
Home » Biology
Category Archives: Biology
A Priori and Empirical Arguments for Multiple Realizability
2500 words
What is the multiple realizability argument?
The multiple realizability argument (MRA) is an argument directed at type-identity theories of mind, while being used for and against functionalist theories of mind. (I think Ross’ 1992 argument in Immaterial Aspects of Thought and Feser’s 2013 arguments refute functionalist theories.) First formulated by Hilary Putnam in 1975, the argument he formulated can be put like this:
P1. If type-physicalism is true, then every mental property can be realized in exactly one physical way.
P2. It is empirically highly plausible that mental properties are capable of multiple realizations.
C1. It is (empirically) highly plausible that the view of type-physicalism is false (modus tollens, P1, P2). (From Just the Arguments, 81. Putnam’s Multiple Realization Argument against Type-Physicalism)
P1 states the scope of type-physicalism—that all mental states are realizable in one and only one physical way. P2 states that it is probable that mental properties are capable of multiple realizations. This premise is an empirical one, and so we need evidence to believe it. Then, the conclusion is that type-physicalism is false sine mental states are multiply realizable. Quite obviously, this argument shows that mental states don’t reduce to brain states, which means that physicalism is false. Since P2 needs defense, I will defend it in this article while giving my own formulation of the MRA.
Here is my formulation of it:
P1: If mental properties were identical to physical properties, then any change in one entails a change in the other.
P2: Mental properties can change without any corresponding change in physical properties.
C: So mental properties aren’t identical to physical properties.
As you can see, like Putnam’s formulation, P2 is an empirical claim and so needs empirical support. So if one mental state can be realized in multiple ways, then type-physicalism (mind-brain identity) is false. I will spend the rest of this article arguing for the truth of P2 and then provide an argument from analogy showing that mental states are multiply realizable.
An a priori argument for multiple realizability
If the MRA were true, then there would be evidence of a specific mental state that is realized in multiple physical ways. While empirical evidence is irrelevant to metaphysical possibility (and to concepts), multiple realizability can be known a priori. Before I give the empirical argument from analogy for multiple realizability, I will give the a priori argument.
P1: Mental states and processes exhibit certain characteristic features and properties like intentionality, subjectivity, and causality.
P2: If mental states are multiply realizable, then they are not reducible to their underlying physical properties.
C: Thus, mental states and processes are not reducible to their underlying physical properties.
Or
P1: M has properties P1, P2, P3…
P2: If M is multiply realizable, then M is not reducible to it’s physical properties.
C: Therefore, M is irreducible to its physical properties.
Premise 1: Mental states and processes are characterized by what they do rather than what they’re “made of.” Intentionality is the ability for mental states to be “about” things, while directed at objects, events or states of affairs like when a belief or proposition is about a certain end goal. So M properties aren’t reducible to any P properties, and intentionality is a property of mental states which set them apart from physical states, since purely physical things can never have the ability to intend. Subjectivity refers to the fact that mental states are experienced through a first-personal perspective which can’t be observed or measured by others. This property sets M states apart from P states, since physical states can be studied and observed from a third-personal perspective. So while we can study brain states, since mental states don’t reduce to brain physiology, then by studying brain states we aren’t studying the mind. Lastly the property of causality refers to the fact that mental states and processes have causal effects on action and behavior, cognition and other mental states and processes. So the distinctive role that mental states and processes play in generating action, behavior, and cognition cannot be captured by studying the brain or the body.
Premise 2: This premise highlights the fact that multiple realizability implies that mental states can be realized in a multitude of physical states and processes without any loss of mental properties. So the Conclusion follows that mental states and processes are irreducible to their underlying physical properties.
So if mental states and processes have characteristic features that distinguish them from other kinds of states and processes, and if they can be realized by multiple physical systems, then they cannot be reduced to one physical system.
Defending P2: Empirical evidence for multiple realizability
The way that Putnam and I have formulated the argument for MR is an empirical claim. So empirical claims require empirical evidence. While the previous argument was a priori, it could therefore be argued without empirical evidence.
The example of visual perception. Most animals on earth have vision. The visual systems of animals have evolved to help them survive in their ecologies. Humans have three cone types in their eyes which allows them to see a range of colors. On the other hand, some birds have four cone types which allow them to see a wider range of color than humans, and the fourth cone thsg birds have allows them to see more colors than humans (Stoddard et al, 2020). Bats have evolved eyesight that allows them to see in low light environments, while eagles have evolved eyesight that allows them to see objects at great distances. So despite differences in the visual systems between animals, they can all recognize objects and visually navigate their ecologies. So different animals have evolved different vision systems that help in a certain niche. Furthermore, different types of photoreceptors have evolved in different animals, with different connections between the eye and the brain, which began evolving around 600 million years ago, with the Cambrian explosion leading to body plans and systems which then supported vision (Lamb, Pugh Jr, and Collin, 2008; Lamb, Collin, and Pugh Jr, 2011; Asteriti, 2015). These photoreceptors come in two kinds—ciliary type (c-type) or rhabdomeric type (r-type); vertebrates seek to have a unique mix of these cone types which allow a wide range of vision (Marshedian and Fain, 2017). Certain eye structures have also evolved independently (Land and Nilsson, 2012). So different animals have different numbers of photoreceptors and cones, which help them to visualize their environment; the diversity of rods and photoreceptors in the animal kingdom is vast (Piechl, 2005). Thus, the evidence cited here shows that different animals have differently-evolved visual systems, but they can still visualize their environments even though the physical systems that allow it are different.
The brain’s ability to compensate from injury and the brain’s of athletes and musicians. After a traumatic brain injury occurs, the brain—being plastic—can rewire itself to carry different functions after an injury. For example, in the blind, the visual cortex is repurposed and processes tactile and auditory stimuli (Elbert et al, 2002; Lane et al, 2015; Gori et al, 2019). The primary motor cortex in musicians is larger than non-musicians, and this is due to constant practice on their instrument of choice (Toyka and Freund, 2006; Watson, 2006; Olszewska et al, 2021). Basketball players have larger cortical areas associated with visual processing and attention (Kim et al, 2022) along with athletes having different cortical neuronal networks than novices (Tan et al, 2017). This then is solid evidence for the claim that learning new skills and continously performing them at an expert level leads to changes in the brain (Park et al, 2015). Further, when it comes to the brain’s ability to heal from an injury, it has been shown that if a certain brain area is impacted, other parts of the brain will pick up the slack of the injured part, which shows the plasticity of the brain and the brain’s ability to compensate for an injury to it by directing and making new neural pathways to carry out new tasks (Nishimura et al, 2009; Su, Veeravuga, and Grant, 2016; Hylin, Kerr, and Holden, 2017). Thus, the evidence cited here shows how the brain can adapt to tasks that a person performs, and how it can adapt to changes to it (like injury) and even repurposing certain parts of itself in people with certain disabilities. This, like the example of visual perception, lends further support for the claim that different physical systems can perform the same mental function.
Brain-computer interfaces. Lastly, we have brain-computer interfaces. These interfaces “acquire brain signals, analyze them, and translate them into commands that are relayed to output devices that carry out desired actions” (Shih et al, 2012). These interfaces allow humans to control things with their thoughts, bypassing the need for physical interfaces; this technology also allows individuals to control certain kinds of devices using brain waves using their mental intentions (Mak and Wolpaw, 2010). People with these interfaces can control prosthetic limbs (Mischenko et al, 2017; Murphy et al, 2017; Asanza et al, 2022). These interfaces have also been explored to give people the ability to communicate with speech again (Brumberg et al, 2010). This also supports MR since brain-computer interfaces which use EEG to record brain activity could translate mental states into movements while interfaces that use implanted electrodes may allow an individual to control a robotic arm. Thus, the development of this technology shows that different mental states can be realized in different physical systems which is then dependent on the type of interface used.
Strappini et al (2020) provide yet more empirical support for MR. They write:
Here, we illustrate some cases that provide empirical evidence in support of MRT. Recently, it has been proposed that foveal agnosic vision, like peripheral vision, can be restored by increasing object parts’ spacing (Crutch and Warrington, 2007; Strappini et al., 2017b). Agnosic fovea and normal periphery are both limited by crowding, which impairs object recognition, and provides the signature of visual integration. Here, we define a psychological property of restored object identification, and we cross-reference the data of visually impaired patients with different etiologies. In particular, we compare the data of two stroke patients, two patients with posterior cortical atrophy, six cases of strabismic amblyopia, and one case with restored sight. We also compare these patients with unimpaired subjects tested in the periphery. We show that integration (i.e., restored recognition) seems to describe quite accurately the visual performance in all these cases. Whereas the patients have different etiologies and different neural correlates, the unimpaired subjects have no neural damage. Thus, similarity in the psychological property given the differences in the neural substrate can be interpreted in relation to MRT and provide evidence in its support
While Booth (2018: 143-144) uses the example of multilingualism to support MR:
First, there are multiple ways of speaking a second-language, based on difference between high proficiency early and late bilinguals. Second, there are multiple ways of being a speaker of a given language, specifically as a monolingual or bilingual speaker of that language, where the language is the bilingual speaker’s L1. These examples meet the conditions advanced by Polger and Shapiro for examples of multiple realization, and should therefore be accepted as genuine cases of multiple realization.
Now that I have given a good overview of the evidence in support of MR, I will not provide the argument.
The empirical argument from analogy for multiple realizability
P1: Different animals have evolved different vision systems to suite their ecologies.
P2: Humans have a trichromatic visual system while some birds have tetrachromatic visual system while some insects have compound eyes.
P3: Despite differences in these visual systems, these animals all are able to perform similar visual tasks, like spatial navigation and object recognition.
P4: Studies of brain damage and neuroplasticity show that different brain regions can take on different functions after injury or training, like blind people using the visual cortex for auditory processing, muscisians having larger motor areas for finger control, and basketball players having larger cortical areas associated with visual processing and attention.
P5: The development of brain-computer interfaces show that mental states can be translated into different forms of output, like movement, speech, and text, using different physical devices.
C: Thus, multiple realizability is true, since the mental state of visual perception (and other mental states) can be realized in different physical systems without affecting functioning.
P1: If mental states can only be realized in a single physical system, then all animals with similar cognitive tasks should have identical neural structures.
P2: Different animals have different neural structures for performing similar cognitive tasks, like visual perception.
C: Thus, mental states cannot be realized in a single physical system.
P1: If mental states can only be realized in a single physical system, then all animals with similar cognitive tasks should have identical neural structures.
P2: If all animal with similar cognitive tasks have identical neural structures, then different animals should not have different neural structures for performing similar cognitive tasks.
P3: Different animals have different neural structures for performing similar cognitive tasks, like spatial navigation, object recognition and visual perception.
C: Thus, mental states cannot be realized by a single physical system.
Conclusion
I have provided both an a priori and a posteriori argument for the MRA. The a priori argument shows that multiple realizability is metaphysically possible, while the empirical evidence I have cited along with the empirical premises in my arguments have shown that multiple realizability is an empirically defensible position. It seems to me that it is intuitive that different mental states can be realized by different physical systems and not only one kind of physical system.
The a priori argument shows that mental states and physical states have different properties; physical states cannot have the properties that mental states have. So this shows that it’s metaphysically possible the MR is true, while the empirical evidence and arguments I have mounted show that it is true in our world as well. Mental states can’t be reduced to physical states, mental states are causally efficacious, and there are multiple ways to achieve the same cognitive function, like visual perception across the animal kingdom. The example of visual perception of different animals, studies of athletes, musicians, and people with traumatic brain injuries, and even brain-computer interfaces show that different mental states can be realized in multiple physical ways.
So if this is true, then multiple realizability is true. If multiple realizability is true, then type-physicalism is false, and therefore identity theories of mind need to find another avenue to prove their thesis. Mind-brain identity is clearly false; mind doesn’t reduce to brain and mental states can be realized by different physical systems. This is yet another argument against physicalism—the attempted reduction of mind to brain. Physicalism is quite clearly false.
The Answer to Hereditarianism is Developmental Systems Theory
4150 words
Introduction
It is claimed that genes (DNA sequences) have a special, privileged role in the development of all traits. But once we understand what genes do and their role in development, then we will understand that the role ascribed to genes by gene-selectionists and hereditarians outright fails. Indeed, the whole “nature vs nurture” debate implies that genes determine traits and that it’s possible to partition the relative contributions to traits in a genetic and environmental way. This, however, is far from reality (like heritability estimates).
DST isn’t a traditional scientific theory—it is more a theoretical perspective on developmental biology, heredity, and evolution, though it does make some general predictions (Griffiths and Hochman, 2015). But aspects of it have been used to generate novel predictions in accordance with the extended evolutionary synthesis (Laland et al, 2015).
Wilson (2018: 65) notes six themes of DST:
Joint determination by multiple causes
Development is a process of multiple interacting sources.
Context sensitivity and contingency
Development depends on the current state of the organism.
Extended inheritance
An organism inherits resources from the environment in addition to genes.
Development as a process of construction
The organism helps shape its own environment, such as the way a beaver builds a dam to raise the water level to build a lodge.
Distributed control
Idea that no single source of influence has central control over an organism’s development.
Evolution as construction
The evolution of an entire developmental system, including whole ecosystems of which organisms are parts, not just the changes of a particular being or population.
Genes (DNA sequences) as resources and outcomes
Hereditarians have a reductionist view of genes and what they do. Genes, to the hereditarian, are causes of not only development but of traits and evolution, too. However the hereditarian is sorely mistaken—there is no a priori justification for treating genes as privileged causes over and above other developmental resources (Noble, 2012). I take Noble’s argument there to mean that strong causal parity is true—where causal parity means that all developmental resources are on par with each other, with no other resource having primacy over another. They all need to “dance in tune” with the “music of life” to produce the phenotype, to borrow Noble’s (2006, 2017) analogy. Hereditarian dogma also has its basis in the neo-Darwinian Modern Synthesis. The modern synthesis has gotten causality in biology wrong. Genes are, simply put, passive, not active, causes:
Genes, as DNA sequences, do not of course form selves in any ordinary sense. The DNA molecule on its own does absolutely nothing since it reacts biochemically only to triggering signals. It cannot even initiate its own transcription or replication. … It would therefore be more correct to say that genes are not active causes; they are, rather, caused to give their information by and to the system that activates them. The only kind of causation that can be attributed to them is passive, much in the way a computer program reads and uses databases. (Noble, 2011)
These ideas, of course, are also against the claim that genes are blueprints or recipes, as Plomin (2018) claims in his most recent book (Joseph, 2022). This implies that they are context-independent; we have known for years that genes are massively context-sensitive. The line of argument that hereditarians push is that genes are context-insensitive, that is they’re context-independent. But since DNA is but one of the developmental resources the physiological system uses to create the phenotype, this claim fails. Genes are not causes on their own.
Behavioral geneticist and evolutionary psychologist J. P. Rushton (1997: 64) claims that a study shows that “genes are like blueprints or recipes providing a template for propelling development forward to some targeted endpoint.” That is, Rushton is saying that there is context-independent “information” in genes, and that genes, in essence, guide development toward a targeted endpoint. Noah Carl (2019) claims that the hereditarian hypothesis “states that these differences [in cognitive ability] are partly or substantially explained by genetics.” When he says the differences are “partly or substantially explained by genetics”, he’s talking about “cognitive ability” being caused by genes. The claim that genes cause (either partly or substantially) cognitive ability—and all traits, for that matter—fails and it fails since genes don’t do what hereditarians think they do. (Nevermind the conceptual reasons.) These claims are laughable, due to what Noble, Oyama, Moore and Jablonka and Lamb have argued. It is outright false that genes are like blueprints or recipes. Rushton’s is reductionist in a sociobiology-type way, while Plomin’s is reductionist in a behavioral genetic type way.
In The Dependent Gene, David Moore (2002: 81) talks about the context-dependency of genes:
Such contextual dependence renders untenable the simplistic belief that there are coherent, long-lived entities called “genes” that dictate instructions to cellular machinery that merely constructs the body accordingly. The common belief that genes contain context-independent “information”—and so are analogous to “blueprints” or “recipes”—is simply false.
Genes are always expressed in context and cannot be divorced from said context, like hereditarians attempt using heritability analyses. Phenotypes aren’t “in the genes”, they aren’t innate. They develop through the lifespan (Blumberg, 2018).
Causal parity and hereditarianism
Hereditarianism can be said to be a form of genetic reductionism (and mind-brain identity). The main idea of reductionism is to reduce the whole to the sum of its parts and then analyze those parts. Humans (the whole) are made up of genes (the parts), so to understand human behavior, and humans as a whole, we must then understand genes, so the story goes.
Cofnas (2020) makes several claims regarding the hereditarian hypothesis and genes:
But if we find that many of the same SNPs predict intelligence in different racial groups, a risky prediction made by the hereditarian hypothesis will have passed a crucial test.
…
But if work on the genetics and neuroscience of intelligence becomes sufficiently advanced, it may soon become possible to give a convincing causal account of how specific SNPs affect brain structures that underlie intelligence (Haier, 2017). If we can give a biological account of how genes with different distributions lead to race differences, this would essentially constitute proof of hereditarianism. As of now, there is nothing that would indicate that it is particularly unlikely that race differences will turn out to have a substantial genetic component. If this possibility cannot be ruled out scientifically, we must face the ethical question of whether we ought to pursue the truth, whatever it may be.
Haier is a reductionist of not only the gene variety but the neuro variety—he attempts to reduce “intelligence” to genes and neurology (brain physiology). I have though strongly criticized the use of fMRI neuroimaging studies regarding IQ; cognitive localizations in the brain are untenable (Uttal, 2001, 2011) and this is because mind-brain identity is false.
Cofnas asks “How can we disentangle the effects of genes and environment?” and states the the behavioral geneticist has two ways—correlations between twins and adoptees and GWAS. Unfortunately for Cofnas, twin and adoption studies show no such thing (see Ho, 2013), most importantly because the EEA is false (Joseph, 2022a, b). GWAS studies are also fatally confounded (Janssens and Joyner, 2019) and PGS doesn’t show what behavioral geneticists need it to show (Richardson, 2017, 2022). The concept of “heritability” is also a bunk notion (Moore and Shenk, 2016). (Also see below for further discussion on heritability.) At the end of the day, we can’t do what the hereditarian needs to be done for their explanations to hold any water. And this is even before we look at the causal parity between genes and other developmental resources. Quite obviously, the hereditarian hypothesis is a gene-centered view, and it is of course a reductionist view. And since it is a reductionist, gene-centered view, it is then false.
Genetic, epigenetic, and environmental factors operate as a system to form the phenotype. Since this is true, therefore, both genetic and epigenetic determinism is false (also see Wagoner and Uller, 2015). It’s false because the genes one is born with, or develops with, don’t dictate or determine anything, especially not academic achievement as hereditarian gene-hunters would so gleefully claim. And one’s early experience need not dictate an expected outcome, since development is a continuous process. Although, that does not mean that environmental maladies that one experiences during childhood won’t have lasting effects into adulthood due to possibly affecting their psychology, anatomy or physiology.
The genome is responsive, that is, it is inert before it is activated by the physiological system. When we put DNA in a petri dish, it does nothing. It does nothing because DNA cannot be said to be a separate replicator from the cell (Noble, 2018). So genes don’t do anything independent of the context they’re in; they do what they do DUE TO the context they’re in. This is like Gottlieb’s (2007) probabilistic epigenesis, where the development of an organism is due to the coaction of irreducible bidirectional biological and environmental influences. David S. Moore, in The Developing Genome: An Introduction to Behavioral Epigenetics states this succinctly:
Genes—that is, DNA segments—are always influenced by their contexts, so there is never a perfect relationship between the presence of a gene and the ultimate appearance of a phenotype. Genes do not determine who we become, because nongenetic factors play critical roles in trait development; genes do what they do at least in part because of their contexts.
What he means by “critical roles in trait development” is clear if one understands Developmental Systems Theory (DST). DST was formulated by Susan Oyama (1985) in her landmark book “The Ontogeny of Information. In the book, she argues that nature and nurture are not antagonistic to each other, they are cooperative in shaping the development of organisms. Genes do not play a unique informational role in development. Thus, nature vs. nurture is a false dichotomy—it’s nature interacting with nurture, or GxE. This interactionism between nature and nurture—genes and environment—is a direct refutation of hereditarianism. What matters is context, and the context is never independent from what is going on during development. Genes aren’t the units of selection, the developmental system is, as Oyama explains in Evolution’s Eye:
If one must have a “unit” of evolution, it would be the interactive developmental system: life cycles of organisms in their niches. Evolution would then be change in the constitution and distribution of these systems (Oyama, 2000b)
Genes are important, of course, for the construction of the organism—but so are other resources. Without genes, there would be nothing for the cell to read to initiate transcription. However, without the cellular environment, we wouldn’t have DNA. Lewontin puts this wonderfully in the introduction to the 2000 edition of Ontogeny:
There are no “gene actions” outside environments, and no “environmental actions” can occur in the absence of genes. The very status of environment as a contributing cause to the nature of an organism depends on the existence of a developing organism. Without organisms there may be a physical world, but there are no environments. In like manner no organisms exist in the abstract without environments, although there may be naked DNA molecules lying in the dust. Organisms are the nexus of external circumstances and DNA molecules that make these physical circumstances into causes of development in the first place. They become causes only at their nexus, and they cannot exist as causes except in their simultaneous action. That is the essence of Oyama’s claim that information comes into existence only in the process of Ontogeny. (2000, 15-16)
Genes aren’t causes on their own, they are resources for development. And being resources for development, they have no privileged level of causation over other developmental resources, such as “methylation patterns, membrane templates, cytoplasmic gradients, centrioles, nests, parental care, habitats, and cultures” (Griffiths and Stotz, 2018). All of these things, and more of course, need to work in concert with each other.
Indeed, this is the causal parity argument—the claim that genes aren’t special developmental resources, that they are “on par” with other developmental resources (Griffiths and Gray, 1994; Griffiths and Stotz, 2018). Gene knockout studies show that the loss of a gene can be compensated by other genes—which is known as “genetic compensation.” None of the developmental resources play a more determinative role than other resources (Noble, 2012; Gamma and Liebrenz, 2019). This causal parity, then, has implications for thinking about trait ontogeny.
The causal parity of genes and other developmental factors also implies that genes cannot constitute sufficient causal routes to traits, let alone provide complete explanations of traits. Full-blown explanations will integrate various kinds of causes across different levels of organizational hierarchy, and across the divide between the internal and the external. The impossibly broad categories of nature vs. nurture that captured the imagination of our intellectual ancestors a century ago are no longer fit for the science of today. (Gamma and Liebrenz, 2019)
Oyama (2000a 40) articulates the casual parity thesis like this:
What I am arguing for here is a view of causality that gives formative weight to all operative influences, since none is alone sufficient for the phenomenon or for any of its properties, and since variation in any or many of them may or may not bring about variation in the result, depending on the configuration of the whole.
While Griffiths and Hochman (2015) formulate it like this:
The ‘parity thesis’ is the claim that if some role is alleged to be unique to nucleic acids and to justify relegating nongenetic factors to a secondary role in explaining development, it will turn out on closer examination that this role is not unique to nucleic acids, but can be played by other factors.
Genes are necessary pre-conditions for trait development, just as the other developmental resources are necessary pre-conditions for trait development. No humans without genes—this means that genes are necessary pre-conditions. If genes then humans—this implies that genes are sufficient for human life, but they are but one part of what makes humans human, when all of the interactants are present, then the phenotype can be constructed. So all of the developmental resources interacting are sufficient.
The nature vs. nurture dichotomy can be construed in such a way that they are competing explanations. However, we now know that the dichotomy is a false one and that the third way—interactionism—is how we should understand development. Despite hereditarian protestations, DST/interactionism refutes their claims. The “information” in the genes, then, cannot explain how organisms are made, since information is constructed dialectically between the resources and the system. There are a multiplicity of causal factors that are involved in this process, and genes can’t be privileged in this process. Thus the phrase “genetic causation” isn’t a coherent concept. Moreover, DNA sequences aren’t even coherent outside of cellular context (Noble, 2008).
Griffiths and Stotz (2018) put the parity argument like this:
In The Ontogeny of Information Oyama pioneered the parity argument, or the ‘parity thesis’, concerning genetic and environmental causes in development (see also Griffiths and Gray 1994; Griffiths and Gray 2005; Griffiths and Knight 1998; Stotz 2006; Stotz and Allen 2012). Oyama relentlessly tracked down failures of parity of reasoning in earlier theorists. The same feature is accorded great significance when a gene exhibits it, only to be ignored when a non-genetic factor exhibits it. When a feature thought to explain the unique importance of genetic causes in development is found to be more widely distributed across developmental causes, it is discarded and another feature is substituted. Griffiths and Gray (1994) argued in this spirit against the idea that genes are the sole or even the main source of information in development. Other ideas associated with ‘parity’ are that the study of development does not turn on a single distinction between two classes of developmental resources, and that the distinctions useful for understanding development do not all map neatly onto the distinction between genetic and non-genetic.
Shea (2011) tries to argue that genes do have a special role, and that is to transport information. Genes are, of course, inherited, but so is every other part of the system (resources). Claiming that there is information “in the genes” is tantamount to saying that there is a special role for DNA in development. But, as I hope will be clear, this claim fails due to the nature of DNA and its role in development.
This line of argument leads to one clear conclusion—genes are followers, they are not leaders; most evolution begins with environmentally-mediated phenotypic change, and then genetic changes occur (West-Eberhard, 2003). Ho and Saunders (1979) state that variation in organisms is constructed during development due to an interaction between genetic and non-genetic factors. That is, they follow what is needed to do by the developmental system, they aren’t leading development, they are but one party in the whole symphony of development. Development can be said to be irreducible, so we cannot reduce development to genes or anything else, as all interactants need to be present for development to be carried out. Since genes are activated by other factors, it is incoherent to talk of “genetic causes.” Genes affect the phenotype only when they are expressed, and other resources, too, affect the phenotype this is, ultimately, an argument genes against as blueprints, codes, recipes, or any other kind of flowery language one can used to impute what amounts to intention to inert DNA.
Even though epigenetics invalidates all genetic reductionism (Lerner and Overton, 2017), genetic reductionist ideas still persist. They give three reasons why genetic reductionist ideas still persist despite the conceptual, methodological, and empirical refutations. (1) Use of terms like “mechanism”, “trait”, and “interaction”; (2) constantly shifting to other genes once their purported “genes for” traits didn’t workout; and (3) they “buried opponents under repetitive results” (Panofsky, quoted in Lerner and Overton, 2017). The fact of the matter is, there are so many lines of evidence and argument that refute hereditarian claims that it is clear the only reason why one would still be a hereditarian in this day and age is that they’re ignorant—that is racist.
Genes, that is, are servants, not masters, of the development of form and individual differences. Genes do serve as templates for proteins: but not under their own direction. And, as entirely passive strings of chemicals, it is logically impossible for them to initiate and steer development in any sense. (Richardson, 2016)
DST and hereditarian behavioral genetics
I would say that DST challenges three claims from hereditarian behavioral genetics (HBG hereafter):
(1) The claim that we can neatly apportion genes and environment into different causes for the ontogeny of traits;
(2) Genes are the only thing that are inherited and that genes are the unit of selection and a unique—that is, special and privileged cause over and above other resources;
(3) That genes vs environment, blank skate vs human nature, are a valid dichotomy.
(1) HBG needs to rely on the attempting to portion out causes of traits into gene and environmental causes. The heritability statistic presumes additivity, thy is, it assumes no interaction. This is patently false. Charney (2016) gives the example of schizophrenia—it is claimed that 50 percent of the heritability of schizophrenia is accounted for by 8000 genes, which means that each SNP accounts for 1/8000 of the half of the heritability. This claim is clearly false, as genetics aren’t additive, and the additivity assumption precludes the interaction of genes with genes, and environment, which create new interactive environments. Biological systems are not additive, they’re interactive. Heritability estimates, therefore, are attempts at dichotomizing what is not dichitomizable (Rose, 2005).
An approach that partitions variance into independent main effects will never resolve the debate because, by definition, it has no choice but to perpetuate it. (Goldhaber, 2012)
This approach, of course, is the approach that attempts to partition variance into G and E components. The assumption is that G and E are additive. But as DST theorists have argued for almost 40 years, they are not additive, they are interactive and so not additive, therefore heritability estimates fail on conceptual grounds (as well as many others). Heritability estimates have been—and continue to today—been at the heart of the continuance of the nature vs nurture distinction, the battle, if you will. But if we accept Oyama’s causal parity argument—and due to the reality of how genes work in the system, I see no reason why we shouldn’t—then we should reject hereditarianism. Hereditarians have no choice but to continue the false dichotomy of nature vs nurture. Their “field” depends on it. But despite the fact that the main tool for the behavioral geneticist lies on false pretenses (twin and adoption studies), they still try to show that heritability estimates are valid in explaining trait variation (Segalowitz, 1999; Taylor, 2006, 2010).
(2) More than genes are inherited. Jablonka and Lamb (2005) argue that there are four dimensions—interactants—to evolution: genetic, epigenetic, behavioral, and symbolic. They show the context-dependency of the genome, meaning that genotype does not determine phenotype. What does determine the phenotype, as can be seen from the discussion here, is the interacting of developmental resources in development. Clearly, there are many other inheritance systems other than genes. There is also the fact that the gene as popularly conceived does not exist—so it should be the end of the gene as we know it.
(3) Lastly, DST throws out the false dichotomy of genes and environment, nature and nurture. DST—in all of its forms—rejects the outright false dichotomy of nature vs nurture. They are not in a battle with each other, attempting to decide who is to be the determining factor in trait ontogeny. They interact, and this interaction is irreducible. So we can’t reduce development to genes or environment (Moore, 2016) Development isn’t predetermined, it’s probabilistic. The stability of phenotypic form isn’t found in the genes (Moore and Lickliter, 2023)
Conclusion
Genes are outcomes, not causes, of evolution and they are not causes of trait ontogeny on their own. The reality is that strong causal parity is true, so genes cannot be regarded as a special developmental resource from other resources—that is, genes are not privileged resources. Since they are not privileged resources, we need to, then, dispense with any and all concepts of development that champion genes as being the leader of the developmental process. The system is, not genes, with genes being but one of many of the interactants that shape phenotypic development.
By relying on the false narrative that genes are causes and that they cause not only our traits but our psychological traits and what we deem “good” and “bad”, we would then be trading social justice for hereditarianism (genetic reductionism).
These recommended uses of bad science reinforce fears of institutionalized racism in America and further the societal marginalization of minority groups; these implications of their recommendations are never publicly considered by those who promulgate these flawed extensions of counterfactual genetic reductionism. (Lerner, 2021)
Such [disastrous societal] applications can only rob people of life chances and destroy social justice. Because developmental science has the knowledge base to change the life course trajectories of people who are often the targets of genetic reductionist ideas, all that remains to eradicate genetic reductionism from scientific discussion is to have sufficient numbers of developmental scientists willing to proclaim loudly and convincingly that the naked truth is that the “emperor” (of genetic reductionism) has no clothes. (Lerner, 2021: 338)
Clearly, hereditarians need the nature vs nurture debate to continue so they can push their misunderstandings about genes ans psychology. However, given our richer understanding of genes and how they work, we now know that hereditarianism is untenable, and DST conceptions of the gene and development as a whole have led us to that conclusion. Lerner (2017) stated that as soon as the failure of one version of genetic reductionism is observed, another one pious up—making it like a game of whack-a-mole.
The cure to hereditarian genetic reductionism is a relational developmental systems (RDS) model. This model has its origins with Uri Bronfenbrenner’s ecological systems theory (Bronfenbrenner and Ceci, 1994; Ceci, 1996; Patel, 2011; Rosa and Tudge, 2013. Development is about the interacting and relation between the individual and environment, and this is where RDS theory comes in. Biology, physiology, culture, and history are studied to explain human development (Lerner, 2021). Hereditarian ideas cannot give us anything like what models derived from developmental systems ideas can. An organism-environment view can lead to a more fruitful, and the organism and environment are inseparable (Jarvilehto, 1998; Griffiths and Gray, 2002). And it is for these reasons, including many, many more, that hereditarian genetic reductionist ideas should become mere sand in the wind.
Having said all that, here’s the argument:
P1: If hereditarianism is true, then strong causal parity is false.
P2: Strong causal parity is true.
C: Therefore hereditarianism must be false.
Not Feeling Pain: What is CIPA (Congenital Insensitivity to Pain with Anhydrosis)?
1750 words
“Congenital Insensitivity to Pain” (CIPA, or congenital analgesia: CIPA hereafter) is an autosomal recessive disease (Indo, 2002) and was first observed in 1932 (Daneshjou, Jafarieh, and Raeeskarami, 2012). It is called a “congenital disorder” since it is present from birth. Since the disease is autosomal recessive, the closer the two parents are in relatedness, the more likely it is they will pass on a recessive disorder since they are more likely to have and pass on autosomal recessive mutations (Hamamy, 2012). First cousins, for example, 1.7-2.8% higher risk of having a child with an autosomal recessive disease (Teeuw et al, 2013). Consanguinity is common in North Africa (Anwar, Khyatti, and Hemminki, 2014) and the Bedouin have a high rate of this disease (Schulman et al, 2001; Lopez-Cortez et al, 2020; Singer et al, 2020). Three mutations in the TrkA (AKA NTRK1) have been shown to induce protein mis-folding which affect the function of the protein. Different mutations in the TrkA gene have been shown to have be associated with different disease outcomes (Franco et al, 2016). Since the mutated gene in question is needed for nerve growth factors, the pain signals cannot be transferred to the brain since there are hardly any of them there (Shin et al, 2016).
Individuals unfortunate enough to be inflicted with CIPA cannot feel pain. Whether it’s biting their tongues, feeling pain from extreme temperatures. People with CIPA have said that while they can feel the difference between extreme temperatures—hot and cold—they cannot feel the pain that is actually associated with the temperatures on their skin see (Schon et al, 2018). When they bump into things, they may not be aware of what happened and injuries may occur which heal incorrectly due to no medical attention and only noticing the fractures and other things that occur due to CIPA years later after they see doctors for what is possibly factors due to having the disease. People with CIPA are thought to be “dumb” because they constantly bump into things. But what is really happening is that, since they cannot feel pain, they have not learned that bumping into things could be damaging to their bodies, as pain is obviously an experience-dependent event. So these people learn, throughout their lives, to fake being in pain as to not draw suspicion to people who may not be aware of the condition. Children with the disease are thought, most of the time, to be victims of child abuse, but when it is discovered that the child who is thought to be a victim of abuse is inflicted with CIPA (van den Bosch et al, 2014; Amroh et al, 2020), treatments shift toward managing the disease.
About twenty percent of people with CIPA live until three years of age (Lear, 2011), while 20 percent of those who die at age 3 die from complications due to hyperpexia (an elevated body temperature over 106. degrees Fahrenheit) (Rosemberg, Marie, and Kliemann, 1994; Schulmann et al, 2001; Indo, 2002; Nabyev et al, 2018). Since they cannot feel the heat and get themselves to cool down, Due to a low life expectancy (many more live until about 25 years of age), this disease is really hard to study (Inoyue, 2007; Daneshjou, Jafarieh, and Raeeskarami, 2012). People hardly make it past that age since they either don’t feel the pain and do things that normal people, through experience, know not to do since we can feel pain and know to not do things that cause us pain and discomfort or they commit suicide since they have no quality of life due to damaged joints. Furthermore, since they cannot feel pain, people with this disease are more likely to self-mutilate since they cannot learn that self-mutilation causes pain (since pain is a deterrent for future action that may in fact cause pain to an individual). They also cannot sweat, meaning that control of the body temperature of one afflicted with CIPA is of utmost precedence (since they could overheat and die). Thus, these cases of deaths of individuals with CIPA do not occur due to CIPA per se, they occur due to, say, not feeling heat and then sweating while not attempting to regulate their body temperature and cool down (whether by naturally sweating due to being too hot or getting out of the extreme hot temperature causing the elevated body temperature). This is known as “hyperpyrexia” and this cause of death affects around 20 percent of CIPA patients (Sasnur, Sasnur, and Ghaus-ul, 2011). Furthermore, they are more likely to have thick, leathery skin and also show little muscular definition.
Not sweating is associated with CIPA and if one cannot sweat, one cannot have their body temperature regulated when they get too hot. So if they get too hot they cannot feel it and they will die of heat stroke. The disease, though, is rare, as only 17-60 people in America currently have it, while there are about 600 cases of the disease worldwide (Inoyue, 2007; Lear, 2011). This disease is quite hard to identify, but clinicians may be able to detect the presence of the disease through the following ways: Infants biting their lips, fingers, cheeks and not crying or showing any instance of being in pain after the event; repeated fractures in older children; a history of burns with no medical attention; observing that a child has many healed joint injuries and bone fractures without the child’s parents seeking medical care; observing that the patient does not react to hot or cold events (though they can say they can feel a difference between the two) they make errors in distinguishing in whether something is hot or cold (Indo, 2008).
Children who have this disease are at a higher risk of having certain kinds of bodily deformations, since they cannot feel the pain that would make them be hesitant to perform a certain action in the future. Due to this, people with this disease must constantly check themselves for cuts, abrasions, broken bones, etc to ensure that they cannot feel when they actually occur to them. They don’t cry, or show any discomfort, when experiencing what should be an event that would cause someone without CIPA to cry. CIPA-afflicted individuals are more likely to have bodily deformations since their joints and bones do not heal correctly after injury. This then leads to their walking and appearance to be affected. This is one of many reasons why the parents of people with CIPA must constantly check their children for signs of bodily harm or unintentional injuries. One thing that needs to be looked out for is what is termed Charcot joint—which is a degenerative joint disorder (Gucev et al, 2020).
A specific form of CIPA—called HSAN-IV—was discovered in a village in southern Finland called Vittangi, where it was traced to the founder of the village itself in the 1600s. Since the village was remote with such a small population, this meant that the only people around to marry and have children with were people who were closely related to each other. This, then, is the reason why this village in Finland has a high rate of people afflicted with this disease (Norberg, 2006; Minde, 2006). This, again, goes back to the above on consanguinity and autosomal recessive diseases—since CIPA is an autosomal recessive disease, one would reason that we would find it in populations that marry close relatives, either due to custom or population density.
Many features have been noted as showing that an individual is afflicted with CIPA: absent pain sensation from birth, the inability to sweat; and mental retardation, lower height and weight for their age (Safari, Khaledi, and Vojdani, 2011; Perez-Lopez et al, 2015). Children with CIPA have lower IQs than children without CIPA, so there is an inverse relationship between IQ and age; the older the age of the child with CIPA, the lower their IQ, while the reverse is true for individuals who are younger (Erez et al, 2010). One girl, for example. had a WISC-III IQ of 49, and she self-mutilated herself by picking at her nails until they were no longer there (Zafeirou et al, 2004). Another girl with CIPA was seen to have an IQ of 52, be afflicted with mental retardation, have a low birth weight, and was microcephalic (Nolano et al, 2000). Others were noted to have IQs in the normal range (Daneshjou, Jafarieh, and Raaeskarami, 2012). People with a specific form of this disease (HSN type II) were observed to have IQs in the normal range (though it is “caused by” a different set of genes than CIPA, HSN type IV; Kouvelas and Terzoglou, 1989). However, it has been noted that the cut-off of 70 for mental retardation is arbitrary (see Arvidsson and Granlund, 2016). While running a full gamut of tests on an individual thought to have CIPA, we can better attempt to ensure a higher quality of life in individuals afflicted with the disease. In sum, IQ scores of CIPA individuals do not reflect that the mutations in TrkA “cause” IQ scores; it is an outcome of a disrupted system (in this case, mutations on the TrkA gene).
There is currently no cure for this disease, and so, the only way to manage complications stemming from CIPA is to work on the injuries that occur to the joints that occur as they happen, to ensure that the individual has a good quality of life. Treatment for CIPA, therefore, is not actually curing the disease, but it is curing what occurs due to the disease (bone breaks, joint destruction), which would then heighten the quality of life of the person with CIPA (Nabiyev, Kara, and Aksoy, 2016). Naloxone may temporarily relieve CIPA (Rose et al, 2018), while others suggest treatments such as remifentanil (Takeuchi et al, 2018). We can treat outcomes that arise from the disease (like self-mutilation), but we cannot outright cure the disease itself (Daneshjou, Jafarieh, and Raaeskarami, 2012). The current best way to manage the disease is to identify the disease early in children and to do full-body scans of afflicted individuals to attempt to cure the by-products of the disease (such as limb/joint damage and other injuries). Maybe one day we can use gene therapy to help the afflicted, but for now, the best way forward is early identification along with frequent check-ups. By managing body temperature, having frequent check-ups, modifying the behavior of the child as to avoid injuries, wearing a mouth guard so they do not grind their teeth or bite their tongue, avoiding hot or cold environments or food, (Indo, 2008; Rose et al, 2018).
CIPA is a very rare—and very interesting—disease. By better understanding its aetiology, we can better help the extremely low number of people in the world who suffer from this disease.
Gould, Bradbury, and Evolutionary Contingency
1800 words
I have been an avid reader and interested in astronomy/space ever since I could remember. I remember really loving Stephen Hawking and his documentaries on black holes. I would read anything I could find on constellations and stars. From there I went on to reading sci-fi. I then recall seeing The Martian Chronicles by Ray Bradbury and from then on I had become interested in sci-fi writing. But, as I grew older, I drifted away from sci-fi and now only read non-fiction. Then when I got older I got into ‘HBD’ (chronicled here) and along with it evolution—but, unlike other ‘HBDers’ I became enamored with the work of Gould, while some of my favorite books come from him. Gould wrote a lot about evolutionary contingency—the degree to which an outcome could be different. Evolutionary contingency is a big topic in philosophy of biology, and Bradbury has a great short story on this type of contingency.
Ray Bradbury is an interesting author—one who has many short stories and regular books. One of my favorite stories from Bradbury is one called A Sound of Thunder which chronicled a time machine company who let people go back in time to hunt any animal they’d like—if you want to take down the ancestor of a whale before it became aquatic, just name the place and they will send you there. They were told to only stay on the path laid out by the time machine company—animals they could shoot were marked with red paint, presumably those animals would have died anyway so killing them would not change any outcomes. The text from Bradbury is worth quoting in full, as it wonderfully captures the thought of evolutionary contingency:
He indicated a metal path that struck off into green wilderness, over streaming swamp, among giant ferns and palms. “And that,” he said, “is the Path, laid by Time Safari for your use, It floats six inches above the earth. Doesn’t touch so much as one grass blade, flower, or tree. It’s an anti-gravity metal. Its purpose is to keep you from touching this world of the past in any way. Stay on the Path. Don’t go off it. I repeat. Don’t go off. For any reason! If you fall off, there’s a penalty. And don’t shoot any animal we don’t okay.”
“Why?” asked Eckels.
They sat in the ancient wilderness. Far birds’ cries blew on a wind, and the smell of tar and an old salt sea, moist grasses, and flowers the color of blood.
“We don’t want to change the Future. We don’t belong here in the Past. The government doesn’t like us here. We have to pay big graft to keep our franchise. A Time Machine is finicky business. Not knowing it, we might kill an important animal, a small bird, a roach, a flower even, thus destroying an important link in a growing species.”
“That’s not clear,” said Eckels.
“All right,” Travis continued, “say we accidentally kill one mouse here. That means all the future families of this one particular mouse are destroyed, right?”
“Right”
“And all the families of the families of the families of that one mouse! With a stamp of your foot, you annihilate first one, then a dozen, then a thousand, a million, a billion possible mice!”
“So they’re dead,” said Eckels. “So what?”
“So what?” Travis snorted quietly. “Well, what about the foxes that’ll need those mice to survive? For want of ten mice, a fox dies. For want of ten foxes a lion starves. For want of a lion, all manner of insects, vultures, infinite billions of life forms are thrown into chaos and destruction. Eventually it all boils down to this: fifty-nine million years later, a caveman, one of a dozen on the entire world, goes hunting wild boar or saber-toothed tiger for food. But you, friend, have stepped on all the tigers in that region. By stepping on one single mouse. So the caveman starves. And the caveman, please note, is not just any expendable man, no! He is an entire future nation. From his loins would have sprung ten sons. From their loins one hundred sons, and thus onward to a civilization. Destroy this one man, and you destroy a race, a people, an entire history of life. It is comparable to slaying some of Adam’s grandchildren. The stomp of your foot, on one mouse, could start an earthquake, the effects of which could shake our earth and destinies down through Time, to their very foundations. With the death of that one caveman, a billion others yet unborn are throttled in the womb. Perhaps Rome never rises on its seven hills. Perhaps Europe is forever a dark forest, and only Asia waxes healthy and teeming. Step on a mouse and you crush the Pyramids. Step on a mouse and you leave your print, like a Grand Canyon, across Eternity. Queen Elizabeth might never be born, Washington might not cross the Delaware, there might never be a United States at all. So be careful. Stay on the Path. Never step off!”
“I see,” said Eckels. “Then it wouldn’t pay for us even to touch the grass?”
“Correct. Crushing certain plants could add up infinitesimally. A little error here would multiply in sixty million years, all out of proportion. Of course maybe our theory is wrong. Maybe Time can’t be changed by us. Or maybe it can be changed only in little subtle ways. A dead mouse here makes an insect imbalance there, a population disproportion later, a bad harvest further on, a depression, mass starvation, and finally, a change in social temperament in far-flung countries. Something much more subtle, like that. Perhaps only a soft breath, a whisper, a hair, pollen on the air, such a slight, slight change that unless you looked close you wouldn’t see it. Who knows? Who really can say he knows? We don’t know. We’re guessing. But until we do know for certain whether our messing around in Time can make a big roar or a little rustle in history, we’re being careful. This Machine, this Path, your clothing and bodies, were sterilized, as you know, before the journey. We wear these oxygen helmets so we can’t introduce our bacteria into an ancient atmosphere.”
This passage from Bradbury wonderfully illustrates evolutionary—historical—contingency. Things could have been different—this is the basis of the contingency argument. The universe does not repeat itself—if we were to replay the tape of life we would get a completely different outcome—Lane (2015) states maybe octopi would rule the earth? We could replay the tape of life, have it go exactly as it did to lead up to today, change ONE SEEMINGLY MINISCULE THING (say, stepping on a bug that did not die) which would then cascade throughout history leading to a change in the future. Evolution is full of passive trends, with no indication that—for example with body plans—that there is a drive to become more complex—it is passive (Gould, 1996: 207):
All the tests provide evidence for a passive trend and no drive to complexity. McShea found twenty-four cases of significant increases or decreases in comparing the range of modern descendants with an ancestor (out of a potential sample of ninety comparisons, or five groups of mammals, each with six variables measured in each of three ways; for the other comparison, average descendants did not differ significantly from ancestors). Interestingly, thirteen of these significant changes led to decreases in complexity, while only nine showed an increase. (The difference between thirteen and nine is not statistically significant, but I am wryly amused, given all traditional expectation in the other direction, that more comparisons show increasing rather than decreasing complexity.
Gould first put forth his contingency argument in Wonderful Life—any replay would be different then the next. Gould critiqued the increasing complexity claim, arguing that diversification is always accompanied by decimation—once a mass extinction (say, an asteroid impact) occurs, there will then be subsequent diversification after the decimation.
We have no idea why certain organisms persisted over others after periods of decimation—and ‘adaptation’ to environments cannot be the whole story. Out of all of Gould’s writing that I have read in my life, this passage is one of my favorites as it perfectly captures the problem at hand:
Wind the tape of life back to Burgess times, and let it play again. If Pikaia does not survive in the replay, we are wiped out of future history—all of us, from shark to robin to orangutan. And I don’t think that any handicapper, given Burgess evidence as known today, would have granted very favorable odds for the persistence of Pikaia.
And so, if you wish to ask the question of the ages—why do humans exist?—a major part of the answer, touching those aspects of the issue that science can treat at all, must be: because Pikaia survived the Burgess decimation. This response does not cite a single law of nature; it embodies no statement about predictable evolutionary pathways, no calculation of probabilities based on general rules of anatomy or ecology. The survival of Pikaia was a contingency of “just history.” I do not think that any “higher” answer can be given, and I cannot imagine that any resolution could be more fascinating. We are the offspring of history, and must establish our own paths in this most diverse and interesting of conceivable universes—one indifferent to our suffering, and therefore offering us maximal freedom to thrive, or to fail, in our own chosen way. (Gould, 1989: 323)
Contingency is about counterfactuals—what could have happened, what could have been, or what would have been had some certain condition changed, with everything before that occurring as usual. Bradbury’s A Sound of Thunder wonderfully illustrates the contingency of the evolutionary process—change one seemingly small, minuscule thing in the past and this could snowball and cascade to huge changes in the future—we may never have existed or we would have existed but have been radically different. If we could go back in time and, say, crush a butterfly and see the changes it would have made, we could say that the event that caused the future to change was the crushing of that butterfly—this could have, eventually, led to the non-existence of a certain group of people or a certain group of animals which would have radically changed the outcome of the world—both the natural and human world.
So, if we could replay life’s tape from the very beginning, I do believe that life as we know it would be different—for if we played it from the beginning, we could have a scenario as described by Bradbury—everything could go exactly the same with one small seemingly minuscule change snowballing into a world that we would barely recognize.
Musicogenic Epilepsy
1800 words
I was watching the program Diagnose Me on Discovery Health and a woman kept having seizures whenever she heard a certain type of music—“alternative high-pitched female singing”, according to the woman—but her doctors didn’t believe her. So her and her husband began looking for specialists who specialize in hard-to-treat epilepsy. He recommended an endocranial EEG (images of such a surgery can be found below), which meant that the top part of her skull would be removed and electrodes would be placed onto the top of her brain. After the electrodes were placed on the brain. they played the music she said triggered her epilepsy—which was “high-pitched female singing”—and she began to seize. The doctor was shocked and he couldn’t believe what he saw. They ended up finding out that a majority—not all—of her seizing was coming from the right temporal lobe. So her and her husband had a choice—live with the seizures (which she couldn’t because she did not know where she would hear the music) or get part of her brain removed. She chose to have part of her right temporal lobe removed and when it was removed she no longer seized from hearing the music that formerly triggered her symptoms.
The condition is called “musicogenic epilepsy” which is a rare form of what is called “reflex epilepsy”—of which, another similar form involved hitting something which then causes seizing in the patient. (It’s called “reflex epilepsy” since the epileptic events occurs after an event—music, hitting something with your foot, seeing something on the television, etc.) This occurs when certain types of music are heard, certain musical notes can trigger electrical brain activity. The cure is to remove the part of the brain that is affecting the patient. (It is worth noting that many individuals throughout the past 100 years have had large sections of their brains removed and had no loss-of-functioning, staying pretty much the same as they were.) It is important to note that the music is not causing the seizures, it is triggering them—it brings them out. Most of the seizing is localized in the right temporal lobe (Kaplan, 2003), further being localized in Heschl’s gyrus (Nagahama et al, 2017). This has been noted by a few researchers since last century (Shaw and Hill, 1946; Fujinawa and Kawai, 1978) while the Joan of Arc was said to have her perception scrambled while hearing church bells; a Chinese poet stated that he became “absent-minded” and “sick” when hearing the flute-playing from the street vendor (Murray, 2010: 173).
The condition was first noted by a doctor in 1937, with the first known reference to this form of epilepsy being observed in the 1600s (Kaplan, 2003: 465). It affects about 1 in 10,000,000 people (Ellis, 2017). Critical reviews state not to underestimate the power of anti-epileptic drugs in the treatment and management of musicogenic epilepsy (Maguire, 2012), but in the case described above, such drugs did nothing to cure the woman’s seizures that occurred each time she heard a certain kind of music. The effect of music on seizing, it seems, is dichotomous with certain kinds of music either helping manage or causing seizing. The same melody, however, could be played in a different key and not cause seizing (Kaplan and Stoker, 2010) and so, it seems that certain types of sound frequencies influence/screw up the electrical activity in the brain which then leads to seizures of this kind. A specialist in epilepsy explains:
In people with reflex epilepsy, the trigger is extremely specific, and the seizure happens soon thereafter. “It can be a specific song by a particular person or even a specific verse of the song,” says Dr. So, who is a past president of the American Epilepsy Society. For some people, the trigger is a touch or motion. “If patients are interrupted in a particular way, if they are walking along and someone steps in front of them, they may have a seizure,” says Dr. So. In Japan, seizures caused by video games have been reported, he says, but they are highly unusual.
Dr. So evaluated a woman from Tennessee who began having seizures during church when she heard highly emotional hymns. She would blank out and drop her hymn book. At other times, Whitney Houston’s “I Will Always Love You” triggered seizures. The woman had a history of small seizures, but having one while hearing music was a new development. She said the seizures would typically begin with a sense of dread and the feeling that someone was lurking by her side. Dr. So and his Mayo Clinic team attached electrodes to the woman’s scalp to study electrical activity while she listened to different types of music. An electroencephalogram (EEG) showed that slow, emotional songs triggered seizure activity in her brain’s temporal lobe, while faster tunes did not. Dr. So diagnosed the woman with musicogenic epilepsy, a type of reflex epilepsy where seizures are caused by specific music or types of music, and prescribed antiseizure medication. He says he’s had another patient whose seizures were triggered by Rihanna’s “Disturbia” and Pharrell Williams’ “Happy.”
Though musicogenic epilepsy is extremely rare, it may be slightly underreported since many people with the disease may not put two and two together and link their seizing with the type of music or sounds they hear in their day-to-day life. One individual with epilepsy also recounts his experience with this type of rare epilepsy:
… but I still find that certain music, high pitched noise set’s off a kind of aura, I feel spaced out, have intense fear and it sounds almost like water rushing and I hear voices.
One case report exists of a man in which his later seizures were induced by music which prompted stress and a bad mood, implying that the aetiology of musicogenic epilepsy involves an association between the seizing and the patient’s mental state (Cheng, 2016).
We can see how the endocranial EEG looks and how it gets done (WARNING: GRAPHIC) by referring to Nagahama et al (2019):
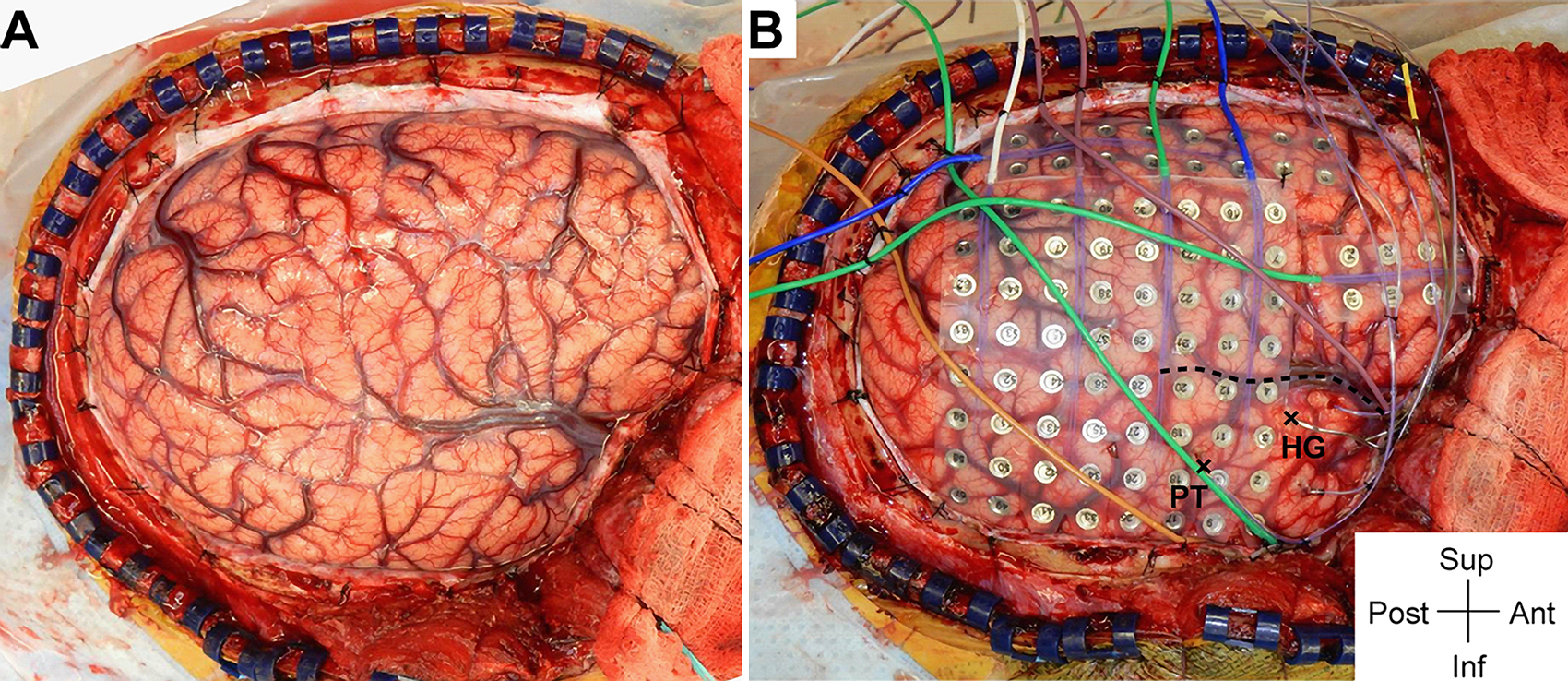
Intraoperative photographs demonstrating exposure and intracranial electrode placement. A right frontotemporoparietal craniotomy (A) allowed proper exposure for placement of grid, strip, and depth of electrodes (B), including the HG depth electrode. The sylvian fissure is marked with a dashed line. The HG depth electrode and PT depth electrose are marked with X symbols anteriorly and posteriorly, respectively, at their entry points at the cortical surface. Ant = anterior; inf = inferior; post = posterior; sup = superior.
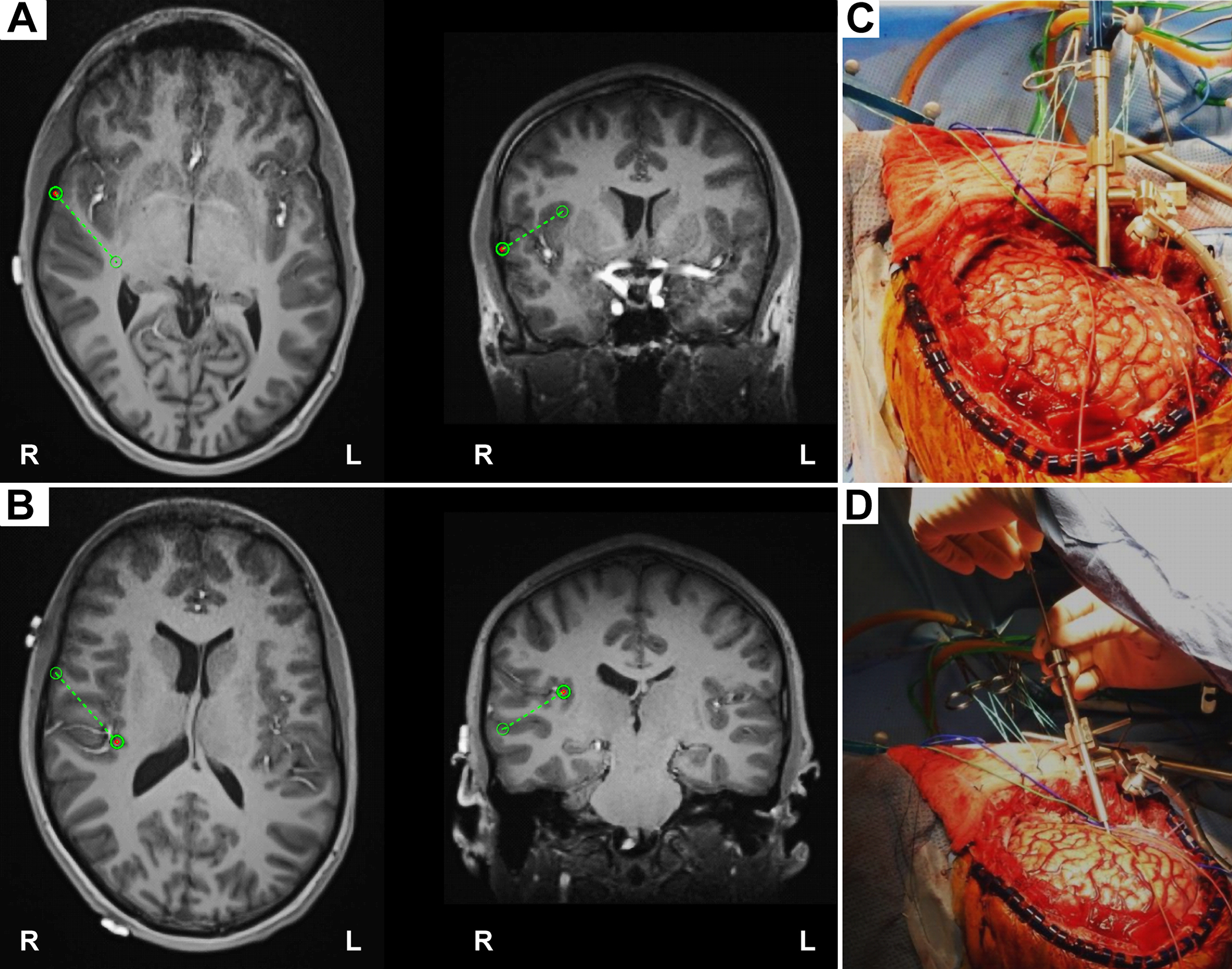
Intraoperative placement of the HG depth electrode. A: The planning view on the frameless stereotactic system (Stealth Navigation, Medtronic) showing the entry point and the trajectory (green circles and dotted lines). B: The similar planning view showing the target and the trajectory. C and D: Intraoperative photographs showing placement of the HG depth electrode. A Stealth Navigus probe was used to select the appropriate trajectory of a guiding tube positioned over the entry point (C). An electrode-guiding cannula was advanced through the tube to the previously determined depth (D). An actual depth electrode was subsequently passed through the cannula, followed by removal of the guiding tube/cannula system. Note the unique anterolateral-to-posteromedial trajectory within the STP for placement of the HG depth electrode.
The average age of onset of musicogenic epilepsy is 28 (Wieser et al, 1997) while the first cases are not reported until around one’s mid-to-late 30s due to the fact that most people are unware that music may be causing their seizures (Pittau et al, 2008; Generalov et al, 2018). This may be due to the fact that seizing may begin several minutes after hearing the music that affects the patient in question (Avanzini, 2003). While the specific tempo and pitch of music seems to have no effect on the beginnings of seizing (Wieser et al, 1997), many patients report that their specific triggers are due to hearing certain lines in songs (Tayah et al, 2006) which implies that it is not the music itself which is causing the seizing, but the emotional response that occurs to the patient after hearing the music and this is supported by the fact that many patients who report such symptoms are interested in music or are musicians themselves (Wieser et al, 1997).
See table 1 from Kaplan (2003: 466) for causes of musicogenic epilepsy in the literature:
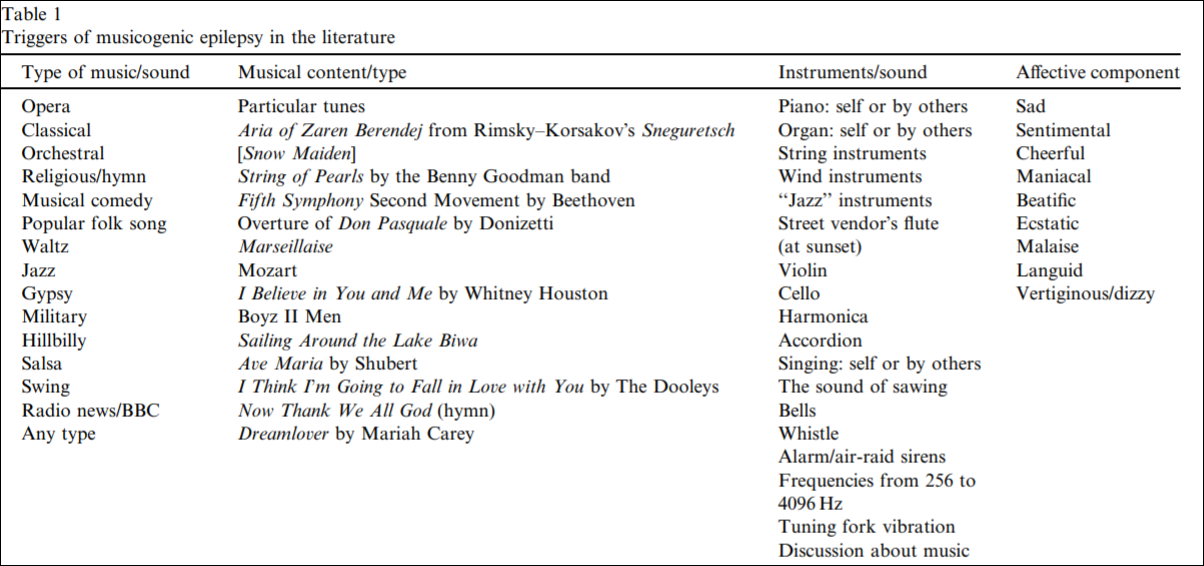
As can be seen by the above table, the mood component is related to the musical type; so the music elicits some sort of emotional state in the individual which would, it seems, to be part of the cause which then triggers the seizure—though the music/emotions are not causing the seizing itself, it is bringing them out.
Going to the shops was fraught with danger. Turning on the television was like playing russian roulette. Even getting into a lift was a gamble. For 23 years my life was hugely restricted because I had epileptic fits whenever I heard music.
If it was more than a few notes, a strange humming would start in my head, immediately followed by a seizure. I didn’t fall to the ground and twitch, but would wander around in a daze, my heart racing, my mind a blank. I also experienced hallucinations: people around me appeared microscopic and it felt as if I had been captured by an invisible force field. It was a terrifying experience and I felt drained for hours afterwards. (Experience: Music gave me seizures)
One woman describes her experience with musicogenic epilepsy for The Guardian. She did everything she could think of to stop the music-induced seizures—from sticking cotton balls into her ears to stop hearing sounds, to staying inside of the house (in case a car driving by played the type of music that triggered her seizing), to having a silent wedding with no music. She ended up getting referred to a specialist and she got her brain checked out. Come to find out, she had scarring on her right temporal lobe and so, surgery was done to fix it. She was cured from her condition and she could then attend social functions in which music was played.
The brain has the capacity to produce electricity, and so, in certain individuals with certain things wrong with the structure of their brains (like in their right temporal lobe), if they hear a certain kind of music or tune, they may then begin seizing. While the condition is rare (around 150 cases have been noted), strides are being made in discovering how and why such things occur. The only cure, it seems, is to remove the affected part of the brain—the right temporal lobe in a majority of cases. Such operations, however, do not always have the same debilitating effects (i.e., causing loss of mental capacity). That the brain’s normal functioning can be affected by sound (music) is very interesting and speaks to the fact that our brains are an enigma which is just beginning to be unraveled.
Life in the Time of Corona
2150 words
(Disclaimer: None of this is medical advice.)
Unless you’ve been living under a rock since December 2019, you should have heard the panic that SARS-CoV (which causes COVID-19—coronavirus disease) is causing ever since it emerged in Wuhan, China (Singhal, 2020). This virus spreads really easily—though asymptomatic transmission is thought to be rare, according to the CDC. There is one case report, though, of an infant who showed no signs of COVID-19 but had a high viral load (Kam et al, 2020). In any case, Trump flip-flopped from calling it a ‘hoax’ to taking it seriously, acknowledging the pandemic. “I’ve felt it was a pandemic long before it was called a pandemic“, Trump said. Ah, of course, It must have been just a facade to say it was a hoax. (Pandering to his base?) The ever prescient Trump knows all.
Speaking of prediction, Cheng et al (2007) stated “The presence of a large reservoir of SARS-CoV-like viruses in horseshoe bats, together with the culture of eating exotic mammals in southern China, is a time bomb. The possibility of the reemergence of SARS and other novel viruses from animals or laboratories and therefore the need for preparedness should not be ignored.” Quite the prediction from 13 years ago—implicating southern China’s “culture of eating exotic mammals”, which is more than likely the origin of this current outbreak.
There has been some discussion on whether or not the coronavirus is “as bad” as they’re saying, which has been criticized, for example, for not bringing up the context-dependency of the numbers. The number of cases in the US, though, as of Friday, March 20, 2020, was at 15,219 with 201 deaths. The number of cases keeps increasing daily. As of 3/22/2020, America has had 26,909 cases with 349 deaths while 178 recovered. Ninety-seven percent are in mild condition right now while three percent are in serious condition.
The current recommendations—social distancing, self-quarantining—are what we are doing to fight the virus, but I think we are going to need more drastic measures. Social distancing and self-quarantining will help to slow the spread of the virus, but the virus is still obviously spreading.
All of the talk about what to call it—Wuhan virus, Chinese virus, China virus, coronavirus—is irrelevant. Call it whatever you’d like, just make sure that whomever you’re communicating with knows what you’re talking about. (And, if you want to ensure they do, just call it “coronavirus” as that seems to be the name that has stuck these past few months.) I understand the want to identify where it began and spread from, but of course, others will use it for racial reasons.
The past few days there has been a lot of attention focused on hydroxychloroquine (HCQ) and azithromycin. These are anti-malarial drugs; a trial was done to see if it would have any effect on COVID-19 (Liu et al, 2020).
For HCQ, there is an “expert consensus” on HCQ treatment and COVID-19, and they state:
It recommended chloroquine phosphate tablet, 500mg twice per day for 10 days for patients diagnosed as mild, moderate and severe cases of novel coronavirus pneumonia and without contraindications to chloroquine.
Chloroquine has been shown to reduce spread and infection of coronaviruses (Vincent et al, 2005; Savarino et al, 2006; Wang et al, 2015; Wang et al, 2020). Wang et al (2015) note that:
HCQ and chloroquine are cellular autophagy modulators that interfere with the pH-dependent steps of endosome-mediated viral entry and late stages of replication of enveloped viruses such as retroviruses, flaviviruses, and coronaviruses (Savarino and others 2003; Vincent and others 2005).
I don’t know what to make of such results, I am awaiting larger trials on the matter. There is some hope in using anti-malarial drugs in the hopes of curbing the disease.
The Chinese knew that this virus was similar to other SARS strains, their scientists were ordered to stop testing on samples and to destroy the evidence. (See here for a timeline of the case.) The scary thing is that this virus has symptoms similar to the common cold that we get every winter so some may brush it off as ‘just the cold.’ I came down with a cold at the end of January and I was out of commission for the week. Thankfully, it was not COVID-19.
Italy and China had a strong trade relationship, which seems to have cost Italy. Italy has one of the oldest populations in the world. Ninety-nine percent of corona deaths in Italy, though, had other health problems, such as being obese, having hypertension, previous heart problems, etc. Italy began locking down cities as early as two weeks ago, though they have reported a staggering 4,825 deaths. This, though, is to be expected when a quarter of the country is aged 65 and older with multiple comorbidities. So if it is that bad in Italy with a smaller population, what does that mean for the US in the coming weeks?
New York and New Jersey banned gatherings of more than 50 people, dining out, gyms, etc in an effort to curb the transmission of the virus. Then, Friday night at midnight (3/21/2020) only essential businesses were allowed to stay open—essentials include healthcare, infrastructure, food (no dining-in, take-out or delivery only), grocery stores, mail, laundromats, law enforcement, etc. In NJ, all businesses were ordered to close down except things like grocery stores, banks, pet stores, convenience stores, etc. This affected me (gyms closed) and so I cannot work. I preempted this a few weeks ago and found a job in logistics, but I got laid off on Friday due to the shut-downs of nonessential businesses (the shut-downs decreased my work). Now, I’m thinking about hunkering down until at least June. Due to what we know about the social determinants of health (Marmot, 2005; Cockerham, 2007; Barr, 2019) we can expect what is associated with low class (poor health, stress, etc) to increase as well.
This is only going to get worse in the coming weeks. I do see a decreased number of people out on the street, and I am glad that states are taking measures to curb the transmission of the virus, but I still see people not really taking it seriously. From the ads on the radio informing us about what is going on around the country in terms of death rate and transmission rate, they are strongly suggesting for people to stay home and to avoid public transportation. Obviously, in places that are enclosed and many people walk in and out in a timely manner, that is a great place for the virus to spread. ….what if we’re doing what the virus ‘wants’? Don’t worry, the evo-psychos are here to tell us just-so stories.
By this account, COVID-19 is turning out to be a remarkably intelligent evolutionary adversary. By exploiting vulnerabilities in human psychology selectively bred by its pathogen ancestors, it has already shut down many of our schools, crashed our stock market, increased social conflict and xenophobia, reshuffled our migration patterns, and is working to contain us in homogenous spaces where it can keep spreading. We should pause to remark that COVID-19 is extraordinarily successful epidemiologically, precisely because it is not extremely lethal. With its mortality rate of 90%, for example, Ebola is a rather stupid virus: It kills its host — and itself — too quickly to spread far enough to reshape other species’ life-ways to cater to its needs. (The Coronavirus Is Much Worse Than You Think)
Ah, the non-lethality of COVID-19 is to its benefit—it can spread more, it is an “intelligent evolutionary adversary” but it is causing a “moral panic” as well. The damage to our psyche, apparently, is worse than what it could do to our lungs. And while I do agree that this could damage our collective psyches, we don’t need to tell just-so stories about it.
When we come out of this pandemic, I can see us being very cautious as we go back to normal life (in places affected, people are still going out where I live but not as much). Then, hundreds of years later, Evolutionary Psychologists notice how averse people are to go outside. “Why are people so introverted? Why do people avoid others?” They ask. “Why are those who wear masks more attractive than those who don’t wear masks?” They then discover the pandemic of the 2020s which ravaged the world. “Ah! Critics won’t be able to say ‘just-so stories’ now! We know the preceding event—we have a record of it happening!” And so, the evo-psychos celebrate.
In all seriousness, if people do take this seriously, there may be some social/cultural customs changes, including how we greet people.
Cao et al (2020) conclude: “The East Asian populations have much higher AFs [allele frequencies] in the eQTL variants associated with higher ACE2 expression in tissues (Fig. 1c), which may suggest different susceptibility or response to 2019-nCoV/SARS-CoV-2 from different populations under the similar conditions.” Asian men smoke more cigarettes than Asian women (Ma et al, 2002, 2004; Chae, Gavin, and Takeuchi, 2006; Tsai et al, 2008). In your lungs you have what is called “cilia fibers’ and these fibers move debris and microbes out while they also protect the bronchus and trap microorganisms. COVID-19 attacks these same cilia fibers that degrade when one smokes. Therefore, the fact that East Asian populations have higher allele frequencies in ACE2 expression tissues along with higher rates of smoking may be why Asian men seem to be affected more than Asian women. In any case, smokers of any race need to exercise caution.
What if after the pandemic is over life does not go back to normal? What if life during the pandemic becomes the ‘new normal’ when the pandemic is over because everyone is paranoid about contracting the virus again? For introverts, like myself, it’s easy to lock-in. I have hundreds of books to choose from to read, so if I do choose to lock in for 2 months (which I am thinking about), then I won’t really be bored. But my thing is this: what’s the point of locking in when everyone else isn’t, the virus still spreads and when you finally go out the pandemic is still going on? The point of quarantining is understandable—but if everyone doesn’t do it, will it really work? Libertarians be damned, we need the government to step in and do these kinds of things right now. It’s not about the individual, but the public as a whole.
On the other hand, it is thought-provoking to think about the fact that the government is ramping up the drama in the news to see how far they can go with social control. What a perfect way to see how far the public would go if they got “suggestions” from the government. Just like the government is “suggesting” we be inside at 8 pm to mitigate viral transmission, for example, it’s just to see what we would accept and how far they can go until they make it mandatory. It is interesting to think about how all of the toilet paper, hand sanitizer, hand soap, etc are being sold out everywhere.
People in my generation have 9/11 to look back to as the “That’s when the world changed” time. Well, kids alive today (around 7-15 years old) are experiencing their “9/11”, as that’s when the world changed for them. But this coronavirus pandemic is not on a country level—it is on the world level. The whole WORLD is affected. So since our Gregorian calendar is based off the birth of Jesus, I propose the following: change 1-December 2019 AD/CE to BC (before Corona) and anything after December 2019 to AC (after Corona).
I hope that, looking back on the current goings-on now that we are not talking about high death tolls and that we can get this under control. The only course of action (for now) is to attempt to stop the transmission of the virus—which is to stop its transmission from human to human. COVID-19 can be said to largely be a social disease since that is how it is most likely to be transmitted, which is why social distancing is so important. Being social is how the virus spreads, so to stop spreading the virus we need to be anti-social.
If we do not heed these warnings, then we will permanently be living in the Time of Corona. Coronavirus will be dictating what we do and when we do it. No one will want to get sick but no one will also want to take the steps needed in order to eradicate the threat. This thing is just getting started, by the end of the month into the first few weeks of April it is only going to get worse. I hope you all are prepared (have food [meat], water, soap, etc) because we’re in for a hell of a ride. With many businesses closing down in an effort to curb the transmission of COVID-19, many people will be out of jobs—many low-income people.
The Hierarchical Nature of Living Systems, Species, and Race
2100 words
Biology is one of the most interesting sciences since, at its core, it is the study of life and living systems. The biological organization of living systems and the ecosystems these living systems find themselves in are interesting to learn about, since we can then discern different species and learn how and when to delineate separate species based on a set of pre-conceived measures. The classification of human races in these systems will be discussed, along with why human races are not different species.
The organization of living systems
Living systems show hierarchical organization, each system—from the physiological to the physical—interacting with each other. However, a key factor in the organization of these interactions is the degree of the complexity of the interactions in question. We can look at the organization of the biological world as hierarchical—that is, each level builds on the preceding level, so we get from atoms to the biosphere and everything in between is what we call “life” and also show how these complex, living biological systems live and exist due to the hierarchical organization of living systems. The point is, life does not have a simple definition, but all living systems share similar characteristics that can describe life. Biologists organize living systems hierarchically, from the subcellular level to the entire biosphere, and then study the interactions that occur which cannot be predicted from just studying the sum of its parts. This is why a holistic—and not reductionistic—approach needs to be taken when studying and describing living systems.
The hierarchy is:
The cellular level, which includes: atoms, molecules, macromolecules, and organelles; the organismal level which include: tissue, organs, the organ system, and the organism; the populational level which includes: the population, species, and the community; and finally the highest level, the ecosystem level which includes the ecosystem and the biosphere.
At the cellular level, we have atoms which are the fundamental elements of matter and are joined together by chemical bonds called molecules. large and complex molecules are called macromolecules, DNA—which stores hereditary information—is a type of macromolecule. Complex biological molecules are then assembled into organelles, where cellular activities are organized. A mitochondrion is, for example, an organelle with a cell that extracted energy from consumed food molecules. And finally, we have cells, which are the basic unit of life.
Next, we have the organismal level, and cells of multicellular organisms make up three levels of organization. Tissues, which are groups of similar cells which function together as a unit. Tissues then are grouped into organs which are structures of the body which are composed of many different kinds of tissues which act in a structural manner and as a unit. Then we have organ systems, such as the nervous system which is the sensory organs, brain and spinal cord, and the network of neurons that convey signals to different parts of the body.
Then we have the populational level. This includes the individual organisms which occupy various hierarchical levels in the biological world. A population is a group of organisms all living in the same place. Together, all populations of a particular kind form a species—members of a species must look similar and be able to interbreed. Then finally, we have the biological community which consists of all of the populations coexisting together in one place.
Lastly, we have the ecosystem level. This is the highest tier of biological organization (the lowest being the cellular level). A biological community and its physical habitat (such as soil composition, available water etc) in which it finds itself in and lives and competes with other organisms constitute an ecosystem while the entire planet is the highest of all levels of biological organization—the biosphere. All of these systems together can be seen as the hierarchical organization of living systems.
(See Mason et al, 2018 for more discussion of the above points.)
Organismal classification
Now, in these differing biological hierarchies, we find differing Eukarya, Prokarya, and Bacteria. The in-use classification system is the Linnean hierarchy. Differences exist between organisms, this is obvious. But it is a bit more tricky to classify these organisms and place them into like groups. Then, in the 1750s, Carolus Linnaeus came along and instituted a binomial classification system for organisms—the most commonly-known binomial being Homo sapiens—which was much simpler than the polynomial names
The hierarchy is as follows:
1. Species;
2. Genus;
3. Family;
4. Order;
5. Class;
6. Phylum;
7. Kingdom; and
8. Domain. Domains can then be split into Archaea, Bacteria, and Eukarya. Domains are the largest taxons, being that they comprise every organism that we know of.
For example, our species is sapiens, our genus is Homo, our family is Hominidae, our order is primates, our class is Mammalia, our phylum is Chordata (with a subphylum Craniata), our kingdom is Animalia and our domain is Eukarya. This is our species’ taxonomic classification.
The traditional classification system—the Linnean system—groups species into genera, families, orders, classes, phyla, and kingdoms. Thus, these systems classify different organisms on the basis of similar traits, and since they consist of a mix of derived and ancestral traits, they do not necessarily take into account different evolutionary relationships.
There are of course limitations to the Linnean hierarchy:
1) Many “higher” taxonomic ranks are not monophyletic and so do not represent real groups (like Reptilia). For something to be a “natural group”, a common ancestor and its descendants must all derive from descent from a common ancestor, so any other type of taxonomic ranks are created by taxonomists, such as paraphyletic and polyphyletic.
2) Linnean ranks are not equivalent. Two families may not represent clades that arose at the same time, because one family may have diverged millions of years before the other family and so the two families had differing amounts of time to diverge and acquire new traits. So comparisons in the Linnean sense may be misleading and we should then use hypotheses of phylogenetic relationships.
What is a species?
It should first be noted that species are, indeed, real. New species arise when isolated organisms of one population become genetically/geographically isolated for a period of time. Over time, as the split population spends time geographically and genetically isolated, they cannot interbreed with the parent population and thusly attain separate species status. This is the received view, the biological species concept.
There are a wide range of species concepts and they all capture the differences that different theorists believe we should emphasize in our classification of organisms.
The phenetic species which appeal to the intrinsic similarities of organisms. The biological species concept which appeals to reproductive isolation (one version of the biological species concept is the recognition concept, which defines species as a system of mating recognition. The cohesion species concept which generalizes the biological species concept and it recognizes that gene flow isn’t the only factor that holds a population together and makes it different from other populations. The ecological species concept which defines species by appealing to the fact that members of a species are in competition with one another because of the need the same resources. And the phylogenetic and evolutionary species concept which define species as segments on the tree of life (the phylogenetic species concept, for instance, holds the term ‘species’ should be reserved for groups of populations that have been evolving independently of other populations.
Sterelny and Griffiths (1999) tackled this in their book Sex and Death: An Introduction to Philosophy of Biology:
While we think cladism presents the best view of systematics, biological classification nevertheless poses an unsolved problem. If we were to accept either evolutionary taxonomy, which builds disparity into its classification system, or phenetic taxonomy, which is based on the idea of nested levels of similarity, traditonal taxonomic levels would be quite defensible. Within those taxonomic pictures, the idea of genus, family, order, and so on makes quite good sense. If cladism is the only defensible picture of systematics, the situation is more troubling. From that perspective, these taxonomic ranks make little sense. Cladists do not think there is a well-defined objective notion of the amount of evolutionary divergence. That, in part, is why they are cladists. Hence, they do not think there will be any robust answer to the questions, when should we call a monophyletic group of species a genus? a family? an order? Only monophyletic groups should be called anything, for they are well-defined chucnks of the tree. But only science greets the question, are the chimps plus humans a genus? It has long been receieved wisdom in taxonomy that there is something arbitrary about taxonomic classification above species. These decisions are judgement calls. So cladists only show a somewhat more extreme version of a skepticism that has long existed. The problem of high taxonomic ranks would not matter except for the importance of the information expressed using them. Hence cladism reinforces the worry that when, for example, we consider divergent extinction and survival patterns, our data may not be tobust, for our units may not be commensurable. Unfortunately, it does this without suggesting much of a cure.
Where does race fit in?
Racehood is simple: A race is a group of humans that: Condition 1; is distinguished from other groups of humans by patterns of visible physical features; Condition 2: is linked by common geographic ancestry which is peculiar to members of this group; and Condition 3: originates from a distinctive geographic location.
So now all we need to do is go through four steps: 1) recognize that there are patterns of visible physical features which correspond to geographic ancestry; 2) observe that these patterns of visible physical features which correspond to geographic ancestry are exhibited between real, existing groups; 3) note that these real existing groups that exhibit these patterns by geographic ancestry satisfy C1-C3; and 4) infer that race exists.
Some may argue that the races are different species, citing the same patterns of visible physical features discussed above. However, if we are referring to the biological species concept, then the human races are not different species at all since all human races can produce fertile offspring with one another. Our genus, of course, is Homo, all of the human races are the same genus; though some may attempt to use the previously-discussed conditions for racehood as conditions for specieshood for humans, the most preferred method for delineating species currently is the biological species concept, and since all of the human races can produce fertile offspring then the human races are not different species.
In keeping with the classification system that is currently used today (see above), where would human races fit into our taxonomy? Falling within our species sapiens seems like a good start, and since the races can interbreed and have fertile offspring, then they are not different species but are the same species, despite phenotypic differences. Thus, human races would be within species but under subspecies. Using this line of logic, human races cannot be different species, despite claims to the contrary that human races are different species based on patterns of visible physical features which correspond to geographic ancestry. That’s enough to denote racehood, not specieshood.
Conclusion
The study of life—in all of its forms and in all of its environments—is one of the most important things we, as humans, can do. From it, we can learn where we came from and even—possibly—where we may be going. Once we understood the biological hierarchy and how upper levels are built from lower levels working together, then we were better able to understand how living systems act on the inside—cellularly and physiologically—to the outside—organismal and environmental interaction. From organismal and environmental interaction, speciation may occur. The highest level of the organization of living systems is the biosphere—and it is so because the living systems that are driven by the smallest cellular interactions interact with other species, the ecosystem and the biosphere.
Species do exist, but there are numerous species concepts—over twenty. One of the more popular species concepts in use is the cladistic species concept. In this species concept, a species is a lineage of populations between two specific branch points. The cladistic concept thusly recognizes differing species by differing branch points and how much change occurs between them (see Ridley, 1989).
The classification of different organisms into different species is pretty straightforward, though it falls prey to oversimplification since it only focuses on similar traits. Species exist, this is established. But races are not species, contrary to some beliefs. Different races can interbreed and, I would argue, that for there to be separate species, human races would not be able to interbreed. Yes, there are physical and morphological differences between races, but, as argued, this is not enough to denote speciation, but it is enough to denote raciation.
