
Home » Posts tagged 'Race' (Page 2)
Tag Archives: Race
Three Arguments for the Existence of Race: Establishing Population Groups as Races
2000 words
At least three arguments establish the existence and reality of biological race:
Argument (1) from Michael Hardimon’s (2017) book “Rethinking Race: The Case for Deflationary Realism” (The Argument for the Existence of Minimalist Races, see Chapters 2, 3, and 4):
The conditions of minimalist racehood are as follows:
(C1) … a group, is distinguished from other groups of human beings by patterns of visible physical features
(C2) [the] members are linked be a common ancestry peculiar to members of that group, and
(C3) [they] originate from a distinctive geographic location (Hardimon, 2017: 31).
This is the argument to prove the existence of minimalist races:
P1) There are differences in patterns of visible physical features which correspond to geographic ancestry
P2) These patterns are exhibited between real groups, existing groups (i.e., individuals who share common ancestry)
P3) These real, existing groups that exhibit these physical patterns by geographic ancestry satisfy the conditions of minimalist race
C) Therefore race exists and is a biological reality
Argument (2) from Michael Hardimon’s (2017) book “Rethinking Race: The Case for Deflationary Realism” (The Argument for the Existence of Populationist Races, see Chapters 5 and 6):
P1) The five populations demarcated by Rosenberg et al (2002) are populationist races; K = 5 demarcates populationist races.
P2) Populationist race=minimalist race.
P3) If populationist race=minimalist race, then everything from showing that minimalist races are a biological reality carries over to populationist races.
P4) Populationist races capture differences in genetic variation between continents and this genetic variation is responsible for the distinctive patterns of visible physical features which correspond to geographic ancestry who belong to biological lines of descent which were initiated by geographically isolated founding populations.
C) Therefore, since populationist races=minmalist races, and visible physical features which correspond to geographic ancestry are genetically transmitted by populations who belong to biological lines of descent, initiated by reproductively isolated founding populations, then populationist races exist and are biologically real.
Argument (3) from Quayshawn Spencer’s (2014) paper “A Radical Solution to the Race Problem” (The argument for the Existence of Blumenbachian Populations):
P1) The term “race” in America refers to biologically real entities; when speaking of race in America, Americans defer to the US Census Bureau who defers to the Office of Management and Budget (OMB).
P2) The OMB refers to race as “sets of” categories, while considering “races” to have 5 members, which correspond to the five major geographic regions.
P3) Rosenberg et al show that, at K = 5, meaningful, though small (~4.3 percent) genetic variation exists between continental-populations
C) Since Americans defer to the US Census Bureau who defers to the OMB, and the OMB refers to race as “sets of” categories which then correspond to five clusters found by Rosenberg et al’s (2002) analysis, race (what Spencer, 2014 terms “Blumenbachian populations”) must exist, though “race” is both socially constructed and biologically real.
Put another way, Spencer’s (2014) argument could also be:
P1) The US meaning of “race” is a referent, which refers to the discourse used by the US Census Bureau; the US Census Bureau refers to the discourse used by the Office of Management and Budget (OMB).
P2) The referent of “race”, in US ontology, refers to a set of human population groups, not a biological kind (sets of human population groups as denoted by the OMB), which refer to “Africans”, “Caucasians”, “East Asians”, “Native Americans”, and Pacific Islanders/Oceanians.
P3) The US meaning of race is both biologically real and socially constructed; Americans refer to real, existing groups when they talk about race.
C) If the US meaning of race is a referent which refers to the discourse used by the US Census Bureau and they refer to the OMB who discuss “sets of” population groups, then when Americans talk about race they talk about Blumenbachian partitions, since race is both biologically real and socially constructed.
The claim “Race exists” is now established. Note how Argument (1) establishes the claim that “races” are real, existing groups who are phenotypically distinct populations with differing geographic ancestry. Note how Argument (2) establishes the claim that populationist race = minimalist race and that “races” are a group of populations that exhibit a distinctive pattern of genetically transmitted phenotypic characters which then correspond to that group’s geographic ancestry who belong to a biological line of descent which was initiated by a geographically separated and reproductively isolated founding population. (This definition of “race” a subdivision of Homo sapiens is the best I’ve come across so far.) Finally, note how Argument (3) establishes the claim that race, in the American sense, is both biologically real and socially constructed. All three arguments are sound and logically valid.
Now, which groups fall into which of the five racial categories?
Caucasians
Caucasians denote a wide-range of groups; Europeans, MENA (Middle Eastern/North African) peoples, Indians are a very diverse group, racially speaking, with “Caucasoids”, “Mongoloids” and “Australoids” (Australoids would mean Pacific Islander/Oceanian) (see Kashyap et al, 2006 for an overview of ethnic, linguistic and geographic affiliations of Indians in the study). Ashkenazi Jews are taken to be a specific race in today’s modern racial ontology, however, Ashkenazi Jews do not exhibit a distinctive pattern of genetically transmitted phenotypic characters which then correspond to their geographic ancestry; they do represent a “geographically isolated and reproductively isolated founding population”, but the fact that they do not exhibit a distinctive pattern of genetically transmitted phenotypic characters means they are not a race, according to Arguments (1) and (2). Ashkenazi Jews are Caucasian, and not their own race. Of course, skin color does not denote race, it is only one marker to use to infer which groups are races.
Africans
Africans comprise all of Sub-Saharan Africa. Africa has the most genetic diversity in the human species (see Campbell and Tishkoff, 2010). Africans, in general, have long, slim bodies with a broad nose, dark skin, kinky hair (lip size is different based on the ethny in question). There are over 3,000 different ethnic groups in Africa, who all comprise the same race. Now, since Africans have the most genetic diversity this does not necessarily mean that they are so phenotypically distinct that there are tens, hundreds, thousands of races on the continent. One only needs to refer back to Arguments (1) and (2) to see that brash claims that “all Xs are Ys” don’t make any sense—especially with the arguments laid out above.
East Asians
East Asians denote a minimalist and populationist race (Arguments (1) and (2)) and Blumenbachian partition (Argument (3)). East Asians denote, obviously, those that derive from East Asia (Chinese, Japanese, Koreans, Vietnamese). These peoples are relatively short, on average, have a distinct yellow-ish tint to their skin (which is why they are sometimes called “yellow”), epicanthic folds and shorter limbs (more likely to have the endomorphic phenotype).
Native Americans
Native Americans are derived from a Siberian population that crossed the Bering Land Bridge about 14kya. They then spread throughout the Americas, becoming the “Natives” we know today. They are what used to be termed “red” people, due to their skin color. Native Americans are derived from Siberians, who share affinities with East Asians. (This will be discussed in further depth below.) They have black hair, and dark-ish skin. Populations that lived in the Americans pre-1492 expansion are part of the Native American racial grouping.
Pacific Islanders/Oceanians
The last racial grouping are Pacific Islanders. Spencer (2014: 1032) writes that we can define Oceanians (Pacific Islanders):
as the most inclusive human population born from East Asians in Oceania (Sahul and the Pacific Islands) and from the original human inhabitants of Oceania. Since Sahul was a single landmass composed of present-day Australia, New Guinea, and Tasmania 50,000–60,000 years ago, when humans first inhabited it, and since we know that the original human inhabitants of Oceania interbred to create modern Oceanians, and since temporal parts of populations are genealogically connected, it should be the case that most Oceanians have genealogical connections to the original peoples of some Pacific island. The only Oceanians who will not will be individuals who became Oceanian from interbreeding alone and Oceanians descended from indigenous peoples of Sahul but not indigenous peoples of a Pacific island (e.g., Aboriginal Australians). The final source of evidence comes from counterfactual cases. [Pacific Islanders and Australian Aborigines share a deep ancestry, see McEvoy et al, 2010.]
A group is in race X, if and only if they share a pattern of visible physical features and common geographic ancestry. If they do not share a pattern of visible physical features which correspond to common geographic ancestry then they do not constitute a race. Keep this in mind for the next two sections.
Are Oceanians black?
One claim that gets tossed around a lot (by black nationalists) is the claim that Oceanians are black due to their skin color, certain phenotypic traits. But this could just as easily be explained by convergent evolution, not that they are, necessarily, the same racial grouping. If this were true, then Australian Aborigines would be black, by proxy, since Australian Aborigines and Oceanian are the same race. The claim, though, holds no water. Just because two groups “look similar” (which I do not see), it does not follow that they are the same race, since other conditions need to be met in order to establish the claim that two separate groups belong to the same race.
Are Native Americans Mongoloid?
Lastly is the claim that Native Americans do not denote an actual racial grouping, they are either Mongoloid or a sub-race of Mongoloids.
Many authors throughout history have presumed that Native Americans were Mongoloid. Franz Boas, for example, said that the Maya Indians were Mongoloid, and that, American populations had features the most similar to Mongoloids, so they are thusly Mongoloid. Wikipedia has a great overview of the history of the “Mongoloid” terminology, with examples from authors throughout history. But that is irrelevant. Native Americans genetically transmit heritable phenotypic characters which correspond with their geographic ancestry and are genetically and geographically isolated population groups.
Although the claim that “Native Americans are Mongoloid” has been echoed for hundreds of years, a simple argument can be erected to take care of the claim:
P1) If Native Americans were East Asian/Mongoloid, then they would look East Asian/Mongoloid.
P2) Native Americans don’t look East Asian/Mongoloid, they have a distinct phenotype which corresponds to their geographic ancestry (See Hardimon’s minimalist/populationist race concepts).
C) Therefore, Native Americans are not East Asian/Mongoloid.
Establishing the claim that Native Americans are not East Asian/Mongoloid is simple. Some authors may make the claim that since they look similar (whatever that means, they don’t look similar to me), that they, therefore according to Arguments (1) and (2) they are a separate race and not a sub-race of East Asians/Mongoloids; Argument (3) further establishes the claim that they are a separate race on the basis that they form a distinct cluster in clustering analyses (Rosenberg et al, 2002) and since, Americans defer to the US Census Beureau and the US Census Beureau defers to the OMB who discusses sets of populations, then when Americans talk about race they talk about Native Americans as separate from East Asians/Mongoloids, since, according to Arguments (1) and (2) they have a distinct phenotype.
Generally, they have distinct skin colors (of course, skin color does not equal race, but it is a big tell), they have similar black, straight hair. But they are, in my opinion, just too phenotypically distinct to call them the same race as Mongoloids/East Asians. For the claim “Native Americans and Mongoloids/East Asians” to be true, they would need to satisfy P1 in Argument (1) and P4 in Argument (2). Native Americans do not satisfy P1 in Argument (1) nor do they satisfy P4 in Argument (2). Therefore, Native Americans are not Mongoloid/East Asian.
Conclusion
The claim “Race exists and is a biological reality” is clearly established by three sound, valid arguments—two from Hardimon (2017; chapters 2-6) and one from Spencer (2014). These arguments show, using the latest of genetic clustering studies, that races, as classicly defined, do indeed, exist and that our old views of race hundreds of years ago were, largely, correct. These arguments establish the existence of the old folk-racial categories. Races have distinct phenotypes which are genetically transmitted and are correlated with geographic ancestry. Some may make certain claims that “Oceanians are black” or “Native Americans are Mongoloid”, but these claims do not hold. These two groups in question are phenotypically distinct, and they come from unique geographic locations, therefore they are not a part of the races that some purport them to be.
Race, Body Fat, and Skin Folds
1250 words
Racial differences in body fat are clear to the naked eye: black women are more likely to carry more body fat than white women; Mexican American women are more likely to carry more body fat than white women, too. Different races/ethnies/genders of these races/ethnies have different formulas to assess body fat through the use of skin-folds. The sites to grasp the skin is different based on gender and race.
Body mass index (BMI) and waist circumference is overestimated in blacks, which means that they need different formulas to assess their BMI and adiposity/lean mass. Race-specific formulas/methods are needed to assess body fat and, along with it, disease risk, since blacks are more likely to be obese (black women, at least, it’s different with black American men with more African ancestry, see below). The fact of the matter is, when matched on a slew of variables, blacks had lower total and abdominal fat mass than whites.
This is even noted in Asian, black and white prepubertal children. He et al (2002) show that sex differences in body fat distribution are present in children who have yet to reach puberty and the differences in body fat in Asians is different than that from blacks and whites which also varies by sex. Asian girls had greater gynoid fat by DXA scan only, with girls having greater gynoid fat than boys. Asian girls had lower adjusted extremity fat and gynoid fat compared to white and black girls. Though, Asian boys had a lower adjusted extremity by fat as shown by DXA (a gold standard in body fat measurement) when compared to whites, but greater gynoid fat than whites and blacks.
Vickery, Cureton, and Collins, (1988), Wagner and Heyward (2000), and Robson, Bazin, and Soderstrom (1971) show that there are considerable body composition differences between blacks and whites. These differences in body composition come down to diet, of course, but there is also a genetic/physiologic component there as well. Combining the known fact that skin-fold testing is not conducive to a good estimate, black American men with more African ancestry are less likely to be obese.
Vickery, Cureton, and Collins (1988) argue that, if accurate estimates of body fat percentages are to be obtained, race-specific formulas need to be developed and used as independent variables to assess racial differences in body fat percentage. Differences in muscularity don’t seem to account for these skinfold differences, nor does greater mesomorphy. One possible explanation for differences in skinfold thickness is that blacks may store most of their body fat subcutaneously. (See Wagner and Heyward, 2000 for a review on fat patterning and body composition in blacks and whites.)
The often-used Durnin-Womersley formula which is used to predict body fat just from skin folds. However, “The 1974 DW equations did not predict %BF(DXA) uniformly in all races or ethnicities” (Davidson et al, 2011). Truesdale et al (2016) even show that numerous formulas used to estimate percent body fat are flawed, even some formulas used on different races. Most of the equations tested showed starkly different conclusions. But, this is based on NHANES data and the only data they provide regarding skin-folds is the tricep and subscapular skinfold so there may still be more problems with all of the equations used to assess body fat percentage between races. (Also see Cooper, 2010.)
Klimentidis et al (2016) show that black men—but not black women—seem to be protected against obesity and central adiposity (fat gain around the midsection) and that race negatively correlated with adiposity. The combo of male gender and West African ancestry predicted low levels of adiposity compared to black Americans with less African ancestry. Furthermore, since black men and women have—theoretically—the same SES, then cultural/social factors would not play as large a role as genetic factors in explaining the differences in adiposity between black men and black women. Black men with more African ancestry had a lower WHR and less central adiposity than black men with less African ancestry. If we assume that they had similar levels of SES and lived in similar neighborhoods, there is only one reason why this would be the case.
Klimentidis et al (2016) write:
One interpretation is that AAs are exposed to environmental and/or cultural factors that predispose them to greater obesity than EAs. Possibly, some of the genes that are inherited as part of their West-African ancestry are protective against obesity, thereby “canceling out” the obesifying effects of environment/culture, but only in men. Another interpretation is that genetic protection is afforded to all individuals of African descent, but this protection is overwhelmed by cultural and/or other factors in women.
Black men do, as is popularly believed, prefer bigger women over smaller women. For example, Freedman et al (2004) showed that black American men were more likely to prefer bigger women. Black American men “are more willing to idealize a woman
of a heavier body size, with more curves, than do their White American counterparts” (Freedman et al, 2004: 197). It is then hypothesized that black American men find these figures attractive (figures with “more curves” (Freedman et al, 2004: 197)) to protect against eating pathologies, such as anorexia and bulimia. So, it has been established that black men have thinner skin folds than whites which leads to skewed lean mass/body fat readings and black men with more African ancestry are less likely to be obese. These average differences between races, of course, contribute to differing disease acquisition.
I have covered differences in body fat in a few Asian ethnies and have come to the obvious conclusion: Asians, at the same height, weight etc as whites and blacks, will have more adipose tissue on their bodies. They, too, like blacks and whites, have different areas that need to be assessed for skin folds to estimate body fat.
Henriques (2016: 29) has a table on the equations for calculating estimated body density from skin fold measures from various populations. Of interest are the ones on blacks or ‘Hispanics‘, blacks or athletes and blacks and whites. (The table is provided from NSCA, 2008 so the references are not in the back of the text.)
For black and ‘Hispanic’ women aged 18-55 years, the sites to use for skin-folds are the chest, abdomen, triceps, subscapular, suprailiac, midaxillary, and the thigh. For blacks or athletes aged 18-61 years, the sites to use are the same as before (but a different equation is used for body fat estimation). For white women or anorexic women aged 18-55, the sites used are just triceps, suprailiac and the thigh. For black and white boys aged 6-17, only the triceps and the calf is used. It is the same for black and white girls, but, again, a different formula is used to assess body fat (Henriques, 2016: 29).
Morrison et al (2012) showed that white girls had a higher percent body fat when compared to black girls at ages 9-12 but every age after, black girls had higher percent body fat (which is related to earlier menarche in black girls since they have higher levels of body fat which means earlier puberty; Kaplowitz, 2008). Black girls, though, had higher levels of fat in their subscapular skin folds than white girls at all ages.
So, it seems, there are population-/race-specific formulas that need to be created to better assess body fat percentage in different races/ethnies and not assume that one formula/way of assessing body fat should be used for all racial/ethnic groups. According to the literature (some reviewed here and in Wagner and Heyward, 2000), these types of formulas are sorely needed to better assess health markers in certain populations. These differences in body fat percentage and distribution then have real health consequences for the races/ethnies in question.
The Concepts of Racialist Race and Socialrace
2500 words
I have chronicled Hardimon’s minimalist and populationist race concepts in a few different articles. They show that race is a biological reality. The arguments that show that race exists are sound. Hardimon, unlike Spencer, distinguishes social from biological concepts of race. For Spencer, race is both a social and biological concept. For Hardimon, race is either a biological or social concept which is why he delineates the social concept from the biological concepts. The two concepts—racialist race and socialrace—are similar, and because they are similar they will both be discussed in the same article.
The racialist concept of race
The racialist concept of race is the first concept of race that Hardimon (2017) defines. He sets out six things that need to be true of human genetics, in his eyes, for the racialist concept of race to be true:
(a) The fraction of human genetic diversity between populations must exceed the fraction of diversity between them.
(b) The fraction of human genetic diversity within populations must be small.
(c) The fraction of diversity between populations must be large.
(d) Most genes must be highly differentiated by race.
(e) The variation in genes that underlie obvious physical differences must be typical of the genome in general.
(f) There must be several important genetic differences between races apart from the genetic differences that underlie obvious physical differences.
Note: (b) says that racialist races are genetically racially homogeneous groups; (c)-(f) say that racialist races are distinguised by major biological differences.
Call (a)-(f) the racialist concept of race’s genetic profile. (Hardimon, 2017: 21)
The racialist race concept, as opposed to the populationist and minimalist race concepts, propose to rank races on traits such as intelligence, morals, and cultural characters to different races. Though, he does strawman the racialist concept (which would be the HBD concept of race, I’d say) because he says things like “To be a member of a particular race would be to be a particular person who is disposed to behave in certain ways. Because of this, if racialist races existed, race would constitute a very significant kind.” This statement aside, though, race does not constitute a very significant kind, it constitutes a modest biological kind.
If one condition for the racialist concept of race is said to be that of an essence of different races, then the racialist concept of race is surely false. This is due to population thinking. Population thinking is a type of thinking that maintains that there is no single way in which genotypes are expressed by genotypes. Thus, there is no phenotypic or genotypic property that could play the role of racial essence. “Since there are no biological essences, there are no racialist race essences, and since there are no racialist race essences, there are no racialist races. The existence of racialist races is incompatible with a broad structural principle of biology” (Hardimon, 2017: 20).
Hardimon has many arguments against the existence of racialist races, including:
The corresponding argument against the truth of the racialist concept of race is that science has not found it to be the case that members of the groups thought to be racialist races share a very large number of important properties by virtue of which they count as members of such groups. Nor does it seem likely that science will find that members of groups thought to be racialist races share a very large number of such properties. The results of Lewontin’s 1972 study and Rosenberh and colleague’s 2002 study strongly suggest that it is extremely unlikely that there are many important genetic differences between races apart from the genetic differences that underlie the obvious physical differences. (pg 24)
Another argument he puts for is that if racialist races exist, then the races would be sharply distinguished between phenotypic and genotypic characteristics. Though, since most of the variation between human races are clinal, he argues, “human populations are not sharply distinguished from one another along a broad range of phenotypic and genotypic dimensions. It follows from this that there are no racialist races” (pg 25).
Hardimon also says that, if racialist races did indeed exist, human populations would be sharply divided by skin color, yet they aren’t. These differences between races are continuous and vary between populations, thus racialist races do not exist. He also says since genetic variation in Homo sapiens is nonconcordant “there are no racialist races” (pg 25).
The likelihood that racialist races exist is especially low relative to the available alternative hypotheses, which indlude the hypothesis that there are no races, period, and the hypothesis that, whereas racialist races do not exist, minimalist races do exist. It is safe to conclude that there are no racialist races, period. (pg 25)
The racialist race concept lacks scientific respectability, it does not represent any “facts of the matter“, and it “supports and legalizes domination” (Hardimon, 2017: 62). It is therefore, socially constructed in a pernicious sense. Racialist races are both ideological and social constructions which then purport to pick out biological kinds.
The concept of socialrace
Socialrace is simply defined as the nonracialist concept of social groups that are taken to be racialist races. Socialraces refer to:
(1) a social group that is taken to be a racialist race,
(2) the social position is occupied by a particular social group that is a socialrace, or
(3) the system of social positions that are socialraces. (Hardimon, 2017: 131)
Socialraces are social groups that are taken or thought to be racialist races. Thus, they are wrongly taken to be racialist races. The two concepts socialrace and racialist race are similar—they both are hierarchical. Since socialrace is a social reality, then it must play a role in our everyday social lives.
Hardimon discusses many things regarding socialrace that many readers—myself included, on some of the things—would disagree with. For instance, he states that “institutional racism”. Hardimon claims that institutional racism “obtains when and where socialrace obtains” (pg 133).
He argues that socialrace is “inter alia a relation of power“, and so the institution is also characterized “by the unequal distribution of social goods such as liberty and opportunity, income and wealth, and the bases of self-respect. Socialrace is a system of advantages (purportedly) based on racialist race” (Hardimon, 2017: 133).
He then argues that differences in socialrace are associated with differences in life outcomes of the socialraces. Thus, “the belief that people are members of a biological race . . . is essential to the social construction of races” (Bernard Boxill, quoted in Hardimon, 2017: 133). Of course, the belief that people are members of a biological race is essential to the social construction of races, because the biological correlate of these socially constructed racial groups is the minimalist concept of race.
Socialrace, of course, has a biological correlate. That biological correlate is minimalist races. Minimalist races can be understood through the populationist race concept. Of course, saying that socialrace has a biological correlate in minimalist race does not necessarily mean that there are corresponding minimalist races for every socialrace. For example, “Hispanics/Latinos” can be said to be a socialrace, but they do not have a corresponding minimalist race because they do not genetically transmit distinct phenotypic characters which correspond to geographic ancestry. Using this terminology, the Irish were, at one point, a socialrace, whereas the Jews are treated as a socialrace today, when they are a subrace of the Caucasian race (they are not their own separate race, that’s like saying the British or Germans are a separate race). Minimalist race then appears to be a necessary condition for socialraces because they pick out real patterns of distinct physical features which correspond to geographic ancestry.
Socialraces, though, do not need to exhibit patterns of visible physical features; even if no such differences existed, groups could still be socialraces, for instance, social classes and other groups can be constructed to be socialraces. The concept of socialrace is a distinctive “race” concept (Hardimon, 2017: 139). Thus, the concept of socialrace is a distinctive concept and it is needed (because it discusses social realities). The idea of a social system based on the racialist concept of race can, therefore, said to be the idea of socialrace (Hardimon, 2017: 140).
Socialrace is clearly separate from the term ‘race’, and so, it is not a ‘race’ concept like the other three concepts, it is distinct with the “socialrace” moniker. This is how we show that, when talking about races in a social sense, there are differences between this concept and the scientific minimalist and populationist race concepts. One can think of it this way: the socialrace concept of race can be of use for sociologists and others whereas the minimalist and populationist concepts of race can be useful for biologists and population geneticists.
Whether or not a group is counted as a socialrace is contingent on whether or not the group in question is treated as a socialrace by the larger society. So, in this sense, “Hispanics/Latinos” can be said to be a socialrace, but, remember, they do not have a corresponding minimalist (populationist) race.
One is “properly” counted a member of a socialrace SR if one in fact satisfies the socially accepted criteria in the correlative putative racialist race. Thus, for example, a person belongs to the socialrace black (in the United States) if he or she has any identifiable sub-Saharan African ancestry because he or she satisfies accepted US criteria for bring a member of the racialist race black.
To “pass” (for example, for white) in a system of socialrace is to be taken to be white (to satisfy the socially accepted criteria for whiteness) despite the dact that one does not satisfy those criteria (for example, by virtue of possesion of “one drop” of “black blood”). The possibility of “passing” (in a given society) points to an important variable in the practical significance of socialrace membership. An individual counts as a member of a socialrace cimply by virtue of satisfying the socially accepted criteria for membership in the corresponding racialist race. Socialrace membership is itself a real social status with real social consequences. In the case of the socialrace black, subjection to antiblack racism is a standing possibility. But the actial practical signifigance of membership in a socialrace will vary with the degree to which the individual is subject to the norms associated with the racialist race to which the individual is taken to belong. (Hardimon, 2017: 144)
Socialraces are a real, social reality. They have a biological correlate in minimalist races and are taken to be racialist races, but racialist races do not exist therefore socialraces are social, not biological, in nature, even though the minimalist race concept can be said to be its biological correlate.
My contention is that the concept I have reflectively uncovered is aleeady in general circulation without being fully recognized as the concept that it is. I have endeavered to make it possible to get a proper hold on the concept and to secure an adequate reflective understanding of its content. If the reader thinks that my account of the concept SOCIALRACE captures a notion she or he has already been using, so much the better. I hope to have clarified that concept. As for the word ‘socialrace’, my hope is that it catches on, that its dissemenation promotes understanding of the phenomenon of socialrace, and that this in turn contribubtes to the dismantling of the latter’s existence.
Hardimon says that the concept of SOCIALRACE can help us better grasp the phenomenon of socialrace. Socialrace is real and illusory, it has “real causal powers (for example, causing people’s death) and that is illusory insofar that it appears to be racialist race” (Hardimon, 2017: 172). Socialraces exist and are a significant social reality, especially since socially determined categories can make biological realities (Kaplan, 2010).
When discussing socialraces, we can say that Jews are sometimes taken to be a socialrace (although they are a part of the Caucasian race) as are Arabs (who are also Caucasian, and thusly not a separate race from Europeans or Jews). However, the US Census Bureau categories take these groups to be “white”; that is the socialgroup “white” in the United States.
Regarding the black socialrace, they comprise numerous “Hispanic/Latino” populations (such as many Dominicans, some Puerto Ricans, Nicaraguans and other Latin American countries with high African admixture). Australian Aborigines and Pacific Islanders can be said to be part of the black socialrace in America, too. The fact of the matter is, socialrace in America just pretty much chooses features that people “think” go with race A, and if they see similar-looking people from two different continents, they will assume that they are part of the same race (as is the case with Aborigines and Pacific Islanders being black, even though they are a distinct group; McEvoy et al, 2010; Spencer, 2014).
Lastly, are Asians. In America, “Asians” are taken to be just East Asians (though in the UK when they talk about “Asians” in their police statistics or in the news, they most probably mean Pakistanis. They are not wrong, they are on the Asian continent. However, that is to mislead the people into thinking that what people term Asian (East Asians) are committing the crimes or whatnot, when it’s Pakistanis. The socialrace of Asian in America comprises East and South Asians (some would include Indians, too but they are Caucasian). The socialraces closely mirror Rushton’s three main races, but they are arbitrary, putting populations into groups where they do not belong.
Conclusion
As can be seen, the concepts of socialrace and racalist race are similar. These two concepts are needed to understand each other. Most people, when talking about race, discuss the socialrace concept so it would do them some good to read up more on the concept itself. The racialist concept of race purports to pick out biological kinds and then rank them on a hierarchy in a slew of different traits (which are not physical). Racialist races also purport that racial essences exist, but since essences as a whole do not exist, then racial essences do not exist either. Further, since genetic variation in Homo sapiens is nonconcordant, racialist races do not exist.
Socialrace is simple. Most likely, however you’ve thought about race throughout your life is the socialrace concept. It has a biological correlate in minimalist races. For socialraces to exist, there must be distinctive visible patterns of visible physical features which correspond to geographic ancestry. Thus, minimalist race is a necessary condition for socialrace. Socialrace is a social reality. And if socialrace is a social reality, then it must play a role in our everyday lives.
When these two concepts are looked at together, we can see how and why both of the concepts have been around for as long as they have: they purport to pick out a biological kind, a distinct biological kind. Racialist races do not exist, but just because racialist races do not exist does not mean that socialraces do not exist, because even if, say, minimalist or populationist races did not exist, the concept of socialrace would still be important because socially determined categories can make biological realities (Kaplan, 2010).
How Much Admixture?: On Social Isolation, the One-Drop Rule, and the Maintenance of Races
3000 words
How much admixture does it take for one race to no longer exist? The answer to the question is intuitive, and using Hardimon’s (2017) minimalist race concept, it is also easily answerable on logical grounds. For example, the answer to the question will show that the “one-drop rule” (that “one drop” of “black blood” makes one black) doesn’t make logical sense. These kinds of holdovers are from the racialist concept. Racialist races do not exist, therefore the concept of the “one-drop rule” does not either, since there are no facts of the matter the two concepts explain.
Social Isolation
The maintenance of the races that current exist depend on, at the moment, social barriers to reproduction, such as racism, segregation, differences in culture and class, role segregation and racial discrimination. Thus, social isolation is important for the maintenance of the current races. Social isolation, like geographic isolation (i.e., oceans, mountains, deserts, etc.) impedes racial interbreeding and thus ensures the continuation of the genetic transmission of distinct patterns of visible physical features which correspond to geographic ancestry.
Social isolation mechanisms have been in effect for hundreds of years, which began with the advent of African slavery to the New World. Laws against miscegenation existed in some states (Phillips, Odunlami, and Bonham, 2007), which is part of the reason why it’s (an unspoken) taboo to racially intermarry and bear children with someone not of their own race. Due to this, the few interracial unions that did produce children were specifically barred—in the eyes of society—to only be able to have children with others of their same socialrace at the lower ends of the social hierarchy.
Social isolation mechanisms have ensured the continuation of human races after the discovery of the New World when the geographic isolation mechanisms began breaking down due to exploring new lands. These isolating mechanisms on the populace ensured little admixture in the European population, but compared to European Americans, African Americans have a higher percentage of the opposite admixture. Understanding racial admixture and the genetic transmission of distinct visible physical features which correspond to geographic ancestry is extremely important to understanding when races “disappear” due to inbreeding.
Therefore, social isolation—ever since 1492—and the laws/rules that came after the breakdowns of geographic isolation between races still ensured the existence of the races as we know them today. Social factors acted as de facto physical barriers that impeded the races from breeding, thusly keeping their visible physical features intact, which means keeping their racial phenotype intact since races are defined—most importantly—on the basis of visible physical features. Social isolation can, clearly, be just about as “strong” as geographic isolation, since the social repercussions of interracial unions may exile them from the groups they were in. Thus, people would be wary of interracial unions, even if—as it seems—our culture in America seems to be swaying towards inclusivity in regard to interracial relationships, people still generally associate with and date people who look like themselves and their parents (see below).
How Much Admixture?
How much admixture can one race take before said race ceases to exist? Since C 1 (a group is distinguished from another group on the basis of distinct visible physical features) doesn’t require sharp lines between said visible physical features, C 2 (members linked by peculiar ancestry) also doesn’t require that all of the ancestors of Rs (races) be Rs.
The best possible example for an answer to the question of “How much admixture?” is simple. Think of Europeans (a subrace of the Caucasian race). When Europeans interbreed with non-Europeans, they begin to lose their distinct pattern of visible physical features which correspond to their geographic ancestry. Thus, in the case of Europeans, the answer to the question of “How much admixture?”, meaning “How much interbreeding can the European subrace take before it is “bred out” of existence?” is, of course, not too much.
Think of a union between a black woman and white man (using the social race designation; their populationist race is African and Caucasian, respectively). The child the woman bears will share some of her physical features, but barely. The baby will look more like the non-European parent, but of course, a baby who is the product of the union between an African and European will share features with both parents, and thus, the baby can “roughly fit the pattern” of a minimalist race. We can easily explain this: mixed-race individuals can err, physically, to one minimalist race over another because they are the products of individuals who do fit the patterns (of visible physical features which correspond to geographic ancestry).
Contrary to the alarmist claims heard in the media and from the altright, trends in interracial marriages do not indicate that minimalist (populationist) races are coming to an end (in this case, the white (social) race).
It is true that in the modoern (post-1492) world there is vastlty more racial interbreeding than there was before 1492. And if one is referring to the very long run, then races are almost certainly on their way out. But it is one thing to say that the human races will cease to exist at some point in the distant future and quite another to say that they are likely to disappear anytime soon. It is by no means clear that we are in an epistemic position to make the latter claim.
Contrary to what some writers suggest, recent trends in racial intermarriage in the United States do not indivate the imminent end of populationist (or minimalist) races. 5 The skyrocketing rates of intermarriage in this country notwithstanding, it remains true that the vast majority of Americans continue to marry within their own conventionally designated racial group. Despite the remarkable fact that the multiracial, multi-ethnic Americans have apparently become the fastest-growing demographic group in the United States, their numbers are still swamped by individuals who are members of a single continental-level minimalist races. 6 I don’t think that the significant fraction of DNA traceable to “Europeans” in most black Americans, and the small but real fraction of DNA traceable to “Africans” in white Americans, makes the end of the populationist (or minimalist) race significantly more imminent.
There is no evidence of which I am aware indicating that the rate at which racial interbreeding in the United States (or anywhere else) is occurring is one that would lead to the elimination of all racial differences—a situation in which no two groups could be distinguished on the basis of patterns of visible physical corresponding to differences in geographic ancestry—in the near future. To sum up: the increase frequency of encountering individuals of mixed racial ancestry does not mean that the concept of race is going to go out of business anytime soon. (Hardimon, 2017: 122)
Yaeger et al (2009) show that, in their sample, self-identification as African American is a reliable indicator of ancestry. Their findings also “suggest that self-reported race and ancestry can predict ancestral clusters, but do not reveal the extent of admixture.” Thus, self-identified race—even in the presence of admixture as is the case with African Americans—can show the racial category that an individual belongs to (based on their ancestry).
Hardimon (2017: 49) articulates a simple rule that employs the minimalist concept of race:
If both parents of an individual belong to one particular racial group R, that individual will belong to R.
What happens, however, if one parent belongs to R1 and the other parent belongs to R2. The minimalist concept of race does not say. Still less does it tell us what one’s race is if one’s grandparents belongs to an R1, another to R2, another to R3, and another to R4. This is a further respect in which the minimalist race concept is vague.
Particular conceptions of race (for example, the infamous “one-drop rule”) may specify the race of the individuals of “mixed” parentage, but the minimalist concept of race does not. The idea that a genune concept of race must specify the race of each individual is a hangover from the racialist race concept. Recall here that the minimalist racehood is not defined in terms of the characteristics of the individuals who belong to races. It is defined in terms of characteristics of groups.
So, the minimalist concept of race is vague, just like the populationist concept. But we can make one claim on the answer to the question “How much admixture?”: “Once a race loses its specific phenotype due to racial interbreeding, then the race ceases to exist.”
One drop?
The one drop rule (also known as the law of hypodescent), is a form of racial essentialism (Perez and Hirschman, 2009), which states that “one drop” of another, inferior (on the basis of racialist races) race’s blood denotes him to the inferior race in the social hierarchy. The one drop rule was created back during the slave days and signified who could breed with who, on the basis of how “pure” their blood was. It was, and still is today, a way for race deniers to deny the existence of race.
The one-drop rule stated that anyone with one black ancestor was classified as black (Pauker et al, 2009). That is, his position on the socialrace hierarchy (a hierarchy since it’s based on the false racialist race concept) is based on the fact that he has one black ancestor. Due to this, and other differing amounts of admixture in certain ethnic groups and other social groups taken to be races, people have—fallaciously—stated that races do not exist since the unions of two separate races “erases” one, or both, of the races in question.
This rule helped to ensure the maintenance of populationist races, since society frowned upon interracial marriage. This, obviously, was a social custom. The Jim Crow laws helped to ensure the maintenance of the physical characteristics of the races in question, though the laws were enacted to ensure the “racial purity” (whatever that is) of the European race, it helped to ensure lower amounts of admixture in black Americans. Thus, black Americans would be expected to self-identify as black (Liebler and Zacher, 2017).
Liebler and Zacher (2017)‘s data “supports the notion that this “rule” has some power even today, as there are almost 30 times as many people reporting that they are racially black with American Indian ancestry (weighted N=522,607) as there are people reporting American Indian race with black ancestry (weighted N=16,226).” Bryc et al (2015) show that, despite the expectations of the one drop rule “individuals identify roughly with the majority of their genetic ancestry.”
Most people in one sample that had less than 20 percent African ancestry identified as white. In the US, “Latinos” (a social-race) were estimated to have 65.1 percent European, 6.2 percent African, and 18.6 percent Native American DNA. Overall, 3.5 percent of European Americans had 1 percent or more African ancestry, while 1.4 percent of self-reported European Americans had were estimated to carry at least 2 percent African ancestry (Bryc et al, 2015).
Importantlty, Guo et al (2014) write:
The one-drop rule represents an important case in which social context trumps bio-ancestry. When asked to classify into a single race, most individuals with 30 % to 60 % African ancestry self-report as black; virtually all respondents with >60 % African ancestry self-classify as black. In contrast, a substantially higher proportion of European ancestry is “required” to self-classify or to be classified by an interviewer as white than the proportion of African ancestry necessary to self-classify or be classified as black. However, when given the option of identifying as multiracial, the majority of individuals with 40 % to 60 % African ancestry in both ROOM and Add Health and substantial proportions of individuals with >60 % African ancestry in ROOM stopped self-classifying as only black and primarily chose a multiracial classification.
“The infamous one-drop rule is peculiar to this country [America] but it is a feature of the American conception of race, not the minimalist concept of race.” (Hardimon, 2017: 56) The one-drop rule is a clear tell to how the socialrace concept acts. It is an essentialist concept, which means that it is necessarily racialist—since “one drop” of black blood makes one black—according to the rule.
The Maintenance of Races
It is possible that one society could take social measures to ensure the existence of their specific racial phenotype (that is, the existence of their minimalist race or subrace). Such a society would have to grapple with the moral and ethical underpinnings of such measures to ensure the maintenance of their phenotype (see Glannon, 2001’s book Genes and Future People for an extensive review of the moral, political, social, and ethical implications of human genetic engineering). This could also include genetic modification, though sound arguments exist that show that the way most people view genetic modification depends on a “strong view” of genetic determinism, which is false (Resnick and Vorhaus, 2006). However, it is possible that, through the will of the people in the society, that social isolation can lead to a de facto “physical” isolation through the social norms of the society in question.
However, since the races as they currently are are in no danger of non-existence, such measures, while they would (presumably) work, do not need to be taken. Such measures, though, do not need to be taken, since most people want to court with others who look like themselves, and those who are more likely to look like themselves are people of their own ethny, which is to say, people of their own populationist race. Thus, social measures to ensure the maintenance of races do not need to be taken.
As noted above, certain concepts from the days of the one drop rule are still in effect today, as a holdover from the days of Jim Crow and before. Some of these holdover concepts, though, help to maintain the races we know today. However, there is a possibility that our populationist races, too, have benefits socially constructed. Hardimon (2017: 126) writes (emphases his):
If populationist races exist, the role human action plays in their maintenance is rather more pronounced then the role it played in their genesis. Insofar as social norms and practices prohibiting or discouraging intermarriage have been the primary mechanisms preventing racial interbreeding since 1492, the maintenance of the separation has been intentional: this outcome is the very point of the discriminatory activity and practices in question. There is thus an especially strong sense in which, if populationist races exist, populationist race has been socially constructed since 1492.
Hardimon (2017: 126) goes on to say that the maintenance of populationist races “is not a natural process outside of human control”, nor is it “immutable or inalterable“, while “its existence is not an invariant, unchangeable,”natural” fact” and “The continued existence of populationist races, if it is a fact, is a fact within our power to change.” Thus, if populationist races exist (and they do), they exist by virtue of existing in nature.
So the races are not in danger of non-existence anytime soon, since the percentage of interracial unions are not too high compared to those who marry within their populationist races. The maintenance of populationist races comes down to—and will come down to, as long as humans are around—to social policies, whether enacted by state/country governments or the people themselves, sans any laws on miscegenation.
It has been said that we are attracted to people “who look like us“, “who look like our parents“, and “‘who are more similar to ourselves“. This means—NECESSARILY—that people are more likely to be attracted to people of their own race/ethnic group. People “who look like us” are co-ethnics and people of the same racial background; people who “look like our parents”, are, again, people who would share the same geographic ancestry. Since the physical features that delineate races are genetically transmitted from parent to offspring, then, people are more likely to be attracted to people of their same race. Finally, “people more similar to ourselves” doesn’t necessarily mean “people more racially/ethnically similar to ourselves”, since, of course, there are many other things that individuals have in common other than their race/ethnic group. However, it has been established that we are attracted more to people who share more similar genes than ourselves (Rushton,1997, 1998; Sebro et al, 2017). Thus, logically, since we are attracted to people who look like ourselves and our parents, we are attracted to people of our own ethnicity/race, as a matter of fact.
Conclusion
The question “How much admixture does it take for one race to no longer exist” is answered simply once the term “RACE” is defined: the amount of admixture it takes for one race to be “bred out” of existence is proportional to the amount of admixture it takes for one race’s physical features which correspond to geographic ancestry which are exhibited by the real group in question (this case being a subrace of a minimalist/populationist race). Europeans can’t take “much”, if any, other admixture, otherwise the traits that make Europeans European (which are, of course, not mutually exclusive to them, but the traits they—and their ethnies—exhibit are distinct) will disappear and so one of the Caucasian subraces will disappear as well. Social isolation, at the moment, is maintaining the races as we know them—and will far into the foreseeable future (there is no evidence that they will disappear anytime soon). “Violations” of the one drop rule abound, but they mean little to the minimalist/populationist concepts of race since the visible physical features which distinguish the races remain intact.
The fact that people are more attracted to people who look like themselves and their parents is an implicit way of saying that people are more attracted to people who are physically similar to themselves—that is, racially/ethnically similar to themselves—and shows that the races will not be going anywhere for the foreseeable future.
Human races will continue to exist as long as the social barriers that impede racial interbreeding remain. (Of course, if these social barriers did not exist, a majority of people still would court people who look like themselves and their families.) This is evidence that, contra social laws that impede or frown upon interracial marriages, we do not need such laws/rules because people stick to their own anyway. Therefore, the races are not in danger of disappearing anytime soon.
Genotypes, Athletic Performance, and Race
2050 words
Everyone wants to know the keys to athletic success, however, as I have argued in the past, to understand elite athletic performance, we must understand how the system works in concert with everything—especially in the environments the biological system finds itself in. To reduce factors down to genes, or training, or X or Y does not make sense; to look at what makes an elite athlete, the method of reductionism, while it does allow us to identify certain differences between athletes, it does not allow us to appreciate the full-range of how and why elite athletes differ in their sport of choice. One large meta-analysis has been done on the effects of a few genotypes on elite athletic performance, and it shows us what we already know (blacks are more likely to have the genotype associated with power performance—so why are there no black Strongmen or any competitors in the World’s Strongest Man?). A few studies and one meta-analysis exist, attempting to get to the bottom of the genetics of elite athletic performance and, while it of course plays a factor, as I have argued in the past, we must take a systems view of the matter.
One 2013 study found that a functional polymorphism in the angiotensinogen (ATG) region was 2 to 3 times more common in elite power athletes than in (non-athlete) controls and elite endurance athletes (Zarebska et al, 2013). This sample tested was Polish, n = 223, 156 males, 67 females, and then they further broke down their athletic sample into tiers. They tested 100 power athletes (29 100-400 m runners; 22 powerlifters; 20 weightlifters; 14 throwers and 15 jumpers) and 123 endurance athletes (4 tri-athletes; 6 race walkers; 14 road cyclists; 6 15 to 50 m cross-country skiers; 12 marathon runners; 53 rowers; 17 3 to 10 km runners; and 11 800 to 1500 m swimmers).
Zarebska et al (2013) attempted to replicate previous associations found in other studies (Buxens et al, 2009) most notably the association with the M235T polymorphism in the AGT (angiotensinogen) gene. Zarebska et al’s (2013) main finding was that there was a higher representation of elite power athletes with the CC and C alleles of the M235T polymorphism compared with endurance athletes and controls, which suggests that the C allele of the M235T gene “may be associated with a predisposition to power-oriented
events” (Zarebska et al, 2013: 2901).
Elite power athletes were more likely to possess the CC genotype; 40 percent of power athletes had the genotype whereas 13 percent of endurance had it and 18 percent of non-athletes had it. So power athletes were more than three times as likely to have the CC genotype, compared to endurance athletes and twice as likely to have it compared to non-athletes. On the other hand, one copy of the C allele was found in 55 percent of the power athletes whereas, for the endurance athletes and non-athletes, the C allele was found in about 40 percent of individuals. (Further, in the elite anaerobic athlete, explosive power was consistently found to be a difference maker in predicting elite sporting performance; Lorenz et al, 2013.)
Now we come to the more interesting parts: ethnic differences in the M235T polymorphism. Zarebska et al (2013: 2901-2902) write:
The M235T allele distribution varies widely according to the subject’s ethnic origin: the T235 allele is by far the most frequent in Africans (;0.90) and in African-Americans (;0.80). It is also high in the Japanese population (0.65–0.75). The T235 (C4027) allele distribution of the control participants in our study was lower (0.40) but was similar to that reported among Spanish Caucasians (0.41), as were the sports specialties of both the power athletes (throwers, sprinters, and jumpers) and endurance athletes (marathon runners, 3- to 10-km runners, and road cyclists), thus mirroring the aforementioned studies.
Zarebska et al (2013: 2902) conclude that their study—along with the study they replicated—supports the hypothesis that the C allele of the M235T polymorphism in the AGT gene may confer a competitive advantage in power-oriented sports, which is partly mediated through ANGII production in the skeletal muscles. Mechanisms can explain the mediation of ANGII production in skeletal muscles, such as a direct skeletal muscle hypertrophic effect, along with the redistribution of between muscle blood flow between type I (slow twitch) and II fibers (fast twitch), which would then augment power and speed. However, it is interesting to note that Zarebska et al (2013) did not find any differences between “top-elite” level athletes who had won medals in international competitions compared to elite-level athletes who were not medalists.
The big deal about this gene is that the AGT gene is part of the renin-angiotensin system which is partly responsible for blood pressure and body salt regulation (Hall, 1991; Schweda, 2014). There seems to be an ethnic difference in this polymorphism, and, according to Zarebska et al (2013), African Americans and Africans are more likely to have the polymorphisms that are associated with elite power performance.
There is also a meta-analysis on genotyping and elite power athlete performance (Weyerstrab et al, 2017). Weyerstrab et al (2017) meta-analyzed 36 studies which attempted to find associations between genotype and athletic ability. One of the polymorphisms studied was the famous ACTN3. It has been noted that, when conditions are right (i.e., the right morphology), the combined effects of morphology along with the contractile properties of the individual muscle fibers contribute to the enhanced performance of those with the RR ACTN3 genotype (Broos et al, 2016), while Ma et al (2013) also lend credence to the idea that genetics influences sporting performance. This is, in fact, the most-replicated association in regard to elite sporting performance: we know the mechanism behind how muscle fibers contract; we know how the fibers contract and the morphology needed to maximize the effectiveness of said fast twitch fibers (type II fibers). (Blacks have a higher proportion of type II fibers [see Caeser and Henry, 2015 for a review].)
Weyerstrab et al (2017) meta-analyzed 35 articles, finding significant associations with genotype and elite power performance. They found that ten polymorphisms were significantly associated with power athlete states. Their most interesting findings, though, were on race. Weyerstrab et al (2017: 6) write:
Results of this meta-analysis show that US African American carriers of the ACE AG genotype (rs4363) were more than two times more likely to become a power athlete compared to carriers of the ACE preferential genotype for power athlete status (AA) in this population.
“Power athlete” does not necessarily have to mean “strength athlete” as in powerlifters or weightlifters (more on weightlifters below).
Lastly, the AGT M235T polymorphism, while associated with other power movements, was not associated with elite weightlifting performance (Ben-Zaken et al, 2018). As noted above, this polymorphism was observed in other power athletes, and since these movements are largely similar (short, explosive movements), one would rightly reason that this association should hold for weightlifters, too. However, this is not what we find.
Weightlifting, compared to other explosive, power sports, is different. The beginning of the lifts take explosive power, but during the ascent of the lift, the lifter moves the weight slower, which is due to biomechanics and a heavy load. Ben-Zaken et al (2018) studied 47 weightlifters (38 male, 9 female) and 86 controls. Every athlete that was studied competed in national and international meets on a regular basis. Thirty of the weightlifters were also classified as “elite”, which entails participating in and winning national and international competitions such as the Olympics and the European and World Championships).
Ben-Zaken et al (2018) did find that weightlifters had a higher prevalence of the AGT 235T polymorphism when compared to controls, though there was no difference in the prevalence of this polymorphism when elite and national-level competitors were compared, which “[suggests] that this polymorphism cannot determine or predict elite competitive weightlifting performance” (Ben-Zaken et al, 2018: 38). Of course, a favorable genetic profile is important for sporting success, though, despite the higher prevalence of AGT in weightlifters compared to controls, this could not explain the difference between national and elite-level competitors. Other polymorphisms could, of course, contribute to weightlifting success, variables “such as training experience, superior equipment and facilities, adequate nutrition, greater familial support, and motivational factors, are crucial for top-level sports development as well” (Ben-Zaken et al, 2018: 39).
I should also comment on Anatoly Karlin’s new article The (Physical) Strength of Nations. I don’t disagree with his main overall point; I only disagree that grip strength is a good measure of overall strength—even though it does follow the expected patterns. Racial differences in grip strength exist, as I have covered in the past. Furthermore, there are associations between muscle strength and longevity, with stronger men being more likely to live longer, fuller lives (Ruiz et al, 2008; Volkalis, Haille, and Meisinger, 2015; Garcia-Hermosa, et al, 2018) so, of course, strength training can only be seen as a net positive, especially in regard to living a longer and fuller life. Hand grip strength does have a high correlation with overall strength (Wind et al, 2010; Trosclair et al, 2011). While handgrip strength can tell you a whole lot about your overall health (Lee et al, 2016), of course, there is no better proxy than actually doing the lifts/exercises to ascertain one’s level of strength.
There are replicated genetic associations between explosive, powerful athletic performance, along with even the understanding of the causal mechanisms behind the polymorphisms and their carry-over to power sports. We know that if morphology is right and the individual has the RR ACTN3 genotype, that they will exceed in explosive sports. We know the causal pathways of ACTN3 and how it leads to differences in sprinting competitions. It should be worth noting that, while we do know a lot more about the genomics of sports than we did 20, even 10 years ago, current genetic testing has zero predictive power in regard to talent identification (Pitsladis et al, 2013).
So, of course, for parents and coaches who wonder about the athletic potential of their children and students, the best way to gauge whether or not they will excel in athletics is…to have them compete and compare them to other kids. Even if the genetics aspect of elite power performance is fully unlocked one day (which I doubt it will be), the best way to ascertain whether or not one will excel in a sport is to put them to the test and see what happens. We are in our infancy in understanding the genomics of sporting performance, but when we do understand which genotypes are more prevalent in regard to certain sports (and of course the interactions of the genotype with the environment and genes), then we can better understand how and why others are better in certain sports.
The genomics of elite sporting performance is very interesting; however, the answer that reductionists want to see will not appear: genes are difference makers (Sterelny and Griffith, 1999), not causes, and along with a whole slew of other environmental and mental factors (Lippi, Favaloro, and Guidi 2008), along with a favorable genetic profile with sufficient training (and everything else that comes along with it) are needed for the athlete to reach their maximum athletic potential (see Guth and Roth, 2013). Genetic and environmental differences between individuals and groups most definitely explain differences in elite sporting performance, though elucidating what causes what and the mechanisms that cause the studied trait in question will be tough.
Just because group A has gene or gene networks G and they compete in competition C does not mean that gene or gene networks G contribute in full—or in part—to sporting success. The correlations could be coincidental and non-functional in regard to the sport in question. Athletes should be studied in isolation, meaning just studying a specific athlete in a specific discipline to ascertain how, what, and why works for the specific athlete along with taking anthropomorphic measures, seeing how bad they want “it”, and other environmental factors such as nutrition and training. Looking at the body as a system will take us away from privileging one part over another—while we also do understand that they do play a role but not the role that reductionists believe.
These studies, while they attempt to show us how genetic factors cause differences at the elite level in power sports, they will not tell the whole story, because we must look at the whole system, not reduce it down to the sum of its parts (Shenk, 2011: chapter 5). While blacks are more likely to have these polymorphisms that are associated with elite power athlete performance, this does not obviously carry over to strongman and powerlifting competition.
Blumenbachian Partitions and Mimimalist Races
2100 words
Race in the US is tricky. On one hand, we socially construct races. On the other, these socially constructed races have biological underpinnings. Racial constructivists, though, argue that even though biological races are false, races have come into existence—and continue to exist—due to human culture and human decisions (see the SEP). Sound arguments exist for the existence of biological races. Biological races exist, and they are real. One extremely strong view is from philosopher of science Quayshawn Spencer. In his paper A Radical Solution to the Race Problem, Spencer (2014) argues that biological races are real; that the term “race” directly refers; that race denotes proper names, not kinds; and these sets of human populations denoted by Americans can be denoted as a partition of human populations which Spencer (2014) calls “the Blumenbach partition”.
To begin, Spencer (2014) defines “referent”: “If, by using appropriate evidential methods (e.g., controlled experiments), one finds that a term t has a logically inconsistent set of identifying conditions but a robust extension, then it is appropriate to identify the meaning
of t as just its referent.” What he means is that the word “race” is just a referent, which means that the term “race” lies in what points out in the world. So, what “race” points out in the world becomes clear if we look at how Americans define “race”.
Spencer (2014) assumes that “race” in America is the “national meaning” of race. That is, the US meaning of race is just the referent to the Census definitions of race, since race-talk in America is tied to the US Census. But the US Census Bureau defers to the Office of Management and Budget (OMB). Therefore, since the US Census Bureau defers to the OMB on matters of race, and since Americans defer to the US Census Bureau, then Americans use the OMB definitions of race.
The OMB describes a “comprehensive set” of categories (according to the OMB) which lead Spencer (2014) to believe that the OMB statements on race are pinpointing Caucasians, Africans, Pacific Islanders, East Asians, and Amerindians. Spencer (2014: 1028-29) thusly claims that race in America “is a term that rigidly designates a particular set of “population groups.” Now, of course, the question is this: are these population groups socially constructed? Do they really exist? Are the populations identified arbitrary? Of course, the answer is that they identify a biologically real set of population groups.
To prove the existence of his Blumenbachian populations, Spencer (2014) invokes populational genetic analyses. Population geneticists first must make the assumption at how many local populations exist in the target species. According to Spencer, “The current estimate for humans is 7,105 ethnic groups, half of which are in Africa and New Guinea.” After the assumptions are made, the next step is to sample the species’ estimated local populations. Then they must test noncoding DNA sequences. Finally, they must attempt to partition the sample so that each partition at each level is unique which then minimizes genetic differences in parts and maximizes genetic differences among parts. There are two ways of doing this: using structure and PCA. For the purposes of this argument, Spencer (2014) chooses structure, invoking a 5-population racial model, (see e.g., Rosenberg et al, 2002).
K = 5 corresponds to 5 populational clusters which denote Africans, Oceanians, East Asians, Amerindians, and Caucasians (Spencer, 2014; Hardimon, 2017b). K = 5 shows that the populations in question are genetically structured—that is, meaningfully demarcated on the basis of genetic markers and only genetic markers. Thus, that the populations in question are meaningfully demarcated on the basis of genetic markers, this is evidence that Hardimon’s (2017b) minimalist races are a biological reality. Furthermore, since Rosenberg et al (2002) used microsatellite markers in their analysis, this is a nonarbitrary way of constructing genetic clusters which then demarcate the continental-level minimalist races (Hardimon, 2017b: 90).
Thus, Spencer (2014) argues to call the partition identified in K = 5 “the Blumenbachian partition” in honor of Johann Blumenbach, anthropologist, physician, physiologist, and naturalist. (Though it should be noted that one of his races “Malays” was not a race, but Oceaninans are, so he “roughly discovered” the population partition.) So we can say that “the Blumenbach partition” is just the US meaning of “race”, the partitions identified by K = 5 (Rosenberg et al, 2002).
Furthermore, like Lewontin (1972), Rosenberg et al (2002) found that a majority of human genetic variation is between individuals, not races. That is, Rosenberg et al (2002) found that only 4.3 percent of human genetic variation was found to lie between the continental-level minimalist races. Thus, minimalist races are a biological kind, “if only a modest one” (Hardimon, 2017b: 91). Thus, Rosenberg et al (2002) support the contention that minimalist races exist and are a biological reality since a fraction of human population variation is due to differences among continental-level minimalist races (Africans, Caucasians, East Asians, Oceanians, and Amerindians). The old canard is true, there really is more genetic variation within races than between them, but, as can be seen, that does not rail against the reality of race, since that small amount of genetic variation shows that humanity is meaningfully clustered in a genetic sense.
Spencer (2014: 1032) then argues why Blumenbachian populations are “race” in the American sense:
It is not hard to generate accessible possible worlds that support the claim that US race terms are just aliases for Blumenbachian populations. For example, imagine a possible world τ where human history unfolded exactly how it did in our world except that every Caucasian in τ was killed by an infectious disease in the year 2013. Presumably, we have access to τ, since it violates no logical, metaphysical, or scientific principles. Then, given that we use ‘white’ in its national American meaning in our world, and given that we use ‘Caucasian’ in its Blumenbachian meaning in our world, it is fair to say that both ‘Caucasian’ and ‘white’ are empty terms in τ in 2014—which makes perfect sense if ‘white’ is just an alias for Caucasians. It is counterfactual evidence like this that strongly suggests that the US meaning of ‘race’ is just the Blumenbach partition.
Contrary to critics, this partition is biologically real and demarcates the five genetically structured populations of the human race. Rosenberg et al (2005) found that if sufficient data are used, “the geographic distribution of the sampled individuals has little effect on the analysis“, while their results verify that genetic clusters “arise from genuine features of the underlying pattern of human genetic variation, rather than as artifacts of uneven sampling along continuous gradients of allele frequencies.”
Some may claim that K = 5 is “arbitrary”, however, constructing genetic clusters using microsatellites is nonarbitrary (Hardimon, 2017b: 90):
Constructing genetic clusters using microsatellites constitutes a nonarbitrary way of demarcating the boundaries of continental-level minimalist races. And the fact that it is possible to construct genetic clusters corresponding to continental-level minimalist races in a nonarbitrary way is itself a reason for thinking that minimalist race is biologically real 62.
It should also be noted that Hardimon writes in note 62 (2017b: 197):
Just to be perfectly clear, I don’t think that the results of the 2002 Rosenberg article bear on the question: Do minimalist races exist? That’s a question that has to be answered separately. In my view, the fundamental question in the philosophy of race on which the results of this study bear is whether minimalist race is biologically real. My contention is that they indicate that minimalist race (or more precisely, continental-level minimalist race) is biologically real if sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races.
Sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races, therefore race is a biological reality. We can pinpoint them on the basis of patterns of visible physical features; these visible physical features correspond to geographic ancestry; this satisfies the criteria for minimalist races; therefore race exists. Race exists as a biological kind.
Furthermore, if these five populations that Rosenberg et al (2002) identified (the Blumenbachian populations) are minimalist races, then minimalist race is “a minor principle of human genetic structure” (Hardimon, 2017b: 92). Since minimalist races constitute a dimension within the small amount of human genetic variation that is captured between the continental-level minimalist races (4.3 percent), then it is completely possible to talk meaningfully about the racial structure of human genetic variation which consists of the human genetic variation which corresponds to continental-level minimalist races.
Thus, the US meaning of race is just a referent; the US meaning of race refers to a particular set of human populations; races in the US are classically-defined races (Amerindian, Caucasian, African, East Asian, and Oceanians; the Blumenbach partition); and race is both a biological reality as well as socially constructed. These populations are biologically real; if these populations are biologically real, then it stands to reason that biological racial realism is true (Hardimon, 2012 2013, 2017a; 2017b; Spencer, 2014, 2015).
Human races exist, in a minimalist biological sense, and there are 5 human races. Defenders of Rushton’s work—who believed there are only 3 primary races: Caucasoids, Mongoloids, and Negroids (while Amerindians and others were thrown into the “Mongoloid race” and Pacific Islanders being grouped with the “Negroid race” (Rushton, 1988, 1997; see also Liberman, 2001 for a critique of Rushton’s tri-racial views)—are forced into a tri-racial theory, since he used this tri-racial theory as the basis for his, now defunct, r/K selection theory. The tri-racial theory, that there are three primary races of man—Caucasoid, Mongoloid, and Negroid—has fallen out of favor with anthropologists for decades. But what we can see from new findings in population genetics since the sequencing of the human genome, however, is that human populations cluster into five populations and these five populations are races, therefore biological racial realism is true.
Biological racial realism (the fact that race exists as a biological reality) is true, however, just like with Hardimon’s minimalist races, they do not denote “superiority”, “inferiority” for one race over another. Most importantly, Blumenbachian populations do not denote those terms because the genetic evidence that is used to support the Blumenbachian partition use noncoding DNA. (It should also be noted that the terms “superior” and “inferior” are nonsensical, when used outside of their anatomic contexts. The head is the most superior part of the human body, the feet are the most inferior part of the human body. This is the only time these terms make sense, thus, using the terms outside of this context makes no sense.)
It is worth noting that, while Hardimon’s and Spencer’s views on race are similar, there are some differences between their views. Spencer sees “race” as a referent, while Hardimon argues that race has a set descriptive meaning on the basis of C (1)-(3); (C1) that, as a group, is distinguished from other groups of human beings by patterns of visible physical features, (C2) whose members are linked be a common ancestry peculiar to members of that group, and (C3) that originates from a distinctive geographic location” (Hardimon, 2017b: 31). Whether or not one prefers Blumenbachian partitions or minimalist races depends on whether or not one prefers race in a descriptive sense (i.e., Hardimon’s minimalist races) or if the term race in America is a referent to the US Census discourse, which means that “race” refers to the OMB definitions which then denote Blumenbachian partitions.
Hardimon also takes minimalist races to be a biological kind, while Spencer takes them to be a proper name for a set of population groups. Both of these differing viewpoints regarding race, while similar, are different in that one is describing a kind, while the other describes a proper name for a population group; these two views regarding population genetics from these two philosophers are similar, they are talking about the same things and hold the same deflationary views regarding race. They are talking about how race is seen in everyday life and where people get their definitions of “race” from and how they then integrate it into their everyday lives.
“Race” in America is a proper name for a set of human population groups, the five population groups identified by K = 5. Americans defer to the US Census Bureau on race, who defers to the Office of Management and Budget to define race. They hold that races are a “set”, and these “sets” are Oceanians, Caucasians, East Asians, Amerindians, and Africans. Race, thusly, refers to a set of population groups; “race” is not a “kind”, but a proper name for known populational groups. K = 5 then shows us that the demarcated clusters correspond to continental-level minimalist races, what is termed “the Blumenbach partition.” This partition is “race” in the US sense of the term, and it is a biological reality, therefore, like Hardimon’s minimalist races, the Blumenbach partition identifies what we in America know to be race. (It’s worth noting that, obviously, the Blumenbach partition/minimalist races are one in the same, Spencer is a deflationary realist regarding race, just like Hardimon.)
Defending Minimalist Races: A Response to Joshua Glasgow
2000 words
Michael Hardimon published Rethinking Race: The Case for Deflationary Realism last year (Hardimon, 2017). I was awaiting some critical assessment of the book, and it seems that at the end of March, some criticism finally came. The criticism came from another philosopher, Joshua Glasgow, in the journal Mind (Glasgow, 2018). The article is pretty much just arguing against his minimalist race concept and one thing he brings up in his book, the case of a twin earth and what we would call out-and-out clones of ourselves on this twin earth. Glasgow makes some good points, but I think he is largely misguided on Hardimon’s view of race.
Hardimon (2017) is the latest defense for the existence of race—all the while denying the existence of “racialist races”—that there are differences in mores, “intelligence” etc—and taking the racialist view and “stripping it down to its barebones” and shows that race exists, in a minimal way. This is what Hardimon calls “social constructivism” in the pernicious sense—racialist races, in Hardimon’s eyes, are socially constructed in a pernicious sense, arguing that racialist races do not represent any “facts of the matter” and “supports and legalizes domination” (pg 62). The minimalist concept, on the other hand, does not “support and legalize domination”, nor does it assume that there are differences in “intelligence”, mores and other mental characters; it’s only on the basis of superficial physical features. These superficial physical features are distributed across the globe geographically and these groups are real and exist who show these superficial physical features across the globe. Thus, race, in a minimal sense, exists. However, people like Glasgow have a few things to say about that.
Glasgow (2018) begins by praising Hardimon (2017) for “dispatching racialism” in his first chapter, also claiming that “academic writings have decisively shown why racialism is a bad theory” (pg 2). Hardimon argues that to believe in race, on not need believe what the racialist concept pushes; one must only acknowledge and accept that there are:
1) differences in visible physical features which correspond to geographic ancestry; 2) these differences in visible features which correspond to geographic ancestry are exhibited between real groups; 3) these real groups that exhibit these differences in physical features which correspond to geographic ancestry satisfy the conditions of minimalist race; C) therefore race exists.
This is a simple enough argument, but Glasgow disagrees. As a counter, Glasgow brings up the “twin earth” argument. Imagine a twin earth was created. On Twin Earth, everything is exactly the same; there are copies of you, me, copies of companies, animals, history mirrored down to exact minutiae, etc. The main contention here is that Hardimon claims that ancestry is important for our conception of race. But with the twin earth argument, since everything, down to everything, is the same, then the people who live on twin earth look just like us but! do not share ancestry with us, they look like us (share patterns of visible physical features), so what race would we call them? Glasgow thusly states that “sharing ancestry is not necessary for a group to count as a race” (pg 3). But, clearly, sharing ancestry is important for our conception of race. While the thought experiment is a good one it fails since ancestry is very clearly necessary for a group to count as a race, as Hardimon has argued.
Hardimon (2017: 52) addresses this, writing:
Racial Twin Americans might share our concept of race and deny that races have different geographical origins. This is because they might fail to understand that this is a component of their race concept. If, however, their belief that races do not have different geographical origins did not reflect a misunderstanding of their “race concept,” then their “race concept” would not be the same concept as the concept that is the ordinary race concept in our world. Their use of ‘race’ would pick out a different subject matter entirely from ours.
and on page 45 writes:
Glasgow envisages Racial Twin Earth in such a way that, from an empirical (that is, human) point of view, these groups would have distinctive ancestries, even if they did not have distinctive ancestries an sich. But if this is so, the groups [Racial Twin Earthings] do not provide a good example of races that lack distinctive ancestries and so do not constitute a clear counterexample to C(2) [that members of a race are “linked by a common ancestry peculiar to members of that group”].
C(2) (P2 in the simple argument for the existence of race) is fine, and the objections from Glasgow do not show that P(C)2 is false at all. The Racial Twin Earth argument is a good one, it is sound. However, as Hardimon had already noted in his book, Glasgow’s objection to C(2) does not rebut the fact that races share peculiar ancestry unique to them.
Next, Glasgow criticizes Hardimon’s viewpoints on “Hispanics” and Brazilians. These two groups, says Glasgow, shows that two siblings with the same ancestry, though they have different skin colors, would be different races in Brazil. He uses this example to state that “This suggests that race and ancestry can be disconnected” (pg 4). He criticizes Hardimon’s solution to the problem of race and Brazilians, stating that our term “race” and the term in Brazil do not track the same things. “This is jarring. All that anthropological and sociological work done to compare Brazil with the rest of the world (including the USA) would be premised on a translation error” (pg 4). Since Americans and Brazilians, in Glasgow’s eyes, can have a serious conversation about race, this suggests to Glasgow that “our concept of race must not require that races have distinct ancestral groups” (pg 5).
I did cover Brazilians and “Hispanics” as regards the minimalist race concept. Some argue that the “color system” in Brazil is actually a “racial system” (Guimaraes 2012: 1160). While they do denote race as ‘COR’ (Brazilian for ‘color), one can argue that the term used for ‘color’ is ‘race’ and that we would have no problem discussing ‘race’ with Brazilians, since Brazilians and Americans have similar views on what ‘race’ really is. Hardimon (2017: 49) writes:
On the other hand, it is not clear that the Brazilian concept of COR is altogether independent of the phenomenon we Americans designate using ‘race.’ The color that ‘COR’ picks out is racial skin color. The well-known, widespread preference for lighter (whiter) skin in Brazil is at least arguably a racial preference. It seems likely that white skin color is preferred because of its association with the white race. This provides a reason for thinking that the minimalist concept of race may be lurking in the background of Brazilian thinking about race.
Since ‘COR’ picks out racial skin color, it can be safely argued that Brazilians and Americans at least are generally speaking about the same things. Since the color system in Brazil pretty much mirrors what we know as racial systems, demarcating races on the basis of physical features, we are, it can be argued, talking about the same (or similar) things.
Further, the fact that “Latinos” do not fit into Hardimon’s minimalist race concepts is not a problem with Hardimon’s arguments about race, but is a problem with how “Latinos” see themselves and racialize themselves as a group. “Latinos” can count as a socialrace, but they do not—can not—count as a minimalist race (such as the Caucasian minimalist race; the African minimalist race; the Asian minimalist race etc), since they do not share visible physical patterns which correspond to differences in geographic ancestry. Since they do not exhibit characters that demarcate minimalist races, they are not minimalist races. Looking at Cubans compared to, say, Mexicans (on average) is enough to buttress this point.
Glasgow then argues that there are similar problems when you make the claim “that having a distinct geographical origin is required for a group to be a race” (pg 5). He says that we can create “Twin Trump” and “Twin Clinton” might be created from “whole cloth” on two different continents, but we would still call them both “white.” Glasgow then claims that “I worry that visible trait groups are not biological objects because the lines between them are biologically arbitrary” (pg 5). He argues that we need a “dividing line”, for example, to show that skin color is an arbitrary trait to divide races. But if we look at skin color as an adaptation to the climate of the people in question (Jones et al, 2018), then this trait is not “arbitrary”, and the trait is then linked to geographic ancestry.
Glasgow then goes down the old and tired route that “There is no biological reason to mark out one line as dividing the races rather than another, simply based on visible traits” (pg 5). He then goes on to discuss the fact that Hardimon invokes Rosenberg et al (2002) who show that our genes cluster in specific geographic ancestries and that this is biological evidence for the existence of race. Glasgow brings up two objections to the demarcation of races on both physical appearance and genetic analyses: picture the color spectrum, “Now thicken the orange part, and thin out the light red and yellow parts on either side of orange. You’ve just created an orange ‘cluster’” (pg 6), while asking the question:
Does the fact that there are more bits in the orange part mean that drawing a line somewhere to create the categories orange and yellow now marks a scientifically principled line, whereas it didn’t when all three zones on the spectrum were equally sized?
I admit this is a good question, and that this objection would indeed go with the visible trait of skin color in regard to race; but as I said above, since skin color can be conceptualized as a physical adaptation to climate, then that is a good proxy for geographic ancestry, whether or not there is a “smooth variation” of skin colors as you move away from the equator or not, it is evidence that “races” have biological differences and these differences start on the biggest organ in the human body. This is just the classic continuum fallacy in action: that X and Y are two different parts of an extreme; there is no definable point where X becomes Y, therefore there is no difference between X and Y.
As for Glasgow’s other objection, he writes (pg 6):
if we find a large number of individuals in the band below 62.3 inches, and another large grouping in the band above 68.7 inches, with a thinner population in between, does that mean that we have a biological reason for adopting the categories ‘short’ and ‘tall’?
It really depends on what the average height is in regard to “adopting the categories ‘short’ and ‘tall’” (pg 6). The first question was better than the second, alas, they do not do a good job of objecting to Hardimon’s race concept.
In sum, Glasgow’s (2018) review of Hardimon’s (2017) book Rethinking Race: The Case for Deflationary Realism is an alright review; though Glasgow leaves a lot to be desired and I do think that his critique could have been more strongly argued. Minimalist races do exist and are biologically real.
I am of the opinion that what matters regarding the existence of race is not biological science, i.e., testing to see which populations have which differing allele frequencies etc; what matters is the philosophical aspects to race. The debates in the philosophical literature regarding race are extremely interesting (which I will cover in the future), and are based on racial naturalism and racial eliminativism.
(Racial naturalism “signifies the old, biological conception of race“; racial eliminativism “recommends discarding the concept of race entirely“; racial constructivism “races have come into existence and continue to exist through “human culture and human decisions” (Mallon 2007, 94)“; thin constructivism “depicts race as a grouping of humans according to ancestry and genetically insignificant, “superficial properties that are prototypically linked with race,” such as skin tone, hair color and hair texture (Mallon 2006, 534); and racial skepticism “holds that because racial naturalism is false, races of any type do not exist“.) (Also note that Spencer (2018) critiques Hardimon’s viewpoints in his book as well, which will also be covered in the future, along with the back-and-forth debate in the philosophical literature between Quayshawn Spencer (e.g., 2015) and Adam Hochman (e.g., 2014).)
Race and Vitamin D Deficiency
1600 words
Vitamin D is an important “vitamin” (it is really a steroid hormone). It is produced when the skin (the largest organ in the body) is exposed to the sun’s UVB rays (Nair and Maseeh, 2012). So this is one of the only ways to get natural levels of UVB. We can then think that, if a population is outside of its natural evolutionary habitat (the habitat where that skin color evolved), then we should note numerous problems caused by the lack of vitamin D in whichever population is studied outside of a location that doesn’t get the correct amount of UVB rays from the sun.
Black Americans are more likely than other ethnies to be deficient in vitamin D (Harris, 2006; Cosman et al, 2007; Nair, 2012; Forest and Stuhldreher, 2014; Taksler et al, 2014). But, paradoxically, low vitamin D levels don’t cause weaker bones in black Americans (O’Conner et al, 2014). However, like with all hypotheses, there are naysayers. For example. Powe et al (2013) argue that vitamin D tests misdiagnose blacks, that blacks have a form of the vitamin that cells can use called 25-hydroxyvitamin D. They conclude: “Community-dwelling black Americans, as compared with whites, had low levels of total 25-hydroxyvitamin D and vitamin D–binding protein, resulting in similar concentrations of estimated bioavailable 25-hydroxyvitamin D. Racial differences in the prevalence of common genetic polymorphisms provide a likely explanation for this observation.” Though there are a whole host of problems here.
The limitations of Powe et al (2013) striking: it was cross-sectional and observational (like most nutrition studies) so they were unable to predict effects of vitamin-D binding protein on bone fractures; no data on the consumption of vitamin D supplements; measurement of bone turnover markers, urinary calcium excretion and levels of 1,25-dihydroxyvitamin D may explain the effect of VDBP (vitamin D-binding protein) on mineral metabolism; and they relied on a calculation, rather than a measurement of 25-hydroxyvitamin D levels.
Powe et al’s (2013) findings, though, have been disputed. Using different measurement tools from Powe et al (2013), Henderson et al (2015) conclude that “Counter to prior observations by immunoassay, VDBG concentrations did not vary by race.” While Bouillon (2014) writes: In our view, black Americans, as compared with white Americans, have lower levels of not only total 25-hydroxyvitamin D but also free or bioavailable 25-hydroxyvitamin D.” And finally, Hollis and Bikle (2014) write: “Specifically, for any given physically measured level of bio-available 25-hydroxyvitamin D, the authors are overestimating bio-available 25-hydroxyvitamin D by 2 to 2.5 times owing to underestimation of vitamin D–binding protein in blacks.”
Either way, even if what Powe et al (2013) conclude is true, that would not mean that black Americans should not supplement with vitamin D, since many diseases and health problems are associated with low vitamin D intake in blacks, including osteoporosis, cardiovascular disease, cancer, diabetes, and other serious conditions (Harris, 2006). An indirect relationship between low levels of vitamin D and hypertension is also noted (Mehta and Agarwal, 2017). Since there is an indirect relationship between vitamin D levels and hypertension, then we should keep an eye on this because black Americans have some of the highest levels of hypertension in the world (Ferdinand and Armani, 2007; see also Fuchs, 2011).
Vitamin D is, of course, important for skeletal and nonskeletal health (Kennel et al, 2010). So if vitamin D is important for skeletal and nonskeletal health, we should see more diseases in black Americans that imply a lack of this steroid in the body. Although blacks have stronger bones even when deficient in vitamin D, it is still observed that black children who break their forearms have less vitamin D circulating in their blood (Ryan et al, 2011). This observation is borne out by the data, since black children are more likely to be deficient in vitamin D compared to other ethnies (Moore, Murphy, and Hollick, 2005). Since black skin predicts vitamin D deficiency (Thomas and Demay, 2000), it seems logical to give vitamin D supplements to children, especially black children, on the basis that it would help lower incidences of bone fractures, even though blacks have stronger bones than whites.
Furthermore, physiologically “normal” levels of vitamin D differ in blacks compared to whites (Wright et al, 2012). They showed that it is indeed a strong possibility that both whites and blacks have different levels of optimum vitamin D. Wright et al (2012) showed that there is a relationship between 25(OH)D levels and intact parathyroid hormone (iPth); for blacks, the threshold in which there was no change was 20 ng/ml whereas for whites it was 30 ng/ml which suggests that there are different levels of optimal vitamin D for each race, and the cause is due to skin color. Thus, physiologically “normal” levels of vitamin D differ for blacks and whites.
There is also a high prevalence of vitamin D deficiency/insufficiency and asthma in black inner-city youth in Washington DC (Freishtat et al, 2010). We can clearly see that, even though black Americans have stronger bones than white Americans and vitamin D predicts bone strength, the fact that blacks have stronger bones than whites even while being deficient in vitamin D on average does not mean that black Americans should not supplement with vitamin D, since it would ameliorate many other problems they have that are related to vitamin D deficiency.
There are also racial differences in prostate cancer (PCa) acquisition too, and vitamin D deficiency may also explain this disparity (Khan and Partin, 2004; Bhardwaj et al, 2017). I have heavily criticized the explanations that testosterone influences PCa, while having indicated that environmental factors such as diet and vitamin D deficiency may explain a large amount of the gap (Batai et al, 2017; but see Stranaland et al, 2017 for a contrary view). Since low vitamin D is related to prostate cancer, by supplementing with vitamin D, it is possible that levels of PCa may decrease. Kristal et al (2014) show that both high and low levels of vitamin D are associated with PCa.
Evidence also exists that vitamin D levels and hypertension are related. Rostand (2010) proposes a unified hypothesis: an important role exists in vitamin D deficiency and the pathogenesis and maintenance of hypertension in blacks (Rostand, 2010).
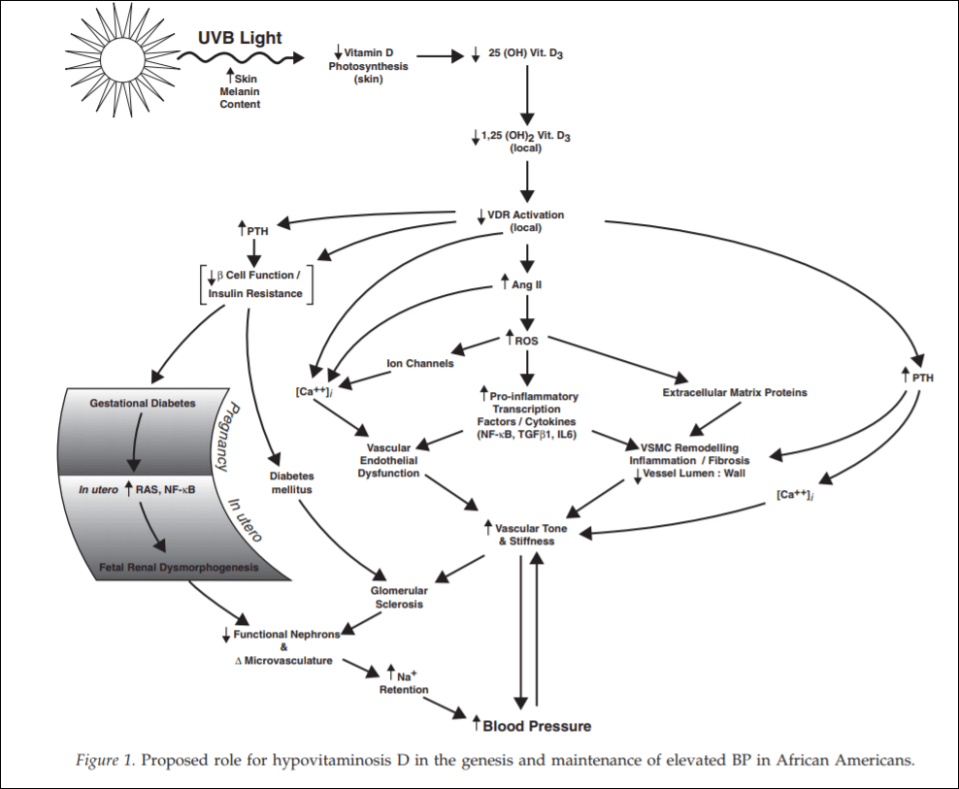
(From Rostand, 2010)
Since black Americans are no longer near the equator, their ability to synthesize vitamin D from UVB rays is diminished. This then probably leads the RAS (renin-angiotensin system) and inflammatory cytokine activation which then leads to vascular endothelial dysfunction along with structural changes to the microvasculature, which have been linked to vascular (arterial) stiffness along with increased vascular resistance, and these changes are shown to precede hypertension, which also occurs early in life. So since blacks are deficient in vitamin D, which even starts in the womb (Bodnar et al, 2007; Dawodu and Wagner, 2007; Lee et al, 2007; Khalessi et al, 2015; Seto et al, 2016), and this vitamin D deficiency most likely produces changes in large and small arteries and arterials, this could be the explanation for higher hypertension in black Americans (Rostand, 2010: 1701).
This would be a large environmental mismatch: since the population is displaced from its ancestral homeland, then this causes problems since it is not the environment where their ancestors evolved. So in this case, since black Americans are concentrated in the southeast corner of the United States, this may explain the high rates of vitamin D deficiency and hypertension in the black American community.
People whose ancestors evolved in locations with fewer UVB rays have lighter skin, whereas people whose ancestors evolved in locations with more UVB rays have darker skin. Thus, by placing populations in their opposite evolutionary environment, we can see how and why deleterious effects would occur in the population that is in the mismatched environment. For whites, skin cancer would occur, whereas for blacks, higher rates of hypertension and low birth weights occur.
Looking at levels of vitamin D deficiency in races is a great way to understand the evolution of certain populations. Because if the vitamin D hypothesis is correct, if skin color is an adaptation to UVB rays, with light skin being an adaptation to low UVB while dark skin is an adaptation to high UVB, then we can safely hypothesize about certain problems that would arise in races that are outside of their natural habitats. We have confirmed these hypotheses—black Americans who are outside of the location that their ancestors evolved in are more likely to have deleterious symptoms, and the symptoms are due to differences in vitamin D production, which come down to differences in skin color and how the skin synthesizes vitamin D in low-light environments.
Even though blacks have stronger bones than whites, this does not mean that they do not experience fractures at a high rate—especially children—and since the association was noticed, then by supplementing with vitamin D, this may lower the disparity of these types of injuries.
Since black Americans, compared to their evolutionary history, live in low-light environments, this then explains the how and why of vitamin D deficiency and why blacks need to supplement with vitamin D; no matter if certain studies show that blacks are ‘healthy’ even though they have low levels of vitamin D. If true (which I strongly doubt), that does not mean that black Americans should not supplement with vitamin D, because numerous other maladies are associated with vitamin D intake. This is one aspect where understanding the evolution of our species and the different races in it would lead to better medical care for individuals and ancestral groups that may need special treatment.
It is clear that race and geography should inform vitamin D intake, for if we do this, many diseases that arise can be ameliorated and quality of life can increase for everyone.
Is Racial Superiority in Sports a Myth? A Response to Kerr (2010)
2750 words
Racial differences in sporting success are undeniable. The races are somewhat stratified in different sports and we can trace the cause of this to differences in genes and where one’s ancestors were born. We can then say that there is a relationship between them since, they have certain traits which their ancestors also had, which then correlate with geographic ancestry, and we can explain how and why certain populations dominate (or would have the capacity to based on body type and physiology) certain sporting events. Critiques of Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It are few and far between, and the few that I am aware of are alright, but this one I will discuss today is not particularly good, because the author makes a lot of claims he could have easily verified himself.
In 2010, Ian Kerr published The Myth of Racial Superiority in Sports, who states that there is a “dark side” to sports, and specifically sets his sights on Jon Entine’s (2000) book Taboo. In this article, Kerr (2010) makes a lot of, in my opinion, giant claims which provide a lot of evidence and arguments in order to show their validity. I will discuss Kerr’s views on race, biology, the “environment”, “genetic determinism”, and racial dominance in sports (which will have a focus on sprinting/distance running in this article).
Race
Since establishing the reality and validity of the concept of race is central to proving Entine’s (2002) argument on racial differences in sports, then I must prove the reality of race (and rebut what Kerr 2010 writes about race). Kerr (2010: 20) writes:
First, it is important to note that Entine is not working in a vacuum; his assertions about race and sports are part of a larger ongoing argument about folk notions of race. Folk notions of race founded on the idea that deep, mutually exclusive biological categories dividing groups of people have scientific and cultural merit. This type of thinking is rooted in the notion that there are underlying, essential differences among people and that those observable physical differences among people are rooted in biology, in genetics (Ossorio, Duster, 2005: 2).
Dividing groups of people does have scientific, cultural and philosophical merit. The concept of “essences” has long been discarded by philosophers. Though there are differences in both anatomy and physiology in people that differ by geographic location, and this then, at the extreme end, would be enough to cause the differences in elite sporting competition that is seen.
Either way, the argument for the existence of race is simple: 1) populations differ in physical attributes (facial, morphological) which then 2) correlate with geographic ancestry. Therefore, race has a biological basis since the physical differences between these populations are biological in nature. Now that we have established that race exists using only physical features, it should be extremely simple to show how Kerr (2010) is in error with his strong claims regarding race and the so-called “mythology” of racial superiority in sports. Race is biological; the biological argument for race is sound (read here and here, and also see Hardimon, 2017).
Genetic determinism
True genetic determinism—as is commonly thought—does not have any sound, logical basis (Resnick and Vorhaus, 2006). So Kerr’s (2010) claims in this section need to be dissected here. This next quote, though, is pretty much imperative to the soundness and validity of his whole article, and let’s just say that it’s easy to rebut and invalidates his whole entire argument:
Vinay Harpalani is one of the most outspoken critics of using genetic determinism to validate notions of inferiority or the superiority of certain groups (in this case Black athletes). He argues that in order for any of Entine’s claims to be valid he must prove that: 1) there is a systematic way to define Black and White populations; 2) consistent and plausible genetic differences between the populations can be demonstrated; 3) a link between those genetic differences and athletic performance can be clearly shown (2004).
This is too easy to prove.
1) While I do agree that the terminology of ‘white’ and ‘black’ are extremely broad, as can be seen by looking at Rosenberg et al (2002), population clusters that cluster with what we call ‘white’ and ‘black’ exist (and are a part of continental-level minimalist races). So is there a systematic way to define ‘Black’ and ‘White’ populations? Yes, there is; genetic testing will show where one’s ancestors came from recently, thereby proving point 1.
2) Consistent and plausible genetic differences between populations can be demonstrated. Sure, there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016). Even these small between-continent/group differences would have huge effects on the tail end of said distribution.
3) I have compiled numerous data on genetic differences between African ethnies and European ethnies and how these genetic differences then cause differences in elite athletic performance. I have shown that Jamaicans, West Africans, Kenyans and Ethiopians (certain subgroups of the two aforementioned countries) have genetic/somatypic differences that then lead to differences in these sporting competitions. So we can say that race can predict traits important for certain athletic competitions.
1) The terminology of ‘White’ and ‘Black’ are broad; but we can still classify individuals along these lines; 2) consistent and plausible genetic differences between races and ethnies do exist; 3) a link between these genetic differences between genes/athletic differences between groups can be found. Therefore Entine’s (2002) arguments—and the validity thereof—are sound.
Kerr (2010) then makes a few comments on the West’s “obsession with superficial physical features such as skin color”, but using Hardimon’s minimalist race concept, skin color is a part of the argument to prove the existence and biological reality of race, therefore skin color is not ‘superficial’, since it is also a tell of where one’s ancestors evolved in the recent past. Kerr (2010: 21) then writes:
Marks writes that Entine is saying one of three things: that the very best Black athletes have an inherent genetic advantage over the very best White athletes; that the average Black athlete has a genetic advantage over the average White athlete; that all Blacks have the genetic potential to be better athletes than all Whites. Clearly these three propositions are both unknowable and scientifically untenable. Marks writes that “the first statement is trivial, the secondly statistically intractable, and the third ridiculous for its racial essentialism” (Marks, 2000: 1077).
The first two, in my opinion (the very best black athletes have an inherent genetic advantage over the very best white athletes and the average black athlete has a genetic advantage over the average white athlete), are true, and I don’t know how you can deny this; especially if you’re talking about AVERAGES. The third statement is ridiculous, because it doesn’t work like that. Kerr (2010), of course, states that race is not a biological reality, but I’ve proven that it is so that statement is a non-factor.
Kerr (2010) then states that “ demonstrating across the board genetic variations between
populations — has in recent years been roundly debunked“, and also says “ Differences in height, skin color, and hair texture are simply the result of climate-related variation.” This is one of the craziest things I’ve read all year! Differences in height would cause differences in elite sporting competition; differences in skin color can be conceptualized as one’s ancestors’ multi-generational adaptation to the climate they evolved in as can hair texture. If only Kerr (2010) knew that this statement here was the beginning of the end of his shitty argument on Entine’s book. Race is a social construct of a biological reality, and there are genetic differences between races—however small (Risch et al, 2002; Tang et al, 2005) but these small differences can mean big differences at the elite level.
The “environment” and biological variability
Kerr (2010) then shifts his focus over to, not genetic differences, but biological differences. He specifically discusses the Kenyans—Kalenjin—stating that “height or weight, which play an instrumental role in helping define an individual’s athletic prowess, have not been proven to be exclusively rooted in biology or genetics.” While estimates of BMI and height are high (both around .8), I think we can disregard the numbers since they came from highly flawed twin studies, since molecular genetic evidence shows lower heritabilities. Either way, surely height is strongly influenced by ‘genes’. Another important caveat is that Kenya has one of the lowest BMIs in the world, 20.7 for Kenyan men, which also is part of the cause of why certain African ethnies dominate running competitions.
I don’t disagree with Kerr (2010) here too much; many papers show that SES/cultural/social factors are very important to Kenyan runners (Onywera et al, 2006; Wilbur and Pistiladis, 2012; Tucker, Onywera, and Santos-Concejero, 2015). You can have all of the ‘physical gifts’ in the world, if it’s not combined with the will to want to do your best, along with cultural and social factors you won’t succeed. But having an advantageous genotype and physique are useless without a strong mind (Lippi, Favaloro, and Guidi, 2008):
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.”
Dissecting this, though, is tougher. Because being born at certain altitudes will cause certain advantageous traits, such as a larger lung capacity (and you will have an advantage in lung capacity when competing at lower altitudes), but certain subpopulations live in these high-altitude areas, so what is it? Genetic? Cultural? Environmental? All three? Nature vs nurture is a false dichotomy; so it is a mixture of the three.
How does one explain, then, the athlete who trains countless hours a day fine-tuning a jump shot, like LeBron James or shaving seconds off sub-four minute miles like Robert Kipkoech Cheruiyot, a four time Boston Marathon winner?
Literally no one denies that elite athletes put in insane amounts of practice; but if everyone has the same amount of practice they won’t have similar abilities.
He also briefly brings up muscle fibers, stating:
These include studies on African fast twitch muscle fibers and development of motor skills. Entine includes these studies to demonstrate irrevocable proof of embedded genetic differences between populations but refuses to accept the fact that any differences may be due to environmental factors or training.
This, again, shows ignorance of the literature. An individual’s muscle fibers are formed during development from the fusion of several myoblasts, with differentiation being completed before birth. Muscle fiber typoing is also set at age 6, no difference in skeletal muscle tissue was found when comparing 6-year-olds and adults, therefore we can state that muscle fiber typing is set by age 6 (Bell et al, 1980). You can, of course, train type II fibers to have similar aerobic capacity to type I fibers, but they’ll never be fully similar. This is something that Kerr (2010) obviously is ignorant to because he’s not well-read on the literature which causes him to make dumb statements like “any differences [in muscle fiber typing] may be due to environmental factors or training“.
Black domination in sports
Finally, Kerr (2010) discusses the fact that whites dominated certain running competitions in the Olympics and that before the 1960s, a majority of distance-running gold medals went to white athletes. He then states that the 2008 Boston Marathon winner was Kenyan; but the next 4 behind him were not. Now, let’s check out the 2017 Marathon winners: Kenya, USA, Japan for the top 3; while 5 Kenyans/Ethiopians are in the top 15 while the same is also true of women; a Kenyan winner, with Kenyans/Ethiopians taking 5 of the top 15 spots. The fact that whites used to do well in running sports is a non-factor; Jesse Owens blew away the competition in the Games in Germany, which showed how blacks would begin to dominate in the US decades later.
Kerr (2010) then ends the article with a ton of wild claims; the wildest one, in my opinion, being that “Kenyans are no more genetically different from any other African or European population on average“, does anyone believe this? Because I have data to the contrary. They have a higher Vo2 max, which of course is trainable but with a ‘genetic’ component (Larsen, 2003), while other authors argue that genetic differences between populations account for differences in success in running competition between populations (Vancini et al, 2014), while male and female Kenyan and Ethiopian runners are the fastest in the half and full marathon (Knechtle et al, 2016). There is a large amount of data out there that speaks about Kenyan/Ethiopian and others’ dominance in running; it seems Kerr (2010) just ignored the data. I agree with Kerr that Kenyanholos show that humans can adapt to their environment; but his conclusion here:
The fact that runners coming from Kenya do so well in running events attests to the fact the combination of intense high altitude training, consumption of a low-fat, high protein diet, and a social and cultural expectation to succeed have created in recent decades an environment which is highly conducive to producing excellent long-distance runners.
is very strong, and while I don’t disagree at all with anything here, he’s disregarding how somatype and genes differ between Kenyans and other populations that compete in these sports that then lead to differences in elite sporting competitions.
Elite sporting performance is influenced by myriad factors, including psychology, ‘environment’, and genetic factors. Something that Kerr (2010) doesn’t understand—because he’s not well-read on this literature—is that many genetic factors that influence sporting performance are known. The ability to become elite depends on one’s capacity for endurance, muscle performance, the ability of the tendons and ligaments to withstand stress and injury, and the attitude to train and push above and beyond what normal people can do (Lippi, Longo, and Maffulli, 2010). We can then extend this to human races; some are better-equipped to excel in running competitions than others.
On its face, Kerr’s (2010) claim that there are no inherent differences between races is wrong. Races differ in somatype, which is due to evolution in different geographic locations for tens of thousands of years. The human body is perfectly adapted to for long distance running (Murray and Costa, 2012), and since our capabilities for endurance running evolved in Africa and they, theoretically, have a musculoskeletal structure similar to the Homo sapiens that left Africa around 70 kya, then it’s only logical to state that African’s, on average, have an inherent ability in running competitions (West and East Africans, while North Africans fare very well in middle distance running, which, again, comes down to living in higher altitudes like Kenyans and Ethiopians).
Wagner and Heyward (2000) reviewed many studies on the physiological differences between blacks and whites. Blacks skew towards mesomorphy; black youths had smaller billiac and bitrochanteric width (the widest measure of the pelvis at the outer edges and the flat process on the femur, respectively), and black infants had longer extremities than white infants (Wagner and Heyward, 2000). We have anatomic evidence that blacks are superior runners (in an American context). Mesomorphic athletes are more likely to be sprinters (Sands et al, 2005; which is also seen in prepubescent children: Marta et al, 2013) Kenyans are ecto-dominant (Vernillo et al, 2013) which helps to explain their success at long-distance running. So just on only looking at the phenotype (a marker for race with geographic ancestry, proving the biological existence of race) we can confidently state, on average just by looking at an individual or a population, how they will fare in certain competitions.
Conclusion
Kerr’s (2010) arguments leave a ton to be desired. Race exists and is a biological reality. I don’t know why this paper got published since it was so full of errors; his arguments were not sound and much of the literature contradicts his claims. What he states at the end about Kenyans is not wrong at all, but to not even bring up genetic/biologic differences as a factor influencing their performance is dishonest.
Of course, a whole slew of factors, be they biological, cultural, psychological, genetic, socioeconomic, anatomic, physiologic etc influence sporting performance, but certain traits are more likely to be found in certain populations, and in the year 2018 we have a good idea of what influences elite sporting performance and what does not. It just so happens that these traits are unevenly distributed between populations, and the cause is evolution in differing climates in differing geographic locations.
Race exists and is a biological reality. Biological anatomic/physiological differences between these races then manifest themselves in elite sporting competition. The races differ, on average, in traits important for success in certain competitions. Therefore, race explains some of the variance in elite sporting competition.
Minimalist Races Exist and are Biologically Real
3050 words
People look different depending on where their ancestors derived from; this is not a controversial statement, and any reasonable person would agree with that assertion. Though what most don’t realize, is that even if you assert that biological races do not exist, but allow for patterns of distinct visible physical features between human populations that then correspond with geographic ancestry, then race—as a biological reality—exists because what denotes the physical characters are biological in nature, and the geographic ancestry corresponds to physical differences between continental groups. These populations, then, can be shown to be real in genetic analyses, and that they correspond to traditional racial groups. So we can then say that Eurasian, East Asian, Oceanian, black African, and East Asians are continental-level minimalist races since they hold all of the criteria needed to be called minimalist races: (1) distinct facial characters; (2) distinct morphologic differences; and (3) they come from a unique geographic location. Therefore minimalist races exist and are a biological reality. (Note: There is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016), but this does not mean that the minimalist biological concept of race has no grounding in biology.)
Minimalist race exists
The concept of minimalist race is simple: people share a peculiar geographic ancestry unique to them, they have peculiar physiognomy (facial features like lips, facial structure, eyes, nose etc), other physical traits (hair/hair color), and a peculiar morphology. Minimalist races exist, and are biologically real since minimalist races can survive findings from population genetics. Hardimon (2017) asks, “Is the minimalist concept of race a social concept?” on page 62. He writes that social concepts are socially constructed in a pernicious sense if and only if it “(i) fails to represent any fact of the matter and (ii) supports and legitimizes domination.” Of course, populations who derive from Africa, Europe, and East Asia have peculiar facial morphology/morphology unique to that isolated population. Therefore we can say that minimalist race does not conform to criteria (i). Hardimon (2017: 63) then writes:
Because it lacks the nasty features that make the racialist concept of race well suited to support and legalize domination, the minimalist race concept fails to satisfy condition (ii). The racialist concept, on the other hand, is socially constructed in the pernicious sense. Since there are no racialist races, there are no facts of the matter it represents. So it satisfies (i). To elaborate, the racialist race concept legtizamizes racial domination by representing the social hierarchy of race as “natural” (in a value-conferring sense): as the “natural” (socially unmediated and inevitable) expression of the talent and efforts of the inidividuals who stand on its rungs. It supports racial domination by conveying the idea that no alternative arrangment of social institutions could possibly result in racial equality and hence that attempts to engage in collective action in the hopes of ending the social hierarchy of race are futile. For these reasons the racialist race concept is also idealogical in the prejorative sense.
Knowing what we know about minimalist races (they have distinct physiognomy, distinct morphology and geographic ancestry unique to that population), we can say that this is a biological phenomenon, since what makes minimalist races distinct from one another (skin color, hair color etc) are based on biological factors. We can say that brown skin, kinky hair and full lips, with sub-Saharan African ancestry, is African, while pale/light skin, straight/wavy/curly hair with thin lips, a narrow nose, and European ancestry makes the individual European.
These physical features between the races correspond to differences in geographic ancestry, and since they differ between the races on average, they are biological in nature and therefore it can be said that race is a biological phenomenon. Skin color, nose shape, hair type, morphology etc are all biological. So knowing that there is a biological basis to these physical differences between populations, we can say that minimalist races are biological, therefore we can use the term minimalist biological phenomenon of race, and it exists because there are differences in the patterns of visible physical features between human populations that correspond to geographic ancestry.
Hardimon then talks about how eliminativist philosophers and others don’t deny that above premises above the minimalist biological phenomenon of race, but they allow these to exist. Hardimon (2017: 68-69) then quotes a few prominent people who profess that there are, of course, differences in physical features between human populations:
… Lewontin … who denies that biological races exist, freely grants that “peoples who have occupied major geographic areas for much of the recent past look different from one another. Sub-Saharan Africans have dark skin and people who have lived in East Asia tend to have a light tan skin and an eye color and eye shape that is difference from Europeans.” Similarly, population geneticist Marcus W. Feldman (final author of Rosenberg et al., “Genetic Stucture of Human Populations” [2002]), who also denies the existence of biological races, acknowledges that “it has been known for centuries that certain physical features of humans are concentrated within families: hair, eye, and skin color, height, inability to digest milk, curliness of hair, and so on. These phenotypes also show obvious variation among people from different continents. Indeed, skin color, facial shape, and hair are examples of phenotypes whose variation among populations from different regions is noticeable.” In the same vein, eliminative anthropologist C. Loring Brace concedes, “It is perfectly true that long term residents of various parts of the world have patterns of features that we can identify as characteristic of they area from which they come.”
So even these people who claim to not believe in “biological races”, do indeed believe in biological races because what they are describing is biological in nature and they, of course, do not deny that people look different while their ancestors came from different places so therefore they believe in biological races. We can then use the minimalist biological phenomenon of race to get to the existence of minimalist races.
Hardimon (2017: 69) writes:
Step 1. Recognize that there are differences in patterns of visible physical features of human beings that correspond to their differences in geographic ancestry.
Step 2. Observe that these patterns are exhibited by groups (that is, real existing groups).
Step 3. Note that the groups that exhibit these patterns of visible physical features correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race.
Step 4. Infer that minimalist race exists.
Those individuals mentioned previously who deny biological races but allow that people with ancestors from differing geographic locales look differently do not disagree with step 1, nor does anyone really disagree with step 2. Step 4’s inference immediately flows from the premise in step 3. “Groups that exhibit patterns or visible physical features that correspond to differences in geographical ancestry satisfy the conditions of the minimalist concept of race. Call (1)-(4) the argument from the minimalist biological phenomenon of race” (Hardimon, 2017: 70). Of course, the argument does not identify which populations may be called races (see further below), it just shows that race is a biological reality. Because if minimalist races exist, then races exist because minimalist races are races. Minimalist races exist, therefore biological races exist. Of course, no one doubts that people come from Europe, sub-Saharan Africa, East Asia, the Americas, and the Pacific Islands, even though the boundaries between them are ‘blurry’. They exhibit patterns of visible physical characters that correspond to their differing geographic ancestry, they are minimalist races therefore minimalist races exist.
Pretty much, the minimalist concept of race is just laying out what everyone knows and arguing for its existence. Minimalist races exist, but are they biologically real?
Minimalist races are biologically real
Of course, some who would assert that minimalist races do not exist would say that there are no ‘genes’ that are exclusive to one certain population—call them ‘race genes’. Of course, these types of genes do not exist. Whether or not one individual is a part of one race or not does not rest on the basis of his physical characters, but is determined by who his parents are, because one of the three premises for the minimalist race argument is ‘must have a peculiar geographic ancestry’. So it’s not that members of races share sets of genes that other races do not, it’s based on the fact that they share a distinctive set of visible physical features that then correspond with geographic ancestry. So of course if the minimalist concept of race is a biological concept then it entails more than ‘genes for’ races.
Of course, there is a biological significance to the existence of minimalist biological races. Consider that one of the physical characters that differ between populations is skin color. Skin color is controlled by genes (about half a dozen within and a dozen between populations). Lack of UV rays for individuals with dark skin will lead to diseases like prostate cancer, while darker skin is a protectant against UV damage to human skin (Brenner and Hearing, 2008; Jablonksi and Chaplin, 2010). Since minimalist race is biologically significant and minimalist races are partly defined by differences in skin color between populations then skin color has both medical and ecological significance.
(1) Consider light skin. People with light skin are more susceptible to skin cancer since they evolved in locations with poor UVR radiation (D’Orazio et al, 2013). The body needs vitamin D to absorb and use calcium for maintaining proper cell functioning. People who evolved near the equator don’t have to worry about this because the doses of UVB they absorb are sufficient for the production of enough previtamin D. While East Asians and Europeans on the other hand, became adapted to low-sunlight locations and therefore over time evolved lighter skin. This loss of pigmentation allowed for better UVB absorption in these new environments. (Also read my article on the evolution of human skin variation and also how skin color is not a ‘tell’ of aggression in humans.)
(2) While darker-skinned people have a lower rate of skin cancer “primarily a result of photo-protection provided by increased epidermal melanin, which filters twice as much ultraviolet (UV) radiation as does that in the epidermis of Caucasians” (Bradford, 2009). Dark skin is thought to have evolved to protect against skin cancer (Greaves, 2014a) but this has been contested (Jablonski and Chaplin, 2014) and defended (Greaves, 2014b). So therefore, using (1) and (2), skin color has evolutionary signifigance.
So as humans began becoming physically adapted to their new niches they found themselves in, they developed new features distinct from the location they previously came from to better cope with the new lifestyle due to their new environments. For instance “Northern Europeans tend to have light skin because they belong to a morphologically marked ancestral group—a minimalist race—that was subject to one set of environmental conditions (low UVR) in Europe” (Hardimon, 2017: 81). Of course explaining how human beings survived in new locations falls into the realm of biology, while minimalist races can explain why this happened.
Minimalist races clearly exist since minimalist races constitute complex biological patterns between populations. Hardimon (2017: 83) writes:
It [minimalist race] also enjoys intrinsic scientific interest because it represents distinctive salient systematic dimension of human biological diversity. To clarify: Minimalist race counts as (i) salient because human differences of color and shape are striking. Racial differences in color and shape are (ii) systematic in that they correspond to differences in geographic ancestry. They are not random. Racial differences are (iii) distinctive in that they are different from the sort of biological differences associated with the other two salient systematic dimensions of human diversity: sex and age.
[…]
An additional consideration: Like sex and age, minimalist race constitutes one member of what might be called “the triumverate of human biodiversity.” An account of human biodiversity that failed to include any one of these three elements would be obviously incomplete. Minimalist race’s claim to be biologically real is as good as the claim of the other members of the triumverate. Sex is biologically real. Age is biologically real. Minimalist race is biologically real.
Real does not mean deep. Compared to the biological associated with sex (sex as contrasted with gender), the biological differences associated with minimalist race are superficial.
Of course, the five ‘clusters’ and ‘populations’ identified by Rosenberg et al’s (2002) K=5 graph, which told structure to produce 5 genetic clusters, corresponds to Eurasia, Africa, East Asia, Oceania, and the Americas, are great candidates for minimalist biological races since they correspond to geographic locations, and even corroborates what Fredrich Blumenbach said about human races back in the 17th century. Hardimon further writes (pg 85-86):
If the five populations corresponding to the major areas are continental-level minimalist races, the clusters represent continental-level minimalist races: The cluster in the mostly orange segment represents the sub-Saharan African continental-level minimalist race. The cluster in the mostly blue segment represents the Eurasian continental-level minimal race. The cluster in the mostly pink segment represents the East Asian continental-level minimalist race. The cluster in the mostly green segment represents the Pacific Islander continental-level minimalist race. And the cluster in the mostly purple segment represents the American continental-level minimalist race.
[…]
The assumption that the five populations are continental-level minimalist races entitles us to interpret structure as having the capacity to assign individuals to continental-level minimalist races on the basis of markers that track ancestry. In constructing clusters corresponding to the five continental-level minimalist races on the basis of objective, race-neutral genetic markers, structure essentially “reconstructs” those races on the basis of a race-blind procedure. Modulo our assumption, the article shows that it is possible to assign individuals to continental-level races without knowing anything about the race or ancestry of the individuals from whose genotypes the microsattelites are drawn. The populations studied were “defined by geography, language, and culture,” not skin color or “race.”
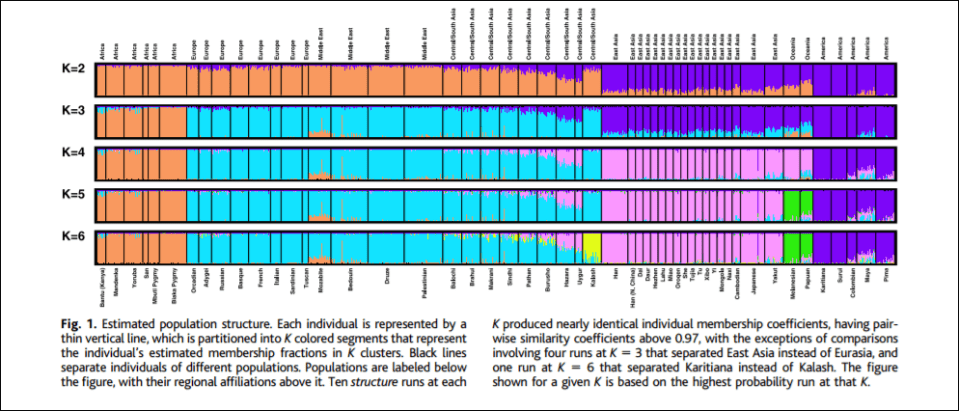
Of course, as critics note, the researchers predetermine how many populations that structure demarcates, for instance, K=5 indicates that the researchers told the program to delineate 5 clusters. Though, these objections do not matter. For the 5 populations that come out in K=5 “are genetically structured … which is to say, meaningfully demarcated solely on the basis of genetic markers” (Hardimon, 2017: 88). K=6 brings one more population, the Kalash, a group from northern Pakistan who speak an Indo-European language. Though “The fact that structure represents a population as genetically distinct does not entail that the population is a race. Nor is the idea that populations corresponding to the five major geographic areas are minimalist races undercut by the fact that structure picks out the Kalash as a genetically distinct group. Like the K=5 graph, the K=6 graph shows that modulo our assumption, continental-level races are genetically structured” (Hardimon, 2017: 88).
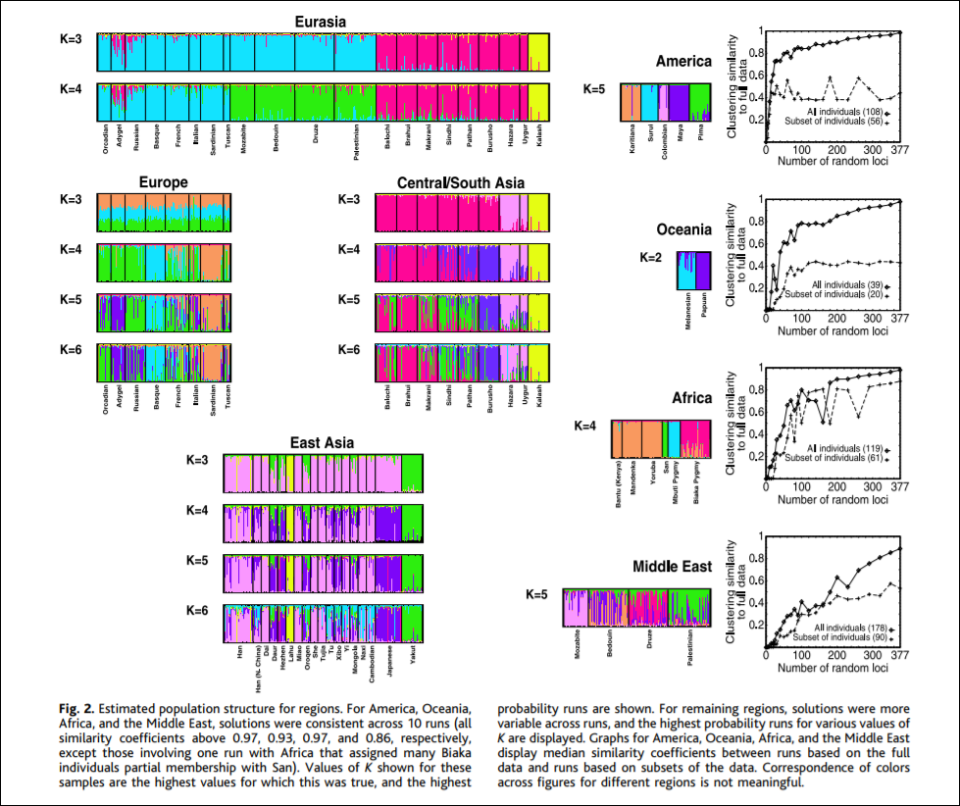
Though of course there are naysayers. Svante Paabo and David Serre, Hardimon writes, state that when individuals are sampled from homogeneous populations from around the world, the gradients of the allele frequencies that are found are distributed randomly across the world rather than clustering discretely. Though Rosenberg et al responded by verifying that the clusters they found are not artifacts of sampling as Paabo and Serre imply, but reflect features of underlying human variation. Though Rosenberg et al agree with Paabo and Serre in that that human genetic diversity consists of clines in variation in allele frequencies (Hardimon, 2017: 89). Other naysayers also state that all Rosenberg et al show is what we can “see with our eyes”. Though a computer does not partition individuals into different populations based on something that can be done with eyes, it’s based on an algorithm.
Hardimon also accepts that black Africans, Caucasians, East Asians, American Indians and Oceanians can be said to be races in the basic sense because “they constitute a partition of the human species“, and that they are distinguishable “at the level of the gene” (Hardimon, 2017: 93). And of course, K=5 shows that the 5 races are genetically distinguishable.
Hardimon finally discusses some medical significance for minimalist races. He states that if you are Caucasian that it is more likely that you have a polymorphism that protects against HIV compared to a member of another race. Meanwhile, East Asians are more likely to carry alleles that make them more susceptible to Steven-Johnson syndrome or another syndrome where their skin falls off. Though of course, the instances where this would matter in a biomedical context are rare, but still should be at the back of everyone’s mind (as I have argued), even though instances where medical differences between minimalist races are rare, there are times where one’s race can be medically significant.
Hardimon finally states that this type of “metaphysics of biological race” can be called “deflationary realism.” Deflationary because it “consists in the repudiation of the ideas that racialist races exist and that race enjoys the kind of biological reality that racialist race was supposed to have” and realism which “consists in its acknowledgement of the existence of minimalist races and the genetically grounded, relatively superficial, but still significant biological reality of minimalist race” (Hardimon, 2017: 95-96).
Conclusion
Minimalist races exist. Minimalist races are a biological reality because distinct visible patterns show differences between geographically isolated populations. This is enough for the classification of the five classic races we know of to be called race, be biologically real, and have a medical significance—however small—because certain biological/physical traits are tied to different geographic populations—minimalist races.
Hardimon (2017: 97) shows an alternative to racialism:
Deflationary realism provides a worked-out alternative to racialism—it it a theory that represents race as a genetically grounded, relatively superficial biological reality that is not normatively important in itself. Deflationary realism makes it possible to rethink race. It offers the promise of freeing ourselves, if only imperfectly, from the racialist background conception of race.
It is clear that minimalist races exist and are biologically real. You do not need to speak about supposed mental traits between these minimalist races, they are irrelevant to the existence of these minimalist biological races. As Hardimon (2017: 67) writes: “No reference is made to normatively important features such as intelligence, sexuality, or morality. No reference is made to essences. The idea of sharp boundaries between patterns of visible physical features or corresponding geographical regions is not invoked. Nor again is reference made to the idea of significant genetic differences. No reference is made to groups that exhibit patterns of visible physical features that correspond to geographic ancestry.”
The minimalist biological concept of race stands up to numerous lines of argumentation, therefore we can say without a shadow of a doubt that minimalist biological race exists and is real.
