
The Physical Reality of Race
1600 words
Rational people can just look at people of different ancestries and see that there is something to what we call “race.” We notice that others look different based on where their ancestors came from and we classify people into different races on the basis of their physical appearance. Anti-biological racial realists may point to the fact that there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016; Hardimon, 2017). While this is true, this does not mean that race is “just a social construct” (a phrase used to deflate the meaning of “race”); it is both a social construct and a biological reality.
The definition of race is simple—a group of populations which genetically transmit heritable characteristics which correspond to that group’s geographic ancestry who also belong to a biological line of descent which was initiated by a geographically isolated and reproductively isolated founding populations (Hardimon, 2017). Note how this definition says nothing about differences in allele frequencies between populations between populations—because, for these purposes, they’re irrelevant for the argument being made. The fact of the matter is, the reality of race hinges on two things: (1) the heritable differences between population groups which were geographically/reproductively isolated and (2) our ability to discern these population groups by their phenotype.
A great book on the history of race, its meaning and how the term was used over the ages is Race: The Reality of Human Differences by Sarich and Miele (2004). For the purposes of this piece, the first two chapters are the most important, since they touch on aspects of race that I have in the past—mainly the fact that we only need phenotype to discern one’s race. People from Europe look phenotypically different from people from Africa who look phenotypically different from people from Asia etc. These differences between these groups are evidence that race exists—these racial differences in phenotype are due, in part, to the climate they evolved in while geographically and reproductively isolated (two conditions for racehood).
Sarich and Miele (2004: 29) write:
Vince [Sarich; one of the authors of the book] naively asked for the legal definition of “race” and was told there wasn’t one.
[…]
As we began working on this book, we discussed the issue of the legal definition of “race” … He informed us that there is still no legal definition of “race”; nor, as far as we know, does it appear that the legal system feels the need for one. Thus, it appears that the most adversarial part of our complex society, the legal system, not only continues to accept the existence of “race” but also relies on the ability of the average individual to sort people into races. Our legal system treats “racial identification” as self-evident …
[…]
The courts have come to accept the commonsense definition of race, and it is this commonsense view that, as we show, best conforms to reality. A look at two recent (2000) cases is illustrative. In both Rice v. Office of Hawaiian Affairs and in Hank v. Rochester School District, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race (even if the racial categories are euphemistically termed “peoples” or “populations”). No special expertise was assumed or granted in defining or recognizing race other than the everyday commonsense usage, as given in the Oxford English Dictionary, that a race is “a group of persons connected by common descent” or “a tribe, nation, or people, regarded as common stock.” The courts and the contending parties, in effect, accepted the existence of race and the ability of the ordinary person to distinguish between races based on a set of physical features.
In Rice v. Office of Hawaiian Affairs, Rice challenged the state of Hawaii since they did not allow him to vote—on the basis that he was not a native Hawaiian, and that the electoral system of Hawaii is for the benefit of Hawaiians and Hawaiians only. Everyone agreed that Rice was a Hawaiian citizen—but he did not have Hawaiian ancestry, so he could not be recognized as “Hawaiian” under state law. However, the SCOTUS overturned the ruling (that Rice should not be allowed to vote on the basis of not having Hawaiian ancestry) 7-2, citing the 15th amendment: “The right of the citizens of the United States to vote shall not be denied or abridged by the United States or any State on account of race, color, or previous condition of servitude.” Sarich and Miele (2004: 31) write “The 15th amendment is explicit—race means what the average person thinks it means—and the majority of the Supreme Court read it that way.” (Also see Hong, 2008 for an overview of the case.)
On the other hand, in Haak v. Rochester School District, the Second Circuit Court of Appeals ruled that a white fourth-grade student named Jessica Haak could transfer from her current district to another district (full of whites) since the transfer program was initiated with the idea of lessening the racial isolation of the adjoining districts. Jessica’s mother cited the 14th amendement, and a district court ruled in their favor but the Second Circuit Court of Appeals overturned the decision. “A “minority pupil” was defined as “a pupil who is of Black or Hispanic origin or is a member of another minority group that historically has been the subject of discrimination” (Sarich and Miele, 2004: 31).
The critical points here are that in both Rice and Haak, neither side raised any questions about the existence of human races or the ability of the average citizen to make valid judgements as to who belongs to which race. No special expertise was assumed or granted in defining or recognizing race other than the everyday usage of the term. In Rice, the court, in effect, took judicial notice of the commonsense definition of race. In Haak, the court accepted physical appearance as a valid means by which the average citizen can recognize races and distinguish among them.
In short, the courts accepted the existence of race, even if the legislature was afraid to use the offending word.
Despite the fact that Sarich and Miele (2004) claim that there is no legal definition of race, Cornell Law School has one definition stating that “the term “racial group” means a set of individuals whose identity as such is distinctive in terms of physical characteristics or biological descent.” While the Law Dictionary, citing the 15th amendment writes that race is “A tribe, people, or nation, belonging or supposed to belong to the same stock or lineage. “Race, color, or previous condition of servitude.” Const U. S., Am. XV.” (Also see Hoffman, 2004 who argues that “race” should not be used in the legal system.)
Notice how Sarich and Miele’s (2004) description of “race” and what “race” is almost—word-for-word—like Spencer’s Blumenbachian partitions (Spencer, 2014). Americans defer to the US Census Bureau on matters of race; the US Census Bureau defers to the Office of Management and Budget (OMB) who speak of sets of populations; these sets of populations correspond to geographic clusters who have distinct phenotypes based on their geographic ancestry, which the average American can discern; therefore race exists. Spencer states that when Americans refer to “race” that Americans refer to both a social construct and a biological reality—that is, Americans socially construct race (think of how Hardimon’s minimalist concept of race is related to the concept of socialrace) but these social constructs do have biological underpinnings which can be discerned in two ways: (1) just observation of phenotypes and (2) looking into the genomes of genetically related individuals who make up these population groups.
Even the ancients distinguished races and sorted them on the basis of hair color/type, skin color, physiognomy etc. “[The Egyptians, Greeks, Romans, Indians, and Chinese] sorted [broad racial groups] based upon the same set of characteristics—skin color, hair form, and head shape” while “it is evident that they relied upon a set of observable features (skin color and form, body build, facial features) quite similar to those used in the commonsense notion of race and the racial classifications of nineteenth-century anthropology to sort the many diverse groups they encountered into a smaller number of categories” (Sarcih and Miele, 2004: 42).
It is very clear that, ever since antiquity at the very least, we have been classifying racial groups on the basis of phenotype—and, come to find out, this is one of the best ways to sort people—and you don’t even need to look at genetic differences between groups. Phenotype is clearly enough to delineate racial groupings, you don’t need genes to delineate race. We only need to recognize that (1) people look different on the basis of where they (or their ancestors) came from; (2) observe that these physical differences between people who come from different places are between real and existing groups; (3) people have common ancestry with others; (4) people derive from distinct geographic locations; so (5) we can infer that race exists.
Race is very clearly a reality—both biologically and socially. At least three sound arguments exist for the existence of race (Sarich and Miele, 2004; Spencer, 2014; Hardimon, 2017; see Hardimon’s and Spencer’s arguments at length). Even those in antiquity delineated races on the basis of physical features—exactly what has been argued by Spencer and Hardimon. Race is physically real—people look different from each other individually, ethnically, and racially.
Biological racial realism is true, and if biological racial realism is true then race exists.
(1) If groups of people look different from each other depending on where their ancestors evolved, then race exists.
(2) Groups of people look different from each other depending on where their ancestors evolved.
(3) Therefore, race exists since people look different depending on where their ancestors evolved.
My Reading List
In no particular order, below is my reading list. This is not an extensive list of the books I’ve read in my life, only what I’ve read (bought) in the past three years
(1) Race: The Reality of Human Differences (Sarich and Miele, 2004)
(2) Beyond Versus: The Struggle to Understand the Interaction Between Nature and Nurture (Tabery, 2014)
(3) Out of Our Heads: Why You Are Not Your Brain, and Other Lessons from the Biology of Consciousness (Noë, 2014)
(4) Limits of Science? Important things we do not know about everything (Beerbower, 2016)
(5) Evolutionary Biology: Conceptual, Ethical, and Religious Issues (Edited by Thompson and Walsh, 2014)
(6) Genes and Future People: Philosophical Issues in Human Genetics (Glannon, 2002)
(7) The Skull Collectors: Race, Science, and America’s Unburied Dead (Fabian, 2010)
(8) The Ontogeny of Information: Developmental Systems and Evolution (Oyama, 1985)
(9) Evolution’s Eye: A System’s View of the Biology-Culture Divide (Oyama, 2000)
(10) I am Not a Brain: Philosophy of Mind for the 21st Century (Gabriel, 2017)
(11) Agents and Goals in Evolution (Okasha, 2018)
(12) Getting Darwin Wrong: Why Evolutionary Psychology Won’t Work (Wallace, 2010)
(13) Sex and Death: An Introduction to Philosophy of Biology (Sterelny and Griffiths, 1999)
(14) What Darwin Got Wrong (Fodor and Piatteli-Palmarini, 2010)
(15) The Sports Gene: Inside the Science of Extraordinary Athletic Performance (Epstein, 2014)
(16) Dance to the Tune of Life: Biological Relativity (Noble, 2017
(17) Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life (Jablonka and Lamb, 2005)
(18) Making Sense of Genes (Kampourakis, 2017)
(19) Did My Neurons Make Me Do It?: Philosophical and Neurobiological Perspectives on Moral Responsibility and Free Will (Murphy and Brown, 2009)
(20) The Essential Davidson (Introduction by Lepore and Ludwig, 2006)
(21) Evolutionary Psychology as Maladapted Psychology (Richardson, 2007)
(22) Ingenious Genes: How Gene Regulation Networks Evolve to Control Development (Sansom, 2011)
(23) Mind and Cosmos: Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False (Nagle, 2012)
(24) Mental Causation: The Mind-Body Problem (Dardis, 2008)
(25) Paleofantasy: What Evolution Really Tells Us About Sex, Diet, and How We Live (Zuk, 2014)
(26) Logic and Philosophy: An Integrated Introduction (Brenner, 1993)
(27) Why Gould Was Wrong (Oeijord, 2003)
(28) Not in Your Genes: The Real Reason Why Children Are Like Their Parents James, 2016)
(29) Evolving Human Nutrition: Implications for Public Health (Ulijaszek, Mann, and Elton, 2012)
(30) The Philosophy of Human Evolution (Ruse, 2012)
(31) Genetics and Philosophy: An Introduction (Griffiths and Stotz, 2013)
(32) What is Philosophy For? (Midgley, 2018)
(33) A Brief History of Everyone Who Ever Lived: The Human Story Retold Through Our Genes (Rutherford, 2017)
(34) Genes, Polymorphisms, and the Making of Societies: How Genetic Behavioral Traits Influence Human Cultures (Kiaris, 2012)
(35) The Trouble with Twin Studies: A Reassessment of Twin Research in the Social and Behavioral Sciences (Joseph, 2015)
(36) Race Unmasked: Biology and Race in the Twentieth Century (Yudell, 2014)
(37) Inventing Intelligence: How America Came to Worship IQ (Castles, 2012)
(38) Beyond Human Nature: How Culture and Experience Shape the Human Mind (Prinz, 2014)
(39) Darwin Deleted: Imagining a World without Darwin (Bowler, 2013)
(40) Getting Science Wrong: Why the Philosophy of Science Matters (Dicken, 2018)
(41) The Philosophy of Cognitive Science (Cain, 2015)
(42) Social By Nature: The Promise and Peril of Sociogenomics (Bliss, 2018)
(43) Extended Heredity: A New Understanding of Inheritance and Evolution (Bonduriansky and Day, 2018)
(44) Genes: A Philosophical Inquiry (Graham, 2002)
(45) Philosophy of Microbiology (O’Malley, 2014)
(46) Genes, Cells, and Brains: The Promethean Promises of the New Biology (Rose and Rose, 2013)
(47) The Genome Factor: What the Social Genomics Revolution Reveals About Ourselves, Our History, and the Future (Coney and Fletcher, 2017)
(48) Endless Forms Most Beautiful: The New Science of Evo Devo (Carroll, 2006)
(49) The Diabetes Code: Prevent and Reverse Type 2 Diabetes Naturally (Fung, 2018)
(50) The Developing Genome: An Introduction to Behavioral Epigenetics (Moore, 2015)
(51) The Epigenetics Revolution: How Biology is Rewriting Our Understanding of Genetics, Disease, and Inheritance (Carey, 2012)
(52) Darwin’s Athletes: How Sport Has Damaged Black America and Preserved the Myth of Race (Hoberman, 1996)
(53) Psychology and Race (Edited by Watson, 1973)
(54) The Human Superorganism: How the Microbiome is Revolutionizing the Pursuit of a Healthy Life (Dietert, 2016)
(55) Reliability in Cognitive Neuroscience: A Meta-Meta-Analysis (Uttal, 2012)
(56) Studying Human Behavior: How Scientists Investigate Aggression and Sexuality (Longino, 2013)
(57) Philosophy of Mind: An Introduction (Jaworski, 2011)
(58) Straightening the Bell Curve: How Stereotypes about Black Masculinity Drive Research on Race and Intelligence (Hilliard, 2012)
(59) Schizophrenia and Genetics: The End of an Illusion (Joseph, 2017)
(60) Disabled Upon Arrival: Eugenics, Immigration, and the Construction of Race and Disability (Dolmage, 2018)
(61) Living Color: The Biological and Social Meaning of Skin Color (Jablonski, 2012)
(62) The Evolved Apprentice: How Evolution Made Humans Unique (Sterelny, 2012)
(63) Who We Are and How We Got Here: Ancient DNA and the New Science of the Human Past (Reich, 2018)
(64) The Gene: From Genetics to Postgenomics (Rheinberger and Müller-Wille, 2018)
(65) Misbehaving Science: Controversy and the Development of Behavior Genetics (Panofsky, 2014)
(66) Postgenomics: Perspectives on Biology After the Genome (Edited by Richardson and Stevens, 2015)
(67) Behave: The Biology of Humans at Our Best and Worst (Sapolsky, 2017)
(68) The Century of the Gene (Keller, 2000)
(69) My Cells Made Me Do It: The Story of Cellular Determinism (Hayes, 2015)
(70) Intelligence and How to Get It: Why Schools and Cultures Count (Nisbett, 2010)
(71) The Obesity Epidemic: Why Diets and Exercise Don’t Work—And What Does (Toomath, 2017)
(72) Genes, Brains, and Human Potential: The Science and Ideology of Intelligence (Richardson, 2017)
(73) Rethinking Race: The Case for Deflationary Realism (Hardimon, 2017)
(74) DNA Is Not Destiny: The Remarkable, Completely Misunderstood Relationship Between You and Your Genes (Heine, 2017)
(75) Everyone Is African: How Science Explodes the Myth of Race (Fairbanks, 2015)
(76) The Story of the Human Body: Evolution, Health, and Disease (Lieberman, 2013)
(77) The End of Overeating: Taking Back the Insatiable American Appetite (Kessler, 2009)
(78) Full House: The Spread of Excellence from Plato to Darwin (Gould, 1996)
(79) Catching Fire: How Cooking Made Us Human (Wrangham, 2009)
(80) J. Phillipe Rushton: A Life History Perspective (Dutton, 2018)
(81) Why Diets Make Us Fat: The Unintended Consequences of Our Obsession with Weight Loss (Aamodt, 2016)
(82) The Blank Slate: The Modern Denial of Human Nature (Pinker, 2003)
(83) Understanding Biology (Mason et al, 2018)
(84) Up From Dragons: The Evolution of Human Intelligence (Skoyles and Sagan, 2002)
(85) Anatomy and Physiology: The Unity of Form an Function (Saladin, 2010)
(86) The Anatomy of Violence: The Biological Roots of Crime (Raine, 2014)
(87) The Selfish Gene (Dawkins, 1976)
(88) Why We Get Fat and What to Do About It (Taubes, 2011)
(89) The Obesity Code: Unlocking the Secrets of Weight Loss (Fung, 2016)
(90) A New History of Life: The Radical New Discoveries About the Origins and Evolution of Life on Earth (Ward and Kirschvink, 2016)
(91) A Troublesome Inheritance: Genes, Race, and Human History (Wade, 2014)
(92) Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It (Entine, 2000)
(93) Monad to Man: The Concept of Progress in Evolutionary Biology (Ruse, 1996)
(94) Descent of Man, and Selection in Relation to Sex (Darwin, 1871)
(95) On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life (Darwin, 1859)
(96) Guns, Germs, and Steel: The Fates of Human Societies (Diamond, 1997)
(97) This is Your Brain on Parasites: How Tiny Creatures Manipulate Our Behavior and Change Our Society (McAuliffe, 2016)
(98) Secrets from the Eating Lab: The Science of Weight Loss, the Myth of Willpower, and Why You Should Never Diet Again (Mann, 2015)
(99) Darwin’s Unfinished Symphony: How Culture Made the Human Mind (Laland, 2017)
(100) Wonderful Life: The Burgess Shale and Natural History (Gould, 1989)
(101) The Evolution of Beauty: How Darwin’s Forgotten Theory of Mate Choice Shapes the Animal World—And Us (Prum, 2017)
(102) Arrival of the Fittest: How Nature Innovates (Wagner, 2015)
(103) This Idea Must Die: Scientific Theories That Are Blocking Progress (Edited by Brockman, 2015)
(104) The Righteous Mind: Why Good People are Divided by Politics and Religion (Haidt, 2012)
(105) Humankind: How Biology and Geography Shape Diversity (Harcourt, 2016)
(106) Rethinking Thin: The New Science of Weight Loss–and the Realities of Dieting (Kolata, 2008)
(107) Why Evolution Is True (Coyne, 2010)
(108) The Greatest Show on Earth: The Evidence for Evolution (Dawkins, 2009)
(109) The Complete Guide to Fasting: Heal Your Body Through Intermittent, Alternate-Day, and Extended Fasting (Fung and Moore, 2016)
(110) Altruism, Socialization, and Society (Rushton, 1980)
(111) The Paradox of Evolution: The Strange Relationship Between Natural Selection and Reproduction (Rothman, 2015)
(112) The Meaning of Human Existence (Wilson, 2015)
(113) The Red Queen: Sex and the Evolution of Human Nature (Ridley, 1993)
(114) The Evolution Delusion: A Scientific Study of Creation and Evolution (Kirkwood, 2015)
(115) The Ten Thousand Year Explosion: How Civilization Accelerated Human Evolution (Cochran and Harpending, 2009)
(116) Moral Tribes: Emotion, Reason, and the Gap Between Us and Them (Greene, 2013)
(117) The God Delusion (Dawkins, 2006)
(118) The Human Advantage: A New Understanding of How Our Brain Became Remarkable (Herculano-Houzel, 2016)
(119) The Testosterone Hypothesis: How Hormones Regulate the Lifecycles of Civilizations (Barzilai, 2015)
(120) Population Wars: A New Perspective on Competition and Coexistence (Graffin, 2016)
(121) An Odyssey in Time: The Dinosaurs of North America (Russell, 1992)
(122) Race, Evolution, and Behavior: A Life History Perspective (Rushton, 1995)
(123) The g Factor: The Science of Mental Ability (Jensen, 1998)
(124) IQ and Human Intelligence (Mackintosh, 1998)
(125) Good Calories, Bad Calories: Fats, Carbs, and the Controversial Science of Diet and Health (Taubes, 2008)
(126) Intelligence in the Flesh: Why Your Mind Needs Your Body Much More Than It Thinks (Claxton, 2015)
(127) The Bell Curve: Intelligence and Class Structure in American Life (Murray and Herrnstein, 1994)
(128) The Genius in All of Us: New Insights into Genetics, Talent, and IQ (Shenk, 2010)
On Individual Variation in Response to Exercise
1500 words
On Twitter, JayMan wrote: “Not talked about much by fitness buffs (a world that’s full of BS anyway): a fair fraction of people respond little to even *negatively* to exercise“. This is the same person that thinks behavior genetics is a science, and that is a field “that’s full of BS anyway”, too. Anyway, the article that JayMan cited was from the website Stronger by Science, titled Hardgainers? What We Know About Non-Responders by Greg Nuckols.
First off, JayMan’s comment that “a fair fraction of people respond … *negatively* to exercise” is, on its face, already false. Most everyone in the study referenced by Nuckols (There Are No Nonresponders to Resistance-Type Training in Older Men and Women; Churchward-Venne et al, 2015) gained strength, but some people’s muscle fibers did not grow, and some apparently shrank (that is, their muscle cross-section area; CSA). But the important thing to note is that ALL gained strength, which implies physiologic adaptation to the stressor placed on the body (something that is overlooked).
Though, even if some people do not respond to certain programs or weight/rep schemes, does not mean that they are “non-responders”. All that needs to be done is to change the program if one “does not respond” to the program created. All exercise programs should be tailored to the individual and their own specific goals. There is no “one-size-fits-all” exercise program, as can be seen from these studies on so-called “hardgainers.”
The best study for this matter, though, is the HERITAGE (HEalth, RIsk factors, exercise, Training, And GEnetics) study, carried out by five universities in Canada and the US, who enlisted 98 two-generation families and then subject each member to five months of the same stationary bike training regimen—three workouts per week with increasing intensity. Each of the 482 individuals in the study was assayed, and so we would also see which genes would play a role in how fit one person would be in comparison to another.
David Epstein, author of The Sports Gene, writes (pg 85):
Despite the fact that every member of the study was on an identical exercise program, all four sites saw a vast and similar spectrum of aerobic capacity improvement, from about 15 percent of participants who showed little or no gain whatsoever after five months of training all the way up to 15 percent of participants who improved dramatically, increasing the amount of oxygen their bodies could use by 50 percent or more.
Amazingly, the amount of improvement that any one person experienced had nothing to do with how good they were to start. In some cases, the poor got relatively poorer (people who started with a low aerobic capacity and improved little); in others, the oxygen rich got richer (people who started with high aerobic capacity and improved rapidly); with all manner of variation in between—exercisers with a high baseline aerobic capacity and little improvement and others with meager starting aerobic capaacity whose bodies transformed drastically.
Though, contrary to JayMan’s claims, “Fortunately, every single HERITAGE subject experienced health benefits from exercise. Even those who did not improve at all in aerobic capacity improved in some other health parameter, like blood pressure, cholesterol, or insulin sensitivity” (Epstein, 2014: 88).
Epstein also writes about another study, undertaken at the University of Alabama-Birmingham’s Core Muscle and Research Laboratory, writing:
Sixty-six people of varying ages were put on a four-month strength training plan—squats, leg press, and leg lifts—all matched for effort level as a percentage of the meximinum they could lift. (A typical set was eleven reps at 75 percent of the maxmimum that could be lifted for a single rep.) At the end of the trainin, the sibjects fell rather neatly into three groups: those whose thigh muscle fibers grew 50 percent in size; those whose fibers grew 25 percent; and those who had no increased in muscle size at all.
[…]
Seventeen weight lifters were “extreme responders” who added muscle furiously; thirty-two were moderate responders, who had decent gains; and seventeen were nonresponders, whose muscle fibers did not grow.* (pg 110)
* “It’s important to keep in mind that the harder the training, the less likely there are to be “nonresponders.” The harder the work, the more likely a subject will get at least some response, even if it is less than her peers” (pg 376).
Those who responded the most to the regimen had the most satellite cells in their quads which were waiting to be activated by training. When one becomes stronger from hypertrophy, the muscle thickness correlates to muscle CSA (Franchi et al, 2018). When one performs a repetition, the muscle fibers break down—this leads to trauma of the cellular proteins in the muscles which must then go under repair. Numerous growth factors influence the growth of skeletal muscle, such as GH (growth hormone), testosterone, protein and carb intake. Skeletal muscle adapts almost immediately after a bout of exercise, but the apparent changes to the muscle (both in the mirror and seeing large gains in strength on any particular movement) will take weeks and months.
There’s one thing about the claims of “exercise nonresponders” that really gets me: everyone responds positively to exercise, even if it’s not the same exact response to another individual doing the same—or different—exercise! I don’t know who made the claim that “people respond the same to any exercise program”, but that’s a claim that hbdchick made, writing “plenty of the “fitness buffs” do [make the claim that everyone would respond the same to the same exercise regimen]. I then asked her, and JayMan, to name three people who made this outrageous claim: but, of course, I got no answer.
Not to mention that Nuckols ended the article writing:
… there were way fewer nonresponders when people were put on personalized training programs instead of one-size-fits-all standardized programs. This study was primarily looking at aerobic fitness, but it also examined strength measures (bench press and leg press 5RM). It found that all the subjects on personalized programs got stronger, while only 64.3% of the subjects on standardized programs got stronger. This gives us more evidence that “nonresponders” in scientific studies aren’t necessarily “true” nonresponders.
Take two people who have similar measures and, say, start at the same weight on one exercise. In 6 months, all else being equal with regard to lifestyle, there will be a difference in strength gained on that particular exercise. However, an increase from t he baseline from when both individuals began, to the 6-month point, shows that they did, indeed, respond to the exercise program at least in some way (see above quotes from Epstein). Thus, the claim that “there are nonresponders to exercise” makes no sense, on the basis that people necessarily respond physiologically to the stressors placed on them, and so, if they do more (and they will) than they did previously from their baseline, then they did adapt to the protocol, implying that they are not “nonresponders” to exercise. It does not matter if Person B does not catch up to Person A on all variables: the fact that there was a difference in each individual from the baseline all the way to 6 months on a specific regimen implies adaptation to the stressors—which implies that there is no such thing “nonresponders”.
JayMan also has views similar to this, which I have responded to last year in the articles Diet and Exercise: Don’t Do It? and Diet and Exercise: Don’t Do It? Part II. Eating well and exercising—although benefits are not the same for each individual (and I do not know who made the claim this was the case)—does ameliorate numerous diseases and can extend lifespan, contrary to the results of certain studies (e.g., the Look AHEAD study; Annuzzi et al 2014).
Claims from people like JayMan who do not know the first thing about dieting and exercise are dangerous—though, all one has to do is have a basic understanding of physiology to understand that the claim “a fair fraction of people respond little to even *negatively* to exercise” is false, since everyone who does something for the first few times will ALWAYS be better in the months after learning the specific movement, implying that there are no nonresponders to exercise.
Of course everyone does not respond the same to exercise regimen A. Other studies found that increasing the frequency, reps, and set scheme lead to changes in the so-called “nonresponders.” Different individuals respond differently to different training programs [be it, strength, conditioning, cardio, plyometrics, balance, and stabilization etc. But it must be stressed that, although not everyone has the same potential for muscle-building/strength-gaining as, say, the IFBB pros or strongmen/powerlifters, everyone can and does benefit from NOT being sedentary, that much is most definitely clear. These studies that show “nonresponders” run people through the same exercise regimen. Anyone with an iota of experience in this industry knows that people do not respond the same to any and every exercise regimen and, so, the program must be tailored to that specific individual. Though, people like JayMan read this stuff and, without understanding what they’re talking about, jump to brash conclusions that are not supported by reality.
Gene-Selectionism vs. Developmental Systems Theory
2300 words
Two dominant theories exist in regard to development, the “gene’s eye view—gene selectionism (GS)—and the developmental view—developmental systems theory (DST). GS proposes that there are two fundamental processes in regard to evolution: replication and interaction. Replicators (the term was coined by Dawkins) are anything that is copied into the next generation whereas interactors (vehicles) are things that only exist to ensure the replicators’ survival. Thus, Dawkins (1976) proposes a distinction between the “vehicle” (organism) and its “riders/replicators” (the genes).
Gene selectionism
Gene selectionists propose a simple hypothesis: evolution through the differential survival of genes, its main premise being that the “gene” is “the ultimate, fundamental unit of natural selection.” Dusek (1999: 156) writes that “Gene selection claims that genes, not organisms, groups of organisms or species, are selected. The gene is considered to be the unit of selection.” The view of gene selectionists is best—and most popularly put—by Richard Dawkins’ seminal book The Selfish Gene (1976), in which he posits that genes “compete” with each other, and that our “selfish actions” are the result of our genes attempting to replicate to the next generation, relegating our bodies to disposable “vehicles” that only house the “replicators” (or “drivers).
Though, just because one is a gene selectionist does not necessarily mean that they are a genetic determinist (both views will be argued below). Gene selectionists are comitted to the view that genes make a distinctive contribution toward building interactors. Dawkins (1982) claims that genetic determinism is not a problem in regard to gene selectionism. Replicators (genes) have a special status to gene selectionists. Gene selectionists argue that adaptive evolution only occurs through cumulative selection, while only the replicators persist through the generations. Gene selectionists do not see organisms as replicators since genes—and not organisms—are what is replicated according to the view.
The gene selectionist view (Dawkins’ 1976 view) can also be said to apply what Okasha (2018) terms “agential thinking”. “Agential thinking” is “treating an evolved organism as if it were an agent pursuing a goal, such as survival or reproduction, and treating its phenotypic traits, including its behaviour, as strategies for achieving that goal, or furthering its biological interests” (Okasha, 2018: 12). Dawkins—and other gene selectionists—treat genes as if they have agency, speaking of “intra-genomic conflict”, as if genes are competing with each other (sure, it’s “just a metaphor”, see below).
Okasha (2018: 71) writes:
To see how this distinction relates to agential thinking, note that every gene is necessarily playing a zero-sum game against other alleles at the same locus: it can only spread in the population if they decline. Therefore every gene, including outlaws, can be thought of as ‘trying’ to outcompete alleles, or having this as its ultimate goal.
Selfish genes also have intermediate goals, which are to maximize fitness, which is done through expression in the organismic phenotype.
Thus, according to Okasha (2018: 73), “… selfish genetic elements have phenotypic effects which can be regarded as adaptations, but only if we apply the notions of agent, benefit, and goal to genes themselves”, though “… only in an evolutionary context [does] it [make] sense to treat genes as agent-like and credit them with goals and interests.” It does not “make sense to treat genes as even “agent-like and credit them with goals and interests since they can only be attributed to humans.
Other genes have as their intermediate goal to enhance the fitness of their host organism’s relatives, by causing altruistic behaviour [genes can’t cause altruistic behavior; it is an action]. However, a small handful of genes have a different intermediate goal, namely to increase their own transmission in their host organism’s gametes, for example, by biasing segregation in their favour, or distorting the sex-ratio, or transposing to new sites in the genome. These are outlaws, or selfish genetic elements.If oulaws are absent or are effectively suppressed, then the genes within a single organism have a common (intermediate) goal, so will cooperate: each gene can onluy benefit by itself by benefiting the whole organism. Agential thinking then can be applied to the organism itself. The organism’s goal—maximizing its fitness—then equates to the intermediate goal of each of the genes within it. (Okasha, 2018: 72)
Attributing agential thinking to anything other than humans is erroneous, since genes are not “selfish.”
The selfish gene is one of the main theories that define the neo-Darwinian paradigm and it is flat out wrong. Genes are not ultimate causes, as the crafters of the neo-Darwinian Modern Synthesis (MS) propose, genes are resources in a dynamic system and can thusly only be seen as causes in a passive, not active, sense (Noble, 2011).
Developmental systems
The alternative to the gene-centric view of evolution is that of developmental systems theory (DST), first proposed by Oyama (1985).
The argument for DST is simple:
(1) Organisms obviously inherit more than DNA from their parents. Since organisms can behave in ways that alter the environment, environments are also passed onto offspring. Thus, it can be said that genes are not the only things inherited, but a whole developmental matrix is.
(2) Genes, according to the orthodox view of the MS, interact with many other factors for development to occur, and so genes are not the only thing that help ‘build’ the organism. Genes can still play some “privileged” role in development, in that they “control”, “direct” or “organize” everything else, but this is up to gene-selectionists to prove. (See Noble, 2012.)
(3) The common claim that genes contain “information” (that is, context-independent information) is untenable, since every reconstruction of genes contain development about information applies directly to all other developmental outcomes. Genes cannot be singled out as privileged causes in development.
(4) Other attempts—such as genes are copied more “directly—are mistaken, since they draw a distinction between development and other factors but fail.
(5) Genes, then, cannot be privileged in development, and are no different than any other developmental factor. Genes, in fact, are just passive templates for protein construction, waiting to be used by the system in a context-dependent fashion (see Moore, 2002; Schneider, 2007). The entire developmental system reconstructs itself “through numerous independent causal pathways” (Sterelny and Griffiths, 1999: 109).
DNA is not the only thing inherited, and the so-called “famed immortality of DNA is actually a property of cells [since] [o]nly cells have the machinery to correct frequent faults that occur in DNA replication.” The thing about replication, though, is that “DNA and the cell must replicate together” (Noble, 2017: 238). A whole slew of developmental tools are inherited and that is what constructs the organism; organisms are, quite obviously, constructed not by genes alone.
Developmental systems, as described by Oyama (1985: 49) do not “have a final form, encoded before its starting point and realized at maturity. It has, if one focuses finely enough, as many forms as time has segments.” Oyama (1985: 61) further writes that “The function of the gene or any other influence can be understood only in relation to the system in which they are involved. The biological relevance or any influence, and therefore the very “information” it conveys, is jointly determined, frequently in a statistically interactive, not additive, manner, by that influence and the system state it influences.”
DNA is, of course, important. For without it, there would be nothing for the cell to read (recall how the genome is an organ of the cell) and so no development would occur. DNA is only “information” about an organism only in the process of cellular functioning.
The simple fact of the matter is this: the development of organs and tissues are not directly “controlled” by genes, but by the exchange signals of the cells. “Details notwithstanding, what is important to note is that whatever kinds of signals it sends out depends on the kind of signals it receives from its immediate environment. Therefore, neighboring cells are interdependent, and its local interactions among cells that drive the developmental processes” (Kampourakis, 2017: 173).
The fact of the matter is that whether or not a trait is realized depends on the developmental processes (and the physiologic system itself) and the environment. Kampourakis, just like Noble (2006, 2012, 2017) pushes a holistic view of development and the system. Kampourakis (2017: 184) writes:
What genetics research consistently shows is that biological phenomena should be approached holistically. at various levels. For example, as genes are expressed and produce proteins, and some of these proteins regulate or affect gene expression, there is absolutely no reason to privilege genes over proteins. This is why it is important to consider developmental processes in order to undertand how characters and disease arise. Genes cannot be considered alone but only in the broader context (cellular, organismal, environmental) in which they exist. And both characters and disease in fact develop; they are not just produced. Therefore, reductionism, the idea that genes provide the ultimate explanation for characters and disease, is also wrong. In order to understand such phenomena, we need to consider influence at various levels of organization, both bottom-up and top-down. This is why current research has adopted a systems biology approach (see Noble, 2006; Voit, 2016 for accessible introductions).
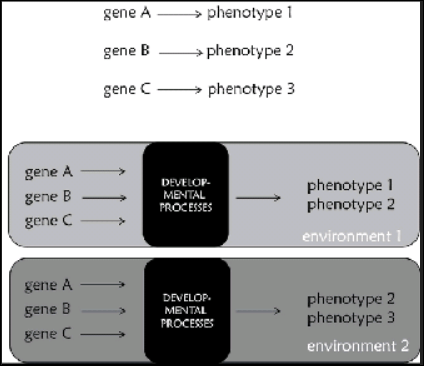
All this shows that developmental processes and interactions play a major role in shaping characters. Organisms can respond to changing environments through changes in their development and eventually their phenotypes. Most interestingly, plastic responses of this kind can become stable and inherited by their offspring. Therefore, genes do not predetermine phenotypes; genes are implicated in the development of phenotypes only through their products, which depends on what else is going on within and outside cells (Jablonka, 2013). It is therefore necessary to replacr the common representation of gene function presented in Figure 9.6a, which we usually find in the public sphere, with others that consider development, such as the one in figure 9.6b. Genes do not determine characters, but they are implicated in their development. Genes are resources that provide cells with a generative plan about the development of the organism, and have a major role in this process through their products. This plan is the resouce for the production of robust developmental outcomes that are at the same time plastic enough to accomodate changes stemming from environmental signals.
Figure 9.6 (a) The common representation of gene function: a single gene determines a single phenotype. It should be clear by what has been present in the book so far that is not accurate. (b) A more accurate representation of gene function that takes development and environment into account. In this case, a phenotype is propduced in a particular environment by developmental processes in which genes are implicated. In a different environment the same genes might contribute tothe development of a different phenotype. Note the “black box” of development.
[Kampourakis also writes on page 188, note 3]
In the original analogy, Wolpert (2011, p. 11) actually uses the term “program.” However, I consider the term “plan” as more accurate and thus more appropriate. In my view, the term “program” impies instructions and their implimentation, whereas the term “plan” is about instructions only. The notion of a genetic program can be very misleading because it implies that, if it were technically feasible, it would be possible to compute an organism by reading the DNA sequence alone (see Lewontin, 2000, pp. 17-18).
Kampourakis is obviously speaking of a “plan” in a context-dependent manner since that is the only way that genes/DNA contain “information” (Moore, 2002; Schneider, 2007). The whole point is that genes, to use Noble’s terminology, are “slaves” to the system, since they are used by and for the (physiological) system. Developmental systems theory is a “wholeheartedly epigenetic approach to development, inheritance and evolution” (Hochman and Griffiths, 2015).
This point is driven home by Richardson (2017:111):
And how did genes eventually become established? Probably not at all as the original recipes, designers, and controllers of life. Instead they arose as templates for molecular components used repeatedly in the life of the cell and the organism: a kind of facility for just-in-time production of parts needed on a recurring basis. Over time, of course, the role of these parts themselves evolved to become key players in the metabolism of the call—but as part of a team, not the boss.
[…]
It is not surprising, then, that we find that variation in form and function has, for most traits, only a tenuous relationship with variation in genes.
[And also writes on page 133]:
There is no direct command line between environments and genes or between genes and phenotypes. Predictions and decisions about form and variation are made through a highly evolved dynamical system. That is why ostensibly the same environment, such as hormonal signal, can initiate a variaety of responses like growth, cell division, differentiation, and migration, depending on deeper context. This reflects more than fixes responses from fixed information in genes, something fatally overlooked in the nature-nurture debate
(Also read Richardson’s article So what is a gene?)
Conclusion
The gene-selectionist point-of-view entails too many (false) assumptions. The DST point of view, on the other hand, does not fall prey to the pitfalls of the gene-selectionist POV; Developmental systems theorists look at the gene, not as the ultimate causes of development—and, along with that, only changes in gene frequency driving evolutionary change—but only as products to be used by and for the system. Genes can only be looked at in terms of development, and in no other way (Kamporuakis, 2017; Noble, 2017). Thus, the gene-selectionists are wrong; the main tenet of the neo-Darwinian Modern Synthesis, gene-selectionism—the selfish gene—has been refuted (Jablonka and Lamb, 2005; Noble, 2006, 2011). The main tenets of the neo-Darwinian Modern Synthesis have been refuted, and so it is now time to replace the Modern Synthesis with a new view of evolution: one that includes the role of genes and development and the role of epigenetics on the developmental system. The gene-selectionist view champions an untenable view of the gene: that the gene is priviliged above any other developmental variables, but Noble and Kampourakis show that this is not the case, since DNA is inherited with the cell; the cell is what is “immortal” to use the language of Dawkins—not DNA itself.
A priori, there is no privileged level of causation, and this includes the gene, which so many place at the top of the hierarchy (Noble, 2012).
Differences in Body Composition Between Asians and Whites
1700 words
Black-white differences get talked about more than Asian-white differences. (For the purposes of this article, “Asian” refers to Koreans, Chinese, Japanese and Filipinos whereas “white” refers to those of European descent.) One interesting racial difference is that of body fatness between ethnies/races. Blacks have thinner skin folds and lower percent body fat than whites at the same height/BMI, and Asians have higher body fat and larger skinfolds than do whites. The interesting thing about this Asian-white difference is the fact that, at the same BMI, Asians have more upper body fat (trunk) than whites. The interesting thing is that there are two good studies, looking at these types of differences between Asians and whites (one study looking at the aforementioned “Asians” I previously identified and whites in the NYC area and another comparing whites and Chinese living in China.)
Wang et al (1994) studied 687 healthy volunteers (445 whites and 242 Asians, ranging from 18-94 years of age with BMIs in the range of 15-38). They defined ethnicity as the birthplace of one’s grandparents. The “Asian” category included 225 Chinese, 9 Japanese, 6 Koreans and 2 Filipinos; 97 percent of this sample was born in Asia. Then, after an overnight fast to better assess body fat differences and skinfold measures, they were weighed and measured, with their back, butt and feet firmly against the wall.
They measured skinfold thickness at the midarm for the average of the triceps and biceps, trunk thickness was the average circumference of the chest, subscapular, umbilicus, abdomen, and suprailiac. The circumference of the arm was measured at the midarm, while the circumference of the trunk was the average circumference of the upper chest, waist, iliac crest, and chest.
Wang et al (1994) found that whites were significantly taller, heavier and had higher BMIs than Asians. Wang et al (1994: 25) write:
For lean and normal BMIs, Asians were fatter than whites in both sexes, but the differences in estimated fat% between whites and Asians varied by BMI in different directions for males and females: fat% increased with BMI for males but decreased with BMI for females.
Whites were had significantly larger circumference in the measured appendages compared to Asians, while in Asian and white females, the circumference of the arms and waist were not different but other circumferences showed a greater difference, favoring whites. Asians had significantly higher levels of subcutaneous trunk fat (upper body fat) than whites, while white females had more lower (thigh) body fat than Asians. In both sexes, Asians had thicker bicep, subscapular, abdomen, and suprailiac skinfolds than whites, in both sexes. White women had higher levels of subcutaneous fat in their thighs. The only difference between white and Asian males in regard to skinfold area was the thigh, with whites having larger thighs, but were similar at the midarm and trunk. Asian men had a larger trunk skinfold area whereas whites had a larger thigh skinfold area while arm fatness did not differ between the races. Women in both races had larger skinfold areas except in the trunk; for whites, there were no differences between the sexes. In both sexes, Asians had higher values in subcutaneous fat (at the midarm, trunk, and midthigh), but white women had a higher value in the thigh than Asian women.
Wang et al (1994) show that there are significant differences in body fatness at different sites of the body, and so, since most (if not all) BMI equations are based on white populations, then, these equations will not work for Asians and will result in substantial error.
Wang et al (2011) studied differences in body composition between Chinese and white males living in the Shenzhen, Guangdong Province, China. They studied 115 Chinese and 114 white males. In this sample, Chinese males were younger, shorter, had a lower body weight and lower BMI than the white sample. Whites had higher fat mass, fat-free mass and bioelectrical impedance (which assess body composition, which measures lean mas in relation to fat mass; but these can be skewed by how much water one has or has not drunk, and so the DXA scan and hydrostatic weighing are, in my opinion, superior assessors). After adjustment for age and BMI, the percentage of fat mass in the trunk and arm was higher in Chinese than white males. Further, Chinese men had higher diastolic blood pressure (DBP), fasting glucose (FG) and triglycerides (TG), while whites had higher fasting total plasma cholesterol (TC) and high-density lipoprotein (HDL). The only statistically significant differences were between FG and HDL. Even after adjustment, Chinese men had 3.0 mmHg higher DBP than whites.
Chinese men had higher percent body fat than whites and more fat stored around their trunks than whites at the same BMI. Chinese men had higher fasting glucose levels (a risk-factor for obesity) but lower HDL levels at the same BMI as whites. Wang et al (2011) write:
In addition, comparing the two nationally representative studies, NHANES III [32] and China National Nutrition and Health Survey 2002 (CNNHS 2002) [33], Chinese men held a relatively 15.0% lower mean value of BMI than that for American white men. While comparison results from two large-scale epidemiological studies, the Shanghai Diabetes Studies (SHDS) [34] and the NHANES III [35], show that the mean value of PBF for American men is relatively 7.4% higher than that for Chinese men. The relative difference of PBF between American and Chinese males is much less than the difference of BMI, implying that the PBF among American men should be lower than that of Chinese men with the same BMI level.
What this implies is that the proportion of overweight/obese Chinese men are severely underestimated since, as noted earlier, most—if not all—BMI equations are created using strictly white populations. This study also provides more evidence that Chinese men had more central (trunk) adiposity than whites (Britons, in this study; Eston, Evans, and Fu, 1994). Central adiposity and risk for type II diabetes and cardiovascular disease is heightened in those of Chinese descent (Weng et al, 2006). It should also be noted that, in a sample of 129 Pacific Islanders, 120 Asians, 91 Maoris, and 91 Europeans aged 12-91, the relationship between bioelectrical impedance analysis (BIA) is ethnicity-dependent, due to the fact the equations developed for fatness estimation using BIA were more accurate than what was recommended by the manufacturer (Sluyter et al, 2010). Cheng (2011) showed that central adiposity was more predictive of cardiovascular diseases in the Chinese population than was BMI, while Hu et al (2007) showed that central obesity was more related to diabetes mellitus and impaired fasting glucose than to overall obesity in the Chinese population.
So, clearly, obesity-related factors appear at lower BMIs for Asians than Europeans (e.g., Huxley et al, 2008). Pan et al (2004) showed that for most BMI values, incidences of hypertension, diabetes, and hyperuricemia were higher in the Taiwanese sample than in the white and black samples. As BMI got higher, the risk for hypertriglyceridemia and hypertension increased. They showed that BMIs of 22.6, 26, and 27.5 were the cutoffs for the best predictabilty in regard to negative and positive variables for Taiwanese, white and black men, respectively. Pan et al (2004: 31) write:
For BMIs 27, 85% of Taiwanese, 66% of whites, and 55% of blacks had at least one of the studied comorbidities. However, a cutoff close to the median of the studied population was often found by maximizing sensitivity and specificity. Reducing BMI from 25 to 25 in persons in the United States could eliminate 13% of the obesity comorbidity studied. The corresponding cutoff in Taiwan is slightly 24.
Pan et al (2004) conclude that, for Taiwanese (Asians) in their study, they should have a lower BMI cutoff than whites and blacks, though it is tough to ascertain where that cutoff would be.
Bell, Adair, and Popkin (2002) show that “at BMI levels less than 25, prevalence difference figures suggested a stronger association between BMI and hypertension in Chinese men and women but not in Filipino women, compared with non-Hispanic Whites” while “[n]on-Hispanic Blacks and Filipino women had a higher prevalence of hypertension at every level of BMI compared with non-Hispanic Whites and Mexican Americans.”
Since Asians have a higher risk of hypertension than whites after controlling for BMI, this indicates that the effects of obesity are not as important as other factors, be they genetic or environmental (or both, which it obviously is). The higher incidence of obesity-related risk-factors in Asian populations with lower BMIs has been attributed to GxE interactions, which, of course, have been intensified with the introduction of the Western Diet (AKA the SAD [Standard American Diet] diet). This can be most notably seen with the explosion of childhood obesity in China, with the number of obese people in China surpassing the US recently, while China is on its way to have the most obese children in the world. The surging obesity epidemic in China is due to increasingly similar lifestyles to what we have (sedentary populations; highly processed, high fat, high carbohydrate foodstuff).
So since the findings in the reviewed studies suggest that, at a lower BMI, Asians are more susceptible to obesity-related risk-factors, and so, BMI standards must be lowered for Asian populations, which would be BMI 24 for overweight and BMI 27 for obese, which was recommended by the Chinese Ministry of Health (Wang et al, 2010). Cheung et al (2018) show that diet quality is inversely associated with obesity in Chinese adults who have type II diabetes.
In conclusion, Asians at the same BMI have higher body fat percentage than whites, and they also have more obesity-related risk-factors than whites at a lower BMI (Pan et al, 2004; WHO expert consultation, 2004; Wang et al, 2010; Hsu et al, 2015), which implies that they need differing BMI scales, just as blacks need different scales in comparison with whites. Here is a good example of two people with the same BMI (22.3) but different DXA results:

This, of course, shows the strong limitations of the use of the same BMI standards calculated in one ethny and used for another. So, just like at the same BMI blacks have lower body fat and thinner skinfolds than whites (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000; Flegal et al, 2010), at the same BMI as whites, Asians have higher body fat and thicker skinfolds (Wang et al, 1994; WHO expert consultation, 2004; Wang et al, 2011).
