
Three Arguments for the Existence of Race: Establishing Population Groups as Races
2000 words
At least three arguments establish the existence and reality of biological race:
Argument (1) from Michael Hardimon’s (2017) book “Rethinking Race: The Case for Deflationary Realism” (The Argument for the Existence of Minimalist Races, see Chapters 2, 3, and 4):
The conditions of minimalist racehood are as follows:
(C1) … a group, is distinguished from other groups of human beings by patterns of visible physical features
(C2) [the] members are linked be a common ancestry peculiar to members of that group, and
(C3) [they] originate from a distinctive geographic location (Hardimon, 2017: 31).
This is the argument to prove the existence of minimalist races:
P1) There are differences in patterns of visible physical features which correspond to geographic ancestry
P2) These patterns are exhibited between real groups, existing groups (i.e., individuals who share common ancestry)
P3) These real, existing groups that exhibit these physical patterns by geographic ancestry satisfy the conditions of minimalist race
C) Therefore race exists and is a biological reality
Argument (2) from Michael Hardimon’s (2017) book “Rethinking Race: The Case for Deflationary Realism” (The Argument for the Existence of Populationist Races, see Chapters 5 and 6):
P1) The five populations demarcated by Rosenberg et al (2002) are populationist races; K = 5 demarcates populationist races.
P2) Populationist race=minimalist race.
P3) If populationist race=minimalist race, then everything from showing that minimalist races are a biological reality carries over to populationist races.
P4) Populationist races capture differences in genetic variation between continents and this genetic variation is responsible for the distinctive patterns of visible physical features which correspond to geographic ancestry who belong to biological lines of descent which were initiated by geographically isolated founding populations.
C) Therefore, since populationist races=minmalist races, and visible physical features which correspond to geographic ancestry are genetically transmitted by populations who belong to biological lines of descent, initiated by reproductively isolated founding populations, then populationist races exist and are biologically real.
Argument (3) from Quayshawn Spencer’s (2014) paper “A Radical Solution to the Race Problem” (The argument for the Existence of Blumenbachian Populations):
P1) The term “race” in America refers to biologically real entities; when speaking of race in America, Americans defer to the US Census Bureau who defers to the Office of Management and Budget (OMB).
P2) The OMB refers to race as “sets of” categories, while considering “races” to have 5 members, which correspond to the five major geographic regions.
P3) Rosenberg et al show that, at K = 5, meaningful, though small (~4.3 percent) genetic variation exists between continental-populations
C) Since Americans defer to the US Census Bureau who defers to the OMB, and the OMB refers to race as “sets of” categories which then correspond to five clusters found by Rosenberg et al’s (2002) analysis, race (what Spencer, 2014 terms “Blumenbachian populations”) must exist, though “race” is both socially constructed and biologically real.
Put another way, Spencer’s (2014) argument could also be:
P1) The US meaning of “race” is a referent, which refers to the discourse used by the US Census Bureau; the US Census Bureau refers to the discourse used by the Office of Management and Budget (OMB).
P2) The referent of “race”, in US ontology, refers to a set of human population groups, not a biological kind (sets of human population groups as denoted by the OMB), which refer to “Africans”, “Caucasians”, “East Asians”, “Native Americans”, and Pacific Islanders/Oceanians.
P3) The US meaning of race is both biologically real and socially constructed; Americans refer to real, existing groups when they talk about race.
C) If the US meaning of race is a referent which refers to the discourse used by the US Census Bureau and they refer to the OMB who discuss “sets of” population groups, then when Americans talk about race they talk about Blumenbachian partitions, since race is both biologically real and socially constructed.
The claim “Race exists” is now established. Note how Argument (1) establishes the claim that “races” are real, existing groups who are phenotypically distinct populations with differing geographic ancestry. Note how Argument (2) establishes the claim that populationist race = minimalist race and that “races” are a group of populations that exhibit a distinctive pattern of genetically transmitted phenotypic characters which then correspond to that group’s geographic ancestry who belong to a biological line of descent which was initiated by a geographically separated and reproductively isolated founding population. (This definition of “race” a subdivision of Homo sapiens is the best I’ve come across so far.) Finally, note how Argument (3) establishes the claim that race, in the American sense, is both biologically real and socially constructed. All three arguments are sound and logically valid.
Now, which groups fall into which of the five racial categories?
Caucasians
Caucasians denote a wide-range of groups; Europeans, MENA (Middle Eastern/North African) peoples, Indians are a very diverse group, racially speaking, with “Caucasoids”, “Mongoloids” and “Australoids” (Australoids would mean Pacific Islander/Oceanian) (see Kashyap et al, 2006 for an overview of ethnic, linguistic and geographic affiliations of Indians in the study). Ashkenazi Jews are taken to be a specific race in today’s modern racial ontology, however, Ashkenazi Jews do not exhibit a distinctive pattern of genetically transmitted phenotypic characters which then correspond to their geographic ancestry; they do represent a “geographically isolated and reproductively isolated founding population”, but the fact that they do not exhibit a distinctive pattern of genetically transmitted phenotypic characters means they are not a race, according to Arguments (1) and (2). Ashkenazi Jews are Caucasian, and not their own race. Of course, skin color does not denote race, it is only one marker to use to infer which groups are races.
Africans
Africans comprise all of Sub-Saharan Africa. Africa has the most genetic diversity in the human species (see Campbell and Tishkoff, 2010). Africans, in general, have long, slim bodies with a broad nose, dark skin, kinky hair (lip size is different based on the ethny in question). There are over 3,000 different ethnic groups in Africa, who all comprise the same race. Now, since Africans have the most genetic diversity this does not necessarily mean that they are so phenotypically distinct that there are tens, hundreds, thousands of races on the continent. One only needs to refer back to Arguments (1) and (2) to see that brash claims that “all Xs are Ys” don’t make any sense—especially with the arguments laid out above.
East Asians
East Asians denote a minimalist and populationist race (Arguments (1) and (2)) and Blumenbachian partition (Argument (3)). East Asians denote, obviously, those that derive from East Asia (Chinese, Japanese, Koreans, Vietnamese). These peoples are relatively short, on average, have a distinct yellow-ish tint to their skin (which is why they are sometimes called “yellow”), epicanthic folds and shorter limbs (more likely to have the endomorphic phenotype).
Native Americans
Native Americans are derived from a Siberian population that crossed the Bering Land Bridge about 14kya. They then spread throughout the Americas, becoming the “Natives” we know today. They are what used to be termed “red” people, due to their skin color. Native Americans are derived from Siberians, who share affinities with East Asians. (This will be discussed in further depth below.) They have black hair, and dark-ish skin. Populations that lived in the Americans pre-1492 expansion are part of the Native American racial grouping.
Pacific Islanders/Oceanians
The last racial grouping are Pacific Islanders. Spencer (2014: 1032) writes that we can define Oceanians (Pacific Islanders):
as the most inclusive human population born from East Asians in Oceania (Sahul and the Pacific Islands) and from the original human inhabitants of Oceania. Since Sahul was a single landmass composed of present-day Australia, New Guinea, and Tasmania 50,000–60,000 years ago, when humans first inhabited it, and since we know that the original human inhabitants of Oceania interbred to create modern Oceanians, and since temporal parts of populations are genealogically connected, it should be the case that most Oceanians have genealogical connections to the original peoples of some Pacific island. The only Oceanians who will not will be individuals who became Oceanian from interbreeding alone and Oceanians descended from indigenous peoples of Sahul but not indigenous peoples of a Pacific island (e.g., Aboriginal Australians). The final source of evidence comes from counterfactual cases. [Pacific Islanders and Australian Aborigines share a deep ancestry, see McEvoy et al, 2010.]
A group is in race X, if and only if they share a pattern of visible physical features and common geographic ancestry. If they do not share a pattern of visible physical features which correspond to common geographic ancestry then they do not constitute a race. Keep this in mind for the next two sections.
Are Oceanians black?
One claim that gets tossed around a lot (by black nationalists) is the claim that Oceanians are black due to their skin color, certain phenotypic traits. But this could just as easily be explained by convergent evolution, not that they are, necessarily, the same racial grouping. If this were true, then Australian Aborigines would be black, by proxy, since Australian Aborigines and Oceanian are the same race. The claim, though, holds no water. Just because two groups “look similar” (which I do not see), it does not follow that they are the same race, since other conditions need to be met in order to establish the claim that two separate groups belong to the same race.
Are Native Americans Mongoloid?
Lastly is the claim that Native Americans do not denote an actual racial grouping, they are either Mongoloid or a sub-race of Mongoloids.
Many authors throughout history have presumed that Native Americans were Mongoloid. Franz Boas, for example, said that the Maya Indians were Mongoloid, and that, American populations had features the most similar to Mongoloids, so they are thusly Mongoloid. Wikipedia has a great overview of the history of the “Mongoloid” terminology, with examples from authors throughout history. But that is irrelevant. Native Americans genetically transmit heritable phenotypic characters which correspond with their geographic ancestry and are genetically and geographically isolated population groups.
Although the claim that “Native Americans are Mongoloid” has been echoed for hundreds of years, a simple argument can be erected to take care of the claim:
P1) If Native Americans were East Asian/Mongoloid, then they would look East Asian/Mongoloid.
P2) Native Americans don’t look East Asian/Mongoloid, they have a distinct phenotype which corresponds to their geographic ancestry (See Hardimon’s minimalist/populationist race concepts).
C) Therefore, Native Americans are not East Asian/Mongoloid.
Establishing the claim that Native Americans are not East Asian/Mongoloid is simple. Some authors may make the claim that since they look similar (whatever that means, they don’t look similar to me), that they, therefore according to Arguments (1) and (2) they are a separate race and not a sub-race of East Asians/Mongoloids; Argument (3) further establishes the claim that they are a separate race on the basis that they form a distinct cluster in clustering analyses (Rosenberg et al, 2002) and since, Americans defer to the US Census Beureau and the US Census Beureau defers to the OMB who discusses sets of populations, then when Americans talk about race they talk about Native Americans as separate from East Asians/Mongoloids, since, according to Arguments (1) and (2) they have a distinct phenotype.
Generally, they have distinct skin colors (of course, skin color does not equal race, but it is a big tell), they have similar black, straight hair. But they are, in my opinion, just too phenotypically distinct to call them the same race as Mongoloids/East Asians. For the claim “Native Americans and Mongoloids/East Asians” to be true, they would need to satisfy P1 in Argument (1) and P4 in Argument (2). Native Americans do not satisfy P1 in Argument (1) nor do they satisfy P4 in Argument (2). Therefore, Native Americans are not Mongoloid/East Asian.
Conclusion
The claim “Race exists and is a biological reality” is clearly established by three sound, valid arguments—two from Hardimon (2017; chapters 2-6) and one from Spencer (2014). These arguments show, using the latest of genetic clustering studies, that races, as classicly defined, do indeed, exist and that our old views of race hundreds of years ago were, largely, correct. These arguments establish the existence of the old folk-racial categories. Races have distinct phenotypes which are genetically transmitted and are correlated with geographic ancestry. Some may make certain claims that “Oceanians are black” or “Native Americans are Mongoloid”, but these claims do not hold. These two groups in question are phenotypically distinct, and they come from unique geographic locations, therefore they are not a part of the races that some purport them to be.
The Black-White Bench Press Study
2200 words
Blacks vs whites: which race is stronger? Unfortunately, actual studies on actual strength lifts between races are few and far between (and if there are any comparisons between races it’s on an ancillary movement such as quadricep extension; for example see Goodpaster et al 2006). There is, though, one study comparing whites and blacks on a major lift: the bench press. The paper is littered with many problems, most of which make me not able to take the paper seriously. I’ve written about this paper in the past but this article will be much more comprehensive.
When searching for racial comparisons in physical strength, Boyce et al (2014) is one of the only papers one would come across looking at racial differences in strength in one specific (main) lift. The authors used a longitudinal design: they had the police officers’ bench press numbers during their first week on the force and then had data 12.5 years later to assess racial and gender differences in the bench press. They assessed strength scores on the bench press, bench press/lean mass, as well as bench press/body mass.
The assessed 327 police officers: 30 women (13 black, 17 white) and 297 males (41 black and 238 white). Officers that could not be classified as either white or black were removed from the sample. Average number of years between the test at recruitment and later into their career was 12.5 years +/- 2 years while the mean age at the initial recruitment was 26.4 years +/- 3.4 years. The officers were mostly college-educated and were low-middle to upper-middle class.
When the recruits were considered for the job, they had their body mass, percent body fat (with skinfold calipers, which will be covered below) and did a 1RM (one-rep max) on the bench press to assess physical strength. The physical fitness battery (for incoming officers) was as follows (in order): body fat, sit-and-reach flexibility (to see how flexible one is in their lower body/hamstrings), 1RM bench press, and finally a 1.5-mile run. For in-service officers, they had their body fat tested, a treadmill test, bench press, muscular endurance (either curls or sit-ups; curls are a contraindicative exercise if used with a straight bar while sit-ups are contraindicative since they put unneeded strain on the lower back).
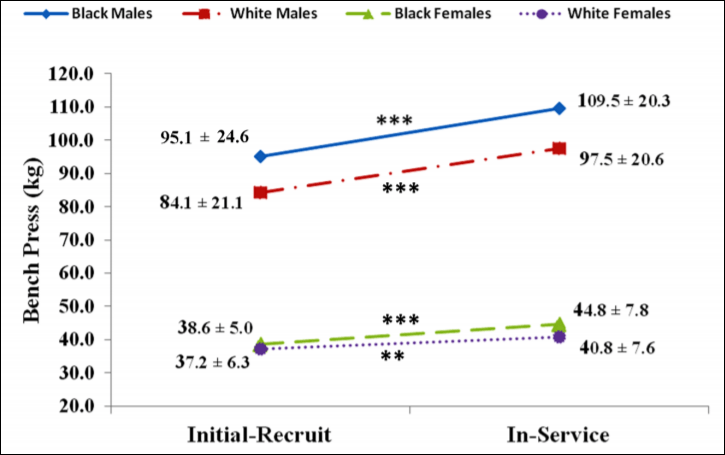
All groups, when combined after 12.5 years, had significant mean gains in strength which were accompanied by gains in body mass, percent fat mass, and lean mass. Black men, at initial recruitment, weighed 187 pounds and benched 210 pounds; white men weighed 180 pounds and benched 185 pounds. Black men benched 1.12 times their body weight whereas white men benched 1.027 times their body weight. At the follow-up assessment, black males weighed 223 pounds and benched 240 pounds. White males weighed 205 pounds and benched 215 pounds. Black males benched 1.07 times their body weight whereas white males benched 1.05 times their body weight. (To find out what percentage of body weight one lifts, take weight lifted and divide it by the weight of the individual.)
Black women, on the other hand at initial recruitment, weighed 130 pounds and benched 85 pounds whereas white women weighed 127 pounds and benched 82 pounds. Black women benched .653 times their body weight whereas white women benched .645 times their body weight. At the follow-up assessment, black women weighed 151 pounds and benched 98 pounds whereas white women weighed 141 pounds and benched 90 pounds. Black women benched .649 times their body weight at the follow-up whereas white women benched .638 times their body weight.
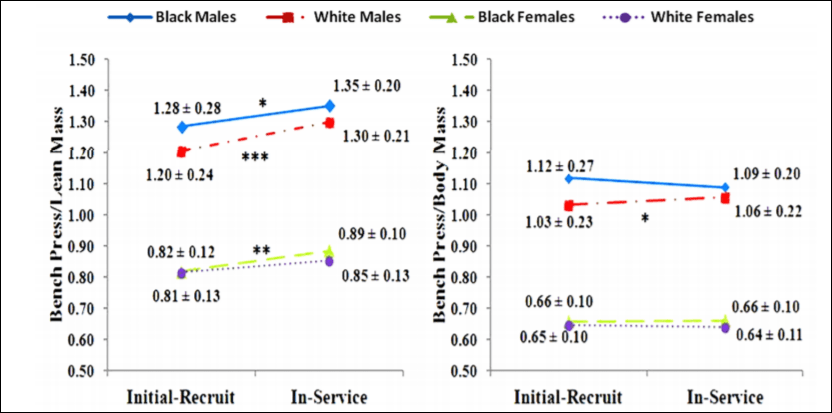
So there was no real difference between black and white female recruits/officers but there was a difference between black and white male recruits/officers. Of course, due to higher levels of testosterone and other hormones compared to women, the males had a changed much more significantly from the initial recruitment. Figures 1 and 2 from Boyce et al (2014: 146) are important too:
Since the study was done over 12.5 years and the individuals were in their mid-20s at initial recruitment, then this study is anomalous since the individuals gained strength and lean mass well into their 30s. Testosterone begins to decrease starting at age 25, though it is not a foregone conclusion that this occurs; age-related declines in testosterone are not inevitable. It just seems that this is the case since, around one’s mid-20s, life changes happen (marriage, kids) which then may cause lifestyle changes (weight gain and the onset of depression). Most of the testosterone decline can be explained by smoking, obesity, depression and other health factors (Shi et al, 2013). So if these officers had good nutrition—and I do not see why they did not—then they can, in effect, bypass most of the so-called age-related declines in testosterone (which is also related to marriage, having children and obesity; Gray, 2011; Pollet, Cobey, and van der Meij, 2013; Mazur et al, 2013).
Gains in lean mass are related to gains in strength, and so, since there was considerable body mass gain and, consequently, lean mass gain, then overall bench pressing strength should have pretty much substantially increased. The strength gains, though, were negated when they divided the weight lifted by the weight of the lifters (pound-for-pound strength, which is what matters more). Black males had the greatest body mass gain over the 12.5 year period, which subsequently corresponded to a decrease in strength on the bench press.
Over the 12.5 year period, black males gained 36 pounds whereas white males gained 25 pounds. Although black males gained more weight over the 12.5 year period—and loss on bench press/body mass—blacks were still very slightly stronger than whites (1.07 compared to 1.05) which is not significant. Black and white females, on the other hand, had no real differences in any of the scores that Boyce et al (2014) did. Furthermore, black and white women, in this study, had similar strength gains and body mass/lean mass gains. Of course, this is a self-selected sample: black women are consistently noted to carry more fat mass than white women (see Rahman et al, 2009).
Both black and white males increased in the bench press throughout the 12.5 year period compared to black and white females, which is due to them being men (higher levels of testosterone and other hormones, larger muscle cross-section area; Miller et al, 1993). The groups who were stronger when they were hired remained the strongest at the follow-up. So, Boyce et al (2014) conclude, the bench press is able to be used as an assessor of upper-body strength since blacks actually, according to their study, are stronger than whites so, therefore, “an upper body strength test such as a bench press will have little adverse impact on blacks, a racially protected group” (Boyce et al, 2014: 148). Though, for women, since they are not as physically strong as men, this will have an “adverse impact on female recruit and incumbent officers, a gender protected group, no matter if they are black or white” (Boyce et al, 2014: 148).
Now time for a few pitfalls (one of which is not the fault of the investigators): height was not assessed; skin-fold calipers were used to assess body fat; and a Smith machine bench press was used.
Height: Due to an ongoing investigation on the Charlotte Mecklenburg Police Department (for apparently discriminating against certain recruits on the basis of height), the height of the officers at recruitment and the follow-up were not noted. This is a problem. If height were known (along with a few more anthropometric variables), then we can infer who had the somatotype that is conducive to bench pressing. In lieu of no height data, we cannot infer this. It is easier for people with shorter limbs to bench press, since the bar has a shorter path to travel, compared to people with longer arms who have to move the weight a further distance. Blacks are more likely to have longer arms and, as I have argued, this would impede them in strength when bench pressing and overhead pressing but will help in the deadlift since they have longer arms and a shorter torso, it is easier for people with this soma to deadlift due to their body proportions.
Skin-fold calipers: Body fat was assessed using skin-fold calipers. Blacks have thinner skin folds than whites, and so since they have thinner skin folds than whites, and the formula for assessing body fat from skin-folds is based on whites, then, it has been argued, that black males need their own BMI scale since they have thinner skin folds and differing levels of fat-free body mass (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000). So since the levels of body fat were off for blacks, then Boyce et al’s (2014) bench press/lean mass is useless since body fat was not assessed correctly. (I know that using calipers is cheaper and easier than sending everyone for a DXA scan, but the difference in body composition between blacks and whites should be known so that, at least when it comes to fat comparisons between different races, bad methods don’t get used and parroted.) Since there are well-known differences in skinfolds as they relate to body fat percentage/lean mass regarding blacks and whites, it’s, again, safe to toss out that part of the study (regarding weight moved/lean mass), since there are huge flaws in regard to assessing body fat through use of conventional measures in blacks compared to whites.
Smith machine bench press: This is, perhaps, one of the most important pitfalls. I can think of a few reasons why this machine was used: (1) you don’t really need to teach someone how to get into proper position to grab the bar. (2) the bar is on a set path, and so people with different anthropometric measures may be uncomfortable while using the machine (which would then affect overall strength). (3) Saeterbakken, Tillaar, and Fimland (2011) assessed three different chest press exercises: the Smith machine bench press, the barbell bench press, and the dumbbell bench press. Those in the study had the highest 1RM on the bench press, followed by the Smith machine and finally were the weakest on dumbbell bench press. The biceps brachii is used for stabilization, and they found that EMG activity in the biceps brachii increased with stability requirements (dumbbell > barbell > Smith). This is due to the fact that, on the two exercises that are not assisted, the muscles need to stabilize the free weight. Since you’re using more muscles to stabilize the weight, then, theoretically, you can move more weight. EMG activity was the same regarding the pectoralis major and anterior deltoids but was different in the triceps and biceps brachii. The prime movers (agonists) of the bench press are the anterior deltoids, triceps, and pecs/serratus. Therefore, the antagonists are the posterior deltoids, the biceps, and the lats/rhomboids.
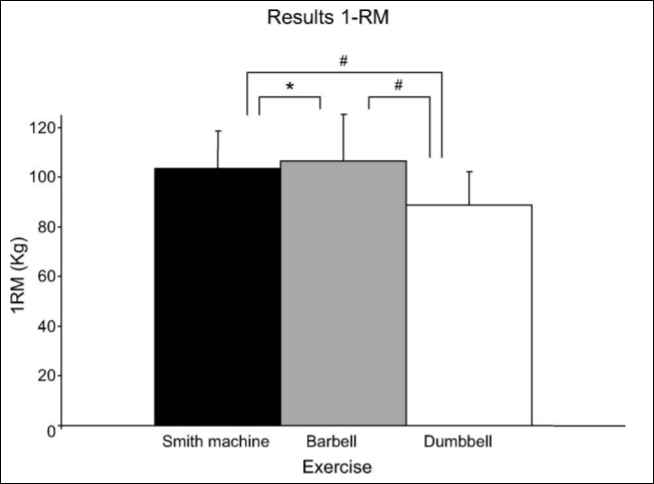
The load lifted on the bench press was three percent higher than on the Smith machine. Since stability requirements are low with the Smith, and the bar is on a set path that cannot be changed, then, theoretically, one should be able to move more weight on the Smith machine (which is my personal experience and the experience of many people I have worked with) compared to the dumbbell and regular bench press. Though, the load lifted on the bench press was about three percent higher than on the Smith machine bench press. The activity of the pectoralis and the anterior deltoid was lower on the eccentric phase (think the negative portion of the lift) when compared to dumbbell and barbell pressing, which is, again, explained by lack of stabilizing muscles used on the Smith machine bench press. So this small study (n=12) shows that there are differences in 1RM between the three lifts studied and that there are differences in the neuromuscular activity of the flexor/extensor muscles of the arm, but showing no difference in the prime movers of the lift (such as the pectoralis major). In the descending phase of the lift, there was less EMG activation, which indicates that a greater neural drive is needed for the eccentric phase of the lift.
The Smith machine used in Boyce et al (2014) was an Atlantis Angled Smith Machine E-155, which I have used personally (and have had hundreds of people use). In my personal experience, weight moved on the Smith is considerably different when compared to a regular bench press, due to the fact that one does not have to stabilize the bar with certain muscles. I will keep an eye out for more Smith machine/bench press/dumbbell press studies in the future, but, from personal experience, one is able to lift more on a Smith than a regular bench.
Conclusion
This is one of the only studies of its kind: assessing racial differences in strength on a major lift. Boyce et al (2014) found that, although blacks had a sharper decrease in pound-for-pound strength, they still kept their slight strength advantage over the 12.5 year period. Both black and white women had similar strength levels and gains in strength and lean mass over the 12.5 year period. Lean mass gain over the period was related to strength gains on the bench; but, as pointed out, they did not correctly assess lean mass for blacks, since calipers (used to test skin folds) are not useful for blacks in the case of figuring out lean mass/body fat levels (Vickery et al, 1988; Wagner and Heyward, 2000).
This study is useless to me. Blacks are not stronger than whites; anthropometric variables play a huge role in strength differences and, due to these differences, blacks are not stronger than whites on certain lifts, as I have documented.
Race, Body Fat, and Skin Folds
1250 words
Racial differences in body fat are clear to the naked eye: black women are more likely to carry more body fat than white women; Mexican American women are more likely to carry more body fat than white women, too. Different races/ethnies/genders of these races/ethnies have different formulas to assess body fat through the use of skin-folds. The sites to grasp the skin is different based on gender and race.
Body mass index (BMI) and waist circumference is overestimated in blacks, which means that they need different formulas to assess their BMI and adiposity/lean mass. Race-specific formulas/methods are needed to assess body fat and, along with it, disease risk, since blacks are more likely to be obese (black women, at least, it’s different with black American men with more African ancestry, see below). The fact of the matter is, when matched on a slew of variables, blacks had lower total and abdominal fat mass than whites.
This is even noted in Asian, black and white prepubertal children. He et al (2002) show that sex differences in body fat distribution are present in children who have yet to reach puberty and the differences in body fat in Asians is different than that from blacks and whites which also varies by sex. Asian girls had greater gynoid fat by DXA scan only, with girls having greater gynoid fat than boys. Asian girls had lower adjusted extremity fat and gynoid fat compared to white and black girls. Though, Asian boys had a lower adjusted extremity by fat as shown by DXA (a gold standard in body fat measurement) when compared to whites, but greater gynoid fat than whites and blacks.
Vickery, Cureton, and Collins, (1988), Wagner and Heyward (2000), and Robson, Bazin, and Soderstrom (1971) show that there are considerable body composition differences between blacks and whites. These differences in body composition come down to diet, of course, but there is also a genetic/physiologic component there as well. Combining the known fact that skin-fold testing is not conducive to a good estimate, black American men with more African ancestry are less likely to be obese.
Vickery, Cureton, and Collins (1988) argue that, if accurate estimates of body fat percentages are to be obtained, race-specific formulas need to be developed and used as independent variables to assess racial differences in body fat percentage. Differences in muscularity don’t seem to account for these skinfold differences, nor does greater mesomorphy. One possible explanation for differences in skinfold thickness is that blacks may store most of their body fat subcutaneously. (See Wagner and Heyward, 2000 for a review on fat patterning and body composition in blacks and whites.)
The often-used Durnin-Womersley formula which is used to predict body fat just from skin folds. However, “The 1974 DW equations did not predict %BF(DXA) uniformly in all races or ethnicities” (Davidson et al, 2011). Truesdale et al (2016) even show that numerous formulas used to estimate percent body fat are flawed, even some formulas used on different races. Most of the equations tested showed starkly different conclusions. But, this is based on NHANES data and the only data they provide regarding skin-folds is the tricep and subscapular skinfold so there may still be more problems with all of the equations used to assess body fat percentage between races. (Also see Cooper, 2010.)
Klimentidis et al (2016) show that black men—but not black women—seem to be protected against obesity and central adiposity (fat gain around the midsection) and that race negatively correlated with adiposity. The combo of male gender and West African ancestry predicted low levels of adiposity compared to black Americans with less African ancestry. Furthermore, since black men and women have—theoretically—the same SES, then cultural/social factors would not play as large a role as genetic factors in explaining the differences in adiposity between black men and black women. Black men with more African ancestry had a lower WHR and less central adiposity than black men with less African ancestry. If we assume that they had similar levels of SES and lived in similar neighborhoods, there is only one reason why this would be the case.
Klimentidis et al (2016) write:
One interpretation is that AAs are exposed to environmental and/or cultural factors that predispose them to greater obesity than EAs. Possibly, some of the genes that are inherited as part of their West-African ancestry are protective against obesity, thereby “canceling out” the obesifying effects of environment/culture, but only in men. Another interpretation is that genetic protection is afforded to all individuals of African descent, but this protection is overwhelmed by cultural and/or other factors in women.
Black men do, as is popularly believed, prefer bigger women over smaller women. For example, Freedman et al (2004) showed that black American men were more likely to prefer bigger women. Black American men “are more willing to idealize a woman
of a heavier body size, with more curves, than do their White American counterparts” (Freedman et al, 2004: 197). It is then hypothesized that black American men find these figures attractive (figures with “more curves” (Freedman et al, 2004: 197)) to protect against eating pathologies, such as anorexia and bulimia. So, it has been established that black men have thinner skin folds than whites which leads to skewed lean mass/body fat readings and black men with more African ancestry are less likely to be obese. These average differences between races, of course, contribute to differing disease acquisition.
I have covered differences in body fat in a few Asian ethnies and have come to the obvious conclusion: Asians, at the same height, weight etc as whites and blacks, will have more adipose tissue on their bodies. They, too, like blacks and whites, have different areas that need to be assessed for skin folds to estimate body fat.
Henriques (2016: 29) has a table on the equations for calculating estimated body density from skin fold measures from various populations. Of interest are the ones on blacks or ‘Hispanics‘, blacks or athletes and blacks and whites. (The table is provided from NSCA, 2008 so the references are not in the back of the text.)
For black and ‘Hispanic’ women aged 18-55 years, the sites to use for skin-folds are the chest, abdomen, triceps, subscapular, suprailiac, midaxillary, and the thigh. For blacks or athletes aged 18-61 years, the sites to use are the same as before (but a different equation is used for body fat estimation). For white women or anorexic women aged 18-55, the sites used are just triceps, suprailiac and the thigh. For black and white boys aged 6-17, only the triceps and the calf is used. It is the same for black and white girls, but, again, a different formula is used to assess body fat (Henriques, 2016: 29).
Morrison et al (2012) showed that white girls had a higher percent body fat when compared to black girls at ages 9-12 but every age after, black girls had higher percent body fat (which is related to earlier menarche in black girls since they have higher levels of body fat which means earlier puberty; Kaplowitz, 2008). Black girls, though, had higher levels of fat in their subscapular skin folds than white girls at all ages.
So, it seems, there are population-/race-specific formulas that need to be created to better assess body fat percentage in different races/ethnies and not assume that one formula/way of assessing body fat should be used for all racial/ethnic groups. According to the literature (some reviewed here and in Wagner and Heyward, 2000), these types of formulas are sorely needed to better assess health markers in certain populations. These differences in body fat percentage and distribution then have real health consequences for the races/ethnies in question.
Just-so Stories: FOXP2
1200 words
FOXP2 is a so-called “gene for” language. The gene is a transcription factor—meaning that it controls the activity of other genes. Thus, changes to FOXP2 will have changes to other genes as well. Thus, the evolution of language in humans was thought to have hinged on mutations on the FOXP2 gene. Humans that have a single-point mutation in FOXP2 “have impaired speech and grammer, but not impaired language comprehension” (Mason, et al, 2018: 403). This gene is found in numerous mammals (e.g., chimpanzees, gorillas, orangutans, rhesus macaques, and mice) but none of those mammals speak. This gene, then, is expressed in the areas of the brain that affects motor functioning, which includes the coordination needed to create words.
Mice and humans at the FOXP2 gene only differ by 3 amino acids. Only one amino acid difference exists between gorillas, chimps, mice, and macaques, who all have identical amino acid sequences on FOXP2. Furthermore, two more amino acid sequences differ between humans and the sequences which is shared by chimpanzees, gorillas, and macaques. Thus, the difference of two amino acids between humans and other primates appears to have made it possible for language to evolve. Evidence exists for strong selective pressures for the two FOXP2 mutations which allow the brain, larynx, and mouth to coordinate to produce speech. These two altered amino acids may change the ability of FOXP2 transcription factor to be phosphorylated—proteins are either activated by phosphorylation or deactivated by dephosphorylation, or the reverse.
Mason et al (2018: 403) write:
Comparative genomics efforts are now extending beyond primates. A role for FOXP2 in songbird singing and vocal learning has been proposed. Mice communicate via squeaks, with lost young mice emitting high-pitched squeaks, FOXP2 mutations leave mice squeakless. For mice and songbirds, it is a stretch to claim that FOXP2 is a language gene—but it is likely needed in the neuromuscular pathway to make sounds.
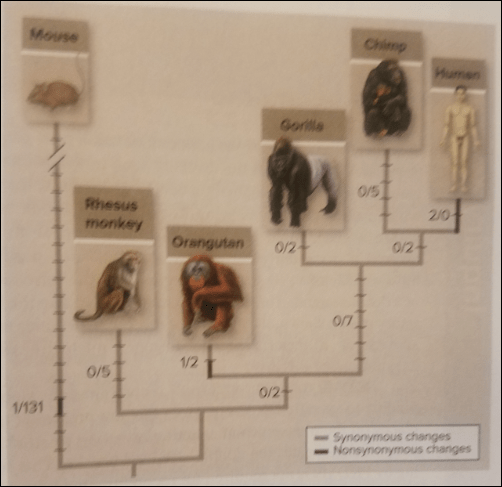
Above is Figure 18.17 from Mason et al (2018: 403). They write:
Comparisons of synonymous and nonsynonymous changes in mouse and primate FOXP2 genes indicate that changing two amino acids in the gene corresponds to the emergence of human language. Black bars represent synonymous changes; gray bars represent nonsynymous changes.
But is that the whole story? Is FOXP2 really a “gene for” language? New results call this hypothesis into question.
In their paper No Evidence for Recent Selection at FOXP2 among Diverse Human Populations, Atkinson et al (2018) did not find evidence for recent positive or balancing selection. Atksinson et al (2018) conclude that they:
do not find evidence that the FOXP2 locus or any previously implicated site within FOXP2 is associated with recent positive selection in humans. Specifically, we demonstrate that there is no evidence that the original two amino-acid substitutions were targeted by a recent sweep limited to modern humans <200 kya as suggested by Enard et al. (2002) … Any modified function of the ROI does not appear to be related to language, however, as modern southern African populations tolerate high minor allele frequencies with no apparent consequences to language faculty. We do not dispute the extensive functional evidence supporting FOXP2’s important role in the neurological processes related to language production (Lai et al., 2001, MacDermot et al., 2005, Torres-Ruiz et al., 2016). However, we show that recent natural selection in the ancestral Homo sapiens population cannot be attributed to the FOXP2 locus and thus Homo sapiens’ development of spoken language.
So the two mutations in exon 7 of FOXP2 weren’t selected and are not responsible for human language. Most likely the accelerated rate is due to loss of function (LoF) (null allele).
The gene was originally discovered in a family that had a history of speech and language disorders (Lai et al, 2001). This “speech gene” was also found in Neanderthals in 2007 (see Krasue et al, 2007). Thus, the modifications to FOXP2 occurred before humans and Neanderthals diverged.
So Atkinson et al (2018) found that the so-called sweep on FOXP2 >200KYA was a statistical artifact which was caused by lumping Africans together Caucasians and other populations. Of course, language is complicated and no one single gene will explain the emergence of human language.
This is a just-so story—that is, an ad hoc hypothesis. Humans had X, others didn’t have X or had a different form of X; therefore X explains human language faculties.
Atkinson et al’s (2018) “results represent a substantial revision to the adaptive history of FOXP2, a gene regarded as vital to human evolution.”
High evolutionary constraint among taxa but variability within Homo sapiens is compatible with a modified functional role for this locus in humans, such as a recent loss of function.
…
Therefore, this SNP must not be necessary for language function as both alleles persist at high frequency in modern human populations. Though perhaps obvious, it is important to note that there is no evidence of differences in language ability across human populations. (Atkinson et al, 2018)
This is another just-so story (Gould and Lewontin, 1976; Lloyd, 1999; Richardson, 2007; Nielsen, 2009) that seems to have bitten the dust. Of course, the functionality of FOXP2 and its role in the neurologic processes related to language; what is disputed (and refuted) is the selectionist just-so story. Selectionist explanations are necessarily ad-hoc. Thus, recent natural selection in our species cannot be attributed to FOXP2, and along with it, our language capabilities.
There is a similar objection, not for FOXP2 and selectionist hypotheses, but for the Lactase gene. Nielsen (2009) puts it succinctly:
The difference in lactose intolerance among human geographic groups, is caused by a difference in allele frequencies in and around the lactase gene (Harvey et al. 1998; Hollox et al. 2001; Enattah et al. 2002; Poulter et al. 2003). … This argument is not erected to dispute the adaptive story regarding the lactase gene, the total evidence in favor of adaptation and selection related to lactose tolerance is overwhelming in this case, but rather to argue that the combination of a functional effect and selection does not demonstrate that selection acted on the specific trait in question. … Although the presence of selection acting on genes underlying a phenotypic trait of interest does help support adaptive stories, it does not establish that selection acted directly on the specific trait of interest.
Even if there were evidence of positive selection of FOXP2 in humans, we cannot logically state that selection acted on the FOXP2 locus; functional effects and selection do not demonstrate that “selection” acted on that trait. Just-so stories (ad hoc hypotheses) “sound good”, but that’s only because they are necessarily true—one can have all the data they want, then they can think up any adaptive story to explain the data and the story will be necessarily true. Therefore, selectionist hypotheses are inherently ad hoc.
In conclusion, another selectionist hypothesis bites the dust. Nevermind the fact that, if FOXP2 were supposedly “selected-for”, there would still be the problem of free-riders (Fodor and Piattelli-Palmarini, 2010). That is, “selection” cannot “select-for” fitness-enhancing traits if/when they are coextensive with other traits—there is no way for selection to distinguish between coextensive traits and thus, it does not explain trait fixation (in this case, the fixation of FOXP2). Ad-hoc hypotheses are necessarily true—that is, they explain the data they purport to explain and only the data they purport to explain. These new results show that there is no support for positive selection at the FOXP2 locus.
