
2050 words
The Modern Synthesis (MS) has entrenched evolutionary thought since its inception in the mid-1950s. The MS is the integreation of Darwinian natural selection and Mendelian genetics. Key assumptions include “(i) evolutionarily significant phenotypic variation arises from genetic mutations that occur at a low rate independently of the strength and direction of natural selection; (ii) most favourable mutations have small phenotypic effects, which results in gradual phenotypic change; (iii) inheritance is genetic; (iv) natural selection is the sole explanation for adaptation; and (v) macro-evolution is the result of accumulation of differences that arise through micro-evolutionary processes” (Laland et al, 2015).
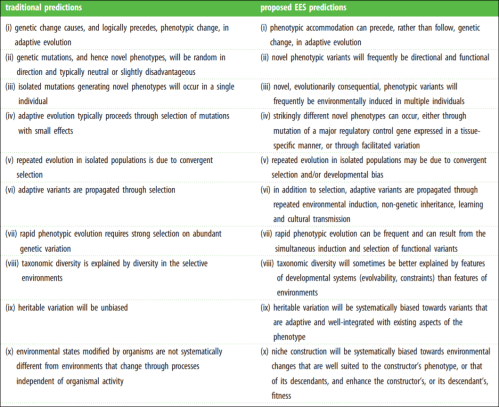
Laland et al (2015) even have a helpful table on core assumptions of both the MS and Extended Evolutionary Synthesis (EES). The MS assumptions are on the left while the EES assumptions are on the right.

Darwinian cheerleaders, such as Jerry Coyne and Richard Dawkins, would claim that neo-Darwinisim can—and already does—account for the assumptions of the EES. However, it is clear that that claim is false. At its core, the MS is a gene-centered perspective whereas the EES is an organism-centered perspective.
To the followers of the MS, evolution occurs through random mutations and change in allele frequencies which then get selected for by natural selection since they lead to an increase in fitness in that organism, and so, that trait that the genes ’cause’ then carry on to the next generation due to its contribution to fitness in that organism. Drift, mutation and gene flow also account for changes in genetic frequencies, but selection is the strongest of these modes of evolution to the Darwinian. The debate about the MS and the EES comes down to gene-selectionism vs developmental systems theory.
On the other hand, the EES is an organism-centered perspective. Adherents to the EES state that the organism is inseparable from its environment. Jarvilehto (1998) describes this well:
The theory of the organism-environment system (Jairvilehto, 1994, 1995) starts with the proposition that in any functional sense organism and environment are inseparable and form only one unitary system. The organism cannot exist without the environment and the environment has descriptive properties only if it is connected to the organism.
At its core, the EES makes evolution about the organism—its developmental system—and relegates genes, not as active causes of traits and behaviors, but as passive causes, being used by and for the system as needed (Noble, 2011; Richardson, 2017).
One can see that the core assumptions of the MS are very much like what Dawkins describes in his book The Selfish Gene (Dawkins, 1976). In the book, Dawkins claimed that we are what amounts to “gene machines”—that is, just vehicles for the riders, the genes. So, for example, since we are just gene machines, and if genes are literally selfish “things”, then all of our actions and behaviors can be reduced to the fact that our genes “want” to survive. But the “selfish gene” theory “is not even capable of direct empirical falsification” (Noble, 2011) because Richard Dawkins emphatically stated in The Extended Phenotype (Dawkins, 1982: 1) that “I doubt that there is any experiment that could prove my claim” (quoted in Noble, 2011).
Noble (2011) goes on to discuss Dawkins’ view that on genes:
Now they swarm in huge colonies, safe inside gigantic lumbering robots, sealed off from the outside world, communicating with it by tortuous indirect routes, manipulating it by remote control. They are in you and me; they created us, body and mind; and their preservation is the ultimate rationale for our existence. (1976, 20)
Noble then switches the analogy: Noble likens genes, not as having a “selfish” attribute, but to that of being “prisoners”, stuck in the body with no way of escape. Noble then says that, since there is no experiment to distinguish between the two views (which Dawkins admitted). Noble then concludes that, instead of being “selfish”, the physiological sciences look at genes as “cooperative”, since they need to “cooperate” with the environment, other genes, gene networks etc which comprise the whole organism.
In his 2018 book Agents and Goals in Evolution Samir Okasha distinguishes between type I and type II agential thinking. “In type 1 [agential thinking], the agent with the goal is an evolved entity, typically an individual organism; in type 2, the agent is ‘mother nature’, a personification of natural selection” (Okasha, 2018: 23). An example of type I agential thinking is Dawkins’ selfish genes, while type II is the personification that one imputes onto natural selection—which Okasha states that this type of thinking “Darwin was himself first to employ” (Okasha, 2018: 36) it.
Okasha states that each gene’s ultimate goal is to outcompete other genes—for that gene in question to increase its frequency in the organism. They also can have intermediate goals which is to maximize fitness. Okasha gives three rationales on what makes something “an agent”: (1) goal-directedness; (2) behavioral flexibility; and (3) adaptedness. So the “selfish” element “constitutes the strongest argument for agential thinking” of the genes (Okasha, 2018: 73). However, as Denis Noble has tirelessly pointed out, genes (DNA sequences) are inert molecules (and are one part of the developing system) and so do not show behavioral flexibility or goal-directedness. Genes can (along with other parts of the system working in concert with them) exert adaptive effects on the phenotype, though when genes (and traits) are coextensive, selection cannot distinguish between the fitness-enhancing trait and the free-riding trait so it only makes logical sense to claim that organisms are selected, not any individual traits (Fodor and Piatteli-Palmarini, 2010a, 2010b).
It is because of this, that the Neo-Darwinian gene-centric paradigm has failed, and is the reason why we need a new evolutionary synthesis. Some only wish to tweak the MS a bit in order to allow what the MS does not incorporate in it, but others want to overhaul the entire thing and extend it.
Here is the main reason why the MS fails: there is absolutely no reason to privilege any level of the system above any other! Causation is multi-level and constantly interacting. There is no a priori justification for privileging any developmental variable over any other (Noble, 2012, 2017). Both downward and upward causation exists in biological systems (which means that molecules depend on organismal context). The organism also able to control stochasticity—which is “used to … generate novelty” (Noble and Noble, 2018). Lastly, there is the creation of novelty at new levels of selection, like with how the organism is an active participant in the construction of the environment.
Now, what does the EES bring that is different from the MS? A whole bunch. Most importantly, it makes a slew of novel predictions. Laland et al (2016) write:
For example, the EES predicts that stress-induced phenotypic variation can initiate adaptive divergence in morphology, physiology and behaviour because of the ability of developmental mechanisms to accommodate new environments (consistent with predictions 1–3 and 7 in table 3). This is supported by research on colonizing populations of house finches [68], water fleas [132] and sticklebacks [55,133] and, from a more macro-evolutionary perspective, by studies of the vertebrate limb [57]. The predictions in table 3 are a small subset of those that characterize the EES, but suffice to illustrate its novelty, can be tested empirically, and should encourage deriving and testing further predictions.
[Table 3]
There are other ways to verify EES predictions, and they’re simple and can be done in the lab. In his book Above the Gene, Beyond Biology: Toward a Philosophy of Epigenetics, philosopher of biology Jan Baedke notes that studies of epigenetic processes which are induced in the lab and those that are observed in nature are similar in that they share the same methodological framework. So we can use lab-induced epigenetic processes to ask evolutionary questions and get evolutionary answers in an epigenetic framework. There are two problems, though. One, that we don’t know whether experimental and natural epigenetic inducements will match up; and two we don’t know whether or not these epigenetic explanations that focus on proximate causes and not ultimate causes can address evolutionary explananda. Baedke (2018: 89) writes:
The first has been addressed by showing that studies of epigenetic processes that are experimentally induced in the lab (in molecular epigenetics) and those observed in natural populations in the field (in ecological or evolutionary epigenetics) are not that different after all. They share a similar methodological framework, one that allows them to pose heuristically fruitful research questions and to build reciprocal transparent models. The second issue becomes far less fundamental if one understands the predominant reading of Mayr’s classical proximate-ultimate distinction as offering a simplifying picture of what (and how) developmental explanations actually explain. Once the nature of developmental dependencies has been revealed, the appropriateness of developmentally oriented approaches, such as epigenetics, in evolutionary biology is secured.
Further arguments for epigenetics from an evolutionary approach can be found in Richardson’s (2017) Genes, Brains, and Human Potential (chapter 4 and 5) and Jablonka and Lamb’s (2005) Evolution in Four Dimensions. More than genes alone are passed on and inherited, and this throws a wrench into the MS.
Some may fault DST for not offering anything comparable to Darwinisim, as Dupre (2003: 37) notes:
Critics of DST complain that it fails to offer any positive programme that has achievements comparable to more orthodox neo-Darwinism, and so far this complaint is probably justified.
But this is irrelevant. For if we look at DST as just a part of the whole EES programme, then it is the EES that needs to—and does—“offer a positive programme that has achievements comparable to more orthodox neo-Darwinism” (Dupre, 2003: 37). And that is exactly what the EES does: it makes novel predictions; it explains what needs to be explained better than the MS; and the MS has shown to be incoherent (that is, there cannot be selection on only one level; there can only be selection on the organism). That the main tool of the MS (natural selection) has been shown by Fodor to be vacuous and non-mechanistic is yet another strike against it.
Since DST is a main part of the EES, and DST is “a wholeheartedly epigenetic approach to development, inheritance and evolution” (Griffiths, 2015) and the EES incorporates epigenetic theories, then the EES will live or die on whether or not its evolutionary epigenetic theories are confirmed. And with the recent slew of books and articles that attest to the fact that there is a huge component to evolutionary epigenetics (e.g., Baedke, 2018; Bonduriansky and Day, 2018; Meloni, 2019), it is most definitely worth seeing what we can find in regard to evolutionary epigenetics studies, since epigenetic changes induced in the lab and those that are observed in natural populations in nature are not that different. This can then confirm or deconfirm major hypotheses of the EES—of which there are many. It is time for Lamarck to make his return.
It is clear that the MS is lacking, as many authors have pointed out. To understand evolutionary history and why organisms have the traits they do, we need much more than the natural selection-dominated neo-Darwinian Modern Synthesis. We need a new synthesis (which has been formulated for the past 15-20 years) and only through this new synthesis can we understand the hows and whys. The MS was good when we didn’t know any better, but the reductionism it assumes is untenable; there cannot be any direct selection on any level (i.e., the gene) so it is a nonsensical programme. Genes are not directly selected, nor are traits that enhance fitness. Whole organisms and their developmental systems are selected and propagate into future generations.
The EES (and DST along with it) hold right to the causal parity thesis—“that genes/DNA play an important role in development, but so do other variables, so there is no reason to privilege genes/DNA above other developmental variables.” This causal parity between all tools of development is telling: what is selected is not just one level of the system, as genetic reductionists (neo-Darwinists) would like to believe; it occurs on the whole organism and what it interacts with (the environment); environments are inherited too. Once we purge the falsities that were forced upon us by the MS in regard to organisms and their relationship with the environment and the MS’s assumptions about evolution as a whole, we can then truly understand how and why organisms evolve the phenotypes they do; we cannot truly understand the evolution of organisms and their phenotypes with genetic reductionist thinking with sloppy logic. So who wins? The MS does not, since it has causation in biology wrong. This only leaves us with the EES as the superior theory, predictor, and explainer.
