian has no response to these two arguments against natural selection, then they cannot logically claim that the differences they describe are due to “natural selection.”
So the hereditarian theorist asserts that those with genes that conferred a fitness advantage had more children than those that didn’t which led to the selection of the genes that became fixed in certain populations. This is a familiar story—and the hereditarian uses this as a basis for the claim that racial differences in traits are the outcome of natural selection. These views are noted in Rushton (2000: 228-231), Jensen (1998: 170, 434-436) and Lynn (2006: Chapters 15, 16, and 17). But as Noble (2012) noted, there is no privileged level of causation—that is, before performing the relevant experiments, we cannot state that genes are causes of traits so this, too, refutes the hereditarian claim.
Rushton’s “Differential K” theory—where Mongoloids, Caucasians, and Africans differ on a suite of traits, which is influenced by their life histories and whether or not they are r- or K-strategists. Rushton (2000: 27) also claimed that “different environments cause, via natural selection, biological differences“, and by this he means that the environment acts as a filter. But the claim that the environment is the filter that causes variation in traits due to genes being “selected against” fails, too. When traits are correlated, the environmental filters (the mechanism by which selection theory purportedly works) cannot distinguish between causes of fitness and mere correlates of causes of fitness. So appealing to environments causing biological differences fails.
But unfortunately for hereditarians, a new analysis by Kevin Bird refutes the claim that natural selection is responsible for racial differences in “IQ” (Bird, 2021). So now, even assuming that genes can be selected-for their contribution to fitness and assuming that psychological traits can be genetically transmitted (which is false), hereditarianism still fails.
Hereditarianism and genetic reductionism
The ideology of IQ-ism is inherently reductionist. Behavioral geneticists, although they claim to be able to partition the relative contributions of genes and environment into nest little percentages, are also reductionists about “traits”—such as “IQ.” Further, if one is an IQ-ist then there is a good chance that they would fall into the reductionist camp of attempting to explain “intelligence” as being reducible to physiological brain states, and parts of the brain (such as Deary, 1996; Deary, Penke and Johnson, 2010; Jung and Haier, 2007; Haier, 2016; Deary, Cox, and Hill, 2021).
Reductionism can be simply stated as the parts have a sort of causal primacy over the whole. When it comes to psychological reduction, it is often assumed that genes would be the ultimate thing that it is reduced to, thereby, explaining how and why psychological traits differ between individuals—most importantly to the IQ-ist, “intelligence.” Behavioral geneticists have been reductionists since the field’s inception which has carried over to the present day (Panofsky, 2014). Even now, in the 3rd decade of the 2020s, reductionist accounts of behavior and psychology are still being pushed and the attempted reduction is reduction to genes. Now, this does not mean that environmental reduction has primacy—although we can and have identified environmental insults that do impede the ontogeny of certain traits.
Deary, Cox, and Hill (2021) argue for a “systems biology” approach to the study of “intelligence.” They review GWAS studies, neuroimaging studies and attempt a to lay the groundwork for a “mechanistic account” of intelligence, attempting to pick up where Jung and Haier (2007) left off. Unfortunately, the claims they make about GWAS fail (Richardson and Jones, 2020; Richardson, 2017b, 2021) and so do the claims they make about neuroreduction (Uttal, 2012).
This kind of genetic reductionism for psychological traits—along social ills such as addiction, violence, etc—then becomes ideological, in thinking that genes can explain how and why we have these kinds of problems. Indeed, this was why the first “IQ” tests were translated and brought to America—to screen and bar immigrants the IQ-ists saw as “feebleminded” (Richardson, 2003, 2011; Allen, 2006; Dolmage, 2018). Such tests were also used to sterilize people in the name of a eugenic ideology that was said to be for the betterment of society (Wilson, 2017). Thus, when such kinds of reductionism are applied to society and become an ideology, we definitely can see how such pseudoscientific beliefs can manifest itself in negative outcomes for the populace.
Ladner (2020:10) “constructed an economic analysis grounded in evolutionary biology.” Ladner claims that “Natural Selection is the main force that determines economic behavior.” Ladner claims that socialism will always fail since authoritarian regimes stifle our selfish proclivities while capitalism is grounded in selfishness and greed and so will always prevail over socialism. This is quite the unique argument… Of course Dawkins gets cited since Ladner is talking about selfishness, and these selfish genes are what cause the selfish behavior that allow capitalism to flourish. But the claim that genes are selfish is not a physiologically testable hypothesis (Noble, 2011) and DNA can’t be regarded as a replicator that’s independent from the cell (Noble, 2018). In any case, the argument in this book is that inequality is due to natural selection and there isn’t much we can do about capitalism since genes make us selfish and capitalism is all about selfishness. But being too selfish leads to such huge wealth inequalities we see in America today. The argument is pretty novel but it fails since it is a just-so story and the claims about “natural selection” are false.
(See Kampourakis, 2017 and Richardson, 2017a for a great overview on what genes are and what they really do.)
Hereditarianism and mind-brain identity
Pairing hereditarianism with physicalism about the brain is an implicit assumption of the theory. Ever since our the power of our neuroimaging methods have increased since the beginning of the new millennium, many studies have come out correlating different psychological traits with different brain states. Processes of the mind, to the mind-brain identity theorist, are identical to states and processes of the brain (Smart, 2000). And in the past two decades, studies correlating physiological brain states and psychological traits have increased in number.
The leading theorists here are Haier and Jung with their P-FIT model. P-FIT stands for the Parieto-Frontal Integration Theory which first proposed by Jung and Haier (2007) who analyzed 37 neuroimaging studies. This, they claim, will “articulate a biology of intelligence.” (Also see Colom et al, 2009.) Again, correlations are expected but we can’t then claim that the brain states cause the trait (in this case, “IQ.” (See Klein, 2009 for a primer on the philosophical issues in neuroimaging.)
But in 2012 psychologist William Uttal published his book Reliability in Cognitive Neuroscience: A Meta-meta Analysis where he argues that pooling these kinds of studies for a meta-analysis (exactly what Jung and Haier (2007) did) “could lead to grossly distorted interpretations that could deviate greatly from the actual biological function of an individual brain.” Pooling multiple studies from different individuals taken at different times of the day under different conditions would lead to a wide variation in physiologies, nevermind the fact that motion artifacts can influence neuroimages, and it emotion and cognition are intertwined (Richardson, 2017a: 193).
The point is, we cannot pool together these types of studies in attempt to localize cognitive processes to states of the brain. This is exactly what P-FIT does (or attempts to do). In any case, the correlations found by Jung and Haier (2007) can be explained by experience. IQ tests are experience-dependent (that is, one must be exposed to the knowledge on the test and they must be familiar with test-taking), and so too are parts of the brain that change based on what the person experiences. We cannot say that the physiological states are the cause of the IQ score—since the items on the test are more likely to be found in the middle-class, they would then be more prepared for test-taking.
Socially disastrous claims
Views from the likes of Robert Plomin—that there’s “not much we can do” about “environmental effects” (Plomin, 2018: 174)—are socially disastrous. If such ideas become mainstream then we may desist with programs that actually help people, on the basis that “it doesn’t work.” But this claim, that environmental effects are “unsystematic and unstable” are derived from conclusions based largely on twin studies. So whatever variance is left is attributed to the environment. (Do note, though, the Plomin’s claim that DNA is a blueprint is false.)
Hereditarians like Plomin then claim that environmental effects derive from one’s genotype so in actuality environmental effects are genetic effects—this is called “genetic nurture.” By using this new concept, the reductionist can skirt around environmental effects and claim that the effect itself is genetic even though it’s environmental in nature. Genes, in this concept, are active causes, actively causing parental behavior. So genes cause parental behavior which then influences how parents treat/parent their children. In this way, behavioral geneticists can claim that environmental effects are genetic effects too. (This is like Joseph’s (2014) Argument A in its circularity.)
By applying and accepting genetic reductionist claims, we rob people of certain life chances and we don’t commit ourselves to social justice. Of course, to the hereditarian, since the environment doesn’t matter then genes do. So we need to look at society from the gene-view. But this view betrays how and why our current social structures are the way they are. “IQ” tests were originally created to show that the current social hierarchy is the “right one” and the hereditarian believes to have shown that the hierarchy is “genetic” and so each group has their place on the social hierarchy on the basis of IQ scores which reduce to genes (Mensh and Mensh, 1991).
But humans are social creatures and although hereditarians attempt to reduce human social life to genes (in a circular manner), they fail. And their failing has led to the destruction of thousands of lives (see the sterilizations in America during the 1900s and around the world eg in Cohen, 2016 and Wilson, 2017). Reductionist attempts of social behavior to genes have been tried over the past 20 years (e.g., Jensen, 1998; Rushton, 2000, Lynn, 2006; Hart, 2007) but they all fail (Lerner, 2018, 2021). Social (environmental) changes cannot undo what the genes have “set” in individuals and so, we need not pour money into social programs.
For instance, many hereditarians and criminologists have espoused eugenic views, like Jensen’s claim that welfare could lead to the genetic enslavement of a part of the population (Jensen, 1969: 95) and that we can “estimate a person’s genetic standing on intelligence” based on their IQ score (Jensen, 1970: 13) to name two things. It is no surprise to me that people who hold such reductionist views of genes and society that they would also hold eugenic views like these. It is, in fact, a logical endpoint of hereditarianism—“phasing out” populations, as Lynn described in his review of Cattell’s Beyondism (see Tucker, 2009).
The answer to hereditarianism
Since we have to reject hereditarianism, then the answer to hereditarian dogma is relational developmental systems (RDS) theory which emphasizes the actions of all developmental resources, not reducing development to one primary developmental resource as hereditarians do. Similar things have been noted by other developmental systems theorists, most notably Oyama (1985/2000). What is selected aren’t genes, or behaviors. What is actually selected are the whole developmental system. Genes aren’t active causes. So if we look at development as a dance with music, as Noble (2006, 2016) does, there are no sufficient causes for development, but there are necessary causes of which genes are but one part of the whole system.
The answer to hereditarianism is to simply show that it fails conceptually, it’s “causal” framework for explaining the differences is unsound (“natural selection”) and to show that multiple interacting factors are responsible for human development in the womb and throughout the life course. “Theories derived from RDS meta-theory focus on the “rules,” the processes that govern, or regulate, exchanges between (the functioning of) individuals and their contexts” (Lerner, 2021: 457). Hereditarianism relies on gene-selectionism. But genes are not leaders in evolution; development is inherently holistic, not reductonist.
Conclusion
The hereditarian program has its beginnings with Francis Galton and then after the first “IQ” test was made (Binet’s), American eugenicists used it to “show” who was a “moron” (meaning, who had a low “IQ” meaning “intelligence”). Tens of thousands of sterilizations were soon carried out since the causes of these problems were in these people’s genes and so, negative eugenics needed to be practiced in order to cull the population of these genes that lead to socially undesirable traits.
The hereditarian hypothesis is, therefore, a racist hypothesis, contra Carl (2019) who argued the hereditarian hypothesis is not racist while citing many arguments from critics. I won’t get into that here, as I have many articles on the matter that Carl (2019) discusses. But what I will say is that the hereditarian hypothesis is racist in virtue of (1) not being logically plausible (reductionism about the mind and physicalism are both false) and (2) the hypothesis ranks races on a scale of “higher to lower” (that is, a hierarchy). Racism “is a system of ranking human beings for the purpose of gaining and justifying an unequal distribution of political and economic power” (Lovechik, 2018). Therefore, the hereditarian hypothesis is a racist hypothesis, contra Carl’s protestations. Hereditarians may claim that their claims are stifled in the public debate, but for behavioral genetics at large, this is false (see Kampourakis, 2017). Carl (2018) claims that “stifling” the debate around race, genes, and IQ can do harm but he is sadly mistaken! By believing that differences that can be changed are “genetic”, they are deemed to be unfixable and the groups who have a higher frequency of which ever genes that are causally efficacious (supposedly) for IQ will then be treated differently.
If neuroreduction (mind-brain reduction) is false, if genetic reduction is false, and if natural selection isn’t a mechanism, then hereditarianism cannot possibly be true, and if heritability . The arguments given here go well with my conceptual arguments against hereditarianism for more force against the hereditarian hypothesis. Just like with my argument to ban IQ tests, we must ban hereditarian research too, since the outcomes can be socially disastrous (Lerner, 2021 part VI, Developmental Theory and the Promotion of Social Justice). By now, these kinds of “theories” and claims have been refuted to hell and back, and so, the only reason to hold these kinds of beliefs is due to racist attitudes (combined with some mental gymnastics).
So for these, and many more, reasons, we must outright reject genetic reductionism (not least because these claims derive from flawed studies with false assumptions like twin studies) along with its partner “natural selection.” We therefore must commit ourselves to social justice to ameliorate the effects of racist attitudes and views.
White Privilege: What It Is and Who Has It?
2550 words
Discussions about whiteness and privilege have become more and more common. Whites, it is argued, have a form of unearned societal privilege which therefore explains certain gaps between whites and non-whites. White privilege is the privilege that whites have in society—this type of privilege does not have to be in America, it can hold for groups that are viewed as ‘white’ in other countries. This, then, perpetrates social views of race, hence these people are realists about race but in a social/political context and do not have to recognize race as biological (although race can become biologicized through social/cultural practices). This article will discuss (1) What white privilege is; (2) Who has white privilege; (3) Arguments against white privilege; and (4) If race doesn’t exist, why does white privilege matter?
What is white privilege?
The concept of white privilege, like most concepts, evolves with the times and current social thought. The concept was originally created in order to account for whites’ (unearned) privileges and the conscious bias that went into creating and then maintaining these privileges, to unconscious favoritism/psychological advantages that whites give other whites (Bennett, 2012: 75). That is, white privilege is “an invisible package of unearned assets that I can count on cashing in each day, but about which I was “meant” to remain oblivious. White privilege is like an invisible weightless knapsack of special provisions, maps, passports, codebooks, visas, clothes, tools , and blank checks” (McIntosh, 1988).
More easily, we can say that white privilege is—the privilege conferred, either consciously or subconsciously, to one based on their skin color or, as Sullivan (2016, 2019) argues, their class status ALONG WITH their whiteness is what we should be talking about—white privilege with CLASS in between ‘white’ and ‘privilege’. In this sense, one’s class status AND their whiteness is explanatory, not only the concept of whiteness (i.e., their socialrace). The concept of whiteness—one’s skin color—as the privilege leaves out numerous intricacies in how whiteness gives and upholds systemic discrimination. When we add the concept of ‘class’ into ‘white privilege’ we get what Sullivan terms ‘white class privilege’.
While yes, one’s race is an important variable in whether or not they have certain privileges, such privileges are held for middle- to upper-middle class whites. Thus, numerous examples of ‘white privilege’ are better understood as examples of ‘white class privilege’, since lower-class whites don’t have the same kinds of privileges, outlooks, and social status as middle- and upper-middle class whites. Of course, though, lower-class whites can benefit from their whiteness—they definitely can. But the force of Sullivan’s concept of ‘white class privilege’ is this: white privilege is not monolithic towards whites, and some non-whites are better-off (economically and in regard to health) than whites. Thus, according to Sullivan, ‘white privilege’ should be amended to ‘white class privilege’.
Who has white privilege?
Lower-class whites could, in a way, be treated differently than middle- and upper-class whites—even though they are of the same race. Lower-class whites can be seen to have ‘white privilege’ on the basis of everyday thought, since most think of the privilege as down to just skin color, yet there is an untalked about class dimension at play here, which, then, even gives blacks an advantage while upholding the privilege of the upper-class whites.
Non-whites who have are of a higher social class than whites would also receive different treatment. Sullivan states that the revised concept of ‘white class privilege’ must be used intersectionally—that is, privilege must be considered interacting with class, gender, national, and other social experiences. Sure, lower-class whites may be treated differently than higher-class blacks in certain contexts, but this does not mean that the lower-class white has ‘more privilege’ than the upper-class black. This shows that we should not assume that lower-class whites have the same kinds of privilege conferred by society as middle- and upper-class whites. Upper-class blacks and ‘Hispanics‘ may attempt to distinguish themselves from lower-class blacks and ‘Hispanics’, as Sullivan (2019: 18-19) explains:
Class privilege shows up as a feature of most if not all racial groups in which members with “more”—more money, education, or whatever else is valued in society—are treated better than those with “less.” For that reason, we might think that white class privilege actually is an intragroup pattern of advantage and disadvantage among whites, rather than an intergroup pattern that gives white people a leg up over non-white people. After all, many Black middle-class and upper-middle-class Americans also go to great lengths to make sure that they are not mistaken for the Black poor in public spaces: when they are shopping, working, walking, or driving in town, and so on (Lacy, 2007). A similar pattern can be found with middle-to-upper-class Hispanic/Latinx people in the United States, who can “protect” themselves from being seen as illegal immigrants by ensuring that they are not identified as poor (Masuoka and Junn, 2013).
Sullivan then goes on to state that these situations are not equivalent, since wealth, fame, and education do not protect upper-class blacks from racial discrimination. The certain privileges that upper-class whites have, thusly, do not transfer to upper-class blacks. Further, middle- to upper-class whites distinguish themselves as ‘good whites’ who are not racist, while dumping all of the racism accusations on lower-class whites. “…the line between “good” and “bad” white people drawn by many (good) white people is heavily classed. Good white people tend to be middle-to-upper-class, and they often dump responsibility for racism onto lower-class white people” (Sullivan, 2019: 35). Even though the lower-class whites get used as a ‘shield’, so to speak, by upper-class whites, they still have some semblance of white privilege, in that they are not assumed to be non-citizens to the US—something that ‘Hispanics’ do have to deal with (no matter their race).
While wealthy white people generally have more affordances than poor white people do, in a society that prizes whiteness all white people have some racial affordances, at least some of the time.
Paradoxically, whites are not the only ones that benefit off of ‘white privilege’—even non-whites can benefit, though it ultimately helps upper-class whites. They can benefit by being brought up in a white home, around whites (like being adopted or having one white parent while spending most of their childhood with their white family). Thus, white privilege can cross racial lines all the while still benefitting whites.
Sullivan (2019: chapter 2) discusses some blacks who benefit from white privilege. One of the people she discusses has a white parent. This is what gives her her lighter skin, but that is not where her privilege comes from (think colorism in the black community where lighter skin is more prized than darker skin). Her privilege came from “her implicit knowledge of white norms, sensibilities, and ways of doing things that came from living with and being accepted by white family members” (Sullivan, 2019: 26). This is what Sullivan calls “family familiarity” and is one of the ways that blacks can benefit from white privilege. Another way in which blacks can benefit from white privilege is due to “ancestral ties to whiteness.”
Colorism is the discrimination within the black community by skin color. Certain blacks may talk about “light-” and “dark-skinned” blacks and they may—ironically or not—discriminate on the basis of skin color. Such colorism is even somewhat instilled in the black community—where darker-skinned black sons and lighter-skinned black daughters report higher-quality parenting. Landor et al (2014) report that their “findings provide evidence that parents may have internalized this gendered colorism and as a result, either consciously or unconsciously, display higher quality of parenting to their lighter skin daughters and darker skin sons.” Thus, even certain blacks—in virtue of being ‘part white’—would benefit from white (skin) privilege within their own (black) community, which would therefore give them certain advantages.
Arguments against white privilege
Two recent articles with arguments against white privilege (Why White Privilege Is Wrong — Quillette and The Fallacy of White Privilege — and How It Is Corroding Society) erroneously argue that since other minority groups quickly rose up upon arrival to America, therefore white privilege is a myth. These kinds of takes, though, are quite confused. It does not follow that since other groups have risen upon entry into America and that since whites have worse outcomes on some—and not other—health outcomes, that therefore the concept of white privilege is ‘fallacious’; we just need something more fine-grained.
For example, the claims that X minority group is over-represented compared to whites in America gets used as a point that ‘white privilege’ does not exist (e.g., Avora’s article). Avora discusses the experiences and data from many black immigrants, proclaiming:
These facts challenge the prevailing progressive notion that America’s institutions are built to universally favor whites and “oppress” minorities or blacks. On the whole, whatever “systemic racism” exists appears to be incredibly ineffectual, or even nonexistent, given the multitude of groups who consistently eclipse whites.
How does that follow? In fact, how does the whole discussion of, for example, Japanese now outperforming whites follow that white privilege therefore is a ‘fallacy’? I ask the question, since Asian immigrants to America are hyper-selected (Noam, 2014; Zhou and Lee, 2017), meaning that what explains higher Asian academic achievement is academic effort (Hsin and Xie, 2014) and the fact that Asians are hyper-selected—meaning that they have a higher chance of having a higher degree.
The educational credentials of these recent [Asian] arrivals are striking. More than six-in-ten (61%) adults ages 25 to 64 who have come from Asia in recent years have at least a bachelor’s degree. This is double the share among recent non-Asian arrivals, and almost surely makes the recent Asian arrivals the most highly educated cohort of immigrants in U.S. history.
Compared with the educational attainment of the population in their country of origin, recent Asian immigrants also stand out as a select group. For example, about 27% of adults ages 25 to 64 in South Korea and 25% in Japan have a bachelor’s degree or more.2 In contrast, nearly 70% of comparably aged recent immigrants from these two countries have at least a bachelor’s degree. (The Rise of Asian Americans)
Avora even discuses some African immigrants, namely Nigerians and Ghanaians. However, just like Asian immigrants to America, Nigerian and Ghanaian immigrants to America are more likely to hold advanced degrees, signifying that they are indeed hyper-selected in comparison to the population that they derive from (Duvivier, Burch, and Boulet, 2017). Thus, to go along with the stats that Avora cites on the children of Nigerian immigrants, their parents already had higher degrees, signifying that they are indeed a hyper-selected group. This means that such ethnic groups cannot be used to show that white privilege is explanatory.
While Avora does discuss “class” in his article, he shows that it’s not only ‘white privilege’, but the class element that comes along with whiteness in America. He therefore unknowingly shows that once you add the ‘class’ factor and create the concept of ‘white class privilege’, that this privilege can cross racial lines and benefit non-whites.
In the Harinam and Henderson Quillette article, they argue that since there are some things that we say are ‘good’ that non-whites have more of than whites, therefore the concept of ‘white privilege’ does not explain the existence of disparities between ethnic groups in the US since some some bad things happen to whites and some good things happen to non-whites—but this is an oversimplification. The fact of the matter is, whites that do receive privileges over other ethnic/racial groups do so not in virtue of their (white) skin privilege, but in virtue of their class privilege. This can be seen with the above citations on class being the explanatory variable regarding Asian academic success (showing how class values get reproduced in the new country which then explains the academic success of Asians in America).
The fact that both of these articles believe that by showing some minority groups in America have more ‘good’ things than whites or better outcomes for bad things (like suicides) misses the point. That whites kill themselves more than other American ethnic groups does not mean that whites do not have privilege in America compared to other groups.
If race doesn’t exist, then why does white privilege matter?
Lastly, those who argue against the concept of white privilege may say that those who are against the concept of white privilege would then at the same time say that race—and therefore whites—do not exist so, in effect, what are they talking about if ‘whites’ don’t exist because race does not exist? This is of course a ridiculous statement. One can indeed reject claims from biological racial realists and believe that race exists and is a socially constructed reality. Thus, one can reject the claim that there is a ‘biological’ European race, and they can accept the claim that there is an ever-changing ‘white’ race, in which groups get added or subtracted based on current social thought (e.g., the Irish, Italians, Jews), changing with how society views certain groups.
Though, it is perfectly possible for race to exist socially and not biologically. So the social creation of races affords the arbitrarily-created racial groups to be in certain areas on the hierarchy of races. Roberts (2011: 15) states that “Race is not a biological category that is politically charged. It is a political category that has been disguised as a biological one.” She argues that we are not biologically separated into races, we are politically separated into them, signifying race as a political construct. Most people believe that the claim “Race is a social construct” means that “Race does not exist.” However, that would be ridiculous. The social constructivist just believes that society divides people into races based on how we look (i.e., how we are born) and then society divides us into races on the basis of how we look. So society takes the phenotype and creates races out of differences which then correlate with certain continents.
So, there is no contradiction in the claim that “Race does not exist” and the claim that “Whites have certain unearned privileges over other groups.” Being an antirealist about biological race does not mean that one is an antirealist about socialraces. Thus, one can believe that whites have certain privileges over other groups, all the while being antirealists about biological races (saying that “Races don’t exist biologically”).
Conclusion
In this article I have explained what white privilege is and who has it. I have also discussed arguments against white privilege and claims that those who argue against race are hypocrites since they still talk about “whites” while claiming that race exists. After showing the conceptual confusions that people have about white privilege, along with the fact that groups that do better than whites in America (the groups that supposedly show that white privilege is “a fallacy”), I then forward Sulllivan’s (2016, 2019) argument on white class privilege. This shows that their whiteness is not the sole reason why they prosper—their whiteness along with their middle-to-upper-middle-class status explains why they prosper. It also, furthermore, shows that while lower-class whites do have some sort of white privilege, they do not have all of the affordances of white privilege due to their class status. Blacks can, too, benefit from white privilege, whether it’s due to their proximity to whiteness or their ancestral heritage.
White privilege does exist, but to fully understand it, we must add in the nexus of class with it.
The Nike Air Native N7—Should We Market Shoes to Specific Demographics?
1400 words
It has come to my attention that near the end of 2007, Nike boasted about releasing a running shoe that specifically targeted Native American communities. Nike developed the shoe to “to address the specific fit and width requirements for the Native American foot.” Since Native Americans have a high rate of obesity and diabetes (“diabesity”), then it seems that it would be a good thing to promote a shoe specifically for and to the population in question. But do such gestures translate to racist ideas or do they translate to a corporation wanting to be seen as promoting health (while their ultimate goal is profit)? Nike, specializing in athletic clothing, surely would be a good organization to spearhead such a movement, right? On what research is this initiative based on and does it hold water?
Through such outreach programs, Nike hopes to be seen to make social and community impacts when it comes to health. As Welch (2019: 12) notes, the N7 imitative hopes “to further promote sport and physical activity in Native American communities.” Such programs and specific items that would catch the eye of the consumers in question to heighten their physical activity and, subsequently, lessen their rates of fatal diseases, should be seen as a good thing, which would be irrespective of the feelings of the groups in question who see such outreach as racist.
The shoe was developed by a podiatrist named Rodney Stapp who served the Native community for his whole life (b. 1961, d. 2016). This was the first—and since then, only—time that Nike developed a shoe for a specific racial group. Was it a good idea? Was it racist? Even if it could be construed as racist, wouldn’t it be negated by targeting a group that has some of the highest rates of diabesity in America, therefore leading to a more active population and mitigation of the diseases in question? (See Broussard et al, 1991; Narayan, 1996; Acton et al, 2002). Since exercise seems to be necessary in managing diabetes and its symptoms (Colberg et al, 2010; Kirwan, Sacks, and Nieuwoudt, 2018; Borhade and Singh, 2020), then it seems that, irrespective of whether or not such gestures are racist, that such outreach and initiatives are a net good for the population in question.
Stapp was a big-name figure in the outreach to Native groups in Texas, and was the podiatrist that Nike consulted with in the development of their Nike Native American N7 shoe. Stapp was the one that contacted Nike to make such a shoe, since the patients that he serviced did not like the black and bulky shoes that were specially developed for diabetics—the efficacy of such shoes, though, have been debated in the literature (e.g., Brunner, 2015), while others have noted that diabetics have stated that the style and appearance of such diabetic shoes are the reasons why there is such low compliance in wearing them (Macfarlane and Jensen, 2003). In any case, wouldn’t marketing shoes toward specific demographics be a net-good, irrespective of the ultimate goals of the company if they would then promote healthier behaviors in the population in question?
Nike, though, has been criticized for the initiative, with Native right’s groups claiming that Nike is using Native plight for profit (Cole, 2008; Sanders, Phillips, and Alexander, 2018). It has been criticized by such groups since they have embroidered the shoe with feathers and sunsets, arrows, and different kinds of symbolism prevalent in Native cultures in the Americas. Here, I would not say that such things are racist on its face, it’s just a marketing ploy to sell more of their shoes. While it can be construed that such marketing is racist in a way, I think that the good such a program and shoe would do to reach at-risk populations outweighs any racist connotations that the shoes and the outreach program makes.
But most would have a problem with the claim that the shoe was developed specifically for “Native American feet”. Stapp claimed that “Indians tend to have a wider foot, but their heels are about average“, which would indicate slippage while running in a normal running shoe. Nike’s press release on the shoe says that “A strong emphasis was placed on providing a performance product that would cater to the specific needs of Native American foot shapes and help provide motivation to Native Americans predisposed to, or suffering from, health issues that can be improved by leading physically active lifestyles“, while also stating that “Research has engaged individuals from over 70 tribes as well as consulting podiatrists and members of Indian Health Services and the National Indian Health Board.“
(I am unable to find the research in question; hopefully someone can point me in the right direction so that I can find it.)
There is a history of such differences in the appendages between North and South Native Americans—where North Americans have longer and more slender feet than South Americans (e.g., Kate, 1918). Nike stated that the reason they developed the shoes were so that they could accommodate Native American’s wider feet, along with combating the diabesity epidemic that affects them. In 2015, though, Stapp stated that he believed the introduction of the shoe dropped amputations from 5-6 per year to 0-1 per year. If it is indeed true that the shoes were related in lowering the incidences of foot amputations in Native communities, then it would seem that the cause of that would be that they are moving more and getting more blood to their lower extremities which would then lead to lowered rates of amputation in these diabetic populations.
The claim that such shoes “racially profile” Natives is ridiculous. Stapp said that Nike asked him if there were differences in the feet of Native groups compared to others to which he answered “Yes.” Apparently, around the time of the marketing for the shoes, Nike was told that Native Americans had problems fitting into Nike’s ‘normal’ running shoes due to the width of their feet (being wider than average). Along with Natives supposedly having wider feet, since diabetes causes inflammation of tissue, which is concentrated in the feet—for instance, with diabetic foot ulcers (Pendsey, 2010; Schoen and Norman, 2014; Tuttolomondo, Maida, and Pinto, 2015; Amin and Doupis, 2016)—would seem that the call for such shoes to be develop would be a net-good for the population.
Though I can see how the claims that the shoe targeted at a specific racial group could be construed as racist, the net-good that a shoe does in getting to certain populations would outweigh the negative connotations that the racist accusation brings on Nike. Indeed, some of the developers of the shoe were Native, worked with Natives, and developed it to specifically target and help Natives manage a debilitating disease that leads to many negative health outcomes—like foot amputation and eventually death. So if exercise is conducive to managing diabetes and diabetic foot and the N7’s would then target certain populations with different average foot morphology, then it seems that the shoe has been a net-good for the population since, according to Stapp, seven years after the introduction of the shoe diabetic foot amputations went from 5-7 to 0-1 per year. While he may have had financial incentives to say that, I don’t think that it underscores the fact that Nike’s N7 program did not have positive benefits—even if they could be construed in a negative way (i.e., claims of racism).
The answer to the question “Should we market shoes to specific demographics” is “Yes.” It would be a good idea to, for example, make more demographic-specific shoes with specific embroideries in order to attempt to target certain at-risk populations that are more likely to acquire certain diseases on the basis of physical inactivity—like the Nike N7 program and Nike Native American N7 shoe attempt to do. It is for these reasons, then (irrespective of whether or not such morphologic claims of the feet of Natives are true) that the initiative in question is a good thing. The moral “should” question on whether or not we “should” market things—in this example, shoes—to certain demographics seems to rest on whether or not the marketing would have a positive effect on the lifestyles of the groups in question. If it does have positive effects, then we should market such programs toward at-risk populations, irrespective of claims that such marketing is racist toward certain groups.
The Social Construction of the ‘Hispanic/Latino/Spanish’ Race
2600 words
In daily social discourse, you may have heard people referred to as ‘Hispanic/Latino/Spanish” (HLS) What does this mean? Are they referents of biological races or are they social only social terms? When we think of HLSs, we think of things that group HLSs together and most people, in the American context, think of HLSs as a race. In this article, I will discuss the social construction of the HLS race and what it means for race discourse in America.
Imagine you hear two people speaking Spanish. When you hear this and you are then asked a question about the two interlocutors, you may refer to them as “those Spanish people.” You are only referring to them as “Spanish people” since they are Speaking Spanish. Now, thinking of it in this way, the referent “Spanish”, like, say, “Italian” may refer to a particular ethnic group or it may not (depending on whether or not an individual has migrated to that country, had children and then raised them in the country they migrated to). However, when we—in our daily social discourse—refer to “Spanish people” are referring to many different countries and the people that inhabit them.
For example, we may refer to Dominicans as HLS, on the basis that they speak Spanish. Would they, then, be racially similar to Chileans or Argentinians despite the radically different histories of the countries in question? No. That they share a language does not indicate that the three countries in question are a part of this HLS—it does not mean that they are a biological race. However, these people from these countries that do speak Spanish—if they, say emigrate to America—may receive the same treatment as each other on the basis of the language they speak.
Take the identity group La Raza. La Raza means “the race” in English, and it refers to the countries that the Spanish invaded and then intermixed with the indigenous inhabitants of said country (and even African slaves they brought when the indigenous population began to dwindle). In this way, La Raza becomes a socially constructed race and what unites those people is the fact that they speak the same language and have similar cultural histories (having Spaniards invade their land and forcing their customs on them). They may say a phrase like “Viva La Raza” which translates to “Long live the race.”
If we are talking about “race” in the American sense, then we are talking about how the OMB uses the term “race” and what it means to Americans in American discourse. That we speak of certain social groups as “races” does not entail that we treat them as biological races. One may be of the same racial group and, a country or area where they are the minority they may then be treated differently on the basis of their customs/language.
Take Argentina. An Argentinean may have most of their ancestry derive from Southern Italy, since Argentina saw a large number of Southern Italian immigrants begin to emigrate in 1857, right as The Kingdom of Two Sicilies became Italy. In any case, if this Argentinian had all of their ancestry from Southern Italy, and, say, their parents emigrated from Argentina to America, then they would speak Spanish. On this basis alone, if this family were in, say, New York or New Jersey (two states with a high Italian population) and they spoke Spanish, the Italian population would discriminate against them on the basis of their language and culture and how it is not like their own. So even though they are of the same ethnic group, just the simple fact that their families emigrated to different countries before/after Italy’s ‘creation’ would mean that one group would discriminate against the other on the basis of cultural differences.
So take a La Raza proponent. On the basis of the phrase “Viva La Raza“, since “La Raza” refers to countries with historically Spaniard influence, they would then be referring to those countries that Spain had invaded in the 1500s. So, a Mexican La Raza proponent may be referring to, and even attempt to give special privilieges to, an Argentnian even if they are not of the same (biological) race. The Mexican La Raza proponent may, for example, owe most of his ancestry to the Aztecs while the Argentinean may of most of his ancestry to South Italians. Although they are of different racial groups, the Mexcian La Raza proponent may see the Argentinian as part of their race, and therefore would treat them differently than a Southern Italian-descended person who spoke English/Italian—meaning that even though the phenotype would (theoretically) be the same, on the basis of culture (i.e., language), the Mexican La Raza proponent would treat the Argentenian different from the American differently, even though they trace their ancestry back to the same place. So what matters for the La Raza proponent is shared language and certain shared aspects of what we term “Hispanic culture.”
In any case, the term “Spanish” refers to Spaniards—people from Spain. “Latino” refers to Latin America. “Hispanic” refers to people who speak Spanish. In this way, we see how all three terms coalesce into a socially constructed race. We can see this with the way certain HLS groups treat each other even though they are not the same race.
In the book White Privilege: Essential Readings on the Other Side of Racism, edited by Rothenberg (2008), Foley (2008) argues in his article Becoming Hispanic: Mexican Americans and Whiteness that Mexican Americans fought for the ‘Hispanic’ term, and so, wanted it to be an ethnic and not racial term. Mexican Americans wanted to be seen as ‘white’ in order to get over de jure segregation and so, from the 30s-50s, Mexicans argued that they were “white” and even supported the segregation of blacks and whites, on the basis of the claim that they too were white and would not have to be treated like blacks. Foley (2008: 56) cites some concerns of the League of United Latin American Citizens (LULAC) and what they thought their race should be in America:
In a 1932 article in the LULAC news titled “Are Texas-Mexicans ‘Americans’?” the author asserted that Mexican Americans were “the first white race to inhabit this vast empire of ours. Another member of LULAC boasted that Mexican Americans were “not only a part and parcel but as well as the sum of substance of the white race.” As self-constituted Whites, LULAC members considered it “an insult” to be associated with Blacks and other “colored” races. In 1936 a LULAC official deplored the practice of hiring “Negro musicians” to play at Mexican bailes (dances) because it led to “illicit relations” between Black men and “ill-informed Mexican girls.” He urged fellow LULAC members to “tell these Negroes that we are not going to permit our manhood and womanhood to mingle with them on an equal social basis.”
It was then in the 60s, however, when Mexican Americans began referring to themselves as ‘Chiacanos/Chicanas” and speaking of their skin color (brown), not trying to integrate into ‘whites.’ So, argues Foley (2008), Mexican Americans wanted “Hispanic” to be an ethnic and not racial category as they wanted to “keep their whiteness” but still be classified on the basis of language, too. It wasn’t until the 1960s that Mexican Americans began referring to “Aztlan” and “La Raza” and started referring to their browness while not trying to integrate with whites. So although Mexican Americans were de facto ‘white’ in America leading up to the Civil Rights Act (CRA) and were seen as ‘white’ in court cases/the census, they were rarely treated like ‘whites’ (Martinez, 1997; Foley, 2008; Ortiz and Telles, 2013).
Spencer (2019: 241) says “…the OMB is very clear … Hispanics are not a race.” Spencer (2019: 241) continues:
So, even if there are contexts in current American life where … Hispanics are treated as races by ordinary people, there are also contexts in current American life where … Hispanics are not treated as races by ordinary people, namely, when ordinary people use OMB race talk.
In agreeing with whites in the years leading up to the Civil Rights Act—that whites and blacks should be segregated—they wanted to then be seen as ‘white’ and so argued that the term ‘Hispanic’ should be an ethnic and not racial category. Funny how today, Mexican Americans are seen as the social term “people of color” (PoC) and are grouped in with black Americans.
But if we really think about what the HLS distinction really means, it refers to a sociolinguistic group. In this way, certain aspects of Central and South American culture may mirror what may be referred to as the ‘mother culture’ in Spain, as those country’s histories are strongly influenced by the Spaniard invasion post-1519. Though, the term ‘Hispanic’ arose in the 20th century as an ethnonym (a term given to an ethnic group), the terms ‘Hispanic’ and ‘Latino’ and even ‘Spanish’—in ordinary American race talk—refer to the same things. They are used as synonyms. Although the self-described ‘Hispanic’ may be a mixture of, say, three races, they attempt to not classify themselves with any of those races and racially mixed groups even though they themselves may be racially mixed.
This is put well by Quiros (2009: 14) who writes:
…women from Puerto Rican and Dominican ancestry with racially mixed backgrounds are often ascribed a single identity of Hispanic, Latina or even “Spanish,” thereby reducing them to a single ethnic category and silencing their complexities. Yet heterogeneity is also found within the Latino culture. Although Latinos may share some cultural similarities, Puerto Ricans, Dominicans, Mexicans, and Cubans, for example, have unique characteristics and histories that set them apart from one another.
These “unique characteristics” come from the ethnic groups the Spaniards brought to the countries in question, along with the types of customs and cultures that came to dominate in those countries. One may even say that the way a Latin American country ‘looks’ is a direct result of how long the Spaniards were there and what peoples (cultural groups) they brought to replace the dwindling indigenous peoples.
For instance, some Puerto Ricans and even Dominicans may speak of “Taino” heritage even though they may have little to no actual Taino heritage. The Taino are an indigenous people to the Carribean. They are what an American may term a ‘Native American’. Now, no matter how the Puerto Rican or Dominican looks, (say the Puerto Rican looks white and the Dominican looks black) they both may talk about this Taino heritage. For the Carribean Latin countries, this amount of ancestry may seem to bring them together and, along with speaking the same language they may believe that they are the same “people.” But, even though PR and DR are close together, they have radically different histories and phenotypes.
Wikipedia, for example, states this about Dominicans:
According to a 2015 genealogical DNA study of 27 Dominican individuals, their genetic makeup was estimated to be 52.15% European, 39.57% Sub-Saharan African, and 8.28% Native American and East Asian overall.[17]
In a 2014 population survey, 70.4% self-identified as mixed (mestizo/indio[a] 58%, mulatto 12.4%), 15.8% as black, 13.5% as white, and 0.3% as “other”.[18][19] A different survey in 2006 reported 67.6% mulatto and indio, 18.3% black, and 13.6% white.[20] However, according to the electoral roll completed in 1996, 82.5% of the adult population were indio, 7.55% white, 4.13% black, and 2.3% mulatto.[21] Historically there has been a reluctance to expressly identify African ancestry, with most identifying or being identified as mestizo or indio rather than mulatto or black.[21][22]
Regarding Puerto Rico, Wikipedia says:
One genetic study on the racial makeup of Puerto Ricans (including all races) found them to be roughly around 61% West Eurasian/North African (overwhelmingly of Spanish provenance), 27% Sub-Saharan African and 11% Native American.[211] Another genetic study from 2007, claimed that “the average genomewide individual (ie. Puerto Rican) ancestry proportions have been estimated as 66%, 18%, and 16%, for European, West African, and Native American, respectively.”[212] Another study estimates 63.7% European, 21.2% (Sub-Saharan) African, and 15.2% Native American; European ancestry is more prevalent in the West and in Central Puerto Rico, African in Eastern Puerto Rico, and Native American in Northern Puerto Rico.[213]
Although in the 2010 census about 61.4 percent of Puerto Ricans reported they were ‘white’. Meanwhile, only 13.5 percent of Dominicans reported they were ‘white’ while 70.4 percent claimed to be racially mixed (58 percent mestizo/indian and 12.4 percent mullato—which is mixed white and black; per Wikipedia).
So what the examples of Mexican Americans and whites and Puerto Ricans and Dominicans show is that although certain aspects of culture may be shared, the demographics of the countries in question are radically different—even the regions of the countries in question have radically different admixtures based on the history they have with Spain and even recent migrations into that area.
Conclusion
I have shown how HLS terms refer to social distinctions and not biological ones. The examples given with Mexican Americans and white Americans and Puerto Ricans and Dominicans show that, although these groups are held to be the same race, important distinctions exist between the two groups. So great, in both cases, that they cannot be the same “race” logically speaking, but they are the same socialrace. Using Hardimon’s socialrace terminology, HLSs are a socialrace and not a biological one. Although socialraces do have mirrored biological races, in this case, HLSs do not have one, as they are racially/ethnically distinct (as argued above). Hardimon (2017: 39) writes:
To deny that Latinos constitute a race is not to deny that individual Latinos or Latinos as a group can be the targets of racism (for example, owing to skin color). Nor is it to deny that Latinos are often regarded as “racially other” (as differing in some essential humanly important way corresponding to skin color) by members of other racialized groups (for example, Anglos). … Nor is it to deny that they constitute a socialrace in my sense of the term. Still less does it imply that Latinos ought not to aspire to a degree of solidarity connoted by the Spanish word raza.
Although in some studies “Hispanics” do cluster together (depending on where the dataset comes from), this does not mean that these clusters are races (a line of thought that Charles Murray would be married to on the basis of his assumption that clusters = races) (eg Tang et al, 2005). Though Risch et al (2002) write that “Hispanics, who represent a recently admixed group between Native American, Caucasian and African, did not form a distinct subgroup, but clustered variously with the other groups.”
The social construction of racial groups is a pretty complicated topic, and you will get different answers depending on who you are having the discussion with. So the answer to the question “Are HLSs a racial or ethnic group?” is “It depends.” If one is operating on the socialrace definition, then they are a racial group. If they are operating on a biological definition, then they are not a racial group, as they would cluster differently based on the majority admixture of that country (which has to do with the Spanish relationship with that country post-1519).
These groups are a good look into how we socially construct certain racial categories. What you see them as depends on what your previous leanings are. There is of course a social element to this, and this social element dictates what race a certain ethny/nationality would call themselves. Although people who speak the same language are socially the same ‘race’ since in American causal conversation they are referred to as one of the three synonyms (HLS), that does not mean that they are the same race, as evidenced with my Puerto Rican and Dominican examples. Thus, the “HLS” race is a social, not biological, construct as it refers to social, not biological, kinds.
Chinese IQ, Cheating, Immigrant Hyper-Selectivity and East Asian “Genetic Superiority”
2650 words
The East Asian race has been held up as what a high “IQ” population can do and, along with the correlation between IQ and standardized testing, “HBDers” claim that this is proof that East Asians are more “intelligent” than Europeans and Africans. Lynn (2006: 114) states that the average IQ of China is 103. There are many problems with such a claim, though. Not least because of the many reports of Chinese cheating on standardized tests. East Asians are claimed to be “genetically superior” to other races as regards IQ, but this claim fails.
Chinese IQ and cheating
Differences in IQ scores have been noted all over China (Lynn and Cheng, 2013), but generally, the consensus is, as a country, that Chinese IQ is 105 while in Singapore and Hong Kong it is 103 and 107 respectively (Lynn, 2006: 118). To explain the patterns of racial IQ scores, Lynn has proposed the Cold Winters theory (of which a considerable response has been mounted against it) which proposes that the harshness of the environment in the ice age selected-for higher ‘general intelligence’ in East Asian and European populations; such a hypothesis is valid to hereditarians since East Asian (“Mongoloids” as Lynn and Rushton call them) consistently score higher on IQ tests than Europeans (eg Lynn and Dzobion, 1979; Lynn, 1991; Herrnstein and Murray, 1994). In a recent editorial in Psych, Lynn (2019) criticizes this claim from Flynn (2019):
While northern Chinese may have been north of the Himalayas during the last Ice Age, the southern Chinese took a coastal route from Africa to China. They went along the Southern coast of the Middle East, India, and Southeast Asia before they arrived at the Yangzi. They never were subject to extreme cold.
In response, Lynn cites Frost’s (2019) article where he claims that “mean intelligence seems to have risen during recorded history at temperate latitudes in Europe and East Asia.” Just-so storytelling about how and why such “abilities” were “selected-for”, the Chinese score higher on standardized tests than whites and blacks, and this deserves an explanation (the Cold Winters Theory fails; it’s a just-so story).
Before continuing, something must be noted about Lynn and his Chinese IQ data. Lynn ignores numerous studies on Chinese IQ—Lynn would presumably say that he wants to test those in good conditions and so disregards those parts of China with bad environmental conditions (as he did with African IQs). Here is a collection of forty studies that Lynn did not refer to—some showing that, even in regions in China with optimum living conditions, IQs below 90 are found (Qian et al, 2005). How could Lynn miss so many of these studies if he has been reading into the matter and, presumably, keeping up with the latest findings in the field? The only answer to the question is that Richard Lynn is dishonest. (I can see PumpkinPerson claiming that “Lynn is old! It’s hard to search through and read every study!” to defend this.)
Although the Chinese are currently trying to stop cheating on standardized testing (even a possible seven-year prison sentence, if caught cheating, does not deter cheating), cheating on standardized tests in China and by the Chinese in America is rampant. The following is but a sample of what could be found doing a cursory search on the matter.
One of the most popular ways of cheating on standardized tests is to have another person take the exam for you—which is rampant in China. In one story, as reported by The Atlantic, students can hire “gunmen” to sit-in on tests for them, though measures are being taken to fight back against that such as voice recognition and finger-printing. It is well-known that much of the cheating on such tests are being done by international students.
Even on the PISA—which is used as an “IQ” proxy since they correlate highly (.89) (Lynn and Mikk, 2009)—though, there is cheating. For the PISA, each country is to select, at random, 5,000 of their 15-year-old children around the country and administer the PISA—they chose their biggest provinces which are packed with universities. Further, score flucuations attract attention which indicates dishonesty. In 2000, more than 2000 people protested outside of a university to protest a new law which banned cheating on tests.
The rift amounted to this: Metal detectors had been installed in schools to route out students carrying hearing or transmitting devices. More invigilators were hired to monitor the college entrance exam and patrol campus for people transmitting answers to students. Female students were patted down. In response, angry parents and students championed their right to cheat. Not cheating, they said, would put them at a disadvantage in a country where student cheating has become standard practice. “We want fairness. There is no fairness if you do not let us cheat,” they chanted. (Chinese students and their parents fight for the right to cheat)
Surely, with rampant cheating on standardized tests in China (and for Chinese Americans), we can trust the Chinese IQ numbers in light of the news that there is a culture of cheating on tests in China and in America.
“Genetic superiority” and immigrant hyper-selectivity
Strangely, some proponents of the concept of “genetic superiority” and “progressive evolution” still exist. PumpkinPerson is one of those proponents, writing articles with titles like “Genetically superior: Are East Asians more socially intelligent too?, More evidence that East Asians are genetically superior, Oriental populations: Genetically superior, even referring to a fictional character on a TV show as a “genetic superior.” Such fantastical delusions come from Rushton’s ridiculous claim that evolution may be progressive and that some populations are, therefore, “more evolved” than others:
One theoretical possibility is that evolution is progressive and that some populations are more “advanced” than others. Rushton, 1992
Such notions of “evolutionary progress” and “superiority“—even back in my “HBD” days—never passed the smell test to me. In any case, how can East Asians be said to be “genetically superior”? What do “superior genes” or a “superior genome” look like? This has been outright stated by, for example, Lynn (1977) who prolcaims—for the Japanese—that his “findings indicate a genuine superiority of the Japanese in general intelligence.” This claim, though, is refuted by the empirical data—what explains East Asian educational achievement is not “superior genes”, but the belief that education is paramount for upward social mobility, and so, to preempt discrimination, this would then be why East Asians overperform in school (Sue and Okazaki, 1990).
Furthermore, the academic achievement of Asian cannot be reduced to Asian culture—the fact that they are hyper-selected is why social class matters less for Asian Americans (Lee and Zhou, 2017).
These counterfactuals illustrate that there is nothing essential about Chinese or Asian culture that promotes exceptional educational outcomes, but, rather, is the result of a circular process unique to Asian immigrants in the United States. Asian immigrants to the United States are hyper-selected, which results in the transmission and recreation of middle-class specific cultural frames, institutions, and practices, including a strict success frame as well as an ethnic system of supplementary education to support the success frame for the second generation. Moreover, because of the hyper-selectivity of East Asian immigrants and the racialisation of Asians in the United States, stereotypes of Asian-American students are positive, leading to ‘stereotype promise’, which also boosts academic outcomes
Inequalities reproduce at both ends of the educational spectrum. Some students are assumed to be low-achievers and undeserving, tracked into remedial classes, and then ‘prove’ their low achievement. On the other hand, others are assumed to be high-achievers and deserving of meeting their potential (regardless of actual performance); they are tracked into high-level classes, offered help with their coursework, encouraged to set their sights on the most competitive four-year universities, and then rise to the occasion, thus ‘proving’ the initial presumption of their ability. These are the spill-over effects and social psychological consequences of the hyper-selectivity of contemporary Asian immigration to the United States. Combined with the direct effects, these explain why class matters less for Asian-Americans and help to produce exceptional academic outcomes. (Lee and Zhou, 2017)
The success of second-generation Chinese Americans has, too, been held up as more evidence that the Chinese are ‘superior’ in their mental abilities—being deemed ‘model minorities’ in America. However, in Spain, the story is different. First- and second-generation Chinese immigrants score lower than the native Spanish population on standardized tests. The ‘types’ of immigrants that have emigrated has been forwarded as an explanation for why there are differences in attainments of Asian populations. For example, Yiu (2013: 574) writes:
Yet, on the other side of the Atlantic, a strikingly different story about Chinese immigrants and their offspring – a vastly understudied group – emerges. Findings from this study show that Chinese youth in Spain have substantially lower educational ambitions and attainment than youth from every other nationality. This is corroborated by recently published statistics which show that only 20 percent of Chinese youth are enrolled in post-compulsory secondary education, the prerequisite level of schooling for university education, compared to 40 percent of the entire adolescent population and 30 percent of the immigrant youth population in Catalonia, a major immigrant destination in Spain (Generalitat de Catalunyan, 2010).
…
… but results from this study show that compositional differences across immigrant groups by class origins and education backgrounds, while substantial, do not fully account for why some groups have higher ambitions than others. Moreover, existing studies have pointed out that even among Chinese American youth from humble, working-class origins, their drive for academic success is still strong, most likely due to their parents’ and even co-ethnic communities’ high expectations for them (e.g., Kao, 1995; Louie, 2004; Kasinitz et al., 2008).
The Chinese in Spain believe that education is a closed opportunity and so, they allocate their energy elsewhere—into entrepreneurship (Yiu, 2013). So, instead of Asian parents pushing for education, they push for entrepreneurship. What this shows is that what the Chinese do is based on context and how they perceive how they will be looked at in the society that they emigrate to. US-born Chinese immigrants are shuttled toward higher education whereas in the Netherlands, the second-generation Chinese have lower educational attainment and the differences come down to national context (Noam, 2014). The Chinese in the U.S. are hyper-selected whereas the Chinese in Spain are not and this shows—the Chinese in the US have a high educational attainment whereas they have a low educational attainment in Spain and the Netherlands—in fact, the Chinese in Spain show lower educational attainment than other ethnic groups (Central Americans, Dominicans, Morrocans; Lee and Zhou, 2017: 2236) which, to Americans would be seen as a surprise
Second-generation Chinese parents match their intergenerational transmission of their ethnocultural emphasis on education to the needs of their national surroundings, which, naturally, affects their third-generation children differently. In the U.S., adaptation implies that parents accept the part of their ethnoculture that stresses educational achievement. (Noam, 2014: 53)
So what explains the higher educational attainment of Asians? A mixture of culture and immigrant (hyper-) selectivity along with the belief that education is paramount for upward mobility (Sue and Okazaki, 1990; Hsin and Xie, 2014; Lee and Zhou, 2017) and the fact that what a Chinese immigrant chooses to do is based on national context (Noam, 2014; Lee and Zhou, 2017). Poor Asians do indeed perform better on scholastic achievement tests than poor whites and poor ‘Hispanics’ (Hsin and Xie, 2014; Liu and Xie, 2016). Teachers even favor Asian American students, perceiving them to be brighter than other students. But what are assumed to be cultural values are actually class values which is due to the hyper-selectivity of Asian immigrants to America (Hsin, 2016).
The fact that the term “Mongoloid idiot” was coined for those with Down syndrome because they looked Asian is very telling (see Hilliard, 2012 for discussion). But, the IQ-ists switched from talking about Caucasian superiority to Asian superiority right as the East began their economic boom (Liberman, 2001). The fact that there were disparate “estimates” of skulls in these centuries points to the fact such “scientific observations” are painted with a cultural brush. See eg table 1 from Lieberman (2001):
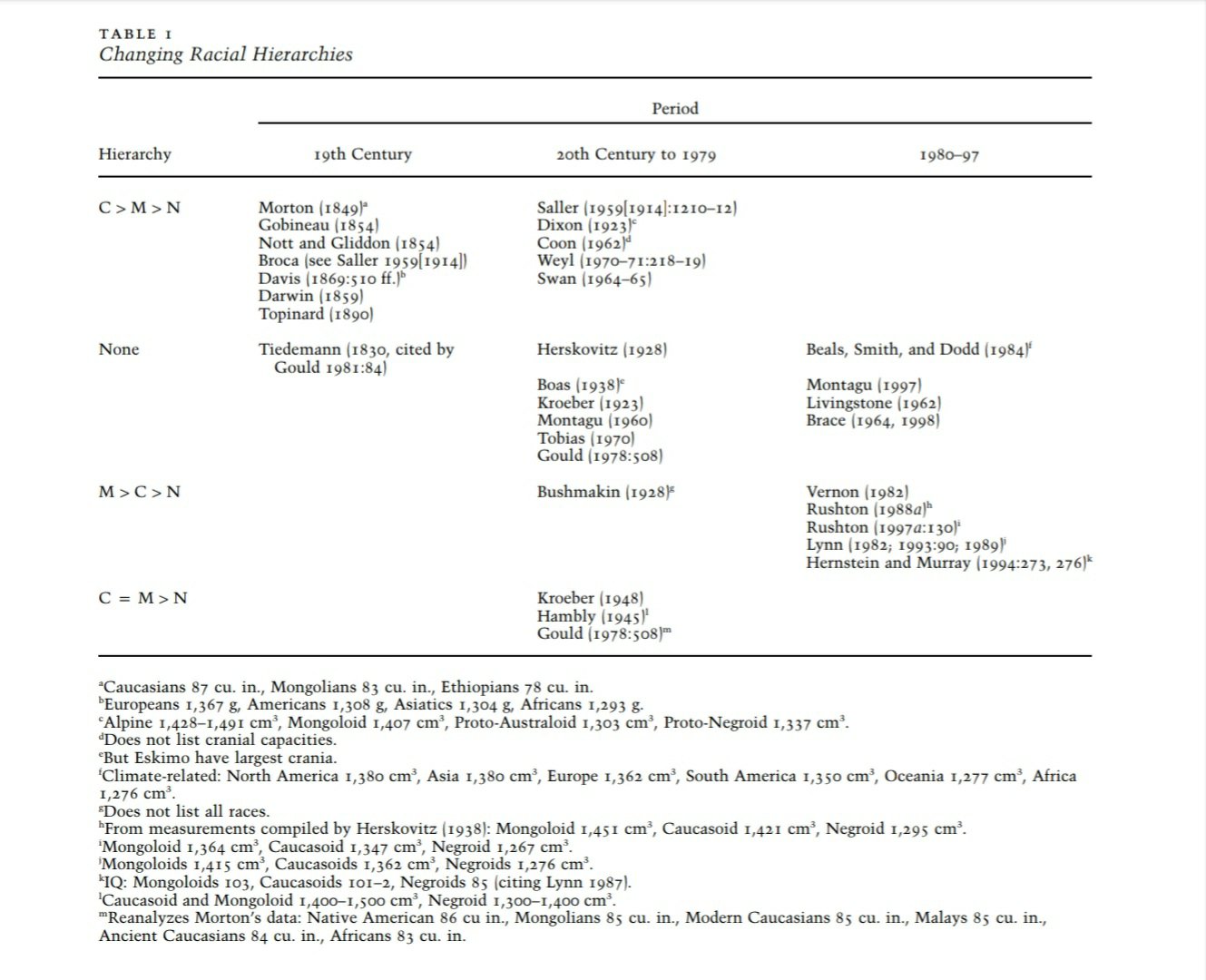
This tells us, again, that our “scientific objectivity” is clouded by political and economic prejudices of the time. This allows Rushton to proclaim “If my work was motivated by racism, why would I want Asians to have bigger brains than whites?” Indeed, what a good question. The answer is that the whole point of “HBD race realism” is to denigrate blacks, so as long as whites are above blacks in their little self-made “hierarchy” no such problem exists for them (Hilliard, 2012).
Note how Rushton’s long debunked- r/K selection theory (Anderson, 1991; Graves, 2002) used the current hierarchy and placed dozens of traits on a hierarchy where it was M > C > N (Mongoloids, Caucasoids, and Negroids respectively, to use Rushton’s outdated terminology). It is a political statement to put the ‘Mongoloids’ at the top of the racial hierarchy; the goal of ‘HBD’ is to denigrate blacks. But, do note that in the late 19th to early 20th century that East Asians were deemed to have small brains, large penises, and that Japanese men, for instance, would “debauch their [white] female classmates” (quoted in Hilliard, 2012: 91).
Conclusion
The “IQ” of China (along with scores on other standardized tests such as TIMMS and PISA), in light of the scandals occurring regarding standardized testing should be suspect. Richard Lynn has failed to report dozens of studies that show low IQ scores for China, thusly inflating their scores. This is, yet again, another nail in the coffin for the ‘Cold Winter Theory’, since the story is formulated on the basis of cherry-picked IQ scores of children. I have noted that if we have different assumptions that we would have different evolutionary stories. Thus, if the other data were provided and, say, Chinese IQ were found to be lower, we would just create a story to justify the score. This is illustrated wonderfully by Flynn (2019):
I will only say that I am suspicious of these because none of us can go back and really evaluate environment and mating patterns. Given free reign, I can supply an evolutionary scenario for almost any pattern of current IQ scores. If blacks had a mean IQ above other races I could posit something like this: they benefitted from exposure to the most rigorous environmental conditions possible, namely, competition from other people. Thanks to greater population pressures on resources, blacks would have benefitted more from this than any of those who left at least for a long time. Those who left eventually became Europeans and East Asians.
The hereditarians point to the academic success of East Asians in America as proof that IQ tests ‘measure’ intelligence, but East Asians in America are a hyper-selected sample. As the references I have provided show, second-generation Chinese immigrants show lower educational attainments than other ethnies (the opposite is true in America) and this is explained by the context that the immigrant family finds themselves in—where do you allocate your energy? Education or entrepreneurship? Such choices seem to be class-based due to the fact education is championed by the Chinese in America and not in Spain and the Netherlands—then dictate, and they also refute any claims of ‘genetic superiority’—they also refute, for that matter, the claim that genes matter for educational attainment (and therefore IQ)—although we did not need to know this to know that IQ is a bunk ‘measure’.
So if Chinese cheat on standardized tests, then we should not accept their IQ scores; the fact that they, for example, provide non-random children from large provinces speaks to their dishonesty. They are like Lynn, in a way, avoiding the evidence that IQ scores are not what they seem—both Lynn and the Chinese government are dishonest cherry-pickers. The ‘fact’ that East Asian educational attainment can be attributed to genes is false; it is attributed to hyper-selectivity and notions of class and what constitutes ‘success’ in the country they emigrate to—so what they attempt is based on (environmental) context.
JP Rushton: Serious Scholar
1300 words
… Rushton is a serious scholar who has amassed serious data. (Herrnstein and Murray, 1996: 564)
How serious of a scholar is Rushton and what kind of “serious data” did he amass? Of course, since The Bell Curve is a book on IQ, H&M mean that his IQ data is “serious data” (I am not aware of Murray’s views on Rushton’s penis “data”). Many people over the course of Rushton’s career have pointed out that Rushton was anything but a “serious scholar who has amassed serious data.” Take, for example, Constance Hilliard’s (2012: 69) comments on Rushton’s escapades at a Toronto shopping mall where he trolled the mall looking for blacks, whites, and Asians (he payed them 5 dollars a piece) to ask them questions about their penis size, sexual frequency, and how far they can ejaculate:
An estimated one million customers pass through the doors of Toronto’s premier shopping mall, Eaton Centre, in any given week. Professor Jean-Phillipe Rushton sought out subjects in its bustling corridors for what was surely one of the oddest scientific studies that city had known yet—one that asked about males’ penis sizes. In Rushton’s mind, at least, the inverse correlation among races between intelligence and penis size was irrefutable. In fact, it was Rushton who made the now famous assertion in a 1994 interview with Rolling Stone magazine: “It’s a trade-off: more brains or more penis. You can’t have everything. … Using a grant from the conservative Pioneer Fund, the Canadian professor paid 150 customers at the Eaton Centre mall—one-third of whom he identified as black, another third white, and the final third Asian—to complete an elaborate survey. It included such questions such as how far the subject could ejaculate and “how large [is] your penis?” Rushton’s university, upon learning of this admittedly unorthodox research project, reprimanded him for not having the project preapproved. The professor defended his study by insisting that approval for off-campus experiments had never been required before. “A zoologist,” he quipped, “doesn’t need permission to study squirrels in his back yard.” [As if one does not need to get approval from the IRB before undertaking studies on humans… nah, this is just an example of censorship from the Left who want to hide the truth of ‘innate’ racial differences!]
(I wonder if Rushton’s implicit assumption here was that, since the brain takes most of a large amount of our consumed energy to power, that since blacks had smaller brains and larger penises that the kcal consumed was going to “power” their larger penis? The world may never know.)
Imagine you’re walking through a mall with your wife and two children. As your shopping, you see a strange man with a combover, holding measuring tape, approaching different people (which you observe are the three different social-racial groups) asking them questions for a survey. He then comes up to you and your family, pulling you aside to ask you questions about the frequency of the sex you have, how far you can ejaculate and how long your penis is.
Rushton: “Excuse me sir. My name is Jean-Phillipe Rushton and I am a psychologist at the University of Western Ontario. I am conducting a research study, surveying individuals in this shopping mall, on racial differences in penis size, sexual frequency, and how far they can ejaculate.”
You: “Errrrr… OK?”, you say, looking over uncomfortably at your family, standing twenty feet away.
Rushton: “First off, sir, I would like to ask you which race you identify as.”
You: “Well, professor, I identify as black, quote obviously“, as you look over at your wife who has a stern look on her face.
Rushton: “Well, sir, my first question for you is: How far can you ejaculate?”
You: “Ummm I don’t know, I’ve never thought to check. What kind of an odd question is that?“, you say, as you try to keep your voice down as to not alert your wife and children to what is being discussed.
Rushton: “OK, sir. How long would you say your penis is?”
You: “Professor, I have never measured it but I would say it is about 7 inches“, you say, with an uncomfortable look on your face. You think “Why is this strange man asking me such uncomfortable questions?
Rushton: “OK, OK. So how much sex would you say you have with your wife? And what size hat do you wear?“, asked Rushton, it seeming like he’s sizing up your whole body, with a twinkle in his eye.
You: “I don’t see how that’s any of your business, professor. What I do with my wife in the confines of my own home doesn’t matter to you. What does my hat size have to do with anything?”, you say, not knowing Rushton’s ulterior motives for his “study.” “I’m sorry, but I’m going to have to cut this interview short. My wife is getting pissed.”
Rushton: “Sir, wait!! Just a few more questions!”, Rushton says while chasing you with the measuring tape dragging across the ground, while you get away from him as quickly as possible, alerting security to this strange man bothering—harasasing—mall shoppers.
If I was out shopping and some strange man started asking me such questions, I’d tell him tough luck bro, find someone else. (I don’t talk to strange people trying to sell me something or trying to get information out of me.) In any case, what a great methodology, Rushton, because men lie about their penis size when asked.
Hilliard (2012: 71-72) then explains how Rushton used the “work” of the French Army Surgeon (alias Dr. Jacobus X):
Writing under the pseudonym Dr. Jacobus X, the author asserted that it was a personal diary that brought together thirty years of medical practice as a French government army surgeon and physician. Rushton was apparently unaware that the book while unknown to American psychologists, was familiar to anthropologists working in Africa and Asia and that they had nicknamed the genre from which it sprang “anthroporn.” Such books were not actually based on scientific research at all; rather, they were a uniquely Victorian style of pornography, thinly disguised as serious medical field research. Untrodden Fields [the title of Dr. Jacobus X’s book that Rushton drew from] presented Jacobus X’s observations and photographs of the presumably lurid sexual practices of exotic peoples, including photographs of the males’ mammoth-size sexual organs.
[…]
In the next fifteen years, Rushton would pen dozens of articles in academic journals propounding his theories of an inverse correlation among the races between brain and genital size. Much of the data he used to “prove” the enormity of the black male organ, which he then correlated inversely to IQ, came from Untrodden Fields. [Also see the discussion of “French Army Surgeon” in Weizmann et al, 1990: 8. See also my articles on penis size on this blog.]
Rushton also cited “research” from the Penthouse forum (see Rushton, 1997: 169). Citing an anonymous “field surgeon”, the Penthouse Forum, and asking random people in a mall questions about their sexual history, penis size and how far they can ejaculate. Rushton’s penis data, and even one of the final papers he penned “Do pigmentation and the melanocortin system modulate aggression and sexuality in humans as they do in other animals?” (Rushton and Templer, 2012) is so full of flaws I can’t believe it got past review. I guess a physiologist was not on the review board when Rushton’s and Templer’s paper went up for review…
Rushton pushed the just-so story of cold winters (which was his main thing and his racial differences hypothesis hinged on it), along with his long-refuted human r/K selection theory (see Anderson, 1991; Graves, 2002). Also watch the debate between Rushton and Graves. Rushton got quite a lot wrong (see Flynn, 2019; Cernovsky and Litman, 2019), as a lot of people do, but he was in no way a “serious scholar”.
Why yes, Mr. Herrnstein and Mr. Murray, Rushton was, indeed, a very serious scholar who has amassed serious data.
Race as a Genuine Kind
1600 words
Race realism can be defined as the belief that our racial categories pick out real kinds in nature. Picking out real kinds in nature is a necessary condition for race realism to be true. I’ve extensively covered Hardimon’s (2017) and Spencer’s (2014) arguments on the existence of race. They definitively prove that, all though there is a social/cultural dimension to race, biological racial realism is true, since our racial categories truly do pick out real kinds in nature.
In the philosophy of race, questions such as “What is race?” and “Is race biological?” are asked. Philosophers of race use many tools at their disposal to attempt to answer these and other questions regarding the metaphysics of race. Four main views on race exist: racial anti-realism, the belief that race does not exist; race realism, the belief that race is real; social constructivism, the belief that race is a social construct; and biological racial realism, the belief that race is biologically real (Spencer, 2011).
Note that social constructivists about race are race realists; they believe that race is real but that it is strictly a social reality. Racial anti-realists take the belief that race does not exist at all; so those who make the claim that social constructivists about race say that race is not real, they really mean to say that racial anti-realists claim that race is not real. People hear the phrase “social construct” and automatically assume that that individual is attempting to argue that race is not real. But social constructivists about race do believe that race is real—that is, they are race realists, but only in a social, not biological, manner.
Spencer (2011: 9) argues that genuine kinds are kinds that contribute to long-term scientific progress, meaning that genuine kinds are “a valid kind in a well-ordered SRP [scientific research program].” Spencer (2011: 9) states that valid kinds in an SRP are “useful for playing an epistemic role of scientific kinds in that SRP, as well as a kind that is adequately epistemically justified in its SRP” while also stating that “ a valid kind in an SRP must be useful for playing an epistemic role of scientific kinds in its SRP.” Kinds are valid in SRPs when they can help scientists generate observations for understanding the natural world. Genuine kinds can be epistemically useful without being inductively useful.
Genuine kinds can be epistemically real or just a human (social) construct. Genuine kinds are real enough to use in scientific research—because they tell us just enough about the world to be epistemically useful in a SRP. That a kind is genuine means that it is “real enough” to be used in scientific research.
Now let’s look at Spencer’s argument that biological racial realism should mean “race is a genuine kind in biology”:
(1) The meaning of ‘biological racial realism’ in the race debate should be a metaphysically minimal interpretation of important scientific kindhood that also does the most justice to what counts as an important scientific kind.
(2) A “metaphysically minimal” interpretation of important scientific kindhood is one that does not adopt unnecessary and contentious metaphysical assumptions.
(3) The interpretation of important scientific kindhood that does the most justice to what counts as an important scientific kind is the one that best captures epistemically important scientific kinds—or ‘EIS kinds’ for short.
(4) The candidates for important scientific kindhood in the race debate are naturalo kinds, naturali kinds, naturalu kinds, naturalp kinds, realp biological kinds, reali biological kinds, and genuine kinds.
(5) No kind of kind in the race debate is both metaphysically minimal and does a better job of capturing EIS kindhood than genuine kinds.
(6) Therefore, the meaning of ‘biological racial realism’ in the race debate should be ‘race is a genuine kind in biology’.
This argument provided by Spencer establishes the fact that biological racial realism in the race debate should be ‘race is a genuine kind in biology.’ Now that I have laid out that an entity being biologically real in the race debate is not whether or not it is objective but whether it is epistemically justified in a biological research program, I will now turn to other author’s views on biological racial realism.
The topic on “Race as a Biological Kind” on PhilPapers states that:
One option is to say that one’s ancestor is a member of race X in virtue of sharing similar phenotypic, or observable, properties specific to other members of one’ s reproductively isolated breeding population. A second option is to say that one’s ancestor is a member of race X in virtue of sharing similar genotypic, or genetic, properties specific to other members of one’s reproductively isolated breeding population.
While “… ancestral relations among reproductively isolated breeding populations and either genotypic or phenotypic properties is one way to develop a naturalistic account of race.” Note how this is almost, to the tee, Hardimon’s (2017) populationist race concept.
Andreason (2000) writes that “Most constructivists assume that biological realism and social constructivism are incompatible views about race; I argue that the two conceptions can be compatible.” Indeed, the claim that race is a social construct of a biological reality is a tempting view to take (and one I take to myself). That we socially construct groups does not mean that there is no biological reality to them, as Spencer (2014) shows with his Blumenbachian partitions. The claim that race is a social construct of a biological reality is completely at ends with the claim that race is purely social/political (as Dorothy Roberts argues) and the claim that race is purely biological. Though, we cannot separate ‘social/political’ and ‘biological’ terminology from our ontology. We must use both in conjunction in order to tease out what ‘race’ truly means. Stating that race is a social construct and only a social construct betrays the biology behind race. Stating that race is only biological betrays the social aspects of race. (Though, Hardimon (2017) has one race concept—the minimalist race concept—in which there is nothing ‘social’ about his proposed concept of race.)
Hardimon has four concepts of race: (1) the racialist concept of race—the claim that significant intellectual and moral differences exist between races (which he dispatched in his book); (2) the minimalist concept of race—the claim that groups that exhibit patterns of visible physical features which correspond to geographic ancestry satisfy the conditions of minimalist race; (3) the populationist race concept of race (the scientization of the minimalist race concept)—the claim that “race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population” (Hardimon, 2017: 99); and (4) the socialrace concept of race—the claim that race is a social, not biological, reality, and that society constructs what ‘race’ is.
Our racial categories pick out real kinds in nature, therefore race realism is true. This is established by both Hardimon (2017) and Spencer (2014), who discuss Rosenberg et al’s (2002) paper on the existence of population clusters deemed continental-level minimalist races by Hardimon (2017) and Blumenbachian partitions by Spencer (2014). Both arguments provided by Hardimon and Spencer are sound—they answer numerous objections that critics bring up to argue against Rosenberg et al’s (2002) study and other’s interpretation of what they say.
This article discusses Rosenberg et al (2002), and has this to say about the Kalash (the sixth cluster in that Rosenberg et al (2002) found):
The sixth population which is composed entirely of members of the Kalash, an isolated population in central Pakistan, is omitted by Wade on the grounds it “makes no genetic or geographic sense”. But the Kalash have a significant degree of genetic isolation from others, comparable in magnitude to that of other groups such as Native Americans. Moreover, they all live in the same place.
This is nonsense. That Structure labels a population as genetically distinct does not entail that that population is a continental-level minimalist race. That Structure picks out the Kalash as a genetically distinct group does not undercut Hardimon’s (2017) arguments on the existence of continental-level minimalist races. So, both K=6 and K=5 show that continental-level minimalist races are genetically structured (Hardimon, 2017: 88). (Further responses to critiques of Rosenberg et al can be found here and in Hardimon, 2017, chapter 5 and Spencer, 2014.)
Lastly, I’ll take some time to respond to commenter Oliver D. Smith’s objections to cranial measures and geographic ancestry.
Smith cited Sierp and Henneberg’s (2015) paper Can ancestry be consistently determined from the skeleton? in which the authors show that “no one individual was identified as belonging to only one ‘racial class’” (Sierp and Henneberg, 2015: 23). I don’t find this to be a problem. If we take Hardimon’s minimalist and populationist race concepts, the three conditions that need to be satisfied to delineate races are differences in phenotype which correspond to geographic ancestry, geographic location, and geographic ancestry. The critique from Sierp and Henneberg (2015) only counts against one parameter—if that—which are skulls. When delineating races, we don’t only look at skulls (one visible physical feature), we look at the whole suite of traits that make a “race” “racial.” Therefore, Sierp and Henneberg’s (2015) critique has no bearing on Hardimon’s (2017) or Spencer’s (2014) arguments.
Is race a genuine kind in biology? Do our racial categories pick out real kinds in nature? The answer to both questions is “yes.” Race is a genuine kind in biology since it captures EIS kindhood; our racial categories do pick out real kinds in nature, as shown by Spencer (2014) and his Blumenbachian partitions. Hardimon’s and Spencer’s arguments are definitive: biological racial realism is true.
(i) If biological racial realism is true, then our racial categories would have to pick out real kinds in nature.
(ii) Our racial categories pick out real kinds in nature.
(iii) Therefore, biological racial realism is true.
Why Biological Racial Realism is True
1650 words
To be a realist about race is to hold that racial categories pick out real kinds in nature. (Smith, 2015: 43; Nature, Human Nature, and Human Difference Race in Early Modern Philosophy)
Claims from biological racial realists are simple: Racial categories pick out real kinds in nature. If racial categories pick out real kinds in nature, then race surely exists.
Racial groups—or groups taken to be racial groups—characterize three conditions from Hardimon (2017): C1: they are distinguished from other groups by patterns of visible physical features; C2: the members are linked by common ancestry which is peculiar to that group; and C3: they derive from a distinct geographic location.
Justifying C1 is simple: groups taken to be ‘racial’ are distinguished from other groups on the basis of physical characters. Someone from Europe looks different than someone from Africa; someone from Africa looks different than someone from Asia; someone from Asia looks different than someone from the Pacific Islands; someone from the Pacific Islands looks different than the Natives of America. Groups taken to be ‘racial’ have different facial features; they have different morphology. Thus, since there are heritable differences between groups taken to be ‘racial’, then this is evidence that race does indeed exist.
It’s important to also discuss what C1 does not demand: it does not demand that racial groups be distinguished by each of their visible physical features; it does not demand that each visible physical features of members of a race be identical; it allows skin color to vary just as much within race as it does between race; finally, it also allows great variation in hair color, skull morphology and skin color. Thus, since Hardimon’s concept is ‘vague’, then one might be able to say that it is “clinal” (that is, these differences vary by geography). But “Physical anthropologist Frank Livingstone’s well-known adage “There are no races, only clines” overlooks the possibility that, logically speaking, races might be clines” (Hardimon, 2017: 38). The claim “There are no races, only clines” is one that is oft-repeated against the reality of biological races.
C2, very simply, shows that differences in visible physical features are not the only things that delineate race: race is also defined in terms of ancestry and is therefore essential to the concept of race (I’d argue that ancestry is essential to any argument that attempts to establish races as biologically real). Races are, clearly, morphologically demarcated ancestry groups. The justification for C2 is thus: it is intuitive. Examples of race articulated in the past also bore this very basic concept: Linneus’ europeaus, asiaticus, afer, and americanus; Blumenbach’s Mongoloid, Caucasoid, Ethiopian, Malay and American; UNESCO’s Negroid, Mongoloid, and Caucasoid (deployed most famously by JP Rushton); and the Office of Management and Budget’s American Indian (or Alaskan native), black, Asian, whites, native Hawaiians (Pacific Islanders) (see Spencer, 2014 for a treatment of the OMB’s views on race and his ‘radical solution to the race problem’).
Now, finally, C3: the condition that groups taken to be ‘racial’ must derive from a distinct geographic location. Race, and the names used to refer to race, and so “The use of typonyms in the naming of racial groups suggests that the thinkers who chose these names were thinking of race as a geographical grouping” (Hardimon, 2017: 50). So, C1 and C2 have been established. This leaves us with C3. Races differ in patterns of visible physical features; these differences are explained by differences in geographic location. If race R1 derives from geographic location G1, and G1 is distinct from G2 which race R2 inhabits, then races R1 and R2 will look physically different.
Thus the groups that we think of when we think about race are groups that genetically transmit heritable characters to their offspring which then correspond to differences in geographic ancestry. So groups that satisfy C1-C3 are ‘races’, in the normal sense of the word. Groups that satisfy C1-C3 are articulated in Hardimon’s (2017) populationist race concept using Rosenberg et al’s (2002) data, and these are, largely, the same groups that Blumenbach pointed out centuries ago (Spencer, 2014).
The Visible Physical Features of Minimalist Race Are Racial
The visible physical features of minimalist race that correspond to geographical ancestry count as “racial” because they are defining features of minimalist races. They no more need to be correlated with normatively important features to be properly counted as racial then minimalist races need to be characterized by normatively important features to be properly counted as races. Just as the concept of minimalist race deflates the concept of RACE, so too it deflates the concept of RACIAL. Visible physical features that correspond to geographical ancestry are eo ipso racial. (Hardimon, 2017: 52)
Hardimon (2017: 99) also articulates one of the best definitions of race I have come across:
A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.
Since Hardimon’s views are new (published in 2017), there are no replies to his argument—excpet one, by Spencer (2018). Spencer doesn’t take to two of Hardimon’s claims: that (1) that the minimalist concept of race is the ordinary concept of race and (2) that minimalist races are biologically real. He grants (1) until Hardimon provides evidence that the minimalist concept of race is the ordinary concept of race. (2), on the other hand, Spencer attacks.
His objection to (2) comes down to the simple fact that 13/17 of the different conceptions of racial groups discussed by Linnaeus, Blumenbach, the OMB, and UNESCO do not fit C1-C3 (Blumenbach’s races do fit C1-C3). Spencer (2018) states that “there’s no ancestor that Eurasians share that’s not also shared by East Asians, Oceanians, and Native Americans […] there’s no ancestor that East Asians share that’s not also shared by Oceanians and Native Americans.” (See Duda and Zrzavy, 2016.) We need to be clear on what Hardimon means by “ancestry.” The dictionary definition of “ancestry” is thus: “one’s family or ethnic descent.” On this definition of “ancestry”, groups taken to be races do have “distinct ancestry“, so defined, and so, Hardimon’s (2017) C2 does indeed hold.
Biological racial realism “should” mean “race is a geniuine kind in biology.” Take the argument from Spencer (2011: 24):
(1) The meaning of ‘biological racial realism’ in the race debate should be a metaphysically minimal interpretation of important scientific kindhood that also does the most justice to what counts as an important scientific kind.
(2) A “metaphysically minimal” interpretation of important scientific kindhood is one that does not adopt unnecessary and contentious metaphysical assumptions.
(3) The interpretation of important scientific kindhood that does the most justice to what counts as an important scientific kind is the one that best captures epistemically important scientific kinds—or ‘EIS kinds’ for short.
(4) The candidates for important scientific kindhood in the race debate are natural kinds, naturali kinds, naturalu kinds, naturalp kinds, realp biological kinds, reali biological kinds, and geniuine kinds.
(5) No kind of kind in the race debate is both metaphysically minimal and does a better job of capturing EIS kindhood than genuine kinds.
(6) Therefore, the meaning of ‘biological racial realism’ in the race debate should be ‘race is a genuine kind in biology’.
Spencer has good critiques of Hardimon’s minimalist/populationist race view, but it does not hold.
Even if we allow Spencer’s views on Hardimon’s arguments for the existence of race to hold, Spencer himself has articulated a sound argument for the existence of race. In his 2014 paper A Radical Solution to the Race Problem, Spencer (2014) shows that Americans defer to the US Census on matters of race; the US Census defers to the OMB; the OMB refers to “sets of” populations—blacks, whites, Asians, Native Americans and Pacific Islanders; these “sets of” populations are not kinds (like what Hardimon argues his racial classifications are); therefore, races are not ‘kinds’, the term ‘race’ refers to sets of population groups. Thus, according to Spencer (2014), race refers to “proper names” for population groups, not “kinds”.
Both of Hardimon’s and Spencer’s arguments show that race is a biological reality; they both show that biological racial realism is true. Their concepts pick out real kinds in nature (Smith, 2016: 43).
Philosopher Justin Smith, in his book 2015 book Nature, Human Nature, and Human Difference Race in Early Modern Philosophy articulates a Hardimonian argument for the existence our habits of distinguishing between human populations:
Now if scientific taxonomy builds on folk taxonomy, and if racial classification builds on this in turn, there might be some basis for supposing that something about the modern habit of distinguishing between human groups on racial grounds is more deep-seated than we have acknowledged it to be. (Smith, 2015: 47)
The reason why there “might be some basis for supposing that something about the modern habit [which is not truly modern; Sarich and Miele, 2004; which I am sure that Smith knows due to the content of his book] of distinguishing between human groups on racial grounds is more deep-seated” than we have acknowledged because we are picking out real kinds that exist in nature.
Conclusion
Race, as a concept, is biologically real. Racial categories pick out real kinds in nature, as argued by Spencer (2014) and Hardimon (2017). Criticisms on Hardimon from Spencer or Spencer from Hardimon do not take away from this one fact: that race exists and is biologically real. Groups taken to be ‘racial’ look different from each other; they look different from each other due to their geographic locations and their ancestry (C1-C3). Since this is true, then race exists. We can argue this view simply:
P1. If groups of people look different from each other depending on where their ancestors evolved, then race exists.
P2. Groups of people look different from each other depending on where their ancestors evolved.
C. Therefore, race exists since people look different depending on where their ancestors evolved (modus ponens, P1, P2).
Biological racial realism is true.
The Physical Reality of Race
1600 words
Rational people can just look at people of different ancestries and see that there is something to what we call “race.” We notice that others look different based on where their ancestors came from and we classify people into different races on the basis of their physical appearance. Anti-biological racial realists may point to the fact that there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016; Hardimon, 2017). While this is true, this does not mean that race is “just a social construct” (a phrase used to deflate the meaning of “race”); it is both a social construct and a biological reality.
The definition of race is simple—a group of populations which genetically transmit heritable characteristics which correspond to that group’s geographic ancestry who also belong to a biological line of descent which was initiated by a geographically isolated and reproductively isolated founding populations (Hardimon, 2017). Note how this definition says nothing about differences in allele frequencies between populations between populations—because, for these purposes, they’re irrelevant for the argument being made. The fact of the matter is, the reality of race hinges on two things: (1) the heritable differences between population groups which were geographically/reproductively isolated and (2) our ability to discern these population groups by their phenotype.
A great book on the history of race, its meaning and how the term was used over the ages is Race: The Reality of Human Differences by Sarich and Miele (2004). For the purposes of this piece, the first two chapters are the most important, since they touch on aspects of race that I have in the past—mainly the fact that we only need phenotype to discern one’s race. People from Europe look phenotypically different from people from Africa who look phenotypically different from people from Asia etc. These differences between these groups are evidence that race exists—these racial differences in phenotype are due, in part, to the climate they evolved in while geographically and reproductively isolated (two conditions for racehood).
Sarich and Miele (2004: 29) write:
Vince [Sarich; one of the authors of the book] naively asked for the legal definition of “race” and was told there wasn’t one.
[…]
As we began working on this book, we discussed the issue of the legal definition of “race” … He informed us that there is still no legal definition of “race”; nor, as far as we know, does it appear that the legal system feels the need for one. Thus, it appears that the most adversarial part of our complex society, the legal system, not only continues to accept the existence of “race” but also relies on the ability of the average individual to sort people into races. Our legal system treats “racial identification” as self-evident …
[…]
The courts have come to accept the commonsense definition of race, and it is this commonsense view that, as we show, best conforms to reality. A look at two recent (2000) cases is illustrative. In both Rice v. Office of Hawaiian Affairs and in Hank v. Rochester School District, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race (even if the racial categories are euphemistically termed “peoples” or “populations”). No special expertise was assumed or granted in defining or recognizing race other than the everyday commonsense usage, as given in the Oxford English Dictionary, that a race is “a group of persons connected by common descent” or “a tribe, nation, or people, regarded as common stock.” The courts and the contending parties, in effect, accepted the existence of race and the ability of the ordinary person to distinguish between races based on a set of physical features.
In Rice v. Office of Hawaiian Affairs, Rice challenged the state of Hawaii since they did not allow him to vote—on the basis that he was not a native Hawaiian, and that the electoral system of Hawaii is for the benefit of Hawaiians and Hawaiians only. Everyone agreed that Rice was a Hawaiian citizen—but he did not have Hawaiian ancestry, so he could not be recognized as “Hawaiian” under state law. However, the SCOTUS overturned the ruling (that Rice should not be allowed to vote on the basis of not having Hawaiian ancestry) 7-2, citing the 15th amendment: “The right of the citizens of the United States to vote shall not be denied or abridged by the United States or any State on account of race, color, or previous condition of servitude.” Sarich and Miele (2004: 31) write “The 15th amendment is explicit—race means what the average person thinks it means—and the majority of the Supreme Court read it that way.” (Also see Hong, 2008 for an overview of the case.)
On the other hand, in Haak v. Rochester School District, the Second Circuit Court of Appeals ruled that a white fourth-grade student named Jessica Haak could transfer from her current district to another district (full of whites) since the transfer program was initiated with the idea of lessening the racial isolation of the adjoining districts. Jessica’s mother cited the 14th amendement, and a district court ruled in their favor but the Second Circuit Court of Appeals overturned the decision. “A “minority pupil” was defined as “a pupil who is of Black or Hispanic origin or is a member of another minority group that historically has been the subject of discrimination” (Sarich and Miele, 2004: 31).
The critical points here are that in both Rice and Haak, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race. No special expertise was assumed or granted in defining or recognizing race other than the everyday usage of the term. In Rice, the court, in effect, took judicial notice of the commonsense definition of race. In Haak, the court accepted physical appearance as a valid means by which the average citizen can recognize races and distinguish among them.
In short, the courts accepted the existence of race, even if the legislature was afraid to use the offending word.
Despite the fact that Sarich and Miele (2004) claim that there is no legal definition of race, Cornell Law School has one definition stating that “the term “racial group” means a set of individuals whose identity as such is distinctive in terms of physical characteristics or biological descent.” While the Law Dictionary, citing the 15th amendment writes that race is “A tribe, people, or nation, belonging or supposed to belong to the same stock or lineage. “Race, color, or previous condition of servitude.” Const U. S., Am. XV.” (Also see Hoffman, 2004 who argues that “race” should not be used in the legal system.)
Notice how Sarich and Miele’s (2004) description of “race” and what “race” is almost—word-for-word—like Spencer’s Blumenbachian partitions (Spencer, 2014). Americans defer to the US Census Bureau on matters of race; the US Census Bureau defers to the Office of Management and Budget (OMB) who speak of sets of populations; these sets of populations correspond to geographic clusters who have distinct phenotypes based on their geographic ancestry, which the average American can discern; therefore race exists. Spencer states that when Americans refer to “race” that Americans refer to both a social construct and a biological reality—that is, Americans socially construct race (think of how Hardimon’s minimalist concept of race is related to the concept of socialrace) but these social constructs do have biological underpinnings which can be discerned in two ways: (1) just observation of phenotypes and (2) looking into the genomes of genetically related individuals who make up these population groups.
Even the ancients distinguished races and sorted them on the basis of hair color/type, skin color, physiognomy etc. “[The Egyptians, Greeks, Romans, Indians, and Chinese] sorted [broad racial groups] based upon the same set of characteristics—skin color, hair form, and head shape” while “it is evident that they relied upon a set of observable features (skin color and form, body build, facial features) quite similar to those used in the commonsense notion of race and the racial classifications of nineteenth-century anthropology to sort the many diverse groups they encountered into a smaller number of categories” (Sarcih and Miele, 2004: 42).
It is very clear that, ever since antiquity at the very least, we have been classifying racial groups on the basis of phenotype—and, come to find out, this is one of the best ways to sort people—and you don’t even need to look at genetic differences between groups. Phenotype is clearly enough to delineate racial groupings, you don’t need genes to delineate race. We only need to recognize that (1) people look different on the basis of where they (or their ancestors) came from; (2) observe that these physical differences between people who come from different places are between real and existing groups; (3) people have common ancestry with others; (4) people derive from distinct geographic locations; so (5) we can infer that race exists.
Race is very clearly a reality—both biologically and socially. At least three sound arguments exist for the existence of race (Sarich and Miele, 2004; Spencer, 2014; Hardimon, 2017; see Hardimon’s and Spencer’s arguments at length). Even those in antiquity delineated races on the basis of physical features—exactly what has been argued by Spencer and Hardimon. Race is physically real—people look different from each other individually, ethnically, and racially.
Biological racial realism is true, and if biological racial realism is true then race exists.
(1) If groups of people look different from each other depending on where their ancestors evolved, then race exists.
(2) Groups of people look different from each other depending on where their ancestors evolved.
(3) Therefore, race exists since people look different depending on where their ancestors evolved.
Do Genes and Polymorphisms Explain the Differences between Eastern and Western Societies?
2150 words
In 2012, biologist Hippokratis Kiaris published a book titled Genes, Polymorphisms, and the Making of Societies: How Genetic Behavioral Traits Influence Human Cultures. His main point is that “the presence of different genes in the corresponding people has actually dictated the acquisition of these distinct cultural and historical lines, and that an alternative outcome might be unlikely” (Kiaris, 2012: 9). This is a book that I have not seen discussed in any HBD blog, and based on the premise of the book (how it purports to explain behavioral/societal outcomes between Eastern and Western society) you would think it would be. The book is short, and he speaks with a lot of determinist language. (It’s worth noting he does not discuss IQ at all.)
In the book, he discusses how genes “affect” and “dictate” behavior which then affects “collective decisions and actions” while also stating that it is “conceivable” that history, and what affects human decision-making and reactions, are also “affected by the genetic identity of the people involved” (Kiaris, 2012: 11). Kiaris argues that genetic differences between Easterners and Westerners are driven by “specific environmental conditions that apparently drove the selection of specific alleles in certain populations, which in turn developed particular cultural attitudes and norms” (Kiaris, 2012: 91).
Kiaris attempts to explain the societal differences between the peoples who adopted Platonic thought and those who adopted Confucian thought. He argues that differences between Eastern and Western societies “are not random and stochastic” but are “dictated—or if this is too strong an argument, they are influenced considerably—by the genes that these people carry.” So, Kiaris says, “what we view as a choice is rather the complex and collective outcome of the influence of people’s specific genes combined with the effects of their specific environment … [which] makes the probability for rendering a certain choice distinct between different populations” (Kiaris, 2012: 50).
The first thing that Kiaris discusses (behavior wise) is DRD4. This allele has been associated with miles migrated from Africa (with a correlation of .85) along with novelty-seeking and hyperactivity (which may cause the association found with DRD4 frequency and miles migrated from Africa (Chen et al, 1999). Kiaris notes, of course, that the DRD4 alleles are unevenly distributed across the globe, with people who have migrated further from Africa having a higher frequency of these alleles. Europeans were more likely to have the “novelty-seeking” DRD7 compared to Asian populations (Chang et al, 1996). But, Kiaris (2012: 68) wisely writes (emphasis mine):
Whether these differences [in DRD alleles] represent the collective and cumulative result of selective pressure or they are due to founder effects related to the genetic composition of the early populations that inhabited the corresponding areas remains elusive and is actually impossible to prove or disprove with certainty.
Kiaris then discusses differences between Eastern and Western societies and how we might understand these differences between societies as regards novelty-seeking and the DRD4-7 distribution across the globe. Westerners are more individualistic and this concept of individuality is actually a cornerstone of Western civilization. The “increased excitability and attraction to extravagance” of Westerners, according to Kiaris, is linked to this novelty-seeking behavior which is also related to individualism “and the tendency to constantly seek for means to obtain satisfaction” (Kiaris, 2012: 68). We know that Westerners do not shy away from exploration; after all, the West discovered the East and not vice versa.
Easterners, on the other hand, are more passive and have “an attitude that reflects a certain degree of stoicism and makes life within larger—and likely collectivistic—groups of people more convenient“. Easterners, compared to Westerners, take things “the way they are” which “probably reflects their belief that there is not much one can or should do to change them. This is probably the reason that these people appear rigid against life and loyal, a fact that is also reflected historically in their relatively high political stability” (Kiaris, 2012: 68-69).
Kiaris describes DRD4 as a “prototype Westerner’s gene” (pg 83), stating that the 7R allele of this gene is found more frequently in Europeans compares to Asians. The gene has been associated with increased novelty-seeking, exploratory activity and human migrations, along with liberal ideology. These, of course, are cornerstones of Western civilization and thought, and so, Kiaris argues that the higher frequency of this allele in Europeans—in part—explains certain societal differences between the East and West. Kiaris (2012: 83) then makes a bold claim:
All these features [novelty-seeking, exploratory activity and migration] indeed tend to characterize Westerners and the culutral norms they developed, posing the intriguing possibility that DRD4 can actually represent a single gene that can “predispose” for what we understand as the stereotypic Western-type behavior. Thus, we could imagine that an individual beating the 7-repeat allele functions more efficiently in Western society while the one without this allele would probably be better suited to a society with Eastern-like structure. Alternatively, we could propose that a society with more individuals bearing the 7-repeat allele is more likely to have followed historical lines and choices more typical of a Western society, while a population with a lower number (or deficient as it is the actual case with Easterners) of individuals with the 7-repeat allele would more likely attend to the collective historical outcome of Eaasterners.
Kiaris (2012: 84) is, importantly, skeptical that having a high number of “novelty-seekers” and “explorers” would lead to higher scientific achievement. This is because “attempts to extrapolate from individual characteristics to those of a group of people and societies possess certain dangers and conceptual limitations.”
Kiaris (2012: 86) says that “collectivistic behavior … is related to the activity of serotonin.” He then goes on to cite a few instances of other polymorphisms which are associated with collective behavior as well. Goldman et al (2010) show ethnic differences in the l and s alleles (from Kiaris, 2012: 86):

It should also be noted that populations (Easterners) that had a higher frequency of the s allele had a lower prevalence of depression than Westerners. So Western societies are more likely to “suffer more frequently from various manifestations of depression and general mood disorders than those of Eastern cultures (Chiao & Blizinsky, 2010)” (Kiaris, 2012: 89).
As can be seen from the table above, Westerners are more likely to have the l allele than Easterners, which should subsequently predict higher levels of happiness in Western compared to Eastern populations. However, “happiness” is, in many ways, subjective; so how would one find an objective way to measure “happiness” cross-culturally? However, Kiaris (2012: 94) writes: “Intuitively speaking, though, I have to admit that I would rather expect Asians to be happier, in general, than Westerners. I cannot support this by specific arguments, but I think the reason for that is related to the individualistic approach of life that the people possess in Western societies: By operating under individualistic norms, it is unavoidably stressful, a condition that operates at the expense of the perception of individuals’ happiness.”
Kiaris discusses catechol-O-methyltransferase (COMT), which is an enzyme responsible for the inactivation of catecholamines. Catecholamines are the hormones dopamine, adrenaline, and noradrenaline. These hormones regulate the “fight or flight” function (Goldstein, 2011). So since catecholamines play a regulatory role in the “fight or flight” mechanism, increased COMT activity results in lower dopamine levels, which is then associated with better performance.
“Warriors” and “worriers” are intrinsically linked to the “fight or flight” mechanism. A “warrior” is someone who performs better under stress, achieves maximal performance despite threat and pain, and is more likely to act efficiently in a threatening environment. A “worrier” is “someone that has an advantage in memory and attention tasks, is more exploratory and efficient in complex environments, but who exhibits worse performance under stressful conditions (Stein et al., 2006)” (Kiaris, 2012: 102).
Kiaris (2012: 107) states that “at the level of society, it can be argued that the specific Met-bearing COMT allele contributes to the buildup of Western individualism. Opposed to this, Easterners’ increased frequency of the Val-bearing “altruistic” allele fits quite well with the construction of a collectivistic society: You have to be an altruist at some degree in order to understand the benefits of collectivism. By being a pure individualist, you only understand “good” as defined and reflected by your sole existence.”
So, Kiaris’ whole point is thus: there are differences in polymorphic genes between Easterners and Westerners (and are unevenly distributed) and that differences in these polymorphisms (DRD4, HTT, MAOA, and COMT) explain behavioral differences between behaviors in Eastern and Western societies. So the genetic polymorphisms associated with “Western behavior” (DRD4) are associated with increased novelty-seeking, tendency for financial risk-taking, distance of OoA migration, and liberal ideology. Numerous different MAOA and 5-HTT polymorphisms are associated with collectivism (e.g., Way and Lieberman, 2006 for MAOA and collectivism). The polymorphism in COMT more likely to be found in Westerners predisposes for “worrier’s behavior”. Furthermore, certain polymorphisms of the CHRNB3 gene are more common in all of the populations that migrated out of Africa, which predisposed for leaders—and not follower—behavior.
| Trait | Gene | Allele frequency |
| Novelty seeking | DRD4 | 7-repeat novelty seeking allele more common in the West |
| Migration | DRD4 | 7-repeat allele is associated with distance from Africa migration |
| Nomads/settlers | DRD4 | 7-repeat allele is associated with nomadic life |
| Political ideology | DRD4 | 7-repeat allele is more common in liberals |
| Financial risk taking | DRD4 | 7-repeat allele is more common in risk takers |
| Individualism/Collectivism | HTT | s allele (collectivistic) of 5-HTT is more common in the East |
| Happiness | HTT | l allele has higher prevalence in individuals happy with their life |
| Individualism/Collectivism | MAOA | 3-repeat allele (collectivistic) more common in the East) |
| Warrior/Worrier | COMT | A-allele (worrier) more common in the West |
| Altruism | COMT | G-allele (warrior) associated with altruism |
| Leader/Follower | CHRBN3 | A-allele (leader) more common in populations Out-of-Africa |
The table above is from Kiaris (2012: 117) who lays out the genes/polymorphisms discussed in his book—what supposedly shows how and why Eastern and Western societies are so different.
Kiaris (2012: 141) then makes a bold claim: “Since we know now that at least a fraction (and likely more than that) of our behavior is due to our genes“, actually “we” don’t “know” this “now”.
The takeaways from the book are: (1) populations differ genetically; (2) since populations differ genetically, then genetic differences correlated with behavior should show frequency differences between populations; (3) since these populations show both behavioral/societal differences and they also differ in genetic polymorphisms which are then associated with that behavior, then those polymorphisms are, in part, a cause of that society and the behavior found in it; (4) therefore, differences in Eastern and Western societies are explained by (some) of these polymorphisms discussed.
Now for a simple rebuttal of the book:
“B iff G” (behavior B is possible if and only if a specific genotype G is instantiated) or “if G, then necessarily B” (genotype G is a sufficient cause for behavior B). Both claims are false; genes are neither a sufficient or necessary cause for any behavior. Genes are, of course, a necessary pre-condition for behavior, but they are not needed for a specific behavior to be instantiated; genes can be said to be difference makers (Sterelny and Kitcher, 1988) (but see Godfrey-Smith and Lewontin, 1993 for a response). These claims cannot be substantiated; therefore, the claims that “if G, then necessarily B” and “B iff G” are false, it cannot be shown that genes are difference makers in regard to behavior, nor can it be shown that particular genes or whatnot.
Conclusion
I’m surprised that I have not come across a book like this sooner; you would expect that there would be a lot more written on this. This book is short, it discusses some good resources, but the conclusions that Kiaris draws, in my opinion, will not come to pass because genes are not neccesary nor sufficient cause of any type of behavior, nor can it be shown that genes are causes of any behavior B. Behavioral differences between Eastern and Western societies, logically, cannot come down to differences in genes, since they are neither necessary nor sufficient causes of behavior (genes are neccessary pre-conditions for behavior, since without genes there is no organism, but genes cannot explain behavior).
Kiaris attempts to show how and why Eastern and Western societies became so different, how and why Western societies are dominated by “Aristotle’s reason and logic”, while Eastern lines of thought “has been dominated by Confucious’s harmony, collectivism, and context dependency” (Kiaris, 2012: 9). While the book is well-written and researched (he talks about nothing new if you’re familiar with the literature), Kiaris fails to prove his ultimate point: that differences in genetic polymorphisms between individuals in different societies explain how and why the societies in question are so different. Though, it is not logically possible for genes to be a necessary nor sufficient cause for any behavior. Kiaris talks like a determinist, since he says that “the presence of different genes in the corresponding people has actually dictated the acquisition of these distinct cultural and historical lines, and that an alternative outcome might be unlikely” (Kiaris, 2012: 9), though that is just wishful thinking: if we were able to start history over again, things would occur differently, “the presence of different genes in the corresponding people” be dammed, since genes do not cause behavior.
