
Home » testosterone
Category Archives: testosterone
The AR Gene, Aggression and Prostate Cancer: Yet Another Hereditarian Reduction-to-Biology Fails
2100 words
Due to the outright failure in linking testosterone to differences in racial genetics, prostate cancer (PCa) and aggression, those who would still claim genetic or biological causation for differences in PCa incidence and aggression/crime which would then be linked to race needed to search other avenues for their long-awaited discovery and mechanism for the proposed relationships. This is where the AR (androgen receptor) gene comes in. The AR gene allows the creation of androgen receptors, and this is where androgen “dock”, if you will, allowing the physiological system to use it to carry out what it needs to. In this article, I will discuss the AR gene, CAG repeats, aggression, PCa, a just-so story and finally what may explain the differences in PCa acquisition and aggression between races, not appealing to genes.
Though, hereditarianism is concerned with the so-called biological/genetic transmission of socially-desired traits and also genetic causation of socially-undesired traits, due to the fall of the testosterone-causes-aggressive-behavior paradigm, surely some other biological mechanism could explain why blacks have higher rates of aggression, crime and PCa along with testosterone? Surely, if testosterone isn’t driving the relationship, it would somehow be implicated with it, through some other mechanism in some other kind of way? This is where the androgen receptor gene (AR gene) comes into play. So since CAG repeat length is assumed to be related to androgen receptor sensitivity, and since one report states that lower CAG repeats are associated with aggression (Simmons and Roney, 2011), this is where the hereditarian looks to next for their proposed relationships between testosterone, aggression and PCa. Simmons and Roney also, similarly to Rushton’s r/K, claim that “shorter AR-CAG repeats would have been beneficial for males inhabiting tropical regions because this genetic trait would have encouraged an androgenic response, reportedly along with higher testosterone levels” (Oubre, 2020: 293). Just-so stories all the way down.
Racial differences in AR gene and aggression/PCa
Such claims that the AR polymorphism followed racial lines and was correlated with PCa incidence began to appear in the late 1990s (eg, Giovannuci et al, 1997; Pettaway, 1999). It has been shown that the number of CAG repeats on the AR gene is related to heightened activity on the androgen receptor, and that blacks are more likely to have fewer CAG repeats on the AR gene (Sartor et al, 1997; Platz et al, 2000; Bennett et al, 2002; Gilligan et al, 2004; Ackerman et al, 2013). (But also see Gilligan et al (2004), Lange et al, (2008), and Sun and Lee (2013) for contrary evidence to these claims.) African populations have shorter CAG repeats than non-African populations on the AR gene (Samtal et al, 2022), and since carriers or short CAG repeats had a higher incidence of PCa (Weng et al, 2017; Qin et al, 2022), then this would be the next-best spot to look after the testosterone/PCa/aggression hypothesis failed so spectacularly. But since “Androgen receptor (AR) mediates the peripheral effects of testosterone” (Tirabassi et al, 2015), this has been a new haven for the hereditarian to go to and look for their relationship between aggression and biology.
Fewer CAG repeats has been linked to self-reported aggression (Mettman et al, 2014; Butovskaya et al, 2015; Fernandez-Castillo and Cormand, 2016). Though unfortunately for hereditarian theorists, they also need to look elsewhere, since the number of CAG repeats wasn’t related to aggressive behavior in men nor in women (Valenzuela et al, 2022). Vermeer (2010) found no relationship between CAG repeats and adolescent risk-taking, depression, dominance, or self-esteem. These findings are contrary to other claims, such as this from Geniole et al (2019): “Testosterone thus appears to promote human aggression through an AR-related mechanism“. Rajender et al (2008) showed that rapists and murders had fewer CAG repeats than controls (18.44 repeats, 17.59 repeats, and 21.19 repeats respectively). This is significant due to what was referenced above about testosterone modulating human aggression through an androgen receptor mechanism. Butovskaya et al (2012) also found no relationship between the AR gene and any of the aggression subscales they used.
Since shorter CAG repeats on the AR gene were also related to the severity of PCa incidence (Giovannuci et al, 1997), then what explains the 2 times higher incidence of PCa in blacks compared to whites and 3 to 4 times higher incidence in Asians (Hinata and Fujisawa, 2022; Yamoah et al, 2022) should be related to AR gene and CAG repeats. (Though shorter CGN repeats don’t increase PCa risk in whites and blacks; Li et al, 2017.) However, when blacks and whites had similar preventative care, differences almost entirely vanished (Dess et al, 2019; Yamoah et al, 2022). Lewis and Cropp (2020) have a good review of PCa incidence in blacks. Thus, external—not internal—factors influenced mortality rates, and even though there may be some biological factors that cause either a higher incidence of PCa or survival once it metastatizes, that doesn’t preclude the possibility of inequities on healthcare which cause this relationship (Reddick, 2018). But how can we explain this in an evolutionary context, either recently or in the deep past? Don’t worry, the just-so storytellers have us covered.
Just-so stories and androgen receptors
Urological surgeon William Aiken (2011), publishing in the prestigious journal Medical Hypotheses “speculated” that slaves thsg survived the Middle Passage were more sensitive to androgens, which would then protect them from the conditions they found themselves in on the slave ships during the Passage. This, he surmised, is why African descendants are way disproportionately represented in sprinting records and why, then, blacks have a higher incidence of PCa than whites. Aiken (2011: 1122) explains the “reasoning” behind his hypothesis:
This hypothesis emerged from an exploration of the possible interplay between historical events and biological mechanisms resulting in the similarity in the disproportionate racial and geographic distributions in seemingly unrelated phenomena such as sprinting ability and prostate cancer. The hypothesis is equally a synthesis of the interpretations of observations of a disparate nature such as the high incidence and mortality rates of prostate cancer amongst men of African descent in the Americas while West Africans residing in urban West African centre’s have a lower prostate cancer incidence and mortality [2], the 3-fold greater prostate cancer incidence in Afro-Trinidadians compared to Indo Asian-Trinidadians despite exposure to largely similar environmental conditions [5], the improvement in athletic sprinting performance observed when athletes take anabolic steroids [3], the observation that both sprinting ability and prostate cancer are related to specific hand patterns which in turn are related to antenatal exposure to high testosterone levels [6,7], the observation that prostate cancer is androgen-dependent and undergoes involution when testosterone is inhibited or withdrawn [4], the observation that West Africans born in West Africa are under-represented amongst the elite sprinters [1] despite their relatively large populations and despite West Africa being the region of origin of the ancestors of today’s elite sprinters and finally the observation that prostate cancer is related to androgen receptor responsiveness which in turn is related to its CAG-repeat length [8].
One of Aiken’s predictions is that black Americans and Caribbean blacks should have lower shorter CAG repeats than the populations of descent in Africa. Unfortunately for him, West Africans seem to have shorter CAG repeats than descendants of the Middle Passage (Kittles et al, 2001). Not least, neither of the two predictions he proposed to explain the relationship are risky or novel. By risky prediction I mean a hypothesis that would disprove the overarching hypothesis should the relationship not hold under scrutiny. By novel fact I mean a predicted fact that’s not used in the construction of the hypothesis. Quite clearly, Aiken’s hypothesis doesn’t meet this criteria, and so it is a just-so story.
But such fantastical, selection-type stories have been in the media relatively recently. Like Oprah’s and Dr. Oz’s assertions that blacks that survived the Middle Passage did so in virtue of their ability to retain salt during the voyage which then, today, leads to higher incidences of hypertension. This is known as the slavery hypertension hypothesis (Lujan and DiCarlo, 2018) and is, of course, also a just-so story. Just like the just-so story cited above, those Africans who took the voyage across the sea had some kind of advantage which explained why they survived and, consequently, explained relationships between maladies in their descendants. These types of stories—no matter how well-crafted—are nothing more than stories that explain what they purport to explain with no novel evidence that would raise the probability of the hypothesis being true.
Aggression
Aggression is related to crime, in that it is surmised that more aggressive individuals would then commit more crimes. I’ve noted the failure of hereditarian explanations over the years, so what do I think best explains the relationship between aggression and crime and, ultimately, criminal activity? Well, since crime is an action, it is therefore irreducible. I would propose a kind of situationism in explaining this.
Situational action theory (SAT) (eg Wilkstrom, 2010, 2019) is a cousin of situationism, and is a kind of moral action theory, placing the agent in situations (environments) which then would lead to criminal action as a discourse to take. “The core principle of SAT is that crime is ultimately the outcome of certain ‘kinds of people’ being exposed to certain ‘kinds of situations’” (Messner, 2012). For instance, a good example of this would be for black Americans. Mazur’s (2016) honor culture hypothesis states that blacks who are constantly vigilant for threats to their status and self have higher rates of testosterone in virtue of the fact that aggression increases testosterone.
So this would then be an example of the kind of relationship that SAT would look for. So SAT and the honor culture hypothesis are interactionist in that they recognize the interaction between the agent and the environment (situations) the agent finds themselves in. Violence is merely situational action (Wikstrom and Treiber, 2009), so to explain a violent crime, we need to know the status of the agent and the environment that the crime occurred in, along with the victim and motivating factors for the action in question. The fact of the matter is, actions are irreducible and what is irreducible isn’t physical, so physical (biological) explanations won’t work here. Further, the longer that people stay in criminogenic environments, the more likely they are to commit crime, due to the situations they find themselves in. Thus, a kind of analytic criminology should be employed to discover how and why crimes occur (Wikstrom and Kroneberg, 2022). Considerations in biology should not be looked at when talking about actions and their causes.
Prostate cancer
I have discussed this in the past: What best explains the incidences in PCa between races is diet. For instance, blacks have lower rates of vitamin D than other races (Guiterrez et al, 2022; Thamattoor, 2021). People with lower levels of vitamin D are more likely to acquire PCa, and those with the lowest levels of vitamin D were more likely to have aggressive PCa (Xie et al, 2017). Since consuming high IUs of vitamin D seems to stave off PCa (Khan and Parton, 2004; Naier-Shalliker et al, 2021), and since there seems to be a dose-response relationship between vitamin D consumption and PCa mortality (Song et al, 2018) along wkth vitamin D seeming to reverse low-grade PCa (Samson, 2015), it stands to reason that the higher incidences of PCa in blacks in comparison to whites are due to socio-environmental dietary factors. We don’t need any assumed biological/genetic factors in order to explain the relationship when we know the etiology of PCa.
Conclusion
Due to the old, 1980s and 1990s explanations from hereditarians on the etiology of PCa and aggression with its link to race and testosterone, researchers had to look to other avenues in order to find the “biological etiology” between the relationships. They then pivoted to the AR gene and CAG repeats to explain the relationship between PCa and testosterone when the original testosterone-causes-PCa-and-aggression claim was refuted (Tricker et al, 1996; Book, Starzyk, and Quinsey, 2001; O’Connor et, 2002; Stattin et al, 2004; Archer, Graham-Kevan, and Davies, 2005; Book and Quinsey, 2005; Michaud, Billups, and Partin, 2015; Boyle et al, 2016).
But as can be seen, again, the relationships between the proposed explanations in order to continue pushing their biological/genetic theories of PCa and aggression linked with testosterone and race also fails. Rushton’s r/K theory, for instance, implicated testosterone as a “master switch” (Rushton, 1999). Attempted reductions to biology were also seen in (the now-retracted) Rushton and Templer (2012) (see responses here and here). Reductions to biology quite clearly fail, but that doesn’t deter the hereditarian from pushing the racist theory that genes and biology explain the poor outcomes of blacks.
Do pigmentation and the melanocortin system modulate aggression and sexuality in humans as they do in other animals? A Response to Rushton and Templer (2012)
2100 words
Rushton et al have kept me pretty busy over the last year or so. I’ve debunked many of their claims that rest on biology—such as testosterone causing crime and aggression. The last paper that Rushton published before he died in October of 2012 was an article with Donald Templer—another psychologist—titled Do pigmentation and the melanocortin system modulate aggression and sexuality in humans as they do in other animals? (Rushton and Templer, 2012) and they make a surfeit of bold claims that do not follow. They review animal studies on skin and fur pigmentation and show that the darker an animal’s skin or fur, the more likely they are to be aggressive and violent. They then conclude that, of course (it wouldn’t be a Rushton article without it), that the long-debunked r/K ‘continuum’ explains the co-variation between human populations in birth rate, longevity, violent crime, infant mortality and rate and acquisition of AIDS/HIV.
In one of the very first articles I wrote on this site, I cited Rushton and Templer (2012) favorably (back when I had way less knowledge of biology and hormones). I was caught by biases and not knowing anything about what was discussed. After I learned more about biology and hormones over the years, I came to find out that the claims in the paper are wrong and that they make huge, sweeping conclusions based on a few correlations. Either way, I have seen the error of my ways and the biases that lead me to the beliefs I held, and when I learned more about hormones and biology I saw how ridiculous some of the papers I have cited in the past truly were.
Rushton and Templer (2012) start off the paper by discussing Ducrest et al (2008) who state that within each species studied, darker-pigmented individuals of said species exhibited higher rates of aggression, sexuality and social dominance (which is caused by testosterone) than lighter-pigmented individuals in that same species. They state that this is due to pleiotropy—when a single gene has to or more phenotypic effects. They then refer to Rushton and Jensen (2005) to reference the claim that low IQ is correlated with skin color (skin color doesn’t cause IQ, obviously).
They then state that in 40 vertebrate species that within each that the darker-pigmented members had higher levels of aggression and sexual activity along with a larger body size, better stress resistance, and are more physically active while grooming (Ducrest, Keller, and Roulin, 2008). Rushton and Templer (2012) then state that this relationship was ‘robust’ across numerous species, specifically 36 species of birds, 4 species of fish, 3 species of mammals, and 4 species of reptiles.
Rushton and Templer (2012) then discuss the “Validation of the pigmentation system as causal to the naturalistic observations was demonstrated by experimentally manipulating pharmacological dosages and by studies of cross-fostering“, citing Ducrest, Keller, and Roulin (2008). They even state that ‘Placing darker versus lighter pigmented individuals with adoptive parents of the opposite pigmentation did not modify offspring behavior.” Seems legit. Must mean that their pigmentation caused these differences. They then state something patently ridiculous: “The genes that control that balance occupy a high level in the hierarchical system of the genome.” Though, unfortunately for their hypothesis, there is no privileged level of causation (Noble, 2016; also see Noble, 2008), so this is a nonsense claim. Genes are not ‘blueprints’ or ‘recipes’ (Oyama, 1985; Schneider, 2007).
They then refer to Ducrest, Keller and Roulin (2008: 507) who write:
In this respect, it is important to note that variation in melanin-based coloration between human populations is primarily due to mutations at, for example, MC1R, TYR, MATP and SLC24A5 [29,30] and that human populations are therefore not expected to consistently exhibit the associations between melaninbased coloration and the physiological and behavioural traits reported in our study.
This quote, however, seems to be ignored by Rushton and Templer (2012) throughout the rest of their article, and so even though they did a brief mentioning of the paper and how one should be ‘cautious’ in interpreting the data in their study, it seems like they just brush it under the rug to not have to contend with it. Rushton and Templer (2012) then cite the famous silver fox study, where tame foxes were bred. They lost their dark fur and became lighter and, apparently, were less aggressive than their darker-pigmented kin. These animal studies are, in my useless when attempting to correlate skin color and the melanocortin system in the modulation of aggressive behavior, so let’s see what they write about human studies.
It’s funny, because Rushton and Templer (2012) cite Ducrest, Keller, and Roulin (2008: 507) to show that caution should be made when assessing any so-called differences in the melanocortin system between human races. They then disregard that by writing “A first examination of whether melanin based pigmentation plays a role in human aggression and sexuality (as seen in non-human animals), is to compare people of African descent with those of European descent and observe whether darker skinned individuals average higher levels of aggression and sexuality (with violent crime the main indicator of aggression).” This is a dumb comparison. Yes, African nations commit more crime than European nations, but does this mean that the skin color (or whatever modulates skin color/melanocortin system) is the cause for this? No. Not at all.
There really isn’t anything to discuss here, though, because they just run through how different African nations have higher levels of crime than European and East Asian nations, how blacks report having more sex and feel less guilty about it. Rushton and Templer (2012) then state that one study “asked married couples how often they had sex each week. Pacific Islanders and Native Americans said from 1 to 4 times, US Whites answered 2–4 times, while Africans said 3 to over 10 times.” They then switch over to their ‘replication’ of this finding, using the data from Alfred Kinsey (Rushton and Bogaert, 1988). Though, unfortunately for Rushton and Bogaert, there are massive problems with this data.
Though, the Kinsey data can hardly be seen as representative (Zuckerman and Brody, 1988), and it is also based on outdated, non-representative, non-random samples (Lynn, 1989). Rushton and Templer (2012) also discuss so-called differences in penis size between races, too. But I have written two response articles on the matter and shown that Rushton used shoddy sources like ‘French Army Surgeon who contradicts himself: “Similarly, while the French Army surgeon announces on p. 56 that he once discovered a 12-inch penis, an organ of that size becomes “far from rare” on p. 243. As one might presume from such a work, there is no indication of the statistical procedures used to compute averages, what terms such as “often” mean, how subjects were selected, how measurements were made, what the sample sizes were, etc” (Weizmann et al, 1990: 8).
Rushton and Templer (2012) invoke, of course, Rushton’s (1985; 1995) r/K selection theory as applied to human races. I have written numerous articles on r/K selection and attempts at reviving it, but it is long dead, especially as a way to describe human populations (Anderson, 1991; Graves, 2002). The theory was refuted in the late 70s (Graves, 2002), and replaced with age-specific mortality (Reznick et al, 2002). Some of his larger claims I will cover in the future (like how r/K relates to criminal activity), but he just goes through all of the same old motions he’s been going through for years, bringing nothing new to the table. In all honesty, testosterone is one of the pillars of Rushton’s r/K selection theory (e.g., Lynn, 1990; Rushton, 1997; Rushton, 1999; Hart, 2007; Ellis, 2017; extensive arguments against Ellis, 2017 can be found here). If testosterone doesn’t do what he believes it does and the levels of testosterone between the races are not as high as believed/non-existent (Gapstur et al, 2002; read my discussion of Gapstur et al 2002; Rohrmann et al, 2007; Richard et al, 2014. Though see Mazur, 2016 and read my interpretation of the paper) then we can safely disregard their claims.
Rushton and Templer (2012: 6) write:
Another is that Blacks have the most testosterone (Ellis & Nyborg, 1992), which
helps to explain their higher levels of athletic ability (Entine, 2000).
As I have said many times in the past, Ellis and Nyborg (1992) found a 3 percent difference in testosterone levels between white and black ex-military men. This is irrelavent. He also, then cites John Entine’s (2002) book Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It, but this doesn’t make sense. Because he literally cites Rushton who cites Ellis and Nyborg (1992) and Ross et al (1986) (stating that blacks have 3-19 percent higher levels of testosterone than whites, citing Ross et al’s 1986 uncorrected numbers)—and I have specifically pointed out numerous flaws in their analysis and so, Ross et al (1986) cannot seriously be used as evidence for high testosterone differences between the races. Though I cited Fish (2013), who wrote about Ellis and Nyborg (1992):
“These uncorrected figures are, of course, not consistent with their racial r- and K-continuum.”
Rushton and Templer (2012) then state that testosterone acts like a ‘master switch’ (Rushton, 1999), implicating testosterone as a cause for aggression, though I’ve shown that this is not true, and that aggression causes testosterone production, testosterone doesn’t cause aggression. Testosterone does control muscle mass, of course. But Rushton’s claim that blacks have deeper voices due to higher levels of testosterone, but this claim does not hold in newer studies.
Rushton and Templer (2012) then shift gears to discuss Templer and Arikawa’s (2006) study on the correlation between skin color and ‘IQ’. However, there is something important to note here from Razib:
we know the genetic architecture of pigmentation. that is, we know all the genes (~10, usually less than 6 in pairwise between population comparisons). skin color varies via a small number of large effect trait loci. in contrast, I.Q. varies by a huge number of small effect loci. so logically the correlation is obviously just a correlation. to give you an example, SLC45A2 explains 25-40% of the variance between africans and europeans.
long story short: it’s stupid to keep repeating the correlation between skin color and I.Q. as if it’s a novel genetic story. it’s not. i hope don’t have to keep repeating this for too many years.
Rushton and Templer (2012: 7) conclude:
The melanocortin system is a physiological coordinator of pigmentation and life history traits. Skin color provides an important marker placing hormonal mediators such as testosterone in broader perspective.
I don’t have a problem with the claim that the melanocortin system is a physiological coordinator of pigmentation, because it’s true and we have a great understanding of the physiology behind the melanocortin system (see Cone, 2006 for a review). EvolutionistX also has a great article, reviewing some studies (mouse studies and some others) showing that increasing melatonin appears to decreases melanin.
Rushton and Templer’s (2012) make huge assumptions not warranted by any data. For instance, Rushton states in his VDare article on the subject, J. Phillipe Rushton Says Color May Be More Than Skin Deep, “But what about humans? Despite all the evidence on color, aggression, and sexuality in animals, there has been little or no discussion of the relationship in people. Ducrest & Co. even warned that genetic mutations may make human populations not exhibit coloration effects as consistently as other species. But they provided no evidence.” All Rushton and Templer (2012) do in their article is just restating known relationships with crime and race, and then attempting to implicate the melanocortin system as a factor driving this relationship, literally off of a slew of animal studies. Even then, the claim that Ducrest, Keller, and Roulin (2008: 507) provide no evidence for their warning is incorrect, because before they stated that, they wrote “In this respect, it is important to note that variation in melanin-based coloration between human populations is primarily due to mutations at, for example, MC1R, TYR, MATP and SLC24A5 [29,30]. . .” Melanin does not cause aggression, it does not cause crime. Rushton and Templer just assume too many things based on no evidence in humans, while their whole hypothesis is structured around a bunch of animal studies.
In conclusion, it seems like Rushton and Templer don’t know anything about the physiology of the melanocortin system if they believe that pigmentation and the melanocortin system modulates aggression and sexual behavior in humans. I know of no evidence (studies, not Rushton and Templer’s 2012 relationships with crime and then asserting that, because these relationships are seen in animals, that it must mean that the melanocortin system in humans modulates the relationships too) for these assertions by Rushton and Templer (2012). The fact that they think that restating relationships between crime and race, country of origin and race, supposed correlations with testosterone and crime and blacks supposedly having higher testosterone than whites, among other things, shows that the proposed relationships are caused by the melanocortin system and Lift History Theory shows their ignorance of the human physiological system.
Steroid Mythconceptions and Racial Differences in Steroid Use
2000 words
Steroids get a bad reputation. It largely comes from movies and people’s anecdotal experiences and repeating stories they hear from the media and other forms of entertainment, usually stating that there is a phenomenon called ‘roid rage’ that makes steroid users violent. Is this true? Are any myths about steroids true, such as a shrunken penis? Are there ways to off-set it? Steroids and their derivatives are off-topic for this blog, but it needs to be stressed that there are a few myths that get pushes about steroids and what it does to behavior, its supposed effects on aggression and so forth.
With about 3 million AAS (ab)users (anabolic-androgenic steroids) in America (El Osta et al, 2016), knowing the effects of steroids and similar drugs such as Winny (a cutting agent) would have positive effects, since, of course, athletes mostly use them.
Shrunken testicles
This is, perhaps, one of the most popular. Though the actual myth is that AAS use causes the penis to shrink (which is not true), in reality, AAS use causes the testicles to shrink by causing the Leydig cells to decrease natural testosterone production which then decreases the firmness and shape of the testicles which then results in a loss of size.
In one study of 772 gay men using 6 gyms between the months of January and February (and you need to think of the type of bias there that those people who are ‘Resolutioners’ would be more likely to go to the gym those months), a questionnaire was given to the men. 15 .2 percent of the men had used, with 11.7 percent of them injecting within the past 12 months. HIV positive men were more likely to have used in the past compared to negative men (probably due to scripts). Fifty-one percent of them reported testicular atrophy, and they were more likely to report suicidal thoughts (Bolding, Sherr, and Elford, 2002). They conclude:
One in seven gay men surveyed in central London gyms in 2000 said they had used steroids in the previous 12 months. HIV positive men were more likely to have used steroids than other men, some therapeutically. Side effects were reported widely and steroid use was associated with having had suicidal thoughts and feeling depressed, although cause and effect could not be established. Our findings suggest that steroid use among gay men may have serious consequences for both physical and mental health.
Of course, those who (ab)use substances have more psychological problems than those who do not. Another study of 203 bodybuilders found that 8 percent (n = 17) found testicular atrophy (for what it’s worth, it was an internet survey of drug utilization) (Perry et al, 2005). Another study found that out of 88 percent of individuals who abused the drug complained of side-effects of AAS use, about 40 percent described testicular atrophy (Evans, 1997), while testicular atrophy was noted in about 50 percent of cases (sample size n = 24) (Darke et al, 2016).
Sperm production
One study of steroid users found that only 17 percent of them had normal sperm levels (Torres-Calleja et al, 2001), this is because exogenous testosterone will result in the atrophy of germinal cells which cause a decrease in spermatogenesis. Though, too, increased AAS (ab)use later into life may lead to infertility later in life. Knuth et al (1989) also studied 41 bodybuilders with an average age of 26.7. They went through a huge laundry list of different types of steroids they have taken over their lives. Nineteen of the men were still using steroids at the time of the investigation (group I), whereas 12 of them (group II) stopped taking steroids 3 months prior, while 10 of them (group III) stopped steroid use 4 to 24 months prior.
They found that only 5 of them had sperm counts below the average of 20 million sperm per square ml, while 24 of the bodybuilders showed these symptoms. No difference between group I and II was noticed and group III (the group that abstained from use for 4 to 24 months) largely had sperm levels in the normal range. So, the data suggests that even in cases of severe decrease of sensitivity to androgens due to AAS (ab)use, spermatogenesis may still continue normally in some men, even when high levels of androgens are administered exogenously, while even after prolonged use it seems it is possible for sperm levels to go back to the normal range (Knuth et al 1989).
Aggression and crime
Now it’s time for the fun part and my reason for writing this article. Does (ab)using steroids cause someone to go into an uncontrollable rage, a la the Incredible Hulk when they inject themselves with testosterone? The media has latched into the mind of many, with films and TV shows showing the insanely aggressive man who has been (ab)using AAS. But how true is this? A few papers have shown that this phenomenon is indeed true (Konacher and Workman, 1989; Pope and Katz, 1994), but how true is it on its own, since AAS (ab)users are known to use multiple substances???
Konacher and Workman (1989) is a case study done on one man who had no criminal history, who began taking AASs three months before he murdered his wife, and they conclude that AAS can be said to be a ‘personality changer’. Piacetino et al (2015) conclude in their review of steroid use and psychopathology in athletes that “AAS use in athletes is associated with mood and anxiety disturbances, as well as reckless behavior, in some predisposed individuals, who are likely to develop various types of psychopathology after long-term exposure to these substances. There is a lack of studies investigating whether the preexistence of psychopathology is likely to induce AAS consumption, but the bulk of available data, combined with animal data, point to the development of specific psycho-pathology, increased aggressiveness, mood destabilization, eating behavior abnormalities, and psychosis after AAS abuse/dependence.” I, too, would add that since most steroid abuse are polysubstance abusers (they use multiple illicit drugs on top of AAS), that the steroids per se are not causing crime or aggressive behavior, it’s the other drugs that the steroid (ab)user is also taking. And there is evidence for this assertion.
Lundholm et al (2015) showed just that: that AAS (ab)use was confounded with other substances used while the individual in question was also taking AAS. They write:
“We found a strong association between self-reported lifetime AAS use and violent offending in a population-based sample of more than 10,000 men aged 20-47 years. However, the association decreased substantially and lost statistical significance after adjusting for other substance abuse. This supports the notion that AAS use in the general population occurs as a component of polysubstance abuse, but argues against its purported role as a primary risk factor for interpersonal violence. Further, adjusting for potential individual-level confounders initially attenuated the association, but did not contribute to any substantial change after controlling for polysubstance abuse.“
Even The National Institute of Health (NIH) writes: “In summary, the extent to which steroid abuse contributes to violence and behavioral disorders is unknown. As with the health complications of steroid abuse, the prevalence of extreme cases of violence and behavioral disorders seems to be low, but it may be underreported or underrecognized.” We don’t know whether steroids cause aggression or more aggressive athletes are more likely to use the substance (Freberg, 2009: 424). Clearly, the claims of steroids causing aggressive behavior and crime are overblown and there has yet to be a scientific consensus on the matter. A great documentary on the matter is Bigger, Stronger, Faster, which goes through the myths of testosterone while chronicling the use of illicit drugs in bodybuilding and powerlifting.
This, too, was even seen in one study where men were administered supraphysiologic doses of testosterone to see its effects on muscle size and strength since it had never been tested; no changes in mood or behavior occurred (Bhasin et al, 1996). Furthermore, injecting individuals with supraphysiological doses of testosterone as high as 200 and 600 mg per week does not cause heightened anger or aggression (Tricker et al, 1996; O’Connor et, 2002). Testosterone is one of the most abused AASs around, and if a heightened level of T doesn’t cause crime, nor can testosterone levels being higher this week compared to last seem to be a trigger for crime, we can safely disregard any claims of ‘roid rage’ since they coincide with other drug use (polysubstance abuse). So since we know that supraphysiologic doses of testosterone don’t cause crime nor aggression, we can say that AAS use, on its own (and even with other drugs) does not cause crime or heightened aggression since aggression elevates testosterone secretion, testosterone doesn’t elevate aggression.
One review also suggests that medical issues associated with AAS (ab)use are exaggerated to deter their use by athletes (Hoffman and Ratamess, 2006). They conclude that “Existing data suggest that in certain circumstances the medical risk associated with anabolic steroid use may have been somewhat exaggerated, possibly to dissuade use in athletes.”
Racial differences in steroid use
Irving et al (2002) found that 2.1 percent of whites used steroids, whereas 7.6 percent of blacks did; 6.1 percent of ‘Hispanics’ use them within the past 12 months, and a whopping 14.1 percent of Hmong Chinese used them; 7.9 percent of ‘other Asians’ used them, and 3,1 percent of ‘Native Americans’ did with 11.3 percent of mixed race people using them within the past 12 months to gain muscle. Middle schoolers were more likely to use than high schoolers, while people from lower SES brackets were more likely to use than people in higher SES brackets.
Stilger and Yesalis (1999: 134) write (emphasis mine):
Of the 873 high school football players participating in the study, 54 (6.3%) reported having used or currently using AAS. Caucasians represented 85% of all subjects in the survey. Nine percent were African-American while the remainder (6%) consisted of Hispanics, Asian, and other. Of the AAS users, 74% were Caucasian, 13% African American, 7% Hispanic, and 3% Asian, x2 (4,854 4) 4.203, p 4 .38. The study also indicated that minorities are twice as likely to use AAS as opposed to Caucasians. Cross tabulated results indicate that 11.2% of all minorities use/used AAS as opposed to 6.5% of all Caucasians (data not displayed).
One study even had whites and blacks reporting the same abuse of steroids in their sample (n = 10,850 ‘Caucasians’ and n = 1,883 black Americans), with blacks reporting, too, lower levels of other drug abuse (Green et al, 2001). Studies indeed find higher rates of drug use for white Americans than other ethnies, in college (McCabe et al, 2007). Black Americans also frequently underreport and lie about their drug use (Ledgerwood et al, 2008; Lu et al, 2001). Blacks are also more likely to go to the ER after abusing drugs than whites (Drug Abuse Warning Network, 2011). Bauman and Ennett (1994) also found that blacks underreport drug use whereas whites overreport.
So can we really believe the black athletes who state that they do not (ab)use AAS? No, we cannot. Blacks like about any and all drug use, so believing that they are being truthful about AAS (ab)use in this specific instance is not called for.
Conclusion
Like with all things you use and abuse, there are always side-effects. Though, the media furor one hears regarding AAS and testosterone (ab)use are largely blown out of proportion. The risks associated with AAS (ab)use are ‘transient’, and will subside after one discontinues using the drugs. Blacks seem to take more AAS than whites, even if they do lie about any and all drug use. (And other races, too, seem to use it at higher rates than whites.) Steroid use does not seem to be ‘bad’ if one knows what they’re doing and are under Doctor’s supervision, but even then, if you want to know the truth about AAS, then you need to watch the documentary Bigger, Stronger, Faster. I chalk this up to the media themselves demonizing testosterone itself, along with the ‘toxic masculinity’ and the ‘toxic jock effect‘ (Miller, 2009; Miller, 2011). Though, if you dig into the literature yourself you’ll see there is scant evidence for AAS and testosterone (ab)use causing crime, that doesn’t stop papers like those two by Miller talking about the effects of ‘toxic jocks’ and in effect, deriding masculine men and with it the hormone that makes Men men: testosterone. If taken safely, there is nothing wrong with AAS/testosterone use.
(Note: Doctor’s supervision only, etc)
Should We End Sex Segregation in Sports? Should Athletes Be Assessed by Anatomy And Physiology?
1400 words
An opinion piece by sociologist Roslyn Kerr, senior lecturer in sociology of sport, from Lincoln University wrote an article on January 18th for The Conversation titled Why it might be time to eradicate sex segregation in sports where she argues against sex segregation in sports. She does publish articles on sports history, leisure studies and sports management and used to be a gymnast so she should have good knowledge—perhaps better than the general public—on anatomy and physiology and how they interact during elite sporting performances. Though is there anything to the argument she provides in her article? Maybe.
The paper is pretty good, though it, of course, uses sociological terms and cites feminist theorists talking about gender binaries in sports and how they’re not ‘fair’. One thing continously brought up in the paper is how there is no way to discern sex regarding sporting competitions (Simpson et al, 1993; Dickinson et al, 2002; Heggie, 2010), with even chromosome-based testing being thrown out (Elsas et al, 2000). which can be seen with the Olympics “still struggling to define gender“. They state that women are put through humiliating tests to discern their sex.
They use this to buttress their own arguments which are based off of what bodies of disables athletes did: whether or not one competed in a particular sport was not on their disability, per se, but the functionality of their own bodies. As an example, sporting bodies used to group people with, say, a similar spinal injury even though they had different physical abilities. Call me crazy, but I most definitely see the logic that these authors are getting at, and not only because I ruminated on something similar back in the summer in an article on transgendered athletes in sports, writing:
This then brings up some interesting implications. Should we segregate competitions by race since the races have strength and weaknesses due to biology and anatomy, such as somatype? It’s an interesting question to consider, but I think we can all agree on one thing: Women should compete with women, and men should compete with men. Thus, transgenders should compete with transgenders.
Of course I posed the question regarding different races since they have different strengths and weaknesses on average due to evolution in different environments. Kerr and Obel (2017) conclude (pg 13):
Numerous authors have noted that the current two-sex classification system is problematic. They argued that it does not include all bodies, such as intersex bodies, and more importantly, does not work to produce fair competition. Instead, some argued that other traits that we know influence sporting success should be used to classify bodies. In this article, we extended this idea through using the ANT concepts of assemblage and black box. Specifically, we interpreted the current understanding of the body that sex segregation is based on as a black box that assumes the constant superiority of the male body over the female. But we argued that with the body understood as an assemblage, this classification could be reassembled so that this black box is no longer given. Instead we argued that by identifying the multiple traits that make up the assemblage of sporting success, sex classification becomes irrelevant and that it is these traits that we should use to classify athletes rather than sex. Drawing on the example of disability sport we noted that the black box of a medical label was undone and replaced with an emphasis on functionality with different effects for each sport. This change had the effect of undoing rigid medical disability label and enabling athletes’ bodies to be viewed as assemblages consisting of various functional and potentially changing physical abilities. We used this discussion to propose a model of classified that eliminated the need for sex segregation and instead used physical measures such as LBM and VO2 capabilities to determine an athlete’s competitive class.
All of their other arguments aside that I disagree with in their paper (their use of ‘feminist theory’, gendered divisions, short discussions and quotes from other authors on the ‘power structure’ of males), I definitely see the logic here and, in my opinion, it makes sense. Anyway, those shortcomings aside, the actual argument of using anatomy and physiology and seeing which different parts work in concert to produce elite athletic performance in certain sports then having some kind of test, say, the Heath-Carter method for somatype (Wilmore, 1970) to a test of Vo2 max (Cureton et al, 1986) to even lean body mass (LBM).
Healy et al (2014) studied 693 elite athletes in a post-competition setting. They assesed testosterone, among other variables such as aerobic performance. They observed a difference of 10 of between men and women’s LBM and that it exclusively accounts for the “observed diffences in strength and aerobic performance seen between the sexes” while they conclude:
We have shown that despite differences in mean testosterone level between genders, there is complete overlap of the range of concentrations seen. This shows that the recent decision of the IOC and IAAF to limit participation in elite events to women with ‘normal’ serum testosterone is unsustainable.
Yes, this testosterone-influences-sports-performance is still ongoing. I’ve covered it a bit last year, and while I believe there is a link between testosterone and athletic ability and have provided some data and a few anecdotes from David Epstein, I do admit that the actual literature is scant with conclusive evidence that testosterone positively influences sport performance. Either way, if testosterone truly does infer an advantage then, of course, the model (which Kerr and Obel admit is simple at the moment) will need to be slightly revised. Arguments and citations can be found in this article written back in the summer on whether or not transgender MtFs should compete with women. This is also directly related to the MtF who dominated women a few months back.
Either way, the argument that once we better identify anatomic and physiologic causes for differences in certain sporting competition, this could, in theory, be used instead of sex segregation. I think it’s a good idea personally and to see how effective it could be there should be a trial run on it. Kerr and Obel state that it would make competition more ‘fair’. However, Sanchez et al, 2014 cite Murray (2010) who writes “fair sports competition does not require that athletes be equal in every imaginable respect.”
At the end of the day, what a lot of this rests on is whether or not testosterone infers athletic advantage at the elite level and there is considerable data for both sides. It’ll be interesting to see how the major sporting bodies handle the question of testosterone in sports and transgenders and hyperandrogenic females.
Personally, I think there may be something to Kerr and Obel’s arguments in their paper (feminist/patriarchy garbage aside) since it’s based on anatomy and physiology which is what we see on the field. However, it can also be argued that sex/gender is manifested in the brain which then infers other advantages/disadvantages in sports. Nonetheless, I think the argument in the paper is sound (the anatomy and physiology arguments only). For instance, we can look at one sport, say, 100 m dash, and we can say “OK, we know that sprinters have meso-ecto somatypes and that combined with the RR ACTN3 genotype, that confers elite athletic performance (Broos et al, 2016).” We could use those two variables along with leg length, foot length etc and then we can test—both in the lab and on the field—which variables infer advantages in certain sports. Another sport we can think of is swimming. Higher levels of body fat with wide clavicles and chest cavity are more conducive to swimming success. We could use those types of variables for swimming and so on.
Of course, this method may not work or it may only work in theory but not work in practice. Using lean body mass, Vo2 max etc etc based on which sport is in question may be better than using the ‘sex binary’, since some women (trust me, I’ve trained hundreds) would be able to compete head-to-head with men and, if for nothing else, it’d be good entertainment.
However, in my opinion, the logic on using anatomy and physiology instead of sex to segregate in sports is intriguing and, if nothing else, would finally give feminists (and non-feminists) the ‘equality’ they ask for.
Race, Testosterone, Aggression, and Prostate Cancer
4050 words
Race, aggression, and prostate cancer are all linked, with some believing that race is the cause of higher testosterone which then causes aggression and higher rates of crime along with maladies such as prostate cancer. These claims have long been put to bed, with a wide range of large analyses.
The testosterone debate regarding prostate cancer has been raging for decades and we have made good strides in understanding the etiology of prostate cancer and how it manifests. The same holds true for aggression. But does testosterone hold the key to understanding aggression, prostate cancer and does race dictate group levels of the hormone which then would explain some of the disparities between groups and individuals of certain groups?
Prostate cancer
For decades it was believed that heightened levels of testosterone caused prostate cancer. Most of the theories to this day still hold that large amounts of androgens, like testosterone and it’s metabolic byproduct dihydrotestosterone, are the two many factors that drive the proliferation of cells and therefore, if a male is exposed to higher levels of testosterone throughout their lives then they are at a high risk of prostate cancer compared to a man with low testosterone levels, so the story goes.
In 1986 Ronald Ross set out to test a hypothesis: that black males were exposed to more testosterone in the womb and this then drove their higher rates of prostate cancer later in life. He reportedly discovered that blacks, after controlling for confounds, had 15 percent higher testosterone than whites which may be the cause of differential prostate cancer mortality between the two races (Ross et al, 1986) This is told in a 1997 editorial by Hugh McIntosh. First, the fact that black males were supposedly exposed to more testosterone in the womb is brought up. I am aware of one paper discussing higher levels of testosterone in black women compared to white women (Perry et al, 1996). Though, I’ve shown that black women don’t have high levels of testosterone, not higher than white women, anyway (see Mazur, 2016 for discussion). (Yes I changed my view on black women and testosterone, stop saying that they have high levels of testosterone it’s just not true. I see people still link to that article despite the long disclaimer at the top.)
Alvarado (2013) discusses Ross et al (1986), Ellis and Nyborg (1992) (which I also discussed here along with Ross et al) and other papers discussing the supposed higher testosterone of blacks when compared to whites and attempts to use a life history framework to explain higher incidences of prostate cancer in black males. He first notes that nutritional status influences testosterone production which should be no surprise to anyone. He brings up some points I agree with and some I do not. For instance, he states that differences in nutrition could explain differences in testosterone between Western and non-Western people (I agree), but that this has no effect within Western countries (which is incorrect as I’ll get to later).
He also states that ancestry isn’t related to prostate cancer, writing “In summation, ancestry does not adequately explain variation among ethnic groups with higher or lower testosterone levels, nor does it appear to explain variation among ethnic groups with high or low prostate cancer rates. This calls into question the efficacy of a disease model that is unable to predict either deleterious or protective effects.”
He then states that SES is negatively correlated with prostate cancer rates, and that numerous papers show that people with low SES have higher rates of prostate cancer mortality which makes sense, since people in a lower economic class would have less access to and a chance to get good medical care to identify problems such as prostate cancer, including prostate biopsies and checkups to identify the condition.
He finally discusses the challenge hypothesis and prostate cancer risk. He cites studies by Mazur and Booth (who I’ve cited in the past in numerous articles) as evidence that, as most know, black-majority areas have more crime which would then cause higher levels of testosterone production. He cites Mazur’s old papers showing that low-class men, no matter if they’re white or black, had heightened levels of testosterone and that college-educated men did not, which implies that the social environment can and does elevate testosterone levels and can keep them heightened. Alvarado concludes this section writing: “Among Westernized men who have energetic resources to support the metabolic costs associated with elevated testosterone, there is evidence that being exposed to a higher frequency of aggressive challenges can result in chronically elevated testosterone levels. If living in an aggressive social environment contributes to prostate cancer disparities, this has important implications for prevention and risk stratification.” He’s not really wrong but on what he is wrong I will discuss later on this section. It’s false that testosterone causes prostate cancer so some of this thesis is incorrect.
I rebutted Ross et al (1986) December of last year. The study was hugely flawed and, yet, still gets cited to this day including by Alvarado (2013) as the main point of his thesis. However, perhaps most importantly, the assay times were done ‘when it was convenient’ for the students which were between 10 am and 3 pm. To not get any wacky readings one most assay the individuals as close to 8:30 am as possible. Furthermore, they did not control for waist circumference which is another huge confound. Lastly, the sample was extremely small (50 blacks and 50 whites) and done on a nonrepresentative sample (college students). I don’t think anyone can honestly cite this paper as any evidence for blacks having higher levels of testosterone or testosterone causing prostate cancer because it just doesn’t do that. (Read Race, Testosterone and Prostate Cancer for more information.)
What may explain prostate cancer rates if not for differences in testosterone like has been hypothesized for decades? Well, as I have argued, diet explains a lot of the variation between races. The etiology of prostate cancer is not known (ACA, 2016) but we know that it’s not testosterone and that diet plays a large role in its acquisition. Due to their dark skin, they need more sunlight than do whites to synthesize the same amount of vitamin D, and low levels of vitamin D in blacks are strongly related to prostate cancer (Harris, 2006). Murphy et al (2014) even showed, through biopsies, that black American men had higher rates of prostate cancer if they had lower levels of vitamin D. Lower concentrations of vitamin D in blacks compared to whites due to dark pigmentation which causes reduced vitamin D photoproduction and may also account for “much of the unexplained survival disparity after consideration of such factors as SES, state at diagnosis and treatment” (Grant and Peiris, 2012).
Testosterone
As mentioned above, testosterone is assumed to be higher in certain races compared to others (based on flawed studies) which then supposedly exacerbates prostate cancer. However, as can be seen above, a lot of assumptions go into the testosterone-prostate cancer hypothesis which is just false. So if the assumptions are false about testosterone, mainly regarding racial differences in the hormone and then what the hormone actually does, then most of their claims can be disregarded.
Perhaps the biggest problem is that Ross et al is a 32-year-old paper (which still gets cited favorably despite its huge flaws) while our understanding of the hormone and its physiology has made considerable progress in that time frame. So it’s in fact not so weird to see papers like this that say “Prostate cancer appears to be unrelated related to endogenous testosterone levels” (Boyle et al, 2016). Other papers also show the same thing, that testosterone is not related to prostate cancer (Stattin et al, 2004; Michaud, Billups, and Partin, 2015). This kills a lot of theories and hypotheses, especially regarding racial differences in prostate cancer acquisition and mortality. So, what this shows is that even if blacks did have 15 percent higher serum testosterone than whites as Ross et al, Rushton, Lynn, Templer, et al believed then it wouldn’t cause higher levels of prostate cancer (nor aggression, which I’ll get into later).
How high is testosterone in black males compared to white males? People may attempt to cite papers like the 32-year-old paper by Ross et al, though as I’ve discussed numerous times the paper is highly flawed and should therefore not be cited. Either way, levels are not as high as people believe and meta-analyses and actual nationally representative samples (not convenience college samples) show low to no difference, and even the low difference wouldn’t explain any health disparities.
One of the best papers on this matter of racial differences in testosterone is Richard et al (2014). They meta-analyzed 15 studies and concluded that the “racial differences [range] from 2.5 to 4.9 percent” but “this modest difference is unlikely to explain racial differences in disease risk.” This shows that testosterone isn’t as high in blacks as is popularly misconceived, and that, as I will show below, it wouldn’t even cause higher rates of aggression and therefore criminal behavior. (Rohrmann et al 2007 show no difference in testosterone between black and white males in a nationally representative sample after controlling for lifestyle and anthropometric variables. Whereas Mazur, 2009 shows that blacks have higher levels of testosterone due to low marriage rates and lower levels of adiposity, while be found a .39 ng/ml difference between blacks and whites aged 20 to 60. Is this supposed to explain crime, aggression, and prostate cancer?)
However, as I’ve noted last year (and as Alvarado, 2013 did as well), young black males with low education have higher levels of testosterone which is not noticed in black males of the same age group but with more education (Mazur, 2016). Since blacks of a similar age group have lower levels of testosterone but are more highly educated then this is a clue that education drives aggression/testosterone/violent behavior and not that testosterone drives it.
Mazur (2016) also replicated Assari, Caldwell, and Zimmerman’s (2014) finding that “Our model in the male sample suggests that males with higher levels of education has lower aggressive behaviors. Among males, testosterone was not associated with aggressive behaviors.” I know this is hard for many to swallow that testosterone doesn’t lead to aggressive behavior in men, but I’ll cover that in the last and final section.
So it’s clear that the myth that Rushton, Lynn, Templer, Kanazawa, et al pushed regarding hormonal differences between the races are false. It’s also with noting, as I did in my response to Rushton on r/K selection theory, that the r/K model is literally predicated on 1) testosterone differences between races being real and in the direction that Rushton and Lynn want because they cite the highly flawed Ross et al (1986) and 2) testosterone does not cause higher levels of aggression (which I’ll show below) which then lead to higher rates of crime along with higher rates of incarceration.
A blogger who goes by the name of ethnicmuse did an analysis of numerous testosterone papers and he found:
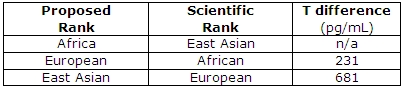 Which, of course, goes against a ton of HBD theory, that is, if testosterone did what HBDers believed it does (it doesn’t). This is what it comes down to: blacks don’t have higher levels of testosterone than whites and testosterone doesn’t cause aggression nor prostate cancer so even if this relationship was in the direction that Rushton et al assert then it still wouldn’t cause any of the explanatory variables they discuss.
Which, of course, goes against a ton of HBD theory, that is, if testosterone did what HBDers believed it does (it doesn’t). This is what it comes down to: blacks don’t have higher levels of testosterone than whites and testosterone doesn’t cause aggression nor prostate cancer so even if this relationship was in the direction that Rushton et al assert then it still wouldn’t cause any of the explanatory variables they discuss.
Last year Lee Ellis published a paper outlining his ENA theory (Ellis, 2017). I responded to the paper and pointed out what he got right and wrong. He discussed strength (blacks aren’t stronger than whites due to body type and physiology, but excel in other areas); circulating testosterone, umbilical cord testosterone exposure; bone density and crime; penis size, race, and crime (Rushton’s 1997 claims on penis size don’t ‘size up’ to the literature as I’ve shown two times); prostate-specific antigens, race, and prostate cancer; CAG repeats; intelligence and education and ‘intelligence’; and prenatal androgen exposure. His theory has large holes and doesn’t line up in some places, as he himself admits in his paper. He, as expected, cites Ross et al (1986) favorably in his analysis.
Testosterone can’t explain all of these differences, no matter if it’s prenatal androgen exposure or not, and a difference of 2.5 to 4.9 percent between blacks and whites regarding testosterone (Richard et al, 2014) won’t explain differences in crime, aggression, nor prostate cancer.
Other authors have attempted to also implicate testosterone as a major player in a wide range of evolutionary theories (Lynn, 1990; Rushton, 1997; Rushton, 1999; Hart, 2007; Rushton and Templer, 2012; Ellis, 2017). However, as can be seen by digging into this literature, these claims are not true and therefore we can discard the conclusions come to by the aforementioned authors since they’re based on false premises (testosterone being a cause for aggression, crime, and prostate cancer and r/K meaning anything to human races, it doesn’t)
Finally, to conclude this section, does testosterone explain racial differences in crime? No, racial differences in testosterone, however small, cannot be responsible for the crime gap between blacks and whites.
Testosterone and aggression
Testosterone and aggression, are they linked? Can testosterone tell us anything about individual differences in aggressive behavior? Surprisingly for most, the answer seems to be a resounding no. One example is the castration of males. Does it completely take away the urge to act aggressively? No, it does not. What is shown when sex offenders are castrated is that their levels of aggression decrease, but importantly, they do not decrease to 0. Robert Sapolsky writes on page 96 of his book Behave: The Biology of Humans at Our Best and Worst (2017) (pg 96):
… the more experience a male has being aggressive prior to castration, the more aggression continues afterward. In other words, the less his being aggressive in the future requires testosterone and the more it’s a function of social learning.
He also writes (pg 96-97):
On to the next issue that lessens the primacy of testosterone: What do individual levels of testosterone have to do with aggression? If one person higher testosterone levels than another, or higher levels this week than last, are they more likely to be aggressive?
Initially the answer seemed to be yes, as studies showed correlation between individual differences in testosterone levels and levels of aggression. In a typical study, higher testosterone levels would be observed in those male prisoners with higher rates of aggression. But being aggressive stimulates testosterone secretion; no wonder more aggressive individuals had higher levels. Such studies couldn’t disentangle chickens and eggs.
Thus, a better question is whether differences in testosterone levels among individuals predict who will be aggressive. And among birds, fish, mammals, and especially other primates, the answer is generally no. This has been studied extensively in humans, examining a variety of measures of aggression. And the answer is clear. To quote British endocrinologist John Archer in a definitive 2006 review, “There is a weak and inconsistent association between testosterone levels and aggression in [human] adults, and . . . administration of testosterone to volunteers typically does not increase aggression.” The brain doesn’t pay attention to testosterone levels within the normal range.
[…]
Thus, aggression is typically more about social learning than about testosterone, differing levels of testosterone generally can’t explain why some individuals are more aggressive than others.
Sapolsky also has a 1997 book of essays on human biology titled The Trouble With Testosterone: And Other Essays On The Biology Of The Human Predicament and he has a really good essay on testosterone titled Will Boys Just Be Boys? where he writes (pg 113 to 114):
Okay, suppose you note a correlation between levels of aggression and levels of testosterone among these normal males. This could be because (a) testosterone elevates aggression; (b) aggression elevates testosterone secretion; (c) neither causes the other. There’s a huge bias to assume option a while b is the answer. Study after study has shown that when you examine testosterone when males are first placed together in the social group, testosterone levels predict nothing about who is going to be aggressive. The subsequent behavioral differences drive the hormonal changes, not the other way around.
Because of a strong bias among certain scientists, it has taken do forever to convince them of this point.
[…]
As I said, it takes a lot of work to cure people of that physics envy, and to see interindividual differences in testosterone levels don’t predict subsequent differences in aggressive behavior among individuals. Similarly, fluctuations in testosterone within one individual over time do not predict subsequent changes in the levels of aggression in the one individual—get a hiccup in testosterone secretion one afternoon and that’s not when the guy goes postal.
And on page 115 writes:
You need some testosterone around for normal levels of aggressive behavior—zero levels after castration and down it usually goes; quadruple it (the sort of range generated in weight lifters abusing anabolic steroids), and aggression typically increases. But anywhere from roughly 20 percent of normal to twice normal and it’s all the same; the brain can’t distinguish among this wide range of basically normal values.
Weird…almost as if there is a wide range of ‘normal’ that is ‘built in’ to our homeodynamic physiology…
So here’s the point: differences in testosterone between individuals tell us nothing about individual differences in aggressive behavior; castration and replacement seems to show that, however broadly, testosterone is related to aggression “But that turns out to not be true either, and the implications of this are lost on most people the first thirty times you tell them about it. Which is why you’d better tell them about it thirty-one times, because it’s the most important part of this piece” (Sapolsky, 1997: 115).
Later in the essay, Sapolsky discusses a discusses 5 monkeys that were given time to form a hierarchy of 1 through 5. Number 3 can ‘throw his weight’ around with 4 and 5 but treads carefully around 1 and 2. He then states to take the third-ranking monkey and inject him with a ton of testosterone, and that when you check the behavioral data that he’d then be participating in more aggressive actions than before which would imply that the exogenous testosterone causes participation in more aggressive behavior. But it’s way more nuanced than that.
So even though small fluctuations in the levels of the hormone don’t seem to matter much, testosterone still causes aggression. But that would be wrong. Check out number 3 more closely. Is he now raining aggression and terror on any and all in the group, frothing in an androgenic glaze of indiscriminate violence. Not at all. He’s still judiciously kowtowing to numbers 1 and 2 but has simply become a total bastard to number 4 and 5. This is critical: testosterone isn’t causing aggression, it’s exaggerating the aggression that’s already there.
The correlation between testosterone and aggression is between .08 and .14 (Book, Starzyk, and Quinsey, 2001; Archer, Graham-Kevan, and Davies, 2005; Book and Quinsey, 2005). Therefore, along with all of the other evidence provided in this article, it seems that testosterone and aggression have a weak positive correlation, which buttresses the point that aggression concurrent increases in testosterone.
Sapolsky then goes on to discuss the amygdala’s role in fear processing. The amygdala has its influence on aggressive behavior through the stria terminalis, which is a bunch of neuronal connections. How the amygdala influences aggression is simple: bursts of electrical excitation called action potentials go up and down the stria terminalis which changes the hypothalamus. You can then inject testosterone right into the brain and will it cause the same action potentials that surge down the stria terminalis? No, it does not turn on the pathway at all. This only occurs only if the amygdala is already sending aggression-provoking action potentials down the stria terminalis with testosterone increasing the rate of action potentials you’re shortening the rest time between them. So it doesn’t turn on this pathway, it exaggerates the preexisting pattern, which is to say, it’s exaggerating the response to environmental triggers of what caused the amygdala to get excited in the first place.
He ends this essay writing (pg 119):
Testosterone is never going to tell us much about the suburban teenager who, in his after-school chess club, has developed a particularly aggressive style with his bishops. And it certainly isn’t going to tell us much about the teenager in some inner-city hellhole who has taken to mugging people. “Testosterone equals aggression” is inadequate for those who would offer a simple solution to the violent male—just decrease levels of those pesky steroids. And “testosterone equals aggression” is certainly inadequate for those who would offer a simple excuse: Boys will be boys and certain things in nature are inevitable. Violence is more complex than a single hormone. This is endocrinology for the bleeding heart liberal—our behavioral biology is usually meaningless outside of the context of social factors and the environment in which it occurs.
Injecting individuals with supraphysiological doses of testosterone as high as 200 and 600 mg per week does not cause heightened anger or aggression (Tricker et al, 1996; O’Connor et, 2002). This, too, is a large blow for the testosterone-induces-aggression hypothesis. Because aggressive behavior heightens testosterone, testosterone doesn’t heighten aggressive behavior. (This is the causality that has been looked for, and here it is. The causality is not in the other direction.) This tells us that we need to be put into situations for our aggression to rise and along with it, testosterone. I don’t even see how people could think that testosterone could cause aggression. It’s obvious that the environmental trigger needs to be there first in order for the body’s physiology to begin testosterone production in order to prepare for the stimulus that caused the heightened testosterone production. Once the trigger occurs, then it can and does stay heightened, especially in areas where dominance contests would be more likely to occur, which would be low-income areas (Mazur, 2006, 2016).
(Also read my response to Batrinos, 2012, my musings on testosterone and race, and my responses to Robert Lindsay and Sean Last.)
Lastly, one thing that gets on my nerves that people point to to attempt to show that testosterone and its derivatives cause violence, aggression etc is the myth of “roid rage” which is when an individual objects himself with testosterone, anabolic steroids or another banned substance, and then the individual becomes more aggressive as a result of more free-flowing testosterone in their bloodstream.
The problem here is that people believe what they hear on the media about steroids and testosterone, and they’re largely not true. One large analysis was done to see the effects of steroids and other illicit drug use on behavior, and what was found was that after controlling for other substance use “Our results suggest that it was not lifetime steroid use per se, but rather co-occurrring polysubstance abuse that most parsimoniously explains the relatively strong association of steroid use and interpersonal violence” (Lundholm et al, 2015). So after controlling for other drugs used, men who use steroids do not go to prison and be convicted of violence after other polysubstance use was controlled for, implying that is what’s driving interpersonal violence, not the substance abuse of steroids.
Conclusion
Numerous myths about testosterone have been propagated over the decades, which are still believed in the new millennium despite numerous other studies and arguments to the contrary. As can be seen, the myths that people believe about testosterone are easily debunked. Numerous papers (with better methodology than Ross et al) attest to the fact that testosterone levels aren’t as high as was believed decades ago between the races. Diet can explain a lot of the variation, especially vitamin D intake. Injecting men with supraphysiological doses of testosterone does not heighten anger nor aggression. It does not even heighten prostate cancer severity.
Racial differences in testosterone are also not as high as people would like to believe, there is even an opposite relationship with Asians having higher levels and whites having lower (which wouldn’t, on average, imply femininity) testosterone levels. So as can be seen, the attempted r/K explanations from Rushton et al don’t work out here. They’re just outright wrong on testosterone, as I’ve been arguing for a long while on this blog.
Testosterone doesn’t cause aggression, aggression causes heightened testosterone. It can be seen from studies of men who have been castrated that the more crime they committed before castration, the more crime they will commit after which implies a large effect of social learning on violent behavior. Either way, the alarmist attitudes of people regarding testosterone, as I have argued, are not needed because they’re largely myths.
Rebutting Ross et al (1986) on Testosterone
1300 words
Ross et al (1986) is one of the most oft-cited papers that HBDers use to attempt to show that blacks have higher levels of testosterone than whites, which then—supposedly—goes on to explain higher rates of crime, aggression, and prostate cancer. However, people 1) say this only from reading the abstract (and not reading the full paper) and 2) even if they could read the paper they would not know where the flaws were to point them out and discredit the study based on flawed methodology. I see this study getting cited every now-and-then and I’m sick of seeing it.
Ross et al (1986: 45) state that they “recently reviewed the evidence that endogenous levels of certain steroid and polypeptide hormones are causally related to a group of human cancers, including cancer of the prostate gland.” I’ve shown how even injecting a man with exogenous testosterone does not worsen his prostate cancer (Eisenberg et al, 2015; Boyle et al, 2016) and testosterone doesn’t cause prostate cancer (Stattin et al, 2003; Michaud, Billups, and Partin, 2015). So this has been falsified. Even if blacks had the testosterone levels that they claimed it still would not cause higher rates of PCa (prostate cancer) incidence.
They solicited study participants from two colleges around the Los Angeles metro area. The two universities they got their sample from were the University of Southern California and California State University of Los Angeles. They recruited individuals through postings on the school bulletin board in in the school newspaper. They got 50 blacks and 50 whites. They then write something that’s troubling to me: “A convenient time for blood collection was arranged, and students were met by a nurse epidemiologist (R. H.) at either the Student Health Center or another mutually convenient location” (Ross et al, 1986: 45). This is dumb. The students were assayed at all times between 10 am and 3 pm; testosterone levels are highest at 8 am though one study on older men shows that assaying between 8 am and 2 pm doesn’t matter (Crawford et al, 2015). However, for the purposes of discussing this paper this is irrelevant.

Table 1 from Ross et al (1986) tells us a lot about the flaws in the study—most importantly, the assay times. A majority were assayed between 10 am and 2 pm—which would depress testosterone though people assayed nearer to 10 am would have higher levels than people assayed nearer to 2 pm. Alcohol consumption only decreases testosterone while drunk, and a majority of the cohort did not consume alcohol within 12 hours of being assayed.
They come to the conclusion that the mean total testosterone level was 19 percent higher than whites whereas free testosterone was 21 percent higher. In regard to the assay collection times, Ross et al (1986: 47) write:
There was a negative correlation between time of sampling (No. of min elapsed since 0600 hr) and testosterone levels for whites (r=-O.4I) but not for blacks (r = -0.08). Adjustment for this variable caused a small reduction in geometric mean differences in levels of testosterone and free testosterone between blacks and whites. After simultaneous adjustment by analysis of covariance for time of sampling and age, weight, alcohol use, smoking, and use of prescription drugs, there
remained a 15% difference in total testosterone levels and a 13% difference in free testosterone levels between blacks and whites.
Even though they ‘adjusted for this variable’, it’s still a huge confound. Testosterone assays must be taken nearer to 8 am; the fact that people were assayed all over the place in the span of a 5 hour time period while testosterone levels decrease throughout the day is a huge red flag.
They then say that they are “uncertain why young black men have higher levels of circulating testosterone than white men“, though small sample sizes, a large range of variation in assay time, and a nonrepresentative sample is why. Other, more robust, analyses show a smaller ‘gap’, about 2.5 to 4.9 percent, favoring blacks (Richard et al, 2014). All in all, this study has huge flaws and should not be pointed to—especially today in 2017—because much larger analyses with much better methodologies have been carried out and some studies show no difference while others show a small difference favoring blacks but that still would not explain higher rates of testosterone, aggression and crime.
Ross et al (1986) is used by hereditarians such as Lynn (1990), Rushton (1997) and Hart (2007). Lynn (1990) states that these large testosterone differences discovered by Ross et al (1986) lend credence to Rushton’s r/K selection theory in which blacks have more children than whites who have more children than East Asians. Evidence for this assertion, states Lynn (1990) is the fact that blacks have higher rates of prostate cancer than whites who have higher levels of testosterone than East Asians, however this has been disproven by ethnicmuse.
Rushton (1997: 170) states that blacks had 19 percent higher levels of testosterone citing Ross et al (1986), however, Rushton didn’t cite the adjusted level which ended up being 15 percent, and, again, doesn’t mean anything to their hypothesis.
And Hart (2007) yet still repeats the same old stuff that “these differences in sexual behavior may be a consequence of the fact that blacks, on average, have higher levels of testosterone than whites“. These three researchers, clearly, are citing this study uncritically because it fits with their ‘racial hierarchy’. In fact, Rushton (1999) asked if testosterone was a ‘master switch’. In this paper, he cites Ellis and Nyborg (1992) who find that blacks had 3 percent higher levels of testosterone than whites. They gave the following values:
For the purposes of Rushton’s interpretation, writes Fish (2013), “These uncorrected figures are, of course, not consistent with their racial r- and K-continuum.” This, of course, is a big deal. Rushton cites this study as if it lends credence to his claims but it actually found the same result as Richard et al (2014). Thirty years after Ross et al (1986) we have numerous other studies showing a small gap between the races or no gap at all. We have numerous other studies showing that testosterone does not cause aggression, crime or violent behavior. However, people will still point to the abstract of Ross et al (1986) and think that they have proven that blacks have higher levels of testosterone than whites which proves how and why they have higher levels of testosterone, commit more crime and are all around more violent.
Though, as I have written about before, what Ross et al (1986) set out find the answer for (testosterone influencing higher levels of prostate cancer) can most definitely be explained by diet and lack of certain vitamins such as vitamin D, since low levels of this steroid hormone (it’s not a vitamin) cause prostate cancer (Schwartz and Hulka, 1990; Zhao and Feldman, 2001; Khan and Partin, 2004; Garland et al, 2006). Diet can explain a lot of the variation, as I have argued in the past.
In sum, Ross et al (1986) is the only study that I’ve found on racial testosterone differences that shows that extreme directionality favoring blacks over whites. This should set off some alarms in some people’s heads. People—psychologists included—need to learn about these hormones, how they’re produced in the body, and what they’re used for. Hormones don’t cause behavior, hormones influence behavior.
This fear of testosterone is largely overblown. We need testosterone for proper normal functioning. We need testosterone to be socially dominant; if you have lower levels you’ll be less socially dominant. This fear of testosterone—especially when it comes to race and it’s so-called causes—is largely pushed by Lynn, Rushton, Hart, and Ellis. I have spent a lot of time and thousands of words showing that they are wrong and testosterone is not a hormone to fear. It does not cause crime. It does not cause aggression. It does not cause prostate cancer. I’ve rebutted quite a few hereditarians on testosterone as well as testosterone and race, and if I come across more I will rebut them as well.
Testosterone, Physical Aggression and Social Dominance in Adolesence
1300 words
Testosterone and social dominance are linked, that is, when one is socially dominant then they will have higher levels of testosterone. But does this necessarily translate into violent behavior? No it does not. One longitudinal study looked at the effect of testosterone in children from kindergarten to the end of primary school (ages 6 to 13). Were high levels of testosterone associated with physical aggression and/or social dominance?
Schaal et al (1996) followed 178 boys from kindergarten to the end of primary school. To attempt to control for certain variables, they “were recruited according to the following criteria: (1) attending school in low socioeconomic areas of Montreal; (2) born from Caucasian, French-speaking parents themselves born in Canada; and (3) living with parents having medium to low educational status” (Schaal et al 1996).
They collected behavioral assessments at age 6, 10 and 13 through teacher and peer ratings. They asked questions such as “Who would you choose as a leader?” and “Who is the toughest?” To assess this, the authors made a game where they had to toss sandbags at targets to win money, with the only rules being they had to be a certain distance away from the targets. At the end of the game, the winner received 2 dollars while the others received 1 dollar. Each individual received a ‘toughness’ and ‘dominance’ score. Schaal et al (1996) write:
The crossing of both toughness and leadership scores, using the median, yielded four groups defined as follows: (1) tough-leader (n = 52); (2) toughnot leader (n = 27); (3) not tough-leader (n = 44); and (4) not tough -not leader (n = 48).
The groups did not differ on demographic or socioeconomic status, however, they showed differing ratings than they did at school in the previous year. Those rated tough by peers unfamiliar to them had also been rated more aggressive by peers who knew them than those who were rated ‘not tough’.
They assessed behavior and anxiety levels using a social behavior questionnaire. They write:
Boys were entered into the following categories according to their behavior rating scores on at least three of four assessments: (1) stable high fighter/stable high anxious (SH F-SHA; n = 20): fighting and anxiety scores at the 70th percentile or greater; (2) stable high fighter/ stable low anxious (SHF-SLA; n = 11): fighting score equaled or exceeded the 70th percentile and anxiety score was equal to or below the 50th percentile; (3) stable low fighter/stable high anxious (SLF-SHA; n = 10): equal to or less than the 50th percentile on the fighting scale and greater than the 70th percentile on the anxiety scale; and (4) stable low fighter/stable low anxious (SLFSLA; n = 25): equal to or lower than the 50th percentile on both fighting and anxiety scores.

Children who were noted by teachers to be more physically aggressive compared with children noted not to be physically aggressive were rated higher on “proactive aggression.” These children were also rated less popular.
They collected saliva to assay for testosterone at 8:30 am (yay), 10 am, 11 am, and 3:30 pm. There is a direct correlation between salivary testosterone levels and free testosterone levels in the body (Wang et al, 1981; Johnson, Joplin, and Burrin, 1987), and since it’s easier and way less of a biohazard to assay saliva for testosterone levels than to assay blood, it’s only logical to assay saliva since it’s easier and less of a chance for contaminated samples.
Now comes the fun part: did the socially dominant, tough leaders who were to have higher levels of testosterone more physically aggressive? To make it short: NO. The children who had LOWER testosterone were seen to be more physically aggressive. The ‘tough-not’ leaders were seen to be more aggressive; they had lower testosterone than both tough-not groups and lower than the tough leaders. Is this not ironic? This completely flys in the face of numerous theories on how and why grade-school children fight: it’s not high testosterone that causes this phenomenon, clearly.
It’s also interesting to note that those boys who were “stable high fighters” had lower testosterone levels compared to boys that were “stable low fighters” at age 13. Forty-one percent of the “stable high fighters “were in a regular classroom at the level appropriate for their age (age 13) compared with 91% of the stable low fighters” (Schaal et al 1996: 1327). The first analysis they conducted showed that, after only having met for three hours, the boys with high salivary testosterone were rated as “tough” and a “leader”. However—and this is where things get good—boys who were consistently rated as having higher levels of aggression between the ages of 6 and 12 had lower levels of testosterone at age 13.
Though, boys with high testosterone at age 13 “appear to be individuals who quickly succeed in imposing their will on peers, sometimes by aggressive behavior, but remain socially attractive over time” (Schaal et al, 1996: 1328). And, of course, boys who were rated as physically aggressive from kindergarten to age 12 (who had lower testosterone levels) were not seen as socially dominant. These children were seen as less popular and also were failing in school.
Elevated testosterone levels at age 13 were seen to be associated with social dominance when assessed after the game and three hour meet discussed above. Further, lower levels of testosterone between the ages of 6 to 12 (and assayed at 13) were associated with higher levels of aggression and more fighting. They state that higher levels of testosterone are only associated with aggression “only if the latter confers a dominant status” (Schaal et al, 1996: 1329). Higher testosterone is, of course, associated with better social well-being whereas lower levels of testosterone was associated with more violence which may be due to social isolation, stable anxiety throughout childhood or both influences influencing the aggressive behavior along with the low testosterone. High testosterone levels in adolescence is not related to antisocial disorder, but it is related to social dominance—as I have stated countless times.
Now for some comments.
I like this study. How they used behavioral questionnaires to assess children’s behavior, as well as using teacher and peer ratings to assess behavior is a great method. It’s even better that they followed them throughout their adolescent life to see any patterns that arose. I hope that there is more data on these children (I will look for any other data on this one point) because this could elucidate so many testosterone relationships with behavior. This study shows that higher testosterone levels were not associated with antisocial behavior but it was, however, associated with greater social well-being.
Clearly, we need higher-ish levels of testosterone—especially since lower levels are correlated with numerous maladies. So, it seems, that the testosterone=crime aggression myth (and in my opinion, crime) is busted. Testosterone is an extremely important hormone for proper endocrine functioning and having levels that are too low. They also state at the end of the paper that “From the perspective of the present study, it can be hypothesized that in a sample of normal adolescent males, those who dominate will tend to react to provocation and to show relatively low frustration tolerance. The results from the Olweus and colleagues’ study might then be interpreted as showing the relationship between dominance and T, rather than between antisocial behavior and T.”
So, again, testosterone is related to dominance; if you have lower testosterone you will be less socially dominant while if you have more you will be more socially dominant (this holds for adults as well). It’s interesting to note that boys perceived as dominant when around people they did not know had higher levels of testosterone than boys who were not. This shows that boys with lower levels of testosterone were seen as less popular, they were failing in school and they had higher levels of aggression. Testosterone levels in children are associated with social success and popularity, rather than antisocial behavior and violent outbursts as other studies have reported.
Testosterone=the dominance hormone—more specifically, the social dominance hormone. It does not lead to social maladjustment.
Musings on Testosterone and Race
1500 words
People don’t understand the relationship between testosterone, aggression, and crime. People hear the sensational media stating that testosterone causes crime, aggression, and anger. However, I have written numerous articles on this blog on the true nature of testosterone, what it’s really needed for and why we need it in high amounts. I’ve mused a lot on this hormone, which is one of my favorites to discuss due to the numerous misconceptions surrounding it.
Which way does causation run in regard to prisoners and their testosterone level?: heightened testosterone > aggression > violence or aggression > heightened testosterone > dominance > possibility (not necessarily, as I have written myself in the past) of violence.
People may use animal studies in support of their contention that testosterone causes aggressive behavior. However, for reasons I have discussed in the past, animal models only show avenues for future research and do not necessarily mean that this is the case for humans (as Mazur, 2006 point out). I don’t use animal studies. They’re good for future research, but to use them as evidence for causation in humans doesn’t make sense.
People may cite Dabbs et al showing that the more violent prisoners had higher levels of testosterone and therefore conclude that higher levels of testosterone drive the violent crime that they commit, however it is much more nuanced than that.
Does being a violent criminal raise testosterone or are violent people more likely to have high testosterone? Dabbs never untangles this; they just showed a correlation, which is small as evidenced by my other citations.
Testosterone is, as evidenced by numerous studies, related to dominance and dominance contests, however, during these dominance contests “a killing is rarely the outcome of a violent dominance contest” (Mazur, 2006: 25). Therefore, this throws a wrench in the testosterone-causes-crime hypothesis.
Some individuals may state that these dominance contests then lead to violence, however, as Mazur (2006) puts it: “Heightened testosterone is not a direct cause of male violence.”
Other animals assert dominance violently but we, necessarily, do not.
Mazur (2006) states that dominance contests rarely escalate to murder. Mazur also states that dominance contests also lead to increased T for the winners and decreased T for the losers, and these contests also don’t necessarily lead to murder/violent behavior. There is a feedback loop with high T causing behavior and behavior causing high T (Mazur, 2006) while this feedback loop may lead to “lethal effects” (Mazur and Booth, 2008).
It’s worth noting that Mazur seems to advocate for ‘testosterone-depressing drugs’. He concludes:
There are strong linkages between macro-level culture and the physiology of
individuals. We may find solutions to some of our social problems by altering these linkage.
Macro-level culture being white culture, black culture, Asian culture, etc.
The physiological differences are due to the preparation for dominance contests. So, his hypothesis goes, the culture of dominance among young black men with no education is why their T is so high. That low education was also associated with low education lends credence to the claim that this is changeable.
However, in his newer article on education, low testosterone and blacks he advocates for more sensible solutions (attempting an environmental change). I don’t know about you but I have big problems with using FDA/Big Pharma drugs to ‘reduce societal problems’, and it seems that Mazur has changed his view there. Mazur (2016) writes:
If high T does facilitate the high violence rate among young black men, there would be a troubling policy question of what, if anything, to do about it. Any notion of a medical or pharmaceutical fix, rather like prescribing Ritalin for hyperactivity, would reek of race-based chemical castration and should be regarded as outside the pale. However, social interventions might be workable and ethically acceptable.
I have railed against measures like this in the past, since proposing measures to attempt to ‘decrease crime through supposedly decreasing one of the main “causes”‘ is very Brave New World-ish, and I am highly against those measures. Social interventions are, in my view, the more sensible measures to undertake.
In regard to low education and testosterone, this same relationship was noticed by Assari, Caldwell, and Zimmerman (2014) where they note that testosterone was not associated with aggression in men, but low education was, which Mazur (2016) replicates, showing that blacks of the same age group with more education had lower levels of testosterone when compared to age-matched blacks. Mazur (2016) cites one study in support for his contention that education can decrease aggressive behavior (Carre et al, 2014)
The correlation is there, I agree. let’s take the middle value of .11 between Archer, Graham-Kevan, and Davies, 2005 at .08; and Book, Starzyk, and Quinsey, 2001 .14. So testosterone explains 3 percent of the the relationship with aggression. Not high at all.
Great evidence against the testosterone-causes-aggressive-behavior hypothesis are data on the Yanomami. About 50 percent of Yanomami men meet their deaths by other Yanomami men. So the Yanomami must have testosterone levels through the roof, right? Wrong. De Lima et al (2015) write:
We observed that Yanomamis present lower levels of testosterone (414 ng/dL) in relation to other ethnic groups (502/512 ng/dL), but still within normal limits (350-1000 ng/dL).
(Note that these values for “normal limits” changed, going into effect at the end of July.)
The Yanomami with an extremely high murder rate with nowhere near a modern society have T levels on the lower end of our range. So….. The Yanomami example is direct evidence against the assertion of testosterone directly causing crime, as some people assert (it is even evidence against an indirect cause). The evidence of the Yanomami having testosterone levels near our lower range is direct evidence against the testosterone/crime hypothesis. Clearly, other variables drive the high violence rate in this society that are not testosterone. More interestingly, these people have had little contact with Western societies, and their T levels are still low compared to ours despite constantly being vigilant for threats from other Yanomami.
Most dominance contests do not end violently in the first-world, there is numerous evidence to attest to this fact. So with the low correlation between testosterone and aggression (Book, Starzyk, and Quinsey, 2001; Archer, Graham-Kevan and Davies, 2005; Book and Quinsey, 2005), along with dominance contests rarely ending in murder/violent crime, then there are way more factors influencing these phenomena.
So the feedback loop goes: Testosterone rises in expectation of a challenge which then, after the dominance contest (which doesn’t always necessarily lead to violence), it affects both individuals differently depending on whether or not they won or lost that dominance contest and these values then persist over time if the dominance contests continuously end up the same.
Let’s say, for argument’s sake, that testosterone is a large cause for aggressive behavior in lower-educated blacks, what should be done about it? Mazur cites evidence that behavioral interventions seem to work to decrease violent behavior during certain circumstances (Carre et al, 2014), so that is a good way to lower violence in populations that have low education.
So heightened testosterone does lead to dominance which then facilitates a dominance contest between two individuals which does not necessarily lead to crime and aggressive, violent behavior (this outcome is rare in dominance contests among “higher primates” [Mazur’s words]) so, therefore, while testosterone does facilitate dominance contests, it rarely leads to violence in our species. Therefore, testosterone does not cause aggressive behavior and crime, but it does cause dominance which, for the most part, do not always result in violent, aggressive, murderous behavior.
I’ve shown that Mazur replicated other analyses that show that testosterone and aggressive behavior are related to lower education. Testosterone wasn’t associated with aggressive behavior in Assari, Caldwell, and Zimmerman’s (2014) study, and, as Mazur (2016) replicates, low education was. So one way to end this relationship is to educate people, as shown by Carre et al (2014), and with this education, crime will begin to fall. Heightened testosterone is not a direct cause of male violence.
(Note: I also believe that other factors such as sleep and depressed nutrition play a factor in crime, as well as racial differences in it. See Birch, 1972; Liu et al, 2003; Liu et al, 2004; Walker et al, 2007; Galler et al, 2011, 2012a, 2012b; Spratt et al, 2012; Gesch, 2013; Kuratko et al, 2013; Waber et al, 2014; Raine et al, 2015; Thompson et al, 2017 for more information, I will cover this in the future. I’m of course not daft enough to believe that no genetic differences between individuals/populations are the cause of a lot of crime between them, however, as I have laid out the case in regard to testosterone and MAOA numerous times, these two explanations for both individual differences in crime as well as racial differences in crime leave a lot to be desired. Other genetic factors, of course, influence these differences, however, I am only worried about refuting the popular notions of ‘testosterone and MAOA, the ‘warrior gene” cause crime. The relationship is a lot more nuanced as I have provided mountains of evidence for.)
Comments on Batrinos (2012)
850 words
Batrinos (2012) is a paper titled Testosterone and Aggressive Behavior in Man. Section 2 reviews studies on prisoners. Small ns, no controls, prison isn’t a natural environment. Similar data was reported, but it has the same problems as above. Studies of sexual offenders are contradictory, with blood T not being associated, then being associated with aggression. An investigation of veterans showed T to raise aggression, again, non-representative sample (and you also have to think of PTSD and other illnesses). Then the money quote:
It is of interest, however, that supraphysiological doses of testosterone in the order of 200 mg weekly (20), or even 600 mg weekly (21), which were administered to normal men had no effect on their aggression or anger levels.
Dominance is related to testosterone increases, and dominance can lead to aggressive behavior then to violent acts, but this is not always true. Mazur and Booth’s (1997) reciprocal model shows a feedback loop on dominance and testosterone:
Ehrenkranz et al. (1974) showed that socially dominant but unaggressive prisoners had relatively high T, not significantly different from the T levels of aggressive prisoners (who may have been dominant too). Nearly all primate studies that have been interpreted as linking T to aggression (Dixson 1980) may as easily be interpreted as liking T with dominance (Mazur 1976). Recent reviewers have questioned whether, among humans, T is related to aggressiveness per se (Archer 1991; Albert et al. 1994).
Testosterone and Dominance in Men
And:
Heightened testosterone is not a direct cause of male violence.
The Role of Testosterone in Male Dominance Contests that Turn Violent
(Much more on this paper soon.)
Small studies have shown that T increases during sports (duh because it’s competition) and that watching your favorite sports team win a game increases T (has been replicated).
Batrinos then cites a study talking about circadian rhythm and testosterone/aggression here:
Interestingly enough, Brown et al (2007) lends credence to my hypothesis that I have discussed in the past. Testosterone decreases at 8pm (most crime is comitted at 10 pm for adults) with increases in testosterone at night:
It is not surprising that T levels at 2000 h and 0800 h would be important for TS-IAB relationships because the 2000 h and 0800 h values represent the evening nadir and morning zenith.
But most crime is not comitted in the morning, for both adults and children (as seen below):
Now, anyone who has read my article on why testosterone doesn’t cause crime knows where I’m going with this:
Look at the times most crimes are committed then think about when T levels are highest (8 am).


Why Testosterone Does Not Cause Crime
This does seem to show a relationship with when most crimes are comitted, a sharp decrease in crime occurs as testosterone levels hit their highest in the day, which is evidence against the testosterone-causes-crime hypothesis.
In regard to the crime chart from the OJJDP, they write:
- In general, the number of violent crimes committed by adults increases hourly from 6 a.m. through the afternoon and evening hours, peaks at 10 p.m., and then drops to a low point at 6 a.m. In contrast, violent crimes by juveniles peak in the afternoon between 3 p.m. and 4 p.m., the hour at the end of the school day.
Using what I gave, what do you draw from the line graph? It’s clear that, since testosterone is highest in the morning aand at its lowest at 8 pm (when most violent crime is being comitted) that testosterone is not directly related to crime, since, as evidenced by Brown et al (2007), testosterone levels are lowest at 8 am with a sharp rise as the night/ morning progresses. Looking at their testosterone chart for the hours between 8 pm and 8 am, testosterone did increase at 8pm and into the night. However, as testosterone levels continued increasing into the night, crime does not linearly increase with the rise in testosterone (see fig. 3 in Brown et al, 2007).
Now, finally, in regards to the claim about “locally produced testosterone”, it is true that the brain can produce testosterone de novo from cholesterol; but wait! Luteinizing hormone signaling in the neurons promotes the secretion and production of steroids (Liu et al, 2007), along with the de novo production of testosterone through cholesterol (literally the only way testosterone can be produced).
Good paper, shaky claims (the prisoner claims suck, circadian rhythm claims suck). The only novel thing in this paper is saying how the brain can produce testosterone de novo from cholesterol (though luteinizing hormones are still involved, see above cite). Injecting a man with supraphysiologic doses of testosterone does not increase aggression nor anger levels. It’s definitive that testosterone does not directly cause crime, as evidenced by the low correlation between testosterone and aggression. Numerous other studies (which I have reviewed in the past), however, show that aggression precedes the testosterone increase which is only seen in certain social situations. These specific situations, by themselves, drive the production of the hormone.
Sex Differences in Aggressive Behavior and Testosterone
1700 words
Many long-time readers may know of the numerous tirades of been on in regards to the “testosterone causes crime and aggression” myth. It’s a fun subject to talk about because the intelligent human physiological system is an amazing system. However, people who are not privy to the literature on testosterone in regards to race, aggression, crime, sex differences etc are only aware of whatever they read in pop science articles. So since they never read the actual papers themselves, they get a clouded view of a subject.
In my last article, I wrote about how there are no “testosterone genes”. In previous articles on the hormone, I have proven that there is no causal link between testosterone and aggression. But when comparing the sexes, how do the results look? Do they look the same with men being more violent while women—who have substantially less testosterone than men—do not have any higher levels of aggression or crime? The most recent study I’m aware of is by Assari, Caldwell, and Zimmerman (2014) titled: Sex Differences in the Association Between Testosterone and Violent Behaviors.
To make a long story short, there was no relationship between testosterone and aggression in men, but a significant relationship between testosterone and aggression in women. This data comes from the Flint Adolescent Study, a longitudinal study conducted between the years of 1994 to 2012. In regards to testosterone collection, saliva was used which has a perfect correlation with circulating testosterone. The eligibility to be included in the testosterone assay was “provided consent for the procedure, not being pregnant, not having anything to eat, drinking nothing except water, and not using tobacco, 1 hour prior to collection” (Assari, Caldwell, and Zimmerman, 2014).
The adolescent who contributed saliva gave a whole slew of demographic factors including SES, demographics, psychological factors, family relations, religion, social relations, behavior, and health. They were aged 14 to 17 years of age. They collected data during face-to-face interviews,
Age and SES were used as control variables in their multivariate analysis. For violent behaviors, the authors write:
Youths were asked how often they had engaged in the following behaviors; ‘had a fight in school’, ‘taken part in a rumble where a group of your friends were against another group’, injured someone badly enough to need bandages or a doctor’, ‘hit a teacher or supervisor at work (work supervisor)’, used a knife or gun or other object (like a club) to get something romantic a person’, ‘carried a knife or razor’, or ‘carried a gun’. All items used a Likert response, ranging from 1 (0 times) to 5 (4 or more times). Responses to each item were averages to calculate the behavior during the last year. Total score was calculated as the average of all items. Higher scores indicated more violent behaviors (a = 0.79). This measure has shown high reliability and validity and it has been used previously in several published reports.
This is a great questionnaire. The only thing I can think of that’s missing is fighting/arguing with parents.
In regards to testosterone assaying, they were assayed after 11 am to “control for changes due to diurnal rhythm” (Assari, Caldwell, and Zimmerman, 2014). I’m iffy on that since testosterone levels are highest at 8 am but whatever. This analysis is robust. Saliva was not taken if the subject had smoked or ingested something other than water or if a subject was pregnant. Assays should be taken as close to 8 am, as that’s when levels are highest. However one study does argue to extend the range to 8 am to 2 pm (Crawford et al, 2015) while other studies show that this only should be the case for older males (Long, Nguyen, and Stevermer, 2015). Even then assays were done at the higher end of the range as stated by Crawford et al (2015), so differences shouldn’t be too much.
86.4 percent of the sample was black whereas 13.4 percent were white. 41.2 percent of the subjects had some college education whereas 58.2 percent of the subjects lived with a partner or relative. 21.4 percent of the subjects were unemployed.
The mean age was 20.5 for both men and women, however, which will be a surprise to some, testosterone did not predict aggressive behavior in men but did in women. Testosterone and aggressive behavior were positively correlated, whereas there was a negative correlation between education and testosterone and aggressive behavior. Though education was associated with aggressive behavior in men but not women. So sex and education was associated with aggressive behavior (the sex link being women more privy to aggressive behavior while men are more privy to aggressive behavior due to lack of education). Females who had high levels of education had lower levels of aggressive behavior. Again: testosterone wasn’t associated with violent behavior in men, but it was in women. This is a very important point to note.
This was a community sample, so, of course, there were different results when compared to a laboratory setting, which is not surprising. Laboratory settings are obviously unnatural settings whereas the environment you live in every day obviously is more realistic.
This study does contradict others, in that it shows that there is no association between testosterone and aggression in men. However, still other research shows that testosterone is not linked to aggression or impulsivity, but to sensation-seeking, sexual experience or sociality (Daitzman and Zuckerman, 1980; Zuckerman, 1984). Clearly, testosterone is a beneficial hormone and due to the low correlation of testosterone with aggression (between .08 and .14; Book, Starzyk, and Quinsey, 2001; Archer, Graham-Kevan and Davies, 2005; Book and Quinsey, 2005). This paper, yet again, buttresses my arguments in regards to testosterone and aggressive behavior.
In regards to the contrast in the literature the authors describe, they write:
One of the many factors that may explain the inconsistency in these findings is the community versus clinical setting, which has been shown to be a determinant of these associations. Literature has previously shown that many of the findings that can be found in clinical samples may not be easily replicated in a community setting (36).
This is like the (in)famous, unreplicable stereotype threat (see Stroessner and Good). It can only be replicated in a lab, not in an actual educational setting. And it also seems that this is the case for testosterone and aggressive behavior.
Just because women have lower testosterone and are less likely to engage in aggressive behavior, that doesn’t mean that a relationship does not exist between females. “It is also plausible to attribute sex differences in the above studies to differential variations in the amount of testosterone among men and women” (Assari, Caldwell, and Zimmerman, 2014). This view supports the case that testosterone is linked to aggression in females, even though their range of testosterone is significantly lower than men’s, while it may also be easier to assay women for testosterone due to less diurnal variation in comparison to men (Book, Starzyk, and Quinsey, 2001).
Assari, Caldwell, and Zimmerman, (2014) also write (which, again, buttresses my arguments):
Age may explain some of the conflicting results across the studies. A meta-analysis of community and selected samples suggested that there might be only low to modest association between testosterone and aggression, with mean weighted correlations ranging from 0.08 to 0.14, in males. Overall, these meta-analyses suggest that the testosterone-aggression association is equally strong in 12 to 21-year-olds, as it is in 22 to 35-year-olds, but that it may be less strong in age groups younger than 12, than in those who are older.
So, testosterone may be associated with aggressive behavior and violence in women but not in men. In men, the significant moderator was education. It’s interesting to note that Mazur (2016) noted that young black males with little education had higher levels of testosterone than age-matched samples of other blacks. This, along with the evidence provided here, may be a clue that if the social environment changes, then so will higher levels of testosterone (as I have argued here).
They, perhaps taking too large of a leap here, argue that “aggressive behaviors may be more social and less biologically based among men” (Assari, Caldwell, and Zimmerman, 2014). Obviously social factors are easier to change than biological ones (in theory), so, they argue, preventative measures may be easier for men than women. More studies need to be done on the complex interactions between sex, testosterone, aggression, biology and the social environment which then shapes the aggressive behaviors of those who live there.
Testosterone and aggression studies are interesting. However, you must know a good amount of the literature to be able to ascertain good studies from the bad, what researchers should and should not have controlled for, time of assay, etc because these variables (some not in the author’s hands, however) can and do lead to false readings if certain variables are not controlled for. All in all, the literature is clearly points to, though other studies contest this at times, the fact that testosterone does not cause aggressive behavior in men. The myth needs to die; the data is piling up for this point of view and those who believe that testosterone causes aggressive behavior and crime (which I have shown it does not, at least for men) will soon be left in the dust as we get a better understanding of this pivotal hormone.
(In case anyone was going to use this as evidence that black women have higher levels of testosterone than white women, don’t do it because it’s not true. You’ll only embarrass yourself like this guy did. Read the comments and see him say that you don’t need scientific measurements, you only need to ‘observe it’ and through ‘observation’ we can deduce that black women have higher levels of testosterone than white women. This is not true. Quoting Mazur, 2016:
The pattern [high testosterone] is not seen among teenage boys or among females.
…
There is no indication of inordinately high T among young black women with low education.
Whoever still pushes that myth is an idealogue; I have retracted my article ‘Black Women and Testosterone‘, but idealogues just gloss over it and read what they think will bolster their views when I have provided the evidence to the contrary. It pisses me off that people selectively read things then cite my article because they think it will confirm their pre-conceived notions. Well too bad, things don’t work like that.)
