
Home » Posts tagged 'Race'
Tag Archives: Race
People Shouldn’t Be Proud of Things They Don’t Accomplish
1500 words
Pride in one’s group seems like it should be valid to have. After all, some (all?) groups have been through some heinous things, and so most may think that it follows that they should be proud of their group, like their race or ethnicity. Take, for example, white pride, Asian price, black pride, Jewish pride, etc. None of them make any sense. They don’t make sense because they aren’t actual accomplishments. They are, in fact, proud of a lucky accident. Something they they themselves don’t choose at all. I will construct an argument I call “the agency-based price argument” to show that it’s nonsense to be proud of something you didn’t accomplish. While it may seem understandable that people may feel pride in their group, I will try to argue that this doesn’t make sense because ones race/ethnicity isn’t an accomplishment.
The argument
(P1) You should only be proud of things you accomplish.
(P2) Your race/ethnicity isn’t an accomplishment.
(C) So you shouldn’t be proud of your race/ethnicity.
The argument is clearly valid (modus ponens), but is it sound? I think so. And while I think there some ways a proponent of racial/ethnic pride (REP) could try to attack the argument, I will focus on the defense of the first premise.
P1 asserts that pride should normatively be tied to personal agency and effort, implying that only accomplishments—outcomes of deliberate actions—justify pride. Quite clearly, this is a merit-based definition of pride. While of course, people do feel pride in their group, that doesn’t make it a rational thing to hold pride about. Some defenses that could possibly be made against premise one fall short.
P1 makes a normative claim about what pride should be. P2 needs no defending since it’s a straightforward observation—one’s race or ethnicity isn’t something they themselves choose or accomplished, it’s just a lucky accident of birth. Since the argument is valid, if all of the premises are true then the argument is sound. Thus, the REP proponent would need to reject P1.
The way I am conceptualizing and defining “pride” in this argument is simple—a positive emotion of self-satisfaction or esteem derived from one’s personal accomplishments, which are outcomes derived from deliberate effort, agency, or responsibility. Thus, pride is warranted if and only if it is tied to actions or results and individual has actively contributed to excluding unchosen circumstances. Pride should be rooted in accomplishments that reflect an individual’s effort, skills, or intentional actions. One must have played a direct causal role in the outcome through their choices or actions, which ensures that pride is earned rather than passively received. So pride is a rational response to merit based on what one does not what one is. This is an agency-based definition of pride, so what matters here are outcomes. Pride requires agency and being born a certain way isn’t agentic.
Clearly, the definition of “pride” I am operating under is an agency-merit one. One may say “That’s just your definition, RR, so what?” But if I can be proud of anything that’s outside of my control, what power does the word even have? My definition preserves the only reason why pride was considered admirable.
So if the word “pride” is removed here when discussing race/ethnicity, which positive emotion are they referring to when they express feeling about their unchosen group status? They are referring to gratitude, belonging/solidarity, happiness, relief at no longer feeling shame, appreciation for their ancestors who have suffered. But none of those emotions requires using the pride concept.
When someone says they are proud of their race, what about other things outside of their control? Would it make sense to say they are proud of any other things that were outside of their control? Why should one feel pride in an unchosen trait?
Defending racial pride means one of two things: (A) membership in their group is itself an achievement or (B) their group is superior and they deserve credit for belonging to it. Let’s say tomorrow it was found out that race X’s achievements had actually been found to have had a worse track record than one who is proud of their race—fewer inventions, more atrocities, less resilience—would they feel less proud? If the answer is “yes” then the individual has admitted that their pride is just basically tribal score keeping, not personal achievement. If the answer is “no”, then pride has nothing to do with the group’s merits at all, so why should the group be brought into it?
The linguistic habit of calling unchosen traits a source of pride is not a harmless linguistic quirk; it is a surrender of the individual to the collective. Pride is one of the only positive emotions that signals personal responsibility. Allowing pride to be triggered by unchosen traits strips the word of its meaning. It no longer means “I earned this”, it then collapses to mean “I like having this.”
Certain other actions and choices people make are not sources of such pride. We treat other actions with shame at worst or neutrality at best. So if pride and shame are to remain to coherent moral emotions, they must track the boundary of agency. Things outside of my control can neither be pride-making or shame-making. Thus, extending pride to unchosen positives while rejecting it for unchosen negatives is arbitrary special pleading.
Pride refers to “I did”, not “I am.” We have words for appreciating unchosen things, like gratitude, contentment, joy, celebration. So why does it make sense to take the one word that uniquely means “I earned this”, and apply it to luck? Therefore, pride must remain tethered to merit. Being proud of your race is like feeling guilty for the weather, feeling jealous of your own possessions.
This is why pride without accomplishment doesn’t make any sense and is a logical contradiction; if asserts a causal responsibility that just does not exist. The emotion doesn’t make any sense when used for things an individual did not accomplish. The REP proponent—no matter if white, black, Asian, or any ethny—has no where to go. This argument holds for any racio-ethnic pride. Although some have argued that racial pride is a valid emotion in response to historical injustices, arguing that it fosters solidarity and resilience, but Frederick Douglas stated that the whole concept of racial pride is ridiculous (Fischer, 2021), and I of course would be included to agree with him.
Why would or should one be proud of their ancestor’s achievements? Would they be proud of the accomplishments of an individual of their same race that is alive at the same time as them? Or is it only for, say, the Roman Empire or the British Empire. Or the Revolutionary War? If one has ancestors from those time periods who were at those events and played pivotal roles in the events, should they be proud of it? No, they shouldn’t. Because there is no causal agency on their part. There is just a causal disconnect. The link between one’s ancestors and racial compatriots is their DNA and phenotype. If your great-great-grandfather signed the Declaration of Independence, should you feel proud? Should the descendants of Benedict Arnold feel shame? Should the grandchildren of SS officers hang their heads in shame? Should Italians feel pride for Caesar but disavow responsibility for Mussolini?
Having pride in historical achievements is like a cafeteria buffet, you’re only choosing what you want and ignoring everything else. Taking pride in the good but ignoring the bad. It’s like a British nationalist taking pride in their great British Empire that they were born into through nothing but luck but disowning slavery, genocide, and conquest. They’re just picking and choosing what to “feel pride for” when it comes to their ancestor’s achievements.
A parent who raises a child can feel proud of their children’s accomplishments because they had a hand in their own development. If a parent helped to create the conditions in which their child’s accomplishments occurred, then the parent is justified in taking pride in their children’s accomplishments, since they have the agency that necessitates such pride. This would be second-order pride, if the parent can point to specific costly actions they took, they grant the child primary ownership of the achievement, and they would have accepted proportionate responsibility had their child failed. Therefore, under these conditions a parent has a right to say that they are proud of their children and the accomplishments they made, since they had a direct hand in it.
Conclusion
Clearly, the argument I’ve mounted here shows that agency is intertwined with pride. Pride belongs only to the hands that did something and the mind that made the choices, not the blood and the luck that made one have the same race or be born in the same place as individuals from the past. One can admire their ancestor’s achievements and study them, but feeling pride for them? That’s obviously ridiculous.
As I’ve shown, pride is the emotion for personal accomplishments. Nothing else qualifies, not genes, skin color, ancestors, empires, co-racial strangers, and no child who’s success is on their own. The only exception is the parent who’s years of deliberate shaping helped the child become successful, but the pride must remain modest and cede the bulk to the child. But everything outside of this is gratitude, admiration, joy, or solidarity.
Racial Differences in Amputation
1850 words
Overview
An amputation is a preventative measure. It is done for a few reasons: To stop the spread of a gangrenous infection and to save more of a limb after there is no blood flow to the limb after a period of time. Other reasons are due to trauma and diabetes. Trauma, infection, and diabetes are leading causes of amputation in developing countries whereas in developed countries it is peripheral vascular disease (Sarvestani and Azam, 2013). Poor circulation to an affected limb leads to tissue death—when the tissue begins turning black, it means that there is no or low blood flow to the tissue, and to save more of the limb, the limb is amputated just above where the infection is. About 1.8 million Americans are living as amputees. After amputation, there is a phenomenon called “phantom limb” where amputees can “feel” their limb they previously had, and even feel pain to it, and it is very common in amputees; about 60-80 percent of amputees report “feeling” a phantom limb (see Collins et al, 2018; Kaur and Guan, 2018). The sensation can occur either immediately after amputation or years after. Phantom limb pain is neuropathic pain—a pain that is caused by damage to the somatosensory system (Subedi and Grossberg, 2011). Amputees even have shorter lifespans. When foot-amputation is performed due to uncontrolled diabetes, mortality ranges between 13-40 percent for year one, 35-65 percent for year 3, and 39-85 percent in year 5 (Beyaz, Guller, and Bagir, 2017).
Race and amputation
Amputation of the lower extremities are the most common amputations (Molina and Faulk, 2020). Minority populations are less likely to receive preventative care, such as preventative vascular screenings and care, which leads to them being more likely to undergo amputations. Such populations are more likely to suffer from disease of the lower extremities, and it is due to this that minorities undergo amputations more often than whites in America. Minorities in America—i.e., blacks and “Hispanics”—are about twice as likely as whites to undergo lower-extremity amputation (Rucker-Whitaker, Feinglass, and Pearce, 2003; Lowe and Tariman, 2008; Lefebvre and Lavery, 2011; Mustapha et al, 2017; Arya et al, 2018)—so it is an epidemic for black America. Blacks are even more likely to undergo repeat amputation (Rucker-Whitaker, Feinglass, and Pearce, 2003). In fact, here is a great essay chronicling the stories of some double-amputee black patients.
Why do blacks undergo amputations more often than whites? One answer is, of course: Physician bias. For example, after controlling for demographic, clinical, and chronic disease status, blacks were 1.7 times more likely than whites to undergo lower-leg amputations (Feinglass et al, 2005; Regenbogen et al, 2007; Lefebvre and Lavery, 2011). What is a cause of this is inequity in healthcare—note that “inequity” here means differences in care that are avoidable and unjust (Sudana and Blas, 2013).
Another reason is due to complications from diabetes. Blacks have higher rates of diabetes than whites (Rodriguez and Campbell, 2007) but see Signorello et al (2007). Muscle fiber differences between races (see also here). Differences in hours-slept between blacks and whites, too, could also explain the severity of the disease. But what could also be driving differences in diabetes between races is the fact that blacks are more likely than whites to live in “food swamps.” Food swamps are where it is hard to find nutritionally-dense food, whereas food deserts are areas where there is little access to healthy, nutritious food. In fact, a neighborhood being a food swamp is more predictive of obesity status of the population in the area than is its being a food desert (Cooksey-Stowers, Schwartz, and Brownell, 2017). Along with the slew of advertisements in that are directed to low-income neighborhoods (see Cassady, Liaw, and Miller, 2015), we can now see how such things like food swamps contribute to high hospitalization rates in low-income neighborhoods (Phillips and Rogriguez, 2019). These amputations are preventable—and so, we can say that there is a lack of equity in healthcare between races which leads to these different rates in amputation—before even thinking about physician bias. Amputation rates for blacks in the southeast can be almost seven times higher than other regions (Goodney et al, 2014).
Stapleton et al (2018: 644) conclude in their study on physician bias and amputation:
Our study demonstrates that such justifications may be unevenly applied across race, suggesting an underlying bias. This may reflect a form of racial paternalism, the general societal perception that minorities are less capable of “taking care of themselves,” even including issues related to health and disease management.23 Underlying bias may prompt more providers to consider amputation for minority patients. Furthermore, unlike in transplant surgery, there is currently no formal process for assessing patient compliance with treatment protocols or self-care in vascular surgery.24 Asking providers to make snap judgments about patient compliance, without a protocol for objective assessment, allows subconscious bias to influence patient care.
Physician bias is pervasive (Hoberman, 2012)—whether it is conscious or unconscious racial bias. Such biases can and do lead to outcomes that should not occur. By attempting to reduce disparities in healthcare that then lead to negative outcomes, we can then attempt to improve the quality of healthcare given to lower-income groups, like blacks. Such biases lead to negative health outcomes for blacks (such as the claim that blacks feel less pain than whites), and if they were addressed and conquered, then we could increase equity between groups until access to healthcare is equal—and physician bias is an impediment to access to equal healthcare due to the a priori biases that physicians may hold about certain racial/ethnic groups. Medical racism, therefore, drives a lot of the amputation differences between blacks and whites. Hospitals that are better equipped to offer revascularization services (attempting to save the limb by increasing blood flow to the affected limb) even had a higher rate of amputations in blacks when compared to whites (Durazzo, Frencher, and Gusberg, 2013).
For example. Mustapha et al (2017) write:
Compared to Caucasian patients, several studies have found that African-Americans with PAD are more likely to be amputated and less likely to have their lower limb revascularized either surgically or via an endovascular approach [3–9]. In an early analysis of data from acute-care hospitals in Florida, Huber et al. reported that the incidence of amputation (5.0 vs. 2.5 per 10,000) was higher and revascularization lower (4.0 vs. 7.1 per 10,000) among African-Americans compared to Caucasians, even though the incidence of any procedure for PAD was comparable (9.0 vs. 9.6 per 10,000) [4]. Other studies have reported that the probability of undergoing a revascularization or angioplasty was reduced by 28–49 % among African-Americans relative to Caucasians [3 6]
Pro-white unconscious biases were also found among physicians, as Kandi and Tan (2020) note:
There is evidence of both healthcare provider racism and unconscious racial biases. Green et al. found significant pro-White bias among internal medicine and emergency medicine residents, while James SA supported this finding, indicating a “pro-white” unconscious bias in physician’s attitudes towards, and interactions with, patients [43,44]. In a survey assessing implicit and explicit racial bias by Emergency Department (ED) providers in care of NA children, it was discovered that many ED providers had an implicit preference for white children compared to those who identified as NA [45]. Indeed, racism and stigmatization are identified as being many American Indians’ experiences in healthcare.
One major cause of the disparity is that blacks are not offered revascularization services at the same rate as whites. Holman et al (2011: 425) write:
Finally, given that patients’ decisions are necessarily confined to the options offered by their physicians, racial differences in limb salvage care might be attributable to differences in physician decision making. There are some data to suggest lower vein graft patency rates in black patients compared to whites.18,19 A patient’s race, therefore, may influence a vascular surgeon’s judgment about the efficacy of revascularization in preventing or delaying amputation. Similarly, a higher proportion of black patients in our sample were of low SES, which correlates with tobacco use,20-22 and we know that continued tobacco use increases the risk of lower extremity graft failure approximately three-fold.23 It is possible that a higher proportion of black patients in our sample were smokers who refused to quit, in which case vascular surgeons would be much less likely to offer them the option of revascularization. While Medicare data include an ICD-9 diagnosis code for tobacco use, the prevalence in our study sample was approximately 2%, suggesting that this code was grossly unreliable as a means of directly measuring and adjusting for tobacco use.
Smoking, of course, could be a reason why revascularization would not be offered to black patients. Though, as I have noted, smoking ads are more likely to be found in lower-income neighborhoods which increases the prevalence of smokers in the community.
With this, I am reminded of two stories I have seen on television programs (I watch Discovery Health a lot—so much so that I have seen most of the programs they show).
In Untold Stories of the ER, a man came in with his hand cut off. He refused medical care. He would not let the doctors attempt to sew his hand back on. Upon the police entering his home to check for evidence (where his hand was found), they searched his computer. It seems that he had a paraphilia called “acrotomophilia” which is where one is sexually attracted to people with amputations. Although he wanted it to be done to himself—he had inflicted the wound on himself. After the doctor tried to reason with the man to have his hand sewed back on, the man would not let up. He did not want his hand sewed back on. I wonder if, years down the line, the man regretted his decision.
In another program (Mystery Diagnosis), a man had said that as a young boy, he had seen a single-legged war veteran amputee. He said that ever since then, he would do nothing but think about becoming an amputee. He lived his whole life thinking about it without doing anything about it. He then went to a psychiatrist and spoke of his desire to become an amputee. After some time, he eventually flew to Taiwan and got the surgery done. He, eventually, found happiness since he had done what he always wanted to.
While these stories are interesting they speak to something deep in the minds of the individuals who mutilate themselves or get surgery to otherwise healthy limbs.
Conclusion
Blacks are more likely than whites to receive amputations in affected limbs than whites and are less likely to receive treatments that may be able to save the affected limb (Holman et al, 2011; Hughes et al, 2013; Minc et al, 2017; Massada et al, 2018). Physician bias is a large driver of this. So, to better public health, we then must attempt to mitigate these biases that physicians have that lead to these kinds of disparities in healthcare. Medical and other kinds of racism have led to this disparity in amputations between blacks and whites. Thus, to attempt to mitigate this disparity, blacks must get the preventative care needed in order to save the affected limb and not immediately go for amputation. Thankfully, such disparities have been noticed and work is being done to decrease said disparities.
So race is a factor in the decision on whether or not to amputate a limb, and blacks are less likely to receive revascularization services.
Racism, Stress, and Physiology
1800 words
The term ‘racism’ has many definitions. What does it mean for a person to be a ‘racist’? What does it mean for a person to have ‘racist beliefs’? What does the term ‘racism’ refer to? The answers to these questions then will inform the next part—what does racism have to do with stress and physiology?
What is ‘racism’?
Racism has many definitions, so many—and so many for uses in different contexts—that it has been argued, for example by those in the far-right, that it is, therefore, a meaningless term. However, just because there are many definitions of the term, it does not then mean that there is no referent for the term we use. A referent is a thing that is signified. In this instance, what is the referent for racism? I will provide a few on-hand definitions and then discuss them.
In Part VI of The Oxford Handbook of Race and Philosophy (edited by Naomi Zack, 2016) titled Racisms and Neo-Racisms, Zack writes (pg 469; my emphasis):
Logically, it would seem as though ideas about race would have to precede racism. But the subject of racism is more broad and complicated than the subject of race, for at least these two historical reasons. First, the kind of prejudice (prejudged cognitions and negative emotions) and discrimination (treating people differently on the grounds of group identities) that constitute racism have a longer history than the modern idea of race, for instance in European anti-Semitism. And second, insofar as modern ideas of race have been in the service of the dominant interests in international and internal interactions, these ideas of race are ideologies that have devalued non-white groups. That is, ideas of race are themselves already inherently racist.
In philosophy, racism has been treated as attitudes and actions of individuals that affect nonwhites unjustly and social structures and institutions that advantage whites and disadvantage nonwhites. The first is hearts-and-minds or classic racism, for instance the use of stereotypes and harmful actions by whites against people of color, as well as negative feelings about them. The second is structural racism, for instance the use of stereotypes or institutional racism, for instance, the facts of how American blacks and Hispanics are, compared to whites, worse off on major measures of human well-being, such as education, income, family wealth, health, family stability, longevity, and rates of incarceration.
John Lovchik in his book Racism: Reality Built on a Myth (2018: 12) notes that “racism is a system of ranking human beings for the purpose of gaining and justifying an unequal distribution of political and economic power.” Note that using this definition, “hereditarianism” (the theory that individual differences between groups and individuals can be reduced to genes; I will give conceptual reasons why hereditarianism is false as what I hope is my final word on the debate) is a racist theory as it attempts to justify the current social hierarchy. (The reason why IQ tests were first brought to America and created by Binet and Simon; see The History and Construction of IQ Tests and The Frivolousness of the Hereditarian-Environmentalist IQ Debate: Gould, Binet, and the Utility of IQ Testing.) This is why hereditarianism saw its resurgence with Jensen’s infamous 1969 paper. Indeed, many prominent hereditarians have held racist beliefs, and were even eugenicists espousing eugenic ideas.
Headley (2000) notes a few definitions of racism—motivational, behavioral, and cognitive racism. Motivational racism is “the infliction of unequal consideration, motivated by the desire to dominate, based on race alone“; behavioral racism is “failure to give equal consideration, based on the fact of race alone”; and cognitive racism is “unequal consideration, out of a belief in the inferiority of another race.”
I have presented six definitions of racism—though there are many more. Now, for the purposes of this article, I will present my own: the ‘inferiorization’ of a racialized group which is then used to explain disparities in things like IQ test scores, social class/SES, education differences, personality, etc. Now, knowing what we know about physiological systems and how they react to the environment around them—the immediate environment and the social environment—how does this then relate to stress and physiology?
Racism, stress, and physiology
Now that we know what racism is, having had a rundown of certain definitions of ‘racism’, I will now discuss the physiological effects such stances could have on groups racialized as ‘races’ (note that I am using socialraces in this article; recall that social constructivists about race need to be realists about race).
The term ‘weathering’ refers to the body’s breaking down due to stress over time. Such stressors can come from one’s immediate environment (i.e., pollution, foodstuffs, etc) or their social environment (a demanding job, how one perceives themselves and how people react to them). So as the body experiences more and more stress it becomes more and more ‘weathered’ which then leads to heightened risk for disease in stressed individuals/populations.
Allostatic states “refer to altered and sustained activity levels of the primary mediators (e.g., glucocorticosteroids) that integrate energetic and associated behaviours in response to changing environments and challenges such as social interactions, weather, disease, predators and pollution” (McEwen, 2005). Examples of allostatic overload such as acceleration of atherosclerosis, hypertension (HTN), stroke, and abdominal obesity (McEwen, 2005) are more likely to be found in the group we racialize as ‘black’ in America—particularly women (Gillum, 1987; Gillum and Hyattsville, 1996; Barnes, Alexander, and Staggers, 1997; Worral et al, 2002; Kataoka et al, 2013).
Geronimus et al (2006) set to find out whether or not the heightened rate of stressors (e.g., racism, environmental pollution, etc) can explain why black bodies are more ‘weathered’ than white bodies. They found that such differences were not explained by poverty, indicating that it even affects well-off blacks. Allostatic load refers to heightened hormonal production in response to stressors. We know that physiological is homeodynamic and therefore changes based on the immediate environment and social environment (for example, when you feel like you’re about to get into a fight, your heart rate increases and you get ready to ‘fight or flight’).
Experiencing racism (environmental stimuli; real or imagined, the outcome is the same) is associated with increased blood pressure (HTN). So if one experiences racism they will them experience an increase in blood pressure, as BP is a physiological variable (Armstead et al, 1987; McNeilly et al, 1995; see Doleszar et al, 2018 for a review). The concept of weathering, then, shows that racial health disparities are, in fact, racist health disparities (Sullivan, 2015: 106). Racism, then, contributes to higher levels of allostasis and, along with it, higher levels of certain hormones associated with higher allostasis.
One way to measure biological age is by measuring the length of telomeres. Telomeres are found at the ends of chromosomes. Since telomere lengths shorten with age (Shammas, 2012), those with shorter telomeres are ‘biologically older’ than those of the same age with longer telomeres. Geronimus et al (2011) showed that black women had shorter telomeres than white women, which was due to subjective and objective stressors (i.e., racism). Black women in the age group 49-55 were 7.5 years ‘older’ than white women. Thus, they had an older physiological age compared to their chronological age. It is known that direct contact with discriminatory events is associated with poor health outcomes. Harrell, Hall, and Taliaferro (2003) note that:
“…physiological set points and the mechanisms governing them are not fixed. External stressors can permanently alter physiological functioning. Racism increases the volume of stress one experiences and may contribute directly to the physiological arousal that is a marker of stress-related diseases.
Social factors can, indeed, influence physiology and there is a wealth of information on how the social becomes biological and how environmental (social) factors influence physiological systems. Forrester et al (2019) replicated Geronimus’ findings, showing that blacks have a higher ‘biological age’ than whites and that psychosocial factors affect blacks more than whites. Simons et al (2020) also replicated Geronimus’ findings, showing that persistent exposure to racism was associated with higher rates of inflammation in blacks which then predicted higher rates of disease in blacks compared to whites. Such discrimination can help to explain differences in birth outcomes (e.g., Jasienska, 2009), stress, inflammation, obesity, stroke rates, etc in blacks compared to whites (Molnar, 2015).
But what is the mechanism by which higher allostatic load scores contribute to negative outcomes and shorter telomeres indicating a higher biological age? When one feels that they are being discriminated against, the sympathetic nervous system activates due to chronic stress and along with it HPA dysfunction. What this means is that there is a loss of the anti-inflammatory effects of cortisol—it becomes blunted. This then increases oxidative stress and inflammation. Thus, the inflammatory processes result in cardiovascular disease and immune and metabolic dysfunction. The HPA axis monitors and responds to stress—allostatic load. When stress hormones are released, the adrenal gland is targeted. When it receives a signal from the pituitary gland, it pumps epinephrine and norepinephrine into the body, causing our hearts to beat faster, causing us to breathe more deeply—what is known as ‘fight or flight.’ Cortisol is also released and is known as a stress hormone, but when the stressful event is over, all three hormones return to baseline. Thus, the higher amount of stress hormones in circulation indicates higher levels of allostatic load—higher levels of stress in the individual in question. We know that blacks have higher levels of allostatic load (i.e., stress-related hormones) than whites (Duru et al, 2012). Barr (2014: 71-72) writes:
Imagine, though, that before the allostatic load has a chance to return to its baseline level, another stressor is sensed by the hypothalamus. The allostatic load will once again increase to the plateau level. Should the perception of stressors be ongoing, the allostatic load will not have the chance to ever fully recharge, and the adrenal gland will be producing an ongoing stream of stress response hormones. The body will experience chronic elevation in its allostatic load. […] A person experiencing repeated stressors, without the opportunity for intervals that are relatively stress-free, will experience a chronically elevated allostatic load, with higher than normal levels of circulating stress response hormones.
Conclusion
What these studies show, then, is that race is a cause of health inequalities, but it’s not inherent in biology but due to social factors that influence the physiology of the individual in question. The term ‘racism’ has many referents, and using one of them identifies ‘hereditarianism’ as a racist ideology (it is inherently ideological). These overviews of studies show that racial health inequalities are due, in part, to perceived discrimination (racism) thus they are racist health disparities. We know that physiology is a dynamic system that can respond to what occurs in the immediate environment—even the social environment (Williams, 1992). Thus, what explains part of the health inequalities between races is perceived discrimination—racism—and how it affects the body’s physiological systems (HPA axis, HTN, etc) and telomeres.
My Citation Count
1200 words
I started this blog almost 5 years ago. Currently (excluding this one), there are 480 articles on this blog. Searching my blog name “notpoliticallycorrect.me” on Google Scholar leads to two citations—one on “IQ” and obesity and the other on inclusionism about race when it comes to medicine. These two cites pretty much perfectly show my views and their change in the past 5 years since the creation of this blog. I will discuss both papers that cited me in turn.
In the journal Social and Human Sciences. Domestic and Foreign Literature (a sociology journal), a 2016 article I published (back in my “HBD” days titled “Race, Obesity, Poverty, and IQ, writing:
income and education (which in the latter case presumably correlates with IQ levels). They have the highest prevalence of type 2 diabetes. In terms of ethnicity, overweight indicators are as follows: 67.3% for whites, 75.6% for African Americans and 77.9% for Latinos. Summing up all this, we obtain, in the words of the authors of the study, “politically incorrect conclusions”: African Americans and Hispanics are more at risk of living in poverty, have lower IQ, higher rates of obesity and a chance of developing diabetes; The main factor in these correlations is the IQ level (Race, obesity, poverty and IQ, 2016).
Almost four years later (after my views have undergone a significant change) I would draw different conclusions. Blacks are 51% more likely to be obese than whites (Lincoln, Abdou, and Lloyd, 2016) with the cause being a multitude of factors. Though it seems that black American men with more African ancestry may be protected against central adiposity (Klimentidis et al, 2016). Racial disparities in obesity are due to an interaction of a multitude of factors (Byrd, Toth, and Stanford, 2018). Interestingly, black kids with obesity don’t perceive themselves as obese (Lankarani and Assani, 2018), which, presumably, is due to higher rates of obesity in the black population. Black girls are more likely to have an earlier menarche than white giris (e.g., Freedman et al, 2000) and it is because black girls are more likely to be obese than white girls which is due to the effects of leptin being permissive for menarche, from the higher levels of body fat in black girls (Salsberry, Reagen, and Pajer, 2010).
We must look to social determinants of health to understand why certain non-white populations are more likely to be obese than others. Looking at “IQ” as causal for obesity—which I used to believe—obscures much more than it helps. We can look to epigenetic effects, for example, regarding biological explanations of obesity (Krueger and Reithner, 2016), for instance high BMI in black women being related to saliva-based DNA methylation, which is used as a marker for aging (Li et al, 2019). Even perceived racism (it does not have to be actual) can have physiologic effects on black women, heigtening cortisol levels, leading to a heigtened obesity risk (Mwendwa et al, 2016).
In any case, it’s cool that I got cited but uncool that it was something that I don’t believe anymore.
The second citation comes from Rossi (2020: 13) in the journal Social Science Information titled New avenues in epigenetic research about race: Online activism around reparations for slavery in the United States citing my article Race, Medicine, and Epigenetics: How the Social Becomes Biological:
Consequently, social scientists’ opinions about epigenetic research dealing with race and slavery have sometimes been scrutinized by blog authors. For example, the article untitled [sic] ‘Race, medicine, and epigenetics: How the social becomes biological’ published in 2019 on the blog Notpoliticallycorrect features a long discussion on whether race could be seen as a viable variable to discuss the epigenetics of trauma, especially relating to slavery in the US.14 After summarizing the views of legal scholar and sociologist Dorothy Roberts, who has argued repeatedly in her works against the use of the concept of race in biomedical sciences, the author sides with philosophers Michael Hardimon and Shannon Sullivan, who are both enthusiastic about the inclusion of race to discuss genetics and epigenetics:
Race and medicine is a tendentious topic. On one hand, you have people like sociologist Dorothy Roberts (2012) who argues against the use of race in a medical context, whereas philosopher of race Michael Hardimon thinks that we should not be exclusionists about race when it comes to medicine. If there are biological races, and there are salient genetic differences between them, then why should we disregard this when it comes to a medically relevant context? [. . .] So, we should not be exclusionists (like Roberts), we should be inclusionists (like Hardimon). [. . .] Furthermore, acknowledging the fact that the social dimensions of race can help us understand how racism manifests itself in biology (for a good intro to this see Sullivan’s (2015) book The Physiology of Racist and Sexist Oppression, for even if the ‘oppression’ is imagined, it can still have very real biological effects that could be passed onto the next generation – and it could particularly affect a developing fetus, too). It seems that there is a good argument that the effects of slavery could have been passed down through the generations manifesting itself in smaller bodies.
Relying also on Jasienska’s research, the author of this blog post therefore dismissed the idea that race should not be applied to the medical field, while using the words and legitimacy of humanities scholars such as Hardimon and Sullivan to back up their claims. These contributions show the way journalists and various blog authors write about epigenetics by mixing together scientific articles in various fields (the social sciences, philosophy, psychiatry, social work) in an effort to bring more legitimacy to the topic. This process highlights the ways in which lay circles produce new connections between various papers and texts dealing with epigenetics, no matter how different their fields of expertise may be.
This shows a very sharp contrast with my current views and my older views on race and obesity. Before, thinking that obesity was “determined” by IQ (e.g., Kanazawa, 2012; Kanazawa, 2014) was an error—people with low “IQs” are more likely to be in poverty and have less access to good foods, along with the abundance of fast food restaurants in areas with a higher concentration of blacks (James et al, 2014). Black women, for instance, have a lower RMR than white women (Gannon, DiPietro, and Poehlman, 2000)
These two articles of mine that were cited (on similar issues, no less) show the evolution of my views over the past four or so years in between the publication of the two articles on this blog. This is a good case study on how the one can view the aetiology of one thing completely different based on the types of views they previously held. The views of obesity and race I hold now are much more complex than the reductive “it’s genes/IQ” kind of guy that I used to be. A more holistic view of obesity disparities, factoring in access to food (food swamps/deserts), income, location etc is more informative than looking just to “IQ” or “genes for” obesity—because even if “genes for” obesity exist and even if “genes for” obesity are distributed unevenly across races, the predominant determinant of weight will be activity level/caloric consumption, which is based on SES and other factors—not “IQ” or “obesity genes.” The social does become biological, and it does have consequences for obesity disparities between and within races.
Charles Murray’s Philosophically Nonexistent Defense of Race in “Human Diversity”
2250 words
Charles Murray published his Human Diversity: The Biology of Gender, Race, and Class on 1/28/2020. I have an ongoing thread on Twitter discussing it.
Murray talks of an “orthodoxy” that denies the biology of gender, race, and class. This orthodoxy, Murray says, are social constructivists. Murray is here to set the record straight. I will discuss some of Murray’s other arguments in his book, but for now I will focus on the section on race.
Murray, it seems, has no philosophical grounding for his belief that the clusters identified in these genomic runs are races—and this is clear with his assumptions that groups that appear in these analyses are races. But this assumption is unfounded and Murray’s assumption that the clusters are races without any sound justification for his belief actually undermines his claim that races exist. That is one thing that really jumped out at me as I was reading this section of the book. Murray discusses what geneticists say, but he does not discuss what any philosophers of race say. And that is to his downfall.
Murray discusses the program STRUCTURE, in which geneticists input the number of clusters they want and, when DNA is analyzed (see also Hardimon, 2017: chapter 4). Rosenberg et al (2002) sampled 1056 individuals from 52 different populations using 377 microsatellites. They defined the populations by culture, geography, and language, not skin color or race. When K was set to 5, the clusters represented folk concepts of race, corresponding to the Americas, Europe, East Asia, Oceania, and Africa. (See Minimalist Races Exist and are Biologically Real.) Yes, the number of clusters that come out of STRUCTURE are predetermined by the researchers, but the clusters “are genetically structured … which is to say, meaningfully demarcated solely on the basis of genetic markers” (Hardimon, 2017: 88).
Races as clusters
Murray then discusses Li et al, who set K to 7 and North Africa and the Middle East were new clusters. Murray then provides a graph from Li et al: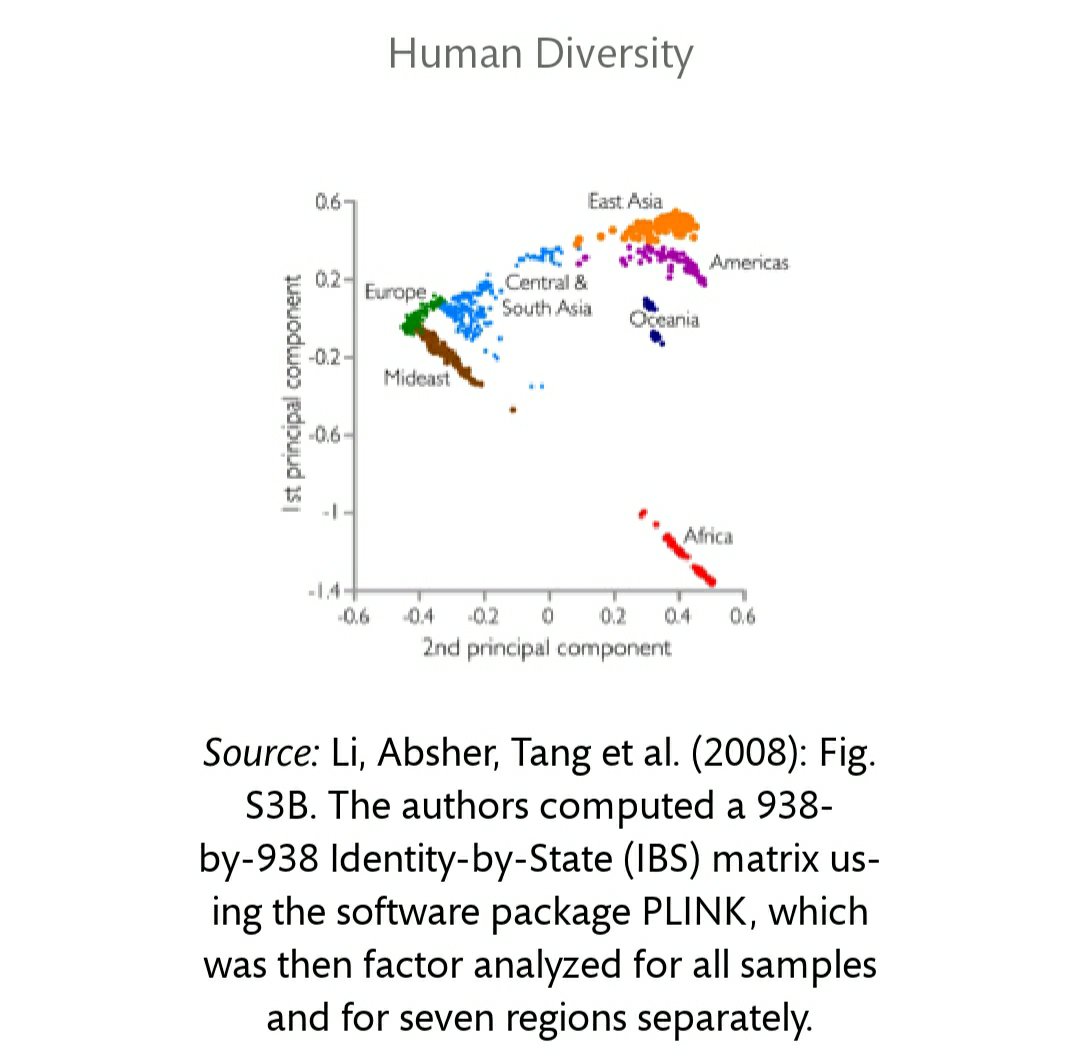
So, Murray’s argument seems to be “(1) If clusters that correspond to concepts of race setting K to 5-7 appear in STRUCTURE and cluster analyses, then (2) race exists. (1). Therefore (2).” Murray is missing a few things here, namely conditions (see below) that would place the clusters into the racial categories. His assumption that the clusters are races—although (partly) true—is not bound by any sound reasoning, as can be seen by his partitioning Middle Easterners and North Africans as separate races. Rosenberg et al (2002) showed the Kalash in K=6, are they a race too?
No, they are not. Just because STRUCTURE identifies a population as genetically distinct, it does not entail that the population in question is a race because they do not fit the criteria for racehood. The fact that the clusters correspond to major areas means that the clusters represent continental-level minimalist races so races, therefore, exist (Hardimon, 2017: 85-86). But to be counted as a continental-level minimalist race, the group must fit the following conditions (Hardimon, 2017: 31):
(C1) … a group is distinguished from other groups of human beings by patterns of visible physical features
(C2) [the] members are linked by a common ancestry peculiar to members of that group, and
(C3) [they] originate from a distinctive geographic location[…]
…what it is for a group to be a race is not defined in terms of what it is for an individual to be a member of a race. What it means to be an individual member of a minimalist race is defined in terms of what it is for a group to be a race.
Murray (paraphrased): “Cluster analyses/STRUCTURE spit out these continental microsatellite divisions which correspond to commonsense notions of race.” What is Murray’s logic for assuming that clusters are races? It seems that there is no logic behind it—just “commonsense.” (See also Fish, below.) Due to not finding any arguments for accepting X number of clusters as the races Murray wants, I can only assume that Murray just chose which one agreed with his notions and use for his book. (If I am in error, then if there is an argument in the book then maybe someone can quote it.) What kind of justification is that?
Compared to Hardimon’s argument and definition. Homo sapiens is:
… a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population. (Hardimon, 2017: 99)
[…]
Step 1. Recognize that there are differences in patterns of visible physical features of human beings that correspond to their differences in geographic ancestry.
Step 2. Observe that these patterns are exhibited by groups (that is, real existing groups).
Step 3. Note that the groups that exhibit these patterns of visible physical features correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race.
Step 4. Infer that minimalist race exists. (Hardimon, 2017: 69)
While Murray is right that the clusters that correspond to the folk races appear in K = 5, you can clearly see that Murray assumes that ALL clusters would then be races and this is where the philosophical emptiness of Murray’s account comes in. Murray has no criteria for his belief that the clusters are races, commonsense is not good enough.
Philosophical emptiness
Murray then lambasts the orthodoxy for claiming that race is a social construct.
Advocates of “race is a social construct” have raised a host of methodological and philosophical issues with the cluster analyses. None of the critical articles has published a cluster analysis that does not show the kind of results I’ve shown.
Murray does not, however, discuss a more critical article of Rosenberg et al (2002)—Mills (2017) – Are Clusters Races? A Discussion of the Rhetorical Appropriation of Rosenberg et al’s “Genetic Structure of Human Populations.” Mills (2017) discusses the views of Neven Sesardic (2010)—philosopher—and Nicholas Wade—science journalist and author of A Troublesome Inheritance (Wade, 2014). Both Wade and Seasardic are what Kaplan and Winther (2014) term “biological racial realists” whereas Rosenberg et al (2002), Spencer (2014), and Hardimon (2017) are bio-genomic/cluster realists. Mills (2017) discusses the “misappropriation” of the bio-genomic cluster concept due to the “structuring of figures [and] particular phrasings” found in Rosenberg et al (2002). Wade and Seasardic shifted from bio-genomic cluster realism to their own hereditarian stance (biological racial realism, Kaplan and Winther, 2014). While this is not a blow to the positions of Hardimon and Spencer, this is a blow to Murray et al’s conception of “race.”
Murray (2020: 144)—rightly—disavows the concept of folk races but wrongly accepting the claim that we dispense with the term “race”:
The orthodoxy is also right in wanting to discard the word race. It’s not just the politically correct who believe that. For example, I have found nothing in the genetics technical literature during the last few decades that uses race except within quotation marks. The reasons are legitimate, not political, and they are both historical and scientific.
Historically, it is incontestably true that the word race has been freighted with cultural baggage that has nothing to do with biological differences. The word carries with it the legacy of nineteenth-century scientific racism combined with Europe’s colonialism and America’s history of slavery and its aftermath.
[…]
The combination of historical and scientific reasons makes a compelling case that the word race has outlived its usefulness when discussing genetics. That’s why I adopt contemporary practice in the technical literature, which uses ancestral population or simply population instead of race or ethnicity …
[Murray also writes on pg 166]
The material here does not support the existence of the classically defined races.
(Nevermind the fact that Murray’s and Herrnstein’s The Bell Curve was highly responsible for bringing “scientific racism” into the 21st century—despite protestations to the contrary that his work isn’t “scientifically racist.”)
In any case, we do not need to dispense with the term race. We only need to deflate the term (Hardimon, 2017; see also Spencer, 2014). Rejecting claims from those termed biological racial realists by Kaplan and Winther (2014), both Hardimon (2017) and Spencer (2014; 2019) deflate the concept of race—that is, their concepts only discuss what we can see, not what we can’t. Their concepts are deflationist in that they take the physical differences from the racialist concept (and reject the psychological assumptions). Murray, in fact, is giving into this “orthodoxy” when he says that we should stop using the term “race.” It’s funny, Murray cites Lewontin (an eliminativist about race) but advocates eliminativism of the word but still keeping the underlying “guts” of the concept, if you will.
We should only take the concept of “race” out of our vocabulary if, and only if, our concept does not refer. So for us to take “race” out of our vocabulary it would have to not refer to any thing. But “race” does refer—to proper names for a set of human population groups and to social groups, too. So why should we get rid of the term? There is absolutely no reason to do so. But we should be eliminativist about the racialist concept of race—which needs to exist if Murray’s concept of race holds.
There is, contra Murray, material that corresponds to the “classically defined races.” This can be seen with Murra’s admission that he read the “genetics technical literature”. He didn’t say that he read any philosophy of race on the matter, and it clearly shows.
To quote Hardimon (2017: 97):
Deflationary realism provides a worked-out alternative to racialism—it is a theory that represents race as a genetically grounded, relatively superficial biological reality that is not normatively important in itself. Deflationary realism makes it possible to rethink race. It offers the promise of freeing ourselves, if only imperfectly, from the racialist background conception of race.
Spencer (2014) states that the population clusters found by Rosenberg et al’s (2002) K = 5 run are referents of racial terms used by the US Census. “Race terms” to Spencer (2014: 1025) are “a rigidly designating proper name for a biologically real entity …” Spencer’s (2019b) position is now “radically pluralist.” Spencer (2019a) states that the set of races in OMB race talk (Office of Management and Budget) is one of many forms “race” can take when talking about race in the US; the set of races in OMB race talk is the set of continental human populations; and the continental set of human populations is biologically real. So “race” should be understood as proper names—we should only care if our terms refer or not.
Murray’s philosophy of race is philosophically empty—Murray just uses “commensense” to claim that the clusters found are races, which is clear with his claim that ME/NA people constitute two more races. This is almost better than Rushton’s three-race model but not by much. In fact, Murray’s defense of race seems to be almost just like Jensen’s (1998: 425) definition, which Fish (2002: 6) critiqued:
This is an example of the kind of ethnocentric operational definition described earlier. A fair translation is, “As an American, I know that blacks and whites are races, so even though I can’t find any way of making sense of the biological facts, I’ll assign people to my cultural categories, do my statistical tests, and explain the differences in biological terms.” In essence, the process involves a kind of reasoning by converse. Instead of arguing, “If races exist there are genetic differences between them,” the argument is “Genetic differences between groups exist, therefore the groups are races.”
So, even two decades later, hereditarians are STILL just assuming that race exists WITHOUT arguments and definitions/theories of race. Rushton (1997) did not define “race”, and also just assumed the existence of his three races—Caucasians, Mongoloids, and Negroids; Levin (1997), too, just assumes their existence (Fish, 2002: 5). Lynn (2006: 11) also uses a similar argument to Jensen (1998: 425). Since the concept of race is so important to the hereditarian research paradigm, why have they not operationalized a definition and rely on just assuming that race exists without argument? Murray can now join the list of his colleagues who also assume the existence of race sans definition/theory.
Conclusion
Hardimon’s and Spencer’s concepts get around Fish’s (2002: 6) objection—but Murray’s doesn’t. Murray simply claims that the clusters are races without really thinking about it and providing justification for his claim. On the other hand, philosophers of race (Hardimon, 2017; Spencer, 2014; 2019a, b) have provided sound justification for the belief in race. Murray is not fair to the social constructivist position (great accounts can be found in Zack (2002), Hardimon (2017), Haslanger (2000)). Murray seems to be one of those “Social constructivists say race doesn’t exist!” people, but this is false: Social constructs are real and the social can does have potent biological effects. Social constructivists are realists about race (Spencer, 2012; Kaplan and Winther, 2014; Hardimon, 2017), contra Helmuth Nyborg.
Murray (2020: 17) asks “Why me? I am neither a geneticist nor a neuroscientist. What business do I have writing this book?” If you are reading this book for a fair—philosophical—treatment for race, look to actual philosophers of race and don’t look to Murray et al who do not, as shown, have a definition of race and just assume its existence. Spencer’s Blumenbachian Partitions/Hardimon’s minimalist races are how we should understand race in American society, not philosophically empty accounts.
Murray is right—race exists. Murray is also wrong—his kinds of races do not exist. Murray is right, but he doesn’t give an argument for his belief. His “orthodoxy” is also right about race—since we should accept pluralism about race then there are many different ways of looking at race, what it is, and its influence on society and how society influences it. I would rather be wrong and have an argument for my belief then be right and appeal to “commonsense” without an argument.
Nature, Nurture, and Athleticism
1600 words
Nature vs nurture can be said to be a debate on what is ‘innate’ and what is ‘acquired’ in an organism. Debates about how nature and nurture tie into athletic ability and race both fall back onto the dichotomous notion. “Athleticism is innate and genetic!”, the hereditarian proclaims. “That blacks of West African ancestry are over-represented in the 100m dash is evidence of nature over nurture!” How simplistic these claims are.
Steve Sailer, in his response to Birney et al on the existence of race, assumes that because those with ancestry to West Africa consistently have produced the most finalists (and winners) in the Olympics that race, therefore, must exist.
I pointed out on Twitter that it’s hard to reconcile the current dogma about race not being a biological reality with what we see in sports, such as each of the last 72 finalists in the Olympic 100-meter dash going all the way back to 1984 nine Olympics ago being at least half sub-Saharan in ancestry.
the abundant data suggesting that individuals of sub-Saharan ancestry enjoy genetic advantages.
[…]
For example, it’s considered fine to suggest that the reason that each new Dibaba is fast is due to their shared genetics. But to say that one major reason Ethiopians keep winning Olympic running medals (now up to 54, but none at any distance shorter than the 1,500-meter metric mile because Ethiopians lack sprinting ability) is due to their shared genetics is thought unthinkable.
Sailer’s argument seems to be “Group X is better than Group Y at event A. Therefore, X and Y are races”, which is similar to the hereditarian arguments on the existence of ‘race’—just assume they exist.
The outright reductionism to genes in Sailer’s view on athleticism and race is plainly obvious. That blacks are over-represented in certain sports (e.g., football and basketball) is taken to be evidence for this type of reductionism that Sailer and others appeal to (Gnida, 1995). Such appeals can be said to be implicitly saying “The reason why blacks succeed at sport is due to genes while whites succeed due to hard work, so blacks don’t need to work as hard as whites when it comes to sports.”
There are anatomic and physiological differences between groups deemed “black” and “white”, and these differences do influence sporting success. Even though this is true, this does not mean that race exists. Such reductionist claims—as I myself have espoused years ago—do not hold up. Yes, blacks have a higher proportion of type II muscle fibers (Caesar and Henry, 2015), but this does not alone explain success in certain athletic disciplines.
Current genetic testing cannot identify an athlete (Pitsiladis et al, 2013). I reviewed some of the literature on power genotypes and race and concluded that there are no genes yet identified that can be said to be a sufficient cause of success in power sports.
Just because group A has gene or gene networks G and they compete in competition C does not mean that gene or gene networks G contribute in full—or in part—to sporting success. The correlations could be coincidental and non-functional in regard to the sport in question. Athletes should be studied in isolation, meaning just studying a specific athlete in a specific discipline to ascertain how, what, and why works for the specific athlete along with taking anthropomorphic measures, seeing how bad they want “it”, and other environmental factors such as nutrition and training. Looking at the body as a system will take us away from privileging one part over another—while we also do understand that they do play a role but not the role that reductionists believe.
No evidence exists for DNA variants that are common to endurance athletes (Rankinen et al, 2016). But they do have one thing in common (which is an environmental effect on biology): those born at altitude have a permanently altered ventilatory response as adults while “Peruvians born at altitude have a nearly 10% larger forced vital capacity compared to genetically matched Peruvians born at sea level” (Brutasaert and Parra, 2009: 16). Certain environmental effects on biology are well-known, and those biological changes do help in certain athletic events (Epstein, 2014). Yan et al (2016) conclude that “conclude that the traditional argument of nature versus nurture is no longer relevant, as it has been clearly established that both are important factors in the road to becoming an elite athlete.”
Georgiades et al (2017) go the other way and what they argue is clear in the title of their paper “Why nature prevails over nurture in the making of the elite athlete.” They continue:
Despite this complexity, the overwhelming and accumulating evidence, amounted through experimental research spanning almost two centuries, tips the balance in favour of nature in the “nature” and “nurture” debate. In other words, truly elite-level athletes are built – but only from those born with innate ability.
They use twin studies as an example stating that since heritability is greater than 50% but lower than 100% means “that the environment is also important.” But this is a strange take, especially from seasoned sports scientists (like Pitsiladis). Attempting to partition traits into a ‘nature’ and ‘nurture’ component and then argue that the emergence of that trait is due more to genetics than environment is an erroneous use of heritability estimates. It is not possible—nor is it feasible—to separate traits into genetic and environmental components. The question does not even make sense.
“… the question of how to separate the native from the acquired in the responses of man does not seem likely to be answered because the question is unintelligible.” (Leonard Carmichael 1925, quoted in Genes, Determinism and God, Alexander, 2017)
Tucker and Collins (2012) write:
Rather, individual performance thresholds are determined by our genetic make-up, and training can be defined as the process by which genetic potential is realised. Although the specific details are currently unknown, the current scientific literature clearly indicates that both nurture and nature are involved in determining elite athletic performance. In conclusion, elite sporting performance is the result of the interaction between genetic and training factors, with the result that both talent identification and management systems to facilitate optimal training are crucial to sporting success.
Tucker and Collins (2012) define training as the realization of genetic potential, while DNA “control the ceiling” of what one may be able to accomplish. “… training maximises
the likelihood of obtaining a performance level with a genetically controlled ‘ceiling’, accounts for the observed dominance of certain populations in specific sporting disciplines” (Tucker and Collins, 2012: 6). “Training” would be the environment here and the “genetically controlled ‘ceiling'” would be genes here. The authors are arguing that while training is important, training is just realizing the ‘potential’ of what is ‘already in’ the genes—an erroneous way of looking at genes. Shenk (2010: 107) explains why:
As the search for athletic genes continues, therefore, the overwhelming evidence suggests that researchers will instead locate genes prone to certain types of interactions: gene variant A in combination with gene variant B, provoked into expression by X amount of training + Y altitude + Z will to win + a hundred other life variables (coaching, injuries, etc.), will produce some specific result R. What this means, of course, What this means, of course, is that we need to dispense rhetorically with thick firewall between biology (nature) and training (nurture). The reality of GxE assures that each person’s genes interacts with his climate, altitude, culture, meals, language, customs and spirituality—everything—to produce unique lifestyle trajectories. Genes play a critical role, but as dynamic instruments, not a fixed blueprint. A seven- or fourteen- or twenty-eight-year-old is not that way merely because of genetic instruction.
The model proposed by Tucker and Collins (2012) is pretty reductionist (see Ericsson, 2012 for a response), while the model proposed by Shenk (2010) is more holistic. The hypothetical model explaining Kenyan distance running success (Wilbur and Pitsiladis, 2012) is, too, a more realistic way of assessing sport dominance:

The formation of an elite athlete comes down to a combination of genes, training, and numerous other interacting factors. The attempt to boil the appearance of a certain trait to either ‘genes’ or ‘environment’ and partition them into percentages is an unsound procedure. That a certain group continuously wins a certain event does not constitute evidence that the group in question is a race, nor does it constitute evidence that ‘genes’ are the cause of the outcome between groups in that event. The holistic model of human athletic performance in which genes contribute to certain physiological processes along with training, and other biomechanical and psychological differences is the correct way to think about sport and race. Actually seeing an athlete in motion in his preferred sport is (and I believe always will be) superior to just genetic analyses. Genetic tests also have “no role to play in talent identification” (Webborn et al, 2015).
One emerging concept is that there are many potential genetic pathways to a given phenotype [41]. This concept is consistent with ideas that biological redundancy underpins complex multiscale physiological responses and adaptations in humans [42]. From an applied perspective, the ideas discussed in this review suggest that talent identification on the basis of DNA testing is likely to be of limited value, and that field testing, which is essentially a higher order ‘bioassay’, is likely to remain a key element of talent identification in both the near and foreseeable future [43]. (Joyner, 2019; Genetic Approaches for Sports Performance: How Far Away Are We?)
Athleticism is irreducible to biology (Louis, 2004). Holistic (nature and nurture) will beat the reductionist (nature vs nurture) views; with how biological systems work, there is no reason to privilege one level over another (Noble, 2012), so there is no reason to privilege the gene over the environment, environment over the gene. The interaction of multiple factors explains sport success.
Race/Ethnicity and Pain
1700 words
There are many superficial physical differences between the races. But differences in pain sensitivity would be one that is not really “superficial”, as you can’t really see it (you can see someone’s reaction to pain, but not see it). “Pain” is defined as physical discomfort caused by injury. There are some myths about pain differences between racial groups, that still persist today. And these myths have bad consequences.
For example, Hoffman et al (2016) state that “people assume a priori that blacks feel less pain than do whites.” Hoffman et al (2016) carried out two studies: (1) using a between-participants design, laymen were asked to assess the pain of white and black subjects and (2) again using a between-participants design, they asked students and medical doctors to assess pain between blacks and whites. In (2) they asked these 15 questions:
1. On average, Blacks age more slowly than Whites.
2. Black people’s nerve-endings are less sensitive than White
people’s nerve-endings.
3. Black people’s blood coagulates more quickly–because of
that, Blacks have a lower rate of hemophilia than Whites.
4. Whites, on average, have larger brains than Blacks.
5. Whites are less susceptible to heart disease like hypertension than Blacks.
6. Blacks are less likely to contract spinal cord diseases like
multiple sclerosis.
7. Whites have a better sense of hearing compared with Blacks.
8. Black people’s skin has more collagen (i.e., it’s thicker) than
White people’s skin.
9. Blacks, on average, have denser, stronger bones than Whites.
10. Blacks have a more sensitive sense of smell than Whites;
they can differentiate odors and detect faint smells better
than Whites.
11. Whites have more efficient respiratory systems than Blacks.
12. Black couples are significantly more fertile than White couples.
13. Whites are less likely to have a stroke than Blacks.
14. Blacks are better at detecting movement than Whites.
15. Blacks have stronger immune systems than Whites and are
less likely to contract colds.
(I’ll cover these questions in a future article.)
Here is the table showing the respondents’ answers to the questions:
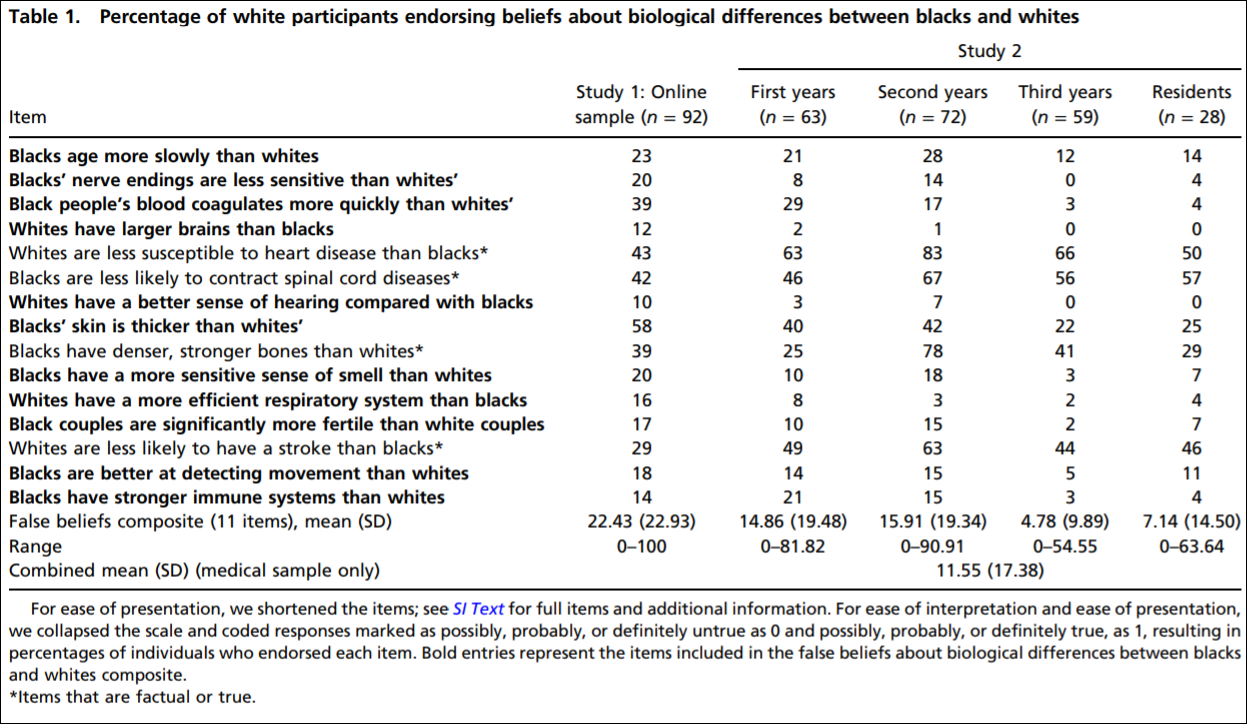
So they established that whites with no medical training hold false beliefs about black-white differences that then carry over to pain management. They showed in study 2 that medical students’ and residents’ apparently false beliefs about racial differences in the questions they answered showed bias in the accuracy of the recommended pain treatments. Hoffman et al (2016) conclude that:
The present work sheds light on a heretofore unexplored source of racial bias in pain assessment and treatment recommendations within a relevant population (i.e., medical students and residents), in a context where racial disparities are well documented (i.e., pain management). It demonstrates that beliefs about biological differences between blacks and whites—beliefs dating back to slavery—are associated with the perception that black people feel less pain than do white people and with inadequate treatment recommendations for black patients’ pain.
(See also the Psychology Today article on the matter.)
Similarly, Hollingshead et al (2016) reported that subjects, regardless of race, rated the white person more sensitive to pain and more likely to report pain than the black person. Whites reported that they were less pain sensitive and less likely to report pain than their peers. Blacks reported that they were more sensitive to pain while reporting more pain than their peers.
Interestingly, Trawalter, Hoffman, and Waytz (2012) state that black NFL players are more likely to play in a subsequent game than whites when injured, and that, as found in many other studies, blacks are more likely to feel less pain than whites. However, what the literature really shows is the opposite: blacks are more likely to feel pain than whites.
Kim et al (2017) showed that blacks, “Hispanics” and Asians had lower pain tolerance, higher pain ratings and greater temporal sensation of pain. They also showed that blacks had lower pain tolerance and higher pain ratings but no differences in pain threshold.
Blacks report greater pain regarding AIDs, glaucoma, migraine, headache, jaw pain, postoperative pain, joint pain and many other types of pain compared to whites (Green et al, 2003; Klonoff, 2009). Riley III et al’s (2002) results indicate that blacks show a stronger link between pain and emotions than whites. Obana and Davis (2016) showed that Native Hawaiian/Pacific Islander male and females reported higher pain scores than whites when it came to joint pain (but they were not significant). Bolen et al (2010) showed that work limitation, severe joint pain, and arthritis-attributable activity were higher for non-“Hispanic” blacks, “Hispanics” and multiracial people compared to non-“Hispanic” whites. Even American Indians, Alaskan natives, and Aboriginal Canadians had a higher prevalence of pain and pain symptoms than Americans (Jimenez et al, 2011).
Chan et al (2011) surveyed older Singaporeans. They found that Malay people had lower pain sensitivity compared to Chinese people, and that Indians reported greater pain sensitivity when compared with Malay and Chinese people. Australian women rated menstrual pain higher and lasting 36 percent longer than Chinese women (Zhu et al, 2010).
When it comes to potential mechanisms, physiological mechanisms are hypothesized by Campbell and Edwards (2012) who write:
For example, in comparison to non-Hispanic whites, African–Americans have reduced nociceptive flexion reflex thresholds [26]; the nociceptive flexion reflex is an electrophysiological, spinally mediated reflex, which is not amenable to voluntary control or subject to issues of response bias that plague self-report of pain experiences. This finding suggests that the observed ethnic differences in pain are unlikely to be fully explainable by sociocultural influences and hints that neurobiological processes may contribute to such differences.
Mossey (2011) shows that “Racial/ethnic minorities consistently receive less adequate treatment for acute and chronic pain than non-Hispanic whites, even after controlling for age, gender, and pain intensity.” Martinez et al (2014) showed that when it comes to colorectal and lung cancer, mixed-race individuals and blacks are more likely to report higher pain severity than whites. (Also see Shavers, Bakos, and Sheppard, 2010.)
All of the literature points in the opposite direction of the myths about pain sensitivity in regard to race: blacks feel more pain than whites and are more likely to have a lower pain tolerance. So the myths people hold about differences in pain between racial groups (mostly blacks and whites) are false. Pain is a subjective experience. And there will be differences in pain thresholds between individuals and racial groups and the causes may be both sociocultural and physiological in nature. However, this bias (in the wrong direction) speaks to what I wrote about last night: physician bias when it comes to blacks and other minorities.
Barr (2014: 183-184) writes:
Based to a certain extent on the attention given to his earlier publication, Todd moved to a faculty position with the Emory University School of Medicine, in Atlanta, Georgia. There he was able to essentially repeat his earlier study, this time examining persons coming tothe emergency room of a large, inner-city community hospital in Atlanta that was affiliated with Emory (Todd et al, 2000). He evaluated the medical records of 217 individuals coming to the emergency room over a 40-month period for treatment of an isolated long-bone fracture. Given the racial makeup of Atlanta, these included 127 blacks and 90 whites. They found that
- 54 of the blacks (43 percent) received no medication for pain during their treatment
- 23 of the whites (26 percent) received no medication for pain during their treatment
As with the earlier study in Los Angeles involving whites and Hispanics, in this study, the blacks were nearly twice as likely to receive no pain medication while in the emergency room. With this study, the authors were keenly aware of the importance of documenting the extent to which the patients expressed painful symptoms. By thoroughly reviewing the medical records of these patients, they found that 54 percent of blacks and 59 percent of whites had a notation in their medical record that they had expressed painful symptoms. The nearly twofold difference in withholding pain medication in blacks and whites was because the doctor didn’t order the medication, not because the patient didn’t want the medication.
This, again, speaks to physician bias when it comes to race in a medical context. Race is a useful tool in medicine, but to hold biases in the complete opposite direction that they exist in is wrong. This study—and many others—speak to the type of bias that physicians have against minorities in a medical context. Understanding that the differences in pain are actually the opposite from what is commonly believed by both laypeople and medical doctors is important: if blacks feel more pain than whites regarding the same injuries and they are not getting the care needed, then this speaks to physician bias. What Barr showed was that blacks were treated at the emergency room based on their ethnicity. This is wrong. Race/ethnicity is a useful tool in medicine, but to outright use it as an assumption for numerous factors makes no sense and could cause more harm than good.
Using race in a medical context is a good thing. But using race in a medical context using essentialist, outdated views about race is wrong and can lead to many horrible outcomes. Of course, using race in this context can and does lead to certain things being discovered over others. For instance, if one’s race is assumed to be “driving” one’s illness (i.e., that one has a disease that that race/ethny is more likely to have), then race can and is a good marker to use—specifically geographic ancestry. However, when it comes to things like pain management, this obviously leads to false ideas about how different groups manage and feel pain.
Views about racial differences in pain affect both laypeople and medical doctors. These views can be and are harmful. The literature points to the case being the opposite of what is believed by people: blacks have lower pain tolerance and higher pain ratings than whites. These types of differences are also found between many other races and ethnic groups. The causes could be both sociocultural and physiological. A person’s response to pain depends on their unique physiology, life experiences, ethnicity and other factors. Understanding how and why physicians are biased toward how blacks feel pain is important, along with addressing the other biases that they have about other minorities when it comes to a medical context. Race and ethnicity are important tools for medicine, but these are some of the ways that the concepts can be used with nothing good coming out of it.
Health by State and Racial Discrimination by Physicians
3000 words
I’m currently reading Health Disparities in the United States: Social Class, Race, Ethnicity and Health by medical doctor and sociologist Donald Barr. In the book, he chronicles differences in health between races and ethnies, talks about the concepts of race used and cites well-known studies to people who read this blog, and he also shows that doctors are—either conscious or not—biased against minorities in certain medical contexts.
In Chapter 1 discusses the fact that, although Americans spend the most money on health care, Americans have a lower life expectancy and higher infant mortality rate than all other developed countries, showing the association in social inequality and health across all income levels and education. In Chapter 2, he asks the question “What is health?”, discussing many concepts of what “health” is. In Chapter 3, he defines “socioeconomic status” and shows the link between poor health and poor SES. In Chapter 4, he discusses the link between inequality and poor health, introducing the concept of “allostatic load”, which is the physiologic response to being in a spot of social disadvantage.
In Chapter 5, he looks at different race concepts, since it is a main premise of the book. In Chapter 6, he shows that minorities are more likely to be in a position of low SES. He asks, if minorities of the same SES as whites are consistently found to be of lower health than whites of the same SES, is it because those with poor health tend to be minorities, that they tend to have lower SES or both? In Chapter 7, he asks the same questions while focusing on children. In Chapter 8, he examines disparities in access to healthcare, showing that even when minorities have the same insurance and doctors that minorities still face worse health outcomes (he shows that they either do not receive appropriate healthcare or receive lower-quality care). In Chapter 9, he shows that physicians treat blacks and other minorities differently, albeit unconsciously. In Chapter 10, he discusses when—if ever—a physician would be justified in using racial/ethnic categories. And in Chapter 11, he states that not all of these disparities need to be eliminated.
In Chapter 2, Barr (2014: 45) presents this table, showing rates of illness and selective rates of death between States in America. Obviously, the one to look at that is different than the others is Mississippi. Mississippi is 37.5% black. 
Wow, I wonder why Mississippi has such a high rate of obesity, diabetes, and hypertension (high blood pressure). Must be all of those obesity, diabetes and hypertension genes (HBDer).
Obesity and diabetes
The first thing to look at is median income. It is substantially lower in Mississippi compared to California, Iowa, and New York. About 23 million people in America live one mile from a supermarket, while black Americans are about half as likely to have access to supermarkets while “Hispanics” are about a third likely to have access to them (New York Law School Racial Justice Project, 2012). So when it comes to those who have to travel more than a mile for fresh fruit and vegetables, they have poorer health (Stack, 2015). So combine lower median income, along with food deserts and one can start to see how minorities have poorer health due in part to their SES. In short, living in a food desert can affect public health.
Blacks are the most obese ethnic group in America, and this relationship is largely driven by black women. Now, it’s not weird that women have higher levels of body fat than men, since women it is needed for physiological functioning. Though, there is something weird here: Black American men with more African ancestry are less likely to be obese (Klimentidis et al, 2016). Since black women and black men in America are in the same economic bracket, there must be something in the West African male physiology that “protects” them against central adiposity, though variation in social, environmental and cultural factors may play a role as well. In any case, the more West African ancestry American blacks have, the less likely they are to be obese. Klimentidis et al’s (2016) study “suggests that there are specific genetic variants and physiological mechanism(s) that differ among West African and European populations.”
Obesity affects more ethnies in America than others: non-Hispanic blacks and “Hispanics” are more likely to be obese than non-“Hispanic” whites and Asians (Hales et al, 2017). This could be due to, in part, to the variation in supermarket access and access to good foods—the concept of food deserts. Look at any low-income area near you. You’ll see a majority of corner stores with cheap, garbage food. The lack of ability to buy good food (along with the education to know what to buy and when to buy it) can explain differences in obesity rates—obviously not all. Obesity is related with diabetes, and sinec the relationship is so strong, the term “diabesity” was coined.
Eating cheap, processed carbohydrates spikes insulin. Repeated insulin spikes over time leads to type II diabetes and, eventually, obesity too. One can be skinny and have diabetes (a phenomenon known as thin on the outside, fat on the inside “TOFI”). However, since both diseases are co-morbid, we need to look at them in similar contexts. The higher rates of obesity can help to explain the higher rates of diabetes and hypertension—since those who are obese have higher blood pressure (Aronow, 2017).
Minorities are more likely to develop type II diabetes (Tuchman, 2011), and the cause of this is access to high-quality foods. But racial differences in obeisty and SES do not fully explain the higher rates of type II diabetes in black Americans; being a black American is a strong, independent factor for developing type II diabetes and this is compounded by low SES (Brancati et al, 1996). Zizi et al (2016) showed that both long and short black sleepers have an increased risk of developing type II diabetes. There are racial differences in sleep, with blacks having higher durations of long and short sleep compared to whites (Adenekan et al, 2013).
Hypertension
Now let’s look at hypertension (blood pressure). Blood pressure is a physiological variable. Since it is a physiological variable, it can and does respond to social/environmental contexts. So blood pressure can be affected by social contexts, too. For example, Williams (1992) cites stress, socioecologic stress, social support, coping patterns, health behavior, sodium and more for reasons why blacks have higher BP than whites. Dressler (1991) shows that the struggle to maintain a middle-class lifestyle is related to higher levels of BP. Similarly, Keith and Herring (1991) show that skin color is a strong predictor of occupational status and that darker-skinned blacks in America are twice as likely to experience racial discrimination than lighter-skinned blacks. This, too, can help to account for higher levels of BP between the races. In any case, Williams (1992) shows, definitively, that the causes of black-white differences in BP lie in the social environment.
Similarly, Non, Gravlee, and Mulligan (2012) show that racial disparities in BP are explained by education, and not genetic ancestry. They show that the association between BP and education was much stronger for blacks than for whites. Their results also support “the minority poverty hypothesis because the worst blood pressures were predicted for people who faced the double burden of being less educated and identifying as African American.” People who face discrimination could, and do, have higher levels of BP due to the stress they feel due to the discrimination. (Note that I take no sides on whether the discrimination is real or imagined, because even if it were imagined, it still leads to real physiologic consequences.)
Do note that there is a just-so story to explain how and why blacks have higher levels of blood pressure than whites: The Slavery Hypertension Hypothesis (Lujan and Dicarlo, 2018). This has all of the hallmarks of a just-so story posited by evolutionary psychologists. The story goes like this: Black slaves who were on the way to America in the Middle Passage had genes that favored better salt retention. So it is noted that black Americans have higher rates of BP than whites, and then they work backward and attempt to posit the best story possible to explain the current-day observation. This is the usual method evolutionary psychologists use—the method of reverse engineering, the inference from function to cause. So (1) note that blacks have higher levels of BP than whites; (2) infer the function to cause (blacks with genes that favored salt retention were more likely to survive; so (3) this is why blacks have higher rates of BP than whites. Though the explanation fails, since education, and not genetic ancestry, explains the difference in BP between blacks and whites (Non, Gravlee, and Mulligan, 2012). One only needs to understand the intricacies of physiology and how our physiological systems respond to what occurs in the greater environment.
So, obesity can explain both the higher rates of diabetes and higher rates of blood pressure, with differences in the immediate social environment explaining the rest of the differences in blood pressure between blacks and whites. (Note that heart disease deaths are directly related to hypertension. Heart disease affects blacks more than whites.)
Breast cancer
In Race, Medicine, and Epigenetics: How the Social Becomes Biological, I shortly discussed breast cancer in black women:
Black women are more likely to die from breast cancer, for example, and racism seems like it can explain a lot of it. They have less access to screening, treatment, care, they receive delays in diagnoses, along with lower-quality treatment than white women. But “implicit racial bias and institutional racism probably play an important role in the explanation of this difficult treatment” (Hardimon, 2017: 166). Furthermore, black women are more than twice as likely to acquire a type of breast cancer called “triple negative” breast cancer (Stark et al, 2010; Howlader et al, 2014; Kohler et al, 2015; DeSantis et al, 2019). Of course, this could be a relevant race-related genetic difference in disease.
Infant mortality
Now note the infant mortality rate between the states: the infant mortality rate in Mississippi is 9.7%. Smith et al (2018) show that black women are at a higher rate of having their infant die at birth. Pre-term births are related to low birth weights, and they both are related to infant mortality. Matoba and Collins (2017) write:
In the United States, African-American infants have significantly worse infant mortality than white infants. Individual risk factors alone do not explain this persistent gap, just as they did not explain the disparity in preterm birth and low birth weight. Recent studies in social determinants provide insight into the contribution of community and environmental factors to the racial disparity. Select community-level factors are potential, but partial, determinants of the racial disparity. Interpersonal and institutionalized racism is an important, and increasingly recognized, stressor for African-American women with damaging consequences to maternal and child health.
The Guardian ran a recent story on infant mortality and race, positing racism as a cause of the disparity. In any case, the social environment can and does play a part in everything discussed here today since the social can and does become biological. Part of the reason why Mississippi has a way higher rate of years of potential life lost (10,214 compared to 5500-5900 for Iowa, New York, and California) is that rates of infant mortality are higher in Mississippi. So the median age of death is 75. If an infant dies at one year of age, then that is 74 years of life lost. Therefore it is not surprising that the State with the highest level of infant mortalities has a higher number of years of potential life lost. Further, one 2017 review found that segregation was associated with increased risk of preterm birth and low birth weight for blacks (Mehra, Boyd, and Ickovis, 2017)
Note how Mississippi has lower rates of asthma. This is explained by the fact that Mississippi is more rural than, say New York. Rates of asthma are associated with living in a metropolitan area (Frazier et al, 2012; Malik, Kumar, and Frieri, 2012). (Note that blacks and other races have higher rates of asthma than other races.)
Physician bias
The lower one’s position is on the social hierarchy the lower their probability of staying healthy and having a high life expectancy; when people have the same type of health insurance and are treated for the same disease in the same hospital by the same doctor, that minority groups get worse health care, either not receiving it or receiving lower standards of quality in care. What could account for such disparities? I asked PumpkinPerson the question, and he said:
1) EGI: Doctors put more effort into saving coethnics: she looks like my italian grandma. I’ll make sure she gets the best medicine.
2) IQ: low IQ populations don’t understand the doctor’s advice and damage their health
3) r/K: some populations have faster life history so don’t live as long, even with good medical care
If (1), then the doctors need to be named, shamed, and have their medical licenses revoked. If (2), then they need better education (since IQ is just an index of middle-class knowledge). (3) is completely irrelevant, since it doesn’t make sense for humans and the concept is long-dead in ecology. In any case, PumpkinPerson danced around the true cause: differences in healthcare brought about by unconscious bias (of which (1) may be a cause). But positing (1) as a cause completely misses the point (and is the usual HBDer reductionism to genes causing most/if not all things). It’s the usual HBD/Rushtonian reductionism to genes. That’s all the HBD worldview reduces to: genes/IQ.
In any case, Reschovsky and O’Malley (2008: 229, 230)
Our results indicate that the minority makeup of physicians’ patient panels is associated with greater reports from physicians of difficulties providing high-quality care. At least some of this relationship appears to be explained by the lower resources flowing to high-minority practices.
The results of this study suggest that racial and ethnic disparities in primary health care are in part systemic in nature, and the lower resources flowing to physicians treating more minority patients are a contributing factor.
Thus, bias—whether conscious or unconscious—by physicians can explain how and why there are differences in health outcomes between people that have the same health insurance and doctor. Barr (2014: 168) states that “for black Americans, where a person lives sems to be associated with access to primary care, the quality of available hospital care, and the quality of available home care.” Barr shows that blacks receive a different level of care for a wide-range of diseases and illnesses compared to whites. For instance, Smedley et al (2003) write that “some evidence suggests that bias, prejudice, and stereotyping on the part of healthcare providers may contribute to differences in care.” Quite clearly, there is racial bias against minorities and it does seem to affect healthcare, whether or not it is intended or unintended (conscious or unconscious) (Williams and Rucker, 2000). Bird and Clinton (2001: 255) write:
Race and class-based structuring of the U.S. health delivery system has combined with other factors, including physicians’ attitudes—perhaps legacies conditioned by their participation in slavery and creation of the scientific myth of black biological and intellectual inferiority—to create a medical-social, health system cultural, and health delivery environment which contributes to the propagation of racial health disparities, and, ultimately, the health system’s race and class dilemma.
Blacks are more likely to take the advice of physicians, and to use the needed services, such as preventative care and are less likely to delay seeking care when the physician is of their own race (Saha et al, 2000; LaVeist, Nuru-Jeter, and Jones, 2008).
Blacks are more likely to perceive racism in healthcare and when they are able to choose their own doctors, they are more satisfied with their level of care (Chen et al, 2005). Chapman, Kaatz, and Carnes (2013) show that increasing awareness of implicit bias in healthcare can lower such disparities, stating that having more black doctors will alleviate such problems since they are less likely to be biased. Having a black doctor lead to more effective care for black men. Quite clearly, the race of the doctor matters for implicit biases and minority doctors lead to more effective healthcare for minorities, since they are less likely to be affected by racial biases. Minorities trust the healthcare system less than whites (Boulware et al, 2003). Black and white physicians even agree that race is a medically relevant data point, but do not agree on why (Bonham et al, 2009).
Conclusion
The table presented by Barr is telling. He purported to show that on certain indices of health, certain states fair worse than others. Rates of illness and rates of death between different states (with differing ethnic compositions) were compared. Using national data, he showed that Mississippi has the highest rates of death and illness (sans asthma). Social factors can and do account for the differences in hypertension between blacks and whites (and States); food deserts (lack of access to good food) can explain higher rates of obesity and diabetes and also higher rates of blood pressure between the races (and States with a higher percentage of certain racial/ethnic groups). Of course, physiological variables are affected by the social environment, so we have to look at differences in the social environment between groups to see how and why there are differences in any physiological variable we look at.
Doctors, whether consciously or not, treat minority patients differently and there is evidence that this leads to differences in health outcomes between ethnic groups in America. PP’s hypotheses don’t cut it (the only one that does it his “EGIs”, but that explanation fails; the cause is bias by the doctors but “EGIs” have nothing to do with the bias). In any case, there are social and cultural reasons why there are such health disparities between States and races/ethnies. Understanding the causes behind them can and will lead to closing the gap between them. The social can and does become biological, and this is the perfect way to show this. There are ways to lower the disparities in a medical context, and education seems to be one of them—for both patient and doctor.
Some states are healthier than others based on objective measures of health and mortality, and understanding the reasons why can and will decrease these differences.
Helmuth Nyborg on the Social Construction of Race
2000 words
Helmuth Nyborg published an article in Psych titled Race as Social Construct (Nyborg, 2019). In the article, he responds to a National Geographic article There’s No Scientific Basis for Race—It’s a Made-Up Label. In the article, Nyborg quotes, what apparently are quotations, from the article. Yet, for example when it comes to this:
‘There’s No Scientific Basis for Race’—‘It’s a Made-Up Label’… ‘Races do not exist because we are equals’, ‘the concept of race is not grounded in genetics’, etc.
The second quote “Races do not exist because we are equals” is not in the article. (Though this is probably a general call-out to so-called “social constructivists about race.”) Now, I won’t’ nit-pick about it, since he is apparently speaking to his critics who make these claims. In any case, Nyborg’s article is titled Race as Social Construct. Where are constructivists about race said to be anti-realists or eliminativists about race? If Nyborg is really speaking to constructivists about race, then he’s strawmanning their position. Because social constructivists about race are realists about race.
Take the new AAPA Statement on Race and Racism, where they write:
… race has become a social reality that structures societies and how we experience the world. In this regard, race is real, as is racism, and both have real biological consequences.
““race” as a social reality — as a way of structuring societies and experiencing the world — is very real.”
So, if constructivists about race claim that “Races do not exist”, then why are social constructivists about race literally saying “race is real” and “”race” as a social reality … is very real”? Weird… Almost as if Nyborg is strawmanning the constructivist position. Nyborg asks if “NG also think[s] of species as a social construct?“. See Elstein (2003) for a view that species are socially constructed. In any case, I don’t think that Nyborg is familiar with the philosophical literature on the status of species.
Here is Nyborg’s first syllogism:
Samuel Morton is a reprehensible model racist with a fixed defintion of race.
1. Samuel Morton is the father of scientific racism.
2. (We “know” that the father of scientific racism has THE correct understanding of race).
3. Morton thinks that races represent separate acts of creation.
4. Morton thinks that races are ranked in a divine hierarchy.
5. Morton did not think that races were closely related.
6. Morton thinks that races has distinct characters which:
(a) Are immutable or “fixed” across generations (i.e., no transmutation, aka evolution).
(b) Are homogenous of “fixed” (in these senses of fixation) across individuals within races.
Ergo,
Morton is wrong about 3-6, and thus represent the opposite of reality. We can then say, given 1-2 and 3-6, that races do not exist.
This is ridiculous. Where has anyone written anything like this, that since Morton was a “racist” that “races do not exist”? Did Gould make that claim in Mismeasure? I personally think that Morton’s analysis was flawed by his own biases, but I do not make the claim that “races do not exist” because of it.
In any case, when it comes to Gould’s critique of Morton’s skulls, contra Jensen (1982), Rushton (1997) , and Lewis et al (2011), Gould’s arguments about Morton were largely correct (Weisberg, 2014; Kaplan, Pigliucci, and Banta, 2015; Weisberg and Paul, 2016). Specifically, Weisberg (2014) writes that “Although Gould made some errors
and overstated his case in a number of places, he provided prima facia evidence, as yet unrefuted, that Morton did indeed mismeasure his skulls in ways that conformed to 19th century racial biases.”
Now when it comes to this one, we’re getting somewhere:
Race does not Relate to Geographic Location
1. There are no fixed traits with specific geographic locations …” because …
2. “… as often as isolation has created differences among populations, migration and mixing have blurred or erased them.
Ergo,
3. “… our pictures of past ‘racial structures’ are almost always wrong” and harmful.
This is a good argument. However, it fails, in my opinion. Yes, there is no sharp delineation in traits between what are purported to be racial groupings. However, for biological racial realism to be true, there do not need to be. Take my article You Don’t Need Genes to Delineate Race. By looking at average facial and morphological features that exist in any continent, we can say that, although there is no sharp gradation and there are clines in phenotypes, that does not mean that there is no what we can say “average look” for the group. (Nyborg discusses “IQ” there, but I won’t get into it.)
Now take this one:
Races do not exist: We are Equals and Africans
1. “… all humans are closely related.”
2. In a very real sense, all people alive today are Africans.”
3. Genetic diversity in Africa is much larger than outside this continent.”
4. Because they [migrants] were just a small subset of Africa’s population, the migrants took with them only a fraction of its genetic diversity.”
5. Admittedly, “… the longer two groups are separated, the more distinctive tweaks [mutations] they will acquire”, BUT …
6. “The concept of race has no genetic or scientific basis.” (NG here refers to a Craig Venter statement at a White House meeting, June 2000; see later).
Ergo,
7. “Science tells us there is no genetic or scientific basis for race. Races do not exist because we are [all] equals.”
1-5 are true; though 6-7 are false. In any case, the existence of race is not a scientific matter. The questions “What is race?”, “Is race real?”, and “If race is real, how many races are there?” are philosophical, not scientific, matters. Nyborg brings up “Lewontin’s fallacy”, but take what Hardimon (2017: 22-23) writes about the matter:
It is worth noting that the force of the argument against the existence of racialist races from Lewontin’s data analysis is unaffected by the critique A.W.F. Edwards made in his 2003 paper “Human Genetic Diversity: Lewontin’s Fallacy.” The fallacy Edwards imputes to Lewontin consists in inferring that racial classification has no taxonomic signifigance from the finding that the between-race component of human genetic diversity is very small. The inference is fallacious because the fact that the between-race component of human genetic diversity is small does not entail that racial classification has no taxonomic signifigance. Lewontin’s locus-by-locus analysis (which does not consider the possibility of a correlation between individual loci) does not preclude the possibility that individual loci might be correlated in such a way that people could be grouped into traditional racial categories. The underlying though is that racial classification would have “taxonomic signifigance” were it possible to group people into traditonal racial categories by making use of correlations between individual loci. However, Lewontin’s argument that there are no racialist races because the component of within-race genetic variation is larger than the component of between-race genetic variation is untouched by Edwards’ objection. That conclusion rests solely on Lewontin’s statistical analysis of human variation (the validity of which Edwards grants) and does not pressupose the absence of correlational structure in the genetic data. In short, Lewontin’s data do not preclude the possibility that raciual classification might have taxonomic signifigance but they do preclude the possibility that racialist races exist.
Nyborg is, obviously, pushing the concept of racialist races, though Hardimon has shown that they do not exist. Nyborg says that “Educability and IQ are arguable [sic] physiological (Spearman, 1927)“. Nope.
Nyborg then presents his next syllogism:
Admixture and Displacement Have Erased All Race Differences
1. Race implies unadmixed groups between which there are fixed—“fix”, in the sense of fixation index—traits.
2. (From Reich (2018) race implies “primeval” groups…separated tens of thousands of years ago”.
3. Genetics shows that mixture and displacement have happened again and again”… and … as a result “Differences have been blurred or erased”.
4. Thus, “there are no fixed traits associated with specific geographic locations…”
5. And “our pictures of past ‘racial structures’ are almost always wrong” and harmful.
Ergo
Since human descent groups are mixed and do not exhibit fixed trait differences and since there are no 10-thousand-year-old primeval groups, there are no races.
This one is strong, and if an eliminativist/anti-realist about race were to use this argument (remember, Nyborg doesn’t understand that social constructivists are realists about race), then it would be strong. But that human populations are mixed and do not exhibit trait differences does not mean that race does not exist, that does not follow. That is a carry-over from the racialist concept of race, which is false.
Nyborg then presents his fifth syllogism:
Race is only Skin Color Deep
1. “When people speak about race, usually they seem to be referring to skin color and, at the same time, to something more than skin color.”
2. “This is the legacy of people such as Morton, who developed the “science” of race to suit his own prejudices and got the actual science totally wrong.”
3. “Science today tells us that the visible differences between people a re accidents of history. They reflect how our ancestors dealt with sun exposure, and not much else.”
4. There is no homogenous African race.
Ergo,
Since race is only based on skin color, it is made up by racists.
I have heard an argument similar to this, and it fails. Race isn’t ONLY BASED ON skin color, but it is a marker of race, along with ancestry and location. Of course, morphology and other phenotypic traits ground the scientific concept of “race” (minimalist/populationist race). Race, of course, does not mean only skin color, there are many other ways to delineate races, with skin color being but one tell.
Nyborg then writes:
Ducrest, Keller, and Rouling, 2008) [79] thus observed that darker color is associated with greater aggressiveness in 10 mammal species, three kinds of birds, and more Lizard forms entirely evaded them. They condemned the color analogue with respect to humans, and reacted forcefully when Rushton and Templer (2009) [80] drew data from no less than 113 countries and found that “… murder, rape, and serious assault were associated with darker skin color, lower IQ, higher birth rate, higher infant mortality, higher HIV/AIDS rate, lower life expectancy, and lower income”
Yea, Ducrest, Keller, and Rouling (2008) is one study that Rushton loved, as it, supposedly, gave a basis for darker color being associated with aggressiveness in a slew of different animals. I rebutted Rushton and Templer, in any case. Their study was ridiculous and they did not even heed what Ducrest, Keller, and Rouling (2008: 507) stated “… that human populations are therefore not expected to consistently exhibit the associations between melanin-based coloration and the physiological and behavioural traits reported in our study.” Must be hard for Rushton and Templer to read.
In sum, Nyborg is wrong that racial constructivists claim that “Races do not exist”, for if they did not exist, then what would constructivists be fighting for? Nyborg seems to be talking to anti-realists/eliminativists about race. Nyborg pushes a racialist concept about race, which was refuted by Hardimon (2017: Chapter 1). Races exist in a minimal sense (Hardimon, 2017) and U.S. sense (Spencer, 2014), but not in the racialist “HBD” sense. In this case, biological racial realism (Spencer, 2011) is true, but if we are going by Kaplan and Winther’s (2014) definitions, Rushton, Jensen, Lynn, and Nyborg would be the biological racial realists, whereas myself, Hardimon and Spencer would be biogenomic/cluster race realists. It seems that Nyborg needs to brush up on the philosophical literature, because what he claims that social constructivists about race believe are not true; he just strawmanned their position. In any case, I’ve shown that constructivists about race do not believe that race is not real. They may not believe that race is real in a biological manner, but they do in a social one, and that is enough for them to be race realists and believe that race exists.
(Also note how Kaplan and Winther (2014) note that “Social racial realism defends the existence of distinct human groups in our ordinary discourse and social interactions. Such groups are often identified and stabilized by “surface” factors such as skin color or facial features.” So, again, those who push a socialrace-type concept do not deny that race exists, on the contrary, they are realists about race. Nyborg got it wrong, and some of his critiques are good against those who deny the reality of race, but his racial ontology is false.
Dorothy Roberts on Race as a Political Entity
2050 words
I recently bought Dorothy Roberts’ Fatal Invention: How Science, Politics and Big Business Re-create Race in the Twenty-First Century (2011) (it was $1.99 in the nook store, couldn’t pass it up), which, how the title of the book explains, discusses how race is recreated today using methods of the past as well as methods of the future. One of her main claims is that race is a political entity. Now, while I don’t disagree here (there are of course social aspects to what we call “races”), she completely rides against biological racial realism (eg Spencer, 2014; Hardimon, 2017). Her concept, though, is similar to Hardimon’s (2017) socialrace concept, and it is already a part of Spencer’s Blumenbachian partitions (since race is both biologically and socially constructed in the American view of race). While I do not believe that you need genes to delineate race, Roberts also goes on the attack on Rosenberg et al (2002), who both Hardimon and Spencer cite to buttress their arguments on the reality of biological races.
Race is not a biological category that is politcally charged. It is a political category that has been disguised as a biological one. (Roberts, 2011: 14)
Note how this is extremely similar to Hardimon’s socialrace concept. In Hardimon’s concept, socialraces have a biological correlate: minimalist races. Hardimon’s concept says, for example, that “Hispanics/Latinos” are socialraces but they are a group that do not have a corresponding minimalist race—because “Hispanics/Latinos” are a mixture of different races. Race IS a biological category that has been politically charged: Groups look different; groups that look different share geographic ancestry; groups that look different that share geographic ancestry are derived from the same geographic location; therefore race is a biological category and is therefore politically charged (one reason) since people do not like the out-group—people that look different from themselves.
This distinction is important because many people misinterpret the phrase “race is socially constructed” to mean that the biological category of race has a social meaning, so that each society interprets differently what is means to belong to a biological race. According to this view, first we are born into a race, and then our society determines the consequences of this natural inheritance. There is, then, no contradiction between seeing race as both biological and socially constructed. (Roberts, 2011: 14)
There, actually, IS NO CONTRADICTION between seeing race as socially and biologically constructed. Racial categories pick out real kinds in nature—which is what “biological racial realism” means. Since our racial categories pick out real kinds in nature, then, when it comes to society and social construction, whatever is believed about certain races in that society will be socially constructed. You can’t, for example, call a Nigerian Caucasian (see more on this below) because it does not make any sense.
Roberts then goes on (p. 14-15) about how “human beings do not fit the zoological definition of race” since a “biological race is a population of organisms that can be distinguished from other populations in the same species based on differences in inherited traits.” And so, since no human groups have this high degree of genetic differentiation, there are no human races, but only one human race.
Though Hardimon (2017: 99) articulates the best definition of race I have come across:
A race is a subdivision of Homo sapiens—a group of populations that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographic ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population.
So we know that (1) populations exhibit distinctive features; (2) these populations that exhibit these distinctive features correspond to that population’s geographic ancestry, (3) these populations that exhibit these distinctive features which correspond to geographic ancestry belong to a biological line of descent which was initiated by reproductive isolated and geographically separated founding populations; so (4) race exists.
We know race is a political grouping because it has its political roots in slavery and colonialism, it has served its political function over the four hundred years since its inceptio, and its boundary lines—how many there are and who belongs to each one—have shifted over time and across nations to suit those political purposes. Who qualifiies as white, black, and Indian has been the matter of countless rule changes and judicial decisions. These racial reclassifications did not occur in response to scientific advances in human biology, but in response to sociopolitical imperatives. They reveal that was is being defined, orgainzed, and interpreted is a political relationship and not an innate classification. (Roberts, 2011: 15)
We can take this two ways here: (1) point out that Roberts is conflating minimalist/populationist races with socialraces (which is exactly what she is describing to the tee). Yes, since race is partly social, then, based on the social attitudes of people which do change over time. Then, in that society, certain groups who were barred from being in another group may be allowed “into” the group. This does not mean that race is not biological. “Oh the Irish were considered “not white” at one point in time, therefore race doesn’t exist since groups can exit group A and become group B based on sociopolitical inclinations.” This, of course, goes over the distinctive phenotypic differences between groups with peculiar geographic ancestry. THAT is what defines race; what Roberts is discussing is important, since race is partly political, but it is not the whole story.
In addition to the grotesque lynchings that terrorized blacks throughout the South, an especially brutal form of reenslavement was the false imprisonment of thousands of black men who were then leased to white farmers, entrepreneurs, and corporations as a source of cheap labor.
It is in this accute distinction that between the political status of whites and blacks, this way of governing the power relationship between them, that we find the origins of race. Colonial landowners inherited slavery as an ancient practive, but they invented race as a modern system of power. They employed Aristotle’s concept of natural slaves and natural rulers to define permanent features of black and white people. Race separated human beings into two fundamentally distinct groups: those who were indelibly born to be lifelong servants and those who were born to be their masters. Race radically transformed not only what it meant to be enslaved, but what it meant to be free. (Roberts, 2011: 23)
Let’s accept Roberts’ argument here that the political status of whites and blacks was a way to govern the power relationship between them: so what? That group A subjugated group B and attempted to justify it with X, Y, and Z doesn’t mean that group A and group B are not biological races—it just means that group A subjugated group B and, in the future, there were social repercussions (which is also a part of the phenomenon of race as a partly social construct).
Roberts then discusses the Census (p. 31-35) and how ever-changing racial definitions undermine the claim that biological racial realism is true. In Spencer’s argument, the US meaning of “race” is just a referent, “specifically the referent of US racial discourse” (Spencer, 2014: 1027). This is because, in America, race-talk is tied to the census. We Americans are familiar with the racial groupings on the census since they are not only in use on the census but numerous other institutions. Spencer (2014) then discusses how we can use “phonetic cues alone (e.g., African American Vernacular English), surnames alone (e.g., “Chen”), first names alone (e.g., “Lakisha”), and visual cues alone (e.g., a person’s face)” (Spencer, 2014: 1027) to know someone’s race. Therefore, according to Spencer, the discourse used in the census is the discourse used nation-wide.
But the census does not set what “race” means on these forms: the OMB (Office of Management Budgeting) does. The OMB refers to race as a “set” of populations, and so this leads Spencer to believe that the “sets” of populations that the OMB is referring to are whites, blacks, Asians, Pacific Islanders, and American Indians. So race is a particular, not a kind as Hardimon argues.
Roberts then argues against the “new racial science”, most forcefully, against Rosenberg et al (2002). She brings up the usual discourse “…the number of genetic clusters is dictated by the computer user, not the computer program” (Roberts, 2011: 74). Roberts says that the clusters are “arbitrary.” Roberts says that Rosenberg et al’s (2002) study failed to verify 18th-century racial typology, but it did confirm what we have known since Lewontin’s (1972) analysis: that there is more genetic variation within races than between them. About 93-95 percent of human genetic variation was found to be within race whereas 5-7 percent of human genetic variation was found to be between groups.
Roberts says that the clusters are “arbitrary.” This is a common critique, but it is irrelevant. The five populations found by Structure are genetically structured—they are meaningfully demarcated on the basis of genetic markers (Hardimon, 2017: 88). Roberts also discusses the K = 6 run, which identified the Kalash people.
The fact that structure represents a population as genetically distinct does not entail that the population is a race. Nor is the idea that populations corresponding to the five major geographic areas are minimalist races undercut by the fact that structure picks out the Kalash as a genetically distinct group. Like the K=5 graph, the K=6 graph shows that modulo our assumption, continental-level races are genetically structured” (Hardimon, 2017: 88).
The five clusters identified by Rosenberg et al (2002) represent continental-level minimalist races so the five populations which correspond to the major geographic locations throughout the world are continental-level minimalist races. So it is, in fact, possible to place individuals into different their continental-level minimalist race without knowing anything about the race or ancestry of the individuals from which the microsatellites were drawn. Rosenberg et al (2002) studied the populations based on language, culture, and geography, not skin color or race.
It is true that Rosenberg et al (2002) found 4.3 percent of the overall human genetic variation to be between races—but this does not ride against claims from biological racial realists. The genetic variation is enough to say that we have partitions at K = 5.
“People are born with ancestry that comes from their parents but are assigned a race” is how Camara Jones, a research director at the Centers for Disease Control (CDC) explains it. (Roberts, 2011: 77)
People are assigned races based on the ethnicity/ancestry of the parents. A Nigerian would not be assigned to the Asian race, since the Nigerian has none of the features which make “Asians” Asian.
This is very simple: if both parents belong to race R, then the child will be race R as well. If parent 1 belongs to R1 and so does parent 2, then the child will belong to R1 as well (since the parents have distinct physical features which correspond with geographic ancestry and their ancestors derived from a distinct geographic location. So, since people are born with ancestry that comes from their parents, then they are assigned their PARENT’S race; they are not assigned A race, as if one can assign any individual to any race. But what if one parent belongs to R1 and the other belongs to R2? Hardimon’s minimalist concept is vague here; it only shows that races exist, it does not say which populations are races. If an individual’s parents belong to R1 and R2, then that individual is mixed race. The existence of mixed race people, of course, does not rail against the existence of biological races.
In sum, Roberts does make some good points (in what I have read of the book so far), but she gets it wrong on race. Hardimon and Spencer have both defended the methodology/concepts used by Rosenberg et al (2002) and in doing so, they successfully argued for the existence of biological races—though their two viewpoints differ. That race is, in part, socially (politically) constructed is irrelevant. What Roberts does not understand is that these socially constructed groups (“white”, “black”) still, very much so, capture biological differences between them. That they are socialraces does not mean that they DO NOT have different physical features which correspond to geographic ancestry. The socialrace concept (which Roberts espouses in her book) is separate from Hardimon’s other scientific race concepts. But it is already inherent in Spencer’s, since his Blumenbachian partitions are social constructs of a biological reality. You don’t need genes to delineate races and minimalist races exist and are biologically real.
(I will cover other things from her book as I get to them. I will discuss race and medicine at length.)
