
Home » Skin Color
Category Archives: Skin Color
Nina Jablonski on Race
1550 words
Nina Jablonski’s work on vitamin D and the implications that lighter skin had not only on our evolution but our health are extremely important for understanding how we evolved after the out of Africa migration. However, Jablonski then takes what she has written about skin color over the past few decades and concludes that race doesn’t exist. Jablonski believes that the term “race” should be discontinued from our lexicon, but as most may know, the term “race” does not need to disappear from our lexicon. (Watch her TED Talk Skin Color is an Illusion.)
In 2014, Nina Jablonski stated that the term “race” was ready for scientific retirement. In the article—and her book (Jablonski, 2012: chapters 9 and 10)—she states that race was a “vague and slippery concept”, eschewing the views of Kant and Hume as “racist”. She talks about how Kant was really one of the first people to recognize and categorize groups of people as “races”, stating that skin color, hair type, skull type etc—along with differing mores, aptitudes, and capacity for civilization—arranged in a hierarchical manner with Europeans at the top. A climatic theory was held, which stated that the original humans were light and became darker since “the transformation from light to dark was a form of degeneration, a departure from the norm” (Jablonski, 2012: 143).
She then discusses how, in Biblical history, skin color was meaningful, meaningful because it was believed that darker-skinned races were descendants of Ham:
And the sons of Noah, that went forth of the ark, were Shem, and Ham, and Japheth: and Ham is the father of Canaan. These are the three sons of Noah: and of them was the whole earth overspread. And Noah began to be an husbandman, and he planted a vineyard: And he drank of the wine, and was drunken; and he was uncovered within his tent. And Ham, the father of Canaan, saw the nakedness of his father, and told his two brethren without. And Shem and Japheth took a garment, and laid it upon both their shoulders, and went backward, and covered the nakedness of their father; and their faces were backward, and they saw not their father’s nakedness. And Noah awoke from his wine, and knew what his younger son had done unto him. And he said, Cursed be Canaan; a servant of servants shall he be unto his brethren. And he said, Blessed be the Lord God of Shem; and Canaan shall be his servant. (Genesis, 9: 18-26)
So Noah’s three sons—Ham, Japheth and Shem—were seen to be the three modern-day races of man—Africans, Europeans, and Asians, respectively. The term “servant of servants” was taken to mean that the descendants of Ham would serve the descendants of Shem and Japheth. This, according to those who believed the authority of the Bible, was enough to justify chattel slavery.
Jablonski—in an interview with the magazine Nautilus—stated that there “are no clean breaks between human populations. Individuals have different groups of genes” and that “Only a tiny fraction of alleles, and a small fraction of allelic combinations, is restricted to a single geographic region, and even less to a single population” which “is why attempts to identify races in humans have failed.” She commits the continuum fallacy, and the argument form is thus: “One extreme is X, at another is Y. There is no definable point where X becomes Y. Therefore, there is no difference between X and Y.” This has also been called the “Argument of the Beard”: at what point does a man not become clean shaven?
The use of the continuum fallacy, that there “are no clean breaks between human populations” shows how far the “race is a social construct” line has come (it is, but that race is a social construct does not also mean that it cannot also be a significant biological reality). The continuum fallacy is one of the most-used fallacies by those who deny race. Though, those who use the continuum fallacy are only attempting to argue that the claim is “too vague” because it is not as precise as they would like it to be. It does not matter that there “are no clean breaks between human populations“; what matters is that patterns of visible physical features correspond to geographic ancestry, and this is what we find.
Her second problem arises when she says that “Only a tiny fraction of alleles, and a small fraction od allelic combinations, is restricted to a single geographic region, and even less to s single population“. That there are no “race genes” or “genes for race” does not mean that race does not exist as a biological reality; these rigid “either this or that” definitions that some people have for race, such as race-specific genes are strawmen: people who believe that race is a significant biological reality do not believe in race-specific genes. That there are no race-specific genes does not mean that race doesn’t exist, as we know that genes are expressed differently in different races.
Finally, she claims that this “is why attempts to identify races in humans have failed“, though these attempts have not failed, of course. So-called races are distinguished by patterns of visible physical features; these patterns are observed between real, existing groups; these real existing groups that share these patterns of visible physical features satisfy the requisites of minimalist race; therefore race exists. Of course, Jablonski has reservations about acknowledging the reality of race due to how the transatlantic slave trade was promogulated through so-called differences that stemmed from Noah and Ham’s curse, but I fail to see why she would discard the argument just provided for the existence of race since differences in mores, intelligence, physical and mental abilities, are not discussed in the argument. ONLY the observable differences between populations are observed, with no value-judgment put onto each race, such as having lower “intelligence” or differing mores compared to another race.
She also states, in an interview with the New York Times, that skin color is not about race, “it’s about sun and how close our ancestors lived to the Equator. Skin color is what regulates our body’s reaction to the sun and its rays. … That shows that color is not a permanent trait.” That the differences in skin color observed in human populations can change over time does not mean that skin color “is not about race” as Jablonski claims. Skin color is one physical trait to delineate races, along with hair type, physiognomy, and anatomy, that groups peoples into groups we call “races”. This is not a good argument against the existence of race; of course anatomy, physiology, and physiognomy can change over time: but this does not mean that race does not exist!
Michael Hardimon’s race concepts (Hardimon, 2017) show that one does not need to believe that races differ in “intelligence”, mores, etc to believe in the existence of race. The concept takes everything from the racialist concept and “minimizes it”, taking the aspect of visible differences in physical features, while leaving the so-called mental differences (“intelligence”, mores) alone. This is enough to recognize that race exists and, as Jablonski has noted for decades in her career, being displaced from the environment where your skin color evolved causes an environmental mismatch which then—in the case of black Americans—may lead to vitamin D deficiency. This is one significant aspect that shows that race has an impact on health policy.
The minimalist concept of race is “deflationary” in that it does not discuss what we “can’t see” with our own eyes; it only discusses physical traits which should be enough for Jablonski to say that race is real and exists as a biological reality. Combined with the known health effects of, for instance, living in differing climates with differing amounts of UV radiation that is not “for” your skin color has further consequences and is why, in some cases, race-based medicine should stick around (though I am aware that, first and foremost, the individual matters first in a medical context, racial membership is secondary).
In sum, Jablonski refers to old and outdated individuals when speaking about the biological reality of race. She does a good job chronicling how and why the concept of race arose, especially through Biblical history and the curse of Ham. However, she takes it too far and claims that race does not exist, nor is it a significant biological reality since there are no “race-specific genes” (also remember that you do not need genes to delineate race, using differences in physical traits and then correlating them to geography is sufficient) and there “are no clean breaks between human populations“. These fallacies aside, it is possible, as I have noted before, to denote racial classifications sans the use of “intelligence” or “mores” in the concept. Skin color is just one of many observable traits that differ by geography that make the basis for separating groups on the basis of race.
The minimalist race concept from Hardimon is non-hierarchical: meaning that it doesn’t discuss anything that would put races in a hierarchy like the racialist concept does (with mores and “intelligence”). If anything, this strictly physical definition of races (and the simple argument for it) should be enough to sway race-deniers to become race-believers.
Race, Testosterone, Aggression, and Prostate Cancer
4050 words
Race, aggression, and prostate cancer are all linked, with some believing that race is the cause of higher testosterone which then causes aggression and higher rates of crime along with maladies such as prostate cancer. These claims have long been put to bed, with a wide range of large analyses.
The testosterone debate regarding prostate cancer has been raging for decades and we have made good strides in understanding the etiology of prostate cancer and how it manifests. The same holds true for aggression. But does testosterone hold the key to understanding aggression, prostate cancer and does race dictate group levels of the hormone which then would explain some of the disparities between groups and individuals of certain groups?
Prostate cancer
For decades it was believed that heightened levels of testosterone caused prostate cancer. Most of the theories to this day still hold that large amounts of androgens, like testosterone and it’s metabolic byproduct dihydrotestosterone, are the two many factors that drive the proliferation of cells and therefore, if a male is exposed to higher levels of testosterone throughout their lives then they are at a high risk of prostate cancer compared to a man with low testosterone levels, so the story goes.
In 1986 Ronald Ross set out to test a hypothesis: that black males were exposed to more testosterone in the womb and this then drove their higher rates of prostate cancer later in life. He reportedly discovered that blacks, after controlling for confounds, had 15 percent higher testosterone than whites which may be the cause of differential prostate cancer mortality between the two races (Ross et al, 1986) This is told in a 1997 editorial by Hugh McIntosh. First, the fact that black males were supposedly exposed to more testosterone in the womb is brought up. I am aware of one paper discussing higher levels of testosterone in black women compared to white women (Perry et al, 1996). Though, I’ve shown that black women don’t have high levels of testosterone, not higher than white women, anyway (see Mazur, 2016 for discussion). (Yes I changed my view on black women and testosterone, stop saying that they have high levels of testosterone it’s just not true. I see people still link to that article despite the long disclaimer at the top.)
Alvarado (2013) discusses Ross et al (1986), Ellis and Nyborg (1992) (which I also discussed here along with Ross et al) and other papers discussing the supposed higher testosterone of blacks when compared to whites and attempts to use a life history framework to explain higher incidences of prostate cancer in black males. He first notes that nutritional status influences testosterone production which should be no surprise to anyone. He brings up some points I agree with and some I do not. For instance, he states that differences in nutrition could explain differences in testosterone between Western and non-Western people (I agree), but that this has no effect within Western countries (which is incorrect as I’ll get to later).
He also states that ancestry isn’t related to prostate cancer, writing “In summation, ancestry does not adequately explain variation among ethnic groups with higher or lower testosterone levels, nor does it appear to explain variation among ethnic groups with high or low prostate cancer rates. This calls into question the efficacy of a disease model that is unable to predict either deleterious or protective effects.”
He then states that SES is negatively correlated with prostate cancer rates, and that numerous papers show that people with low SES have higher rates of prostate cancer mortality which makes sense, since people in a lower economic class would have less access to and a chance to get good medical care to identify problems such as prostate cancer, including prostate biopsies and checkups to identify the condition.
He finally discusses the challenge hypothesis and prostate cancer risk. He cites studies by Mazur and Booth (who I’ve cited in the past in numerous articles) as evidence that, as most know, black-majority areas have more crime which would then cause higher levels of testosterone production. He cites Mazur’s old papers showing that low-class men, no matter if they’re white or black, had heightened levels of testosterone and that college-educated men did not, which implies that the social environment can and does elevate testosterone levels and can keep them heightened. Alvarado concludes this section writing: “Among Westernized men who have energetic resources to support the metabolic costs associated with elevated testosterone, there is evidence that being exposed to a higher frequency of aggressive challenges can result in chronically elevated testosterone levels. If living in an aggressive social environment contributes to prostate cancer disparities, this has important implications for prevention and risk stratification.” He’s not really wrong but on what he is wrong I will discuss later on this section. It’s false that testosterone causes prostate cancer so some of this thesis is incorrect.
I rebutted Ross et al (1986) December of last year. The study was hugely flawed and, yet, still gets cited to this day including by Alvarado (2013) as the main point of his thesis. However, perhaps most importantly, the assay times were done ‘when it was convenient’ for the students which were between 10 am and 3 pm. To not get any wacky readings one most assay the individuals as close to 8:30 am as possible. Furthermore, they did not control for waist circumference which is another huge confound. Lastly, the sample was extremely small (50 blacks and 50 whites) and done on a nonrepresentative sample (college students). I don’t think anyone can honestly cite this paper as any evidence for blacks having higher levels of testosterone or testosterone causing prostate cancer because it just doesn’t do that. (Read Race, Testosterone and Prostate Cancer for more information.)
What may explain prostate cancer rates if not for differences in testosterone like has been hypothesized for decades? Well, as I have argued, diet explains a lot of the variation between races. The etiology of prostate cancer is not known (ACA, 2016) but we know that it’s not testosterone and that diet plays a large role in its acquisition. Due to their dark skin, they need more sunlight than do whites to synthesize the same amount of vitamin D, and low levels of vitamin D in blacks are strongly related to prostate cancer (Harris, 2006). Murphy et al (2014) even showed, through biopsies, that black American men had higher rates of prostate cancer if they had lower levels of vitamin D. Lower concentrations of vitamin D in blacks compared to whites due to dark pigmentation which causes reduced vitamin D photoproduction and may also account for “much of the unexplained survival disparity after consideration of such factors as SES, state at diagnosis and treatment” (Grant and Peiris, 2012).
Testosterone
As mentioned above, testosterone is assumed to be higher in certain races compared to others (based on flawed studies) which then supposedly exacerbates prostate cancer. However, as can be seen above, a lot of assumptions go into the testosterone-prostate cancer hypothesis which is just false. So if the assumptions are false about testosterone, mainly regarding racial differences in the hormone and then what the hormone actually does, then most of their claims can be disregarded.
Perhaps the biggest problem is that Ross et al is a 32-year-old paper (which still gets cited favorably despite its huge flaws) while our understanding of the hormone and its physiology has made considerable progress in that time frame. So it’s in fact not so weird to see papers like this that say “Prostate cancer appears to be unrelated related to endogenous testosterone levels” (Boyle et al, 2016). Other papers also show the same thing, that testosterone is not related to prostate cancer (Stattin et al, 2004; Michaud, Billups, and Partin, 2015). This kills a lot of theories and hypotheses, especially regarding racial differences in prostate cancer acquisition and mortality. So, what this shows is that even if blacks did have 15 percent higher serum testosterone than whites as Ross et al, Rushton, Lynn, Templer, et al believed then it wouldn’t cause higher levels of prostate cancer (nor aggression, which I’ll get into later).
How high is testosterone in black males compared to white males? People may attempt to cite papers like the 32-year-old paper by Ross et al, though as I’ve discussed numerous times the paper is highly flawed and should therefore not be cited. Either way, levels are not as high as people believe and meta-analyses and actual nationally representative samples (not convenience college samples) show low to no difference, and even the low difference wouldn’t explain any health disparities.
One of the best papers on this matter of racial differences in testosterone is Richard et al (2014). They meta-analyzed 15 studies and concluded that the “racial differences [range] from 2.5 to 4.9 percent” but “this modest difference is unlikely to explain racial differences in disease risk.” This shows that testosterone isn’t as high in blacks as is popularly misconceived, and that, as I will show below, it wouldn’t even cause higher rates of aggression and therefore criminal behavior. (Rohrmann et al 2007 show no difference in testosterone between black and white males in a nationally representative sample after controlling for lifestyle and anthropometric variables. Whereas Mazur, 2009 shows that blacks have higher levels of testosterone due to low marriage rates and lower levels of adiposity, while be found a .39 ng/ml difference between blacks and whites aged 20 to 60. Is this supposed to explain crime, aggression, and prostate cancer?)
However, as I’ve noted last year (and as Alvarado, 2013 did as well), young black males with low education have higher levels of testosterone which is not noticed in black males of the same age group but with more education (Mazur, 2016). Since blacks of a similar age group have lower levels of testosterone but are more highly educated then this is a clue that education drives aggression/testosterone/violent behavior and not that testosterone drives it.
Mazur (2016) also replicated Assari, Caldwell, and Zimmerman’s (2014) finding that “Our model in the male sample suggests that males with higher levels of education has lower aggressive behaviors. Among males, testosterone was not associated with aggressive behaviors.” I know this is hard for many to swallow that testosterone doesn’t lead to aggressive behavior in men, but I’ll cover that in the last and final section.
So it’s clear that the myth that Rushton, Lynn, Templer, Kanazawa, et al pushed regarding hormonal differences between the races are false. It’s also with noting, as I did in my response to Rushton on r/K selection theory, that the r/K model is literally predicated on 1) testosterone differences between races being real and in the direction that Rushton and Lynn want because they cite the highly flawed Ross et al (1986) and 2) testosterone does not cause higher levels of aggression (which I’ll show below) which then lead to higher rates of crime along with higher rates of incarceration.
A blogger who goes by the name of ethnicmuse did an analysis of numerous testosterone papers and he found:
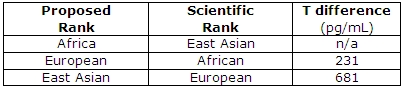 Which, of course, goes against a ton of HBD theory, that is, if testosterone did what HBDers believed it does (it doesn’t). This is what it comes down to: blacks don’t have higher levels of testosterone than whites and testosterone doesn’t cause aggression nor prostate cancer so even if this relationship was in the direction that Rushton et al assert then it still wouldn’t cause any of the explanatory variables they discuss.
Which, of course, goes against a ton of HBD theory, that is, if testosterone did what HBDers believed it does (it doesn’t). This is what it comes down to: blacks don’t have higher levels of testosterone than whites and testosterone doesn’t cause aggression nor prostate cancer so even if this relationship was in the direction that Rushton et al assert then it still wouldn’t cause any of the explanatory variables they discuss.
Last year Lee Ellis published a paper outlining his ENA theory (Ellis, 2017). I responded to the paper and pointed out what he got right and wrong. He discussed strength (blacks aren’t stronger than whites due to body type and physiology, but excel in other areas); circulating testosterone, umbilical cord testosterone exposure; bone density and crime; penis size, race, and crime (Rushton’s 1997 claims on penis size don’t ‘size up’ to the literature as I’ve shown two times); prostate-specific antigens, race, and prostate cancer; CAG repeats; intelligence and education and ‘intelligence’; and prenatal androgen exposure. His theory has large holes and doesn’t line up in some places, as he himself admits in his paper. He, as expected, cites Ross et al (1986) favorably in his analysis.
Testosterone can’t explain all of these differences, no matter if it’s prenatal androgen exposure or not, and a difference of 2.5 to 4.9 percent between blacks and whites regarding testosterone (Richard et al, 2014) won’t explain differences in crime, aggression, nor prostate cancer.
Other authors have attempted to also implicate testosterone as a major player in a wide range of evolutionary theories (Lynn, 1990; Rushton, 1997; Rushton, 1999; Hart, 2007; Rushton and Templer, 2012; Ellis, 2017). However, as can be seen by digging into this literature, these claims are not true and therefore we can discard the conclusions come to by the aforementioned authors since they’re based on false premises (testosterone being a cause for aggression, crime, and prostate cancer and r/K meaning anything to human races, it doesn’t)
Finally, to conclude this section, does testosterone explain racial differences in crime? No, racial differences in testosterone, however small, cannot be responsible for the crime gap between blacks and whites.
Testosterone and aggression
Testosterone and aggression, are they linked? Can testosterone tell us anything about individual differences in aggressive behavior? Surprisingly for most, the answer seems to be a resounding no. One example is the castration of males. Does it completely take away the urge to act aggressively? No, it does not. What is shown when sex offenders are castrated is that their levels of aggression decrease, but importantly, they do not decrease to 0. Robert Sapolsky writes on page 96 of his book Behave: The Biology of Humans at Our Best and Worst (2017) (pg 96):
… the more experience a male has being aggressive prior to castration, the more aggression continues afterward. In other words, the less his being aggressive in the future requires testosterone and the more it’s a function of social learning.
He also writes (pg 96-97):
On to the next issue that lessens the primacy of testosterone: What do individual levels of testosterone have to do with aggression? If one person higher testosterone levels than another, or higher levels this week than last, are they more likely to be aggressive?
Initially the answer seemed to be yes, as studies showed correlation between individual differences in testosterone levels and levels of aggression. In a typical study, higher testosterone levels would be observed in those male prisoners with higher rates of aggression. But being aggressive stimulates testosterone secretion; no wonder more aggressive individuals had higher levels. Such studies couldn’t disentangle chickens and eggs.
Thus, a better question is whether differences in testosterone levels among individuals predict who will be aggressive. And among birds, fish, mammals, and especially other primates, the answer is generally no. This has been studied extensively in humans, examining a variety of measures of aggression. And the answer is clear. To quote British endocrinologist John Archer in a definitive 2006 review, “There is a weak and inconsistent association between testosterone levels and aggression in [human] adults, and . . . administration of testosterone to volunteers typically does not increase aggression.” The brain doesn’t pay attention to testosterone levels within the normal range.
[…]
Thus, aggression is typically more about social learning than about testosterone, differing levels of testosterone generally can’t explain why some individuals are more aggressive than others.
Sapolsky also has a 1997 book of essays on human biology titled The Trouble With Testosterone: And Other Essays On The Biology Of The Human Predicament and he has a really good essay on testosterone titled Will Boys Just Be Boys? where he writes (pg 113 to 114):
Okay, suppose you note a correlation between levels of aggression and levels of testosterone among these normal males. This could be because (a) testosterone elevates aggression; (b) aggression elevates testosterone secretion; (c) neither causes the other. There’s a huge bias to assume option a while b is the answer. Study after study has shown that when you examine testosterone when males are first placed together in the social group, testosterone levels predict nothing about who is going to be aggressive. The subsequent behavioral differences drive the hormonal changes, not the other way around.
Because of a strong bias among certain scientists, it has taken do forever to convince them of this point.
[…]
As I said, it takes a lot of work to cure people of that physics envy, and to see interindividual differences in testosterone levels don’t predict subsequent differences in aggressive behavior among individuals. Similarly, fluctuations in testosterone within one individual over time do not predict subsequent changes in the levels of aggression in the one individual—get a hiccup in testosterone secretion one afternoon and that’s not when the guy goes postal.
And on page 115 writes:
You need some testosterone around for normal levels of aggressive behavior—zero levels after castration and down it usually goes; quadruple it (the sort of range generated in weight lifters abusing anabolic steroids), and aggression typically increases. But anywhere from roughly 20 percent of normal to twice normal and it’s all the same; the brain can’t distinguish among this wide range of basically normal values.
Weird…almost as if there is a wide range of ‘normal’ that is ‘built in’ to our homeodynamic physiology…
So here’s the point: differences in testosterone between individuals tell us nothing about individual differences in aggressive behavior; castration and replacement seems to show that, however broadly, testosterone is related to aggression “But that turns out to not be true either, and the implications of this are lost on most people the first thirty times you tell them about it. Which is why you’d better tell them about it thirty-one times, because it’s the most important part of this piece” (Sapolsky, 1997: 115).
Later in the essay, Sapolsky discusses a discusses 5 monkeys that were given time to form a hierarchy of 1 through 5. Number 3 can ‘throw his weight’ around with 4 and 5 but treads carefully around 1 and 2. He then states to take the third-ranking monkey and inject him with a ton of testosterone, and that when you check the behavioral data that he’d then be participating in more aggressive actions than before which would imply that the exogenous testosterone causes participation in more aggressive behavior. But it’s way more nuanced than that.
So even though small fluctuations in the levels of the hormone don’t seem to matter much, testosterone still causes aggression. But that would be wrong. Check out number 3 more closely. Is he now raining aggression and terror on any and all in the group, frothing in an androgenic glaze of indiscriminate violence. Not at all. He’s still judiciously kowtowing to numbers 1 and 2 but has simply become a total bastard to number 4 and 5. This is critical: testosterone isn’t causing aggression, it’s exaggerating the aggression that’s already there.
The correlation between testosterone and aggression is between .08 and .14 (Book, Starzyk, and Quinsey, 2001; Archer, Graham-Kevan, and Davies, 2005; Book and Quinsey, 2005). Therefore, along with all of the other evidence provided in this article, it seems that testosterone and aggression have a weak positive correlation, which buttresses the point that aggression concurrent increases in testosterone.
Sapolsky then goes on to discuss the amygdala’s role in fear processing. The amygdala has its influence on aggressive behavior through the stria terminalis, which is a bunch of neuronal connections. How the amygdala influences aggression is simple: bursts of electrical excitation called action potentials go up and down the stria terminalis which changes the hypothalamus. You can then inject testosterone right into the brain and will it cause the same action potentials that surge down the stria terminalis? No, it does not turn on the pathway at all. This only occurs only if the amygdala is already sending aggression-provoking action potentials down the stria terminalis with testosterone increasing the rate of action potentials you’re shortening the rest time between them. So it doesn’t turn on this pathway, it exaggerates the preexisting pattern, which is to say, it’s exaggerating the response to environmental triggers of what caused the amygdala to get excited in the first place.
He ends this essay writing (pg 119):
Testosterone is never going to tell us much about the suburban teenager who, in his after-school chess club, has developed a particularly aggressive style with his bishops. And it certainly isn’t going to tell us much about the teenager in some inner-city hellhole who has taken to mugging people. “Testosterone equals aggression” is inadequate for those who would offer a simple solution to the violent male—just decrease levels of those pesky steroids. And “testosterone equals aggression” is certainly inadequate for those who would offer a simple excuse: Boys will be boys and certain things in nature are inevitable. Violence is more complex than a single hormone. This is endocrinology for the bleeding heart liberal—our behavioral biology is usually meaningless outside of the context of social factors and the environment in which it occurs.
Injecting individuals with supraphysiological doses of testosterone as high as 200 and 600 mg per week does not cause heightened anger or aggression (Tricker et al, 1996; O’Connor et, 2002). This, too, is a large blow for the testosterone-induces-aggression hypothesis. Because aggressive behavior heightens testosterone, testosterone doesn’t heighten aggressive behavior. (This is the causality that has been looked for, and here it is. The causality is not in the other direction.) This tells us that we need to be put into situations for our aggression to rise and along with it, testosterone. I don’t even see how people could think that testosterone could cause aggression. It’s obvious that the environmental trigger needs to be there first in order for the body’s physiology to begin testosterone production in order to prepare for the stimulus that caused the heightened testosterone production. Once the trigger occurs, then it can and does stay heightened, especially in areas where dominance contests would be more likely to occur, which would be low-income areas (Mazur, 2006, 2016).
(Also read my response to Batrinos, 2012, my musings on testosterone and race, and my responses to Robert Lindsay and Sean Last.)
Lastly, one thing that gets on my nerves that people point to to attempt to show that testosterone and its derivatives cause violence, aggression etc is the myth of “roid rage” which is when an individual objects himself with testosterone, anabolic steroids or another banned substance, and then the individual becomes more aggressive as a result of more free-flowing testosterone in their bloodstream.
The problem here is that people believe what they hear on the media about steroids and testosterone, and they’re largely not true. One large analysis was done to see the effects of steroids and other illicit drug use on behavior, and what was found was that after controlling for other substance use “Our results suggest that it was not lifetime steroid use per se, but rather co-occurrring polysubstance abuse that most parsimoniously explains the relatively strong association of steroid use and interpersonal violence” (Lundholm et al, 2015). So after controlling for other drugs used, men who use steroids do not go to prison and be convicted of violence after other polysubstance use was controlled for, implying that is what’s driving interpersonal violence, not the substance abuse of steroids.
Conclusion
Numerous myths about testosterone have been propagated over the decades, which are still believed in the new millennium despite numerous other studies and arguments to the contrary. As can be seen, the myths that people believe about testosterone are easily debunked. Numerous papers (with better methodology than Ross et al) attest to the fact that testosterone levels aren’t as high as was believed decades ago between the races. Diet can explain a lot of the variation, especially vitamin D intake. Injecting men with supraphysiological doses of testosterone does not heighten anger nor aggression. It does not even heighten prostate cancer severity.
Racial differences in testosterone are also not as high as people would like to believe, there is even an opposite relationship with Asians having higher levels and whites having lower (which wouldn’t, on average, imply femininity) testosterone levels. So as can be seen, the attempted r/K explanations from Rushton et al don’t work out here. They’re just outright wrong on testosterone, as I’ve been arguing for a long while on this blog.
Testosterone doesn’t cause aggression, aggression causes heightened testosterone. It can be seen from studies of men who have been castrated that the more crime they committed before castration, the more crime they will commit after which implies a large effect of social learning on violent behavior. Either way, the alarmist attitudes of people regarding testosterone, as I have argued, are not needed because they’re largely myths.
The Evolution of Human Skin Variation
4050 words
Human skin variation comes down to how much UV radiation a population is exposed to. Over time, this leads to changes in genetic expression. If that new genotype is advantageous in that environment, it will get selected for. To see how human skin variation evolved, we must first look to chimpanzees since they are our closest relative.
The evolution of black skin
Humans and chimps diverged around 6-12 mya. Since we share 99.8 percent of our genome with them, it’s safe to say that when we diverged, we had pale skin and a lot of fur on our bodies (Jablonski and Chaplin, 2000). After we lost the fur on our bodies, we were better able to thermoregulate, which then primed Erectus for running (Liberman, 2015). The advent of fur loss coincides with the appearance of sweat glands in Erectus, which would have been paramount for persistence hunting in the African savanna 1.9 mya, when a modern pelvis—and most likely a modern gluteus maximus—emerged in the fossil record (Lieberman et al, 2006). This sets the stage for one of the most important factors in regards to the ability to persistence hunt—mainly, the evolution of dark skin to protect against high amounts of UV radiation.
After Erectus lost his fur, the unforgiving UV radiation beamed down on him. Selection would have then occurred for darker skin, as darker skin protects against UV radiation. Dark skin in our genus also evolved between 1 and 2 mya. We know this since the melanocortin 1 receptor promoting black skin arose 1-2 mya, right around the time Erectus appeared and lost its fur (Lieberman, 2015).
However, other researchers reject Greaves’ explanation for skin cancer being a driver for skin color (Jablonksi and Chaplin, 2014). They cite Blum (1961) showing that skin cancer is acquired too late in life to have any kind of effect on reproductive success. Skin cancer rates in black Americans are low compared to white Americans in a survey from 1977-8 showing that 30 percent of blacks had basal cell carcinoma while 80 percent of whites did (Moon et al, 1987). This is some good evidence for Greaves’ hypothesis; that blacks have less of a rate of one type of skin cancer shows its adaptive benefits. Black skin evolved due to the need for protection from high levels of UVB radiation and skin cancers.
Highly melanized skin also protects against folate destruction (Jablonksi and Chaplin, 2000). As populations move away from high UV areas, the selective constraint to maintain high levels of folate by blocking high levels of UV is removed, whereas selection for less melanin prevails to allow enough radiation to synthesize vitamin D. Black skin is important near the equator to protect against folate deficiency. (Also see Nina Jablonski’s Ted Talk Skin color is an illusion.)
The evolution of white skin
The evolution of white skin, of course, is much debated as well. Theories range from sexual selection, to diet, to less UV radiation. All three have great explanatory power, and I believe that all of them did drive the evolution of white skin, but with different percentages.
The main driver of white skin is living in colder environments with fewer UV rays. The body needs to synthesize vitamin D, so the only way this would occur in areas with low UV rays.
White skin is a recent trait in humans, appearing only 8kya. A myriad of theories have been proposed to explain this, from sexual selection (Frost, 2007), which include better vitamin D synthesis to ensure more calcium for pregnancy and lactation (which would then benefit the intelligence of the babes) (Jablonski and Chaplin, 2000); others see light skin as the beginnings of more childlike traits such as smoother skin, a higher pitched voice and a more childlike face which would then facilitate less aggressiveness in men and more provisioning (Guthrie, 1970; from Frost, 2007); finally, van den Berghe and Frost (1986) proposed that selection for white skin involved unconscious selection by men for lighter-skinned women which is used “as a measure of hormonal status and thus childbearing potential” (Frost, 2007). The three aforementioned hypotheses have sexual selection for lighter skin as a proximate cause, but the ultimate cause is something completely different.
The hypothesis that white skin evolved to better facilitate vitamin D synthesis to ensure more calcium for pregnancy and lactation makes the most sense. Darker-skinned individuals have a myriad of health problems outside of their ancestral climate, one of which is higher rates of prostate cancer due to lack of vitamin D. If darker skin is a problem in cooler climates with fewer UV rays, then lighter skin, since it ensures better vitamin D synthesis, will be selected for. White skin ensures better and more vitamin D absorption in colder climates with fewer UV rays, therefore, the ultimate cause of the evolution of white skin is a lack of sunlight and therefore fewer UV rays. This is because white skin absorbs more UV rays which is better vitamin D synthesis.
Peter Frost believes that Europeans became white 11,000 years ago. However, as shown above, white skin evolved around 8kya. Further, contrary to popular belief, Europeans did not gain the alleles for white skin from Neanderthals (Beleza et al, 2012). European populations did not lose their dark skin immediately upon entering Europe—and Neanderthal interbreeding didn’t immediately confer the advantageous white skin alleles. There was interbreeding between AMH and Neanderthals (Sankararaman et al, 2014). So if interbreeding with Neanderthals didn’t infer white skin to proto-Europeans, then what did?
A few alleles spreading into Europe that only reached fixation a few thousand years ago. White skin is a relatively recent trait in Man (Beleza et al, 2012). People assume that white skin has been around for a long time, and that Europeans 40,000 ya are the ancestors of Europeans alive today. That, however, is not true. Modern-day European genetic history began about 6,500 ya. That is when the modern-day European phenotype arose—along with white skin.
Furthermore, Eurasians were still a single breeding population 40 kya, and only diverged recently, about 25,000 to 40,000 ya (Tateno et al, 2014). The alleles that code for light skin evolved after the Eurasian divergence. Polymorphisms in the genes ASIP and OCA2 may code for dark and light skin all throughout the world, whereas SLC24A5, MATP, and TYR have a predominant role in the evolution of light skin in Europeans but not East Asians, which suggests recent convergent evolution of a lighter pigmentation phenotype in European and East Asian populations (Norton et al, 2006). Since SLC24A5, MATP, and TYR are absent in East Asian populations, then that means that East Asians evolved light skin through completely different mechanisms than Europeans. So after the divergence of East Asians and Europeans from a single breeding population 25-40kya, there was convergent evolution for light pigmentation in both populations with the same selection pressure (low UV).
Some populations, such as Arctic peoples, don’t have the skin color one would predict they should have based on their ancestral environment. However, their diets are high in shellfish which is high in vitamin D, which means they can afford to remain darker-skinned in low UV areas. UV rays reflect off of the snow and ice in the summer and their dark skin protects them from UV light.
Black-white differences in UV absorption
If white skin evolved to better synthesize vitamin D with fewer (and less intense) UV rays, then those with blacker skin would need to spend a longer time in UV light to synthesize the same amount of vitamin D. Skin pigmentation, however, is negatively correlated with vitamin D synthesis (Libon, Cavalier, and Nikkels, 2013). Black skin is less capable of vitamin D synthesis. Furthermore, blacks’ skin color leads to an evolutionary environmental mismatch. Black skin in low UV areas is correlated with rickets (Holick, 2006), higher rates of prostate cancer due to lower levels of vitamin D (Gupta et al, 2009; vitamin D supplements may also keep low-grade prostate cancer at bay).
Libon, Cavalier, and Nikkels, (2013) looked at a few different phototypes (skin colors) of black and white subjects. The phototypes they looked at were II (n=19), III (n=1), and VI (n-11; whites and blacks respectively). Phototypes are shown in the image below.
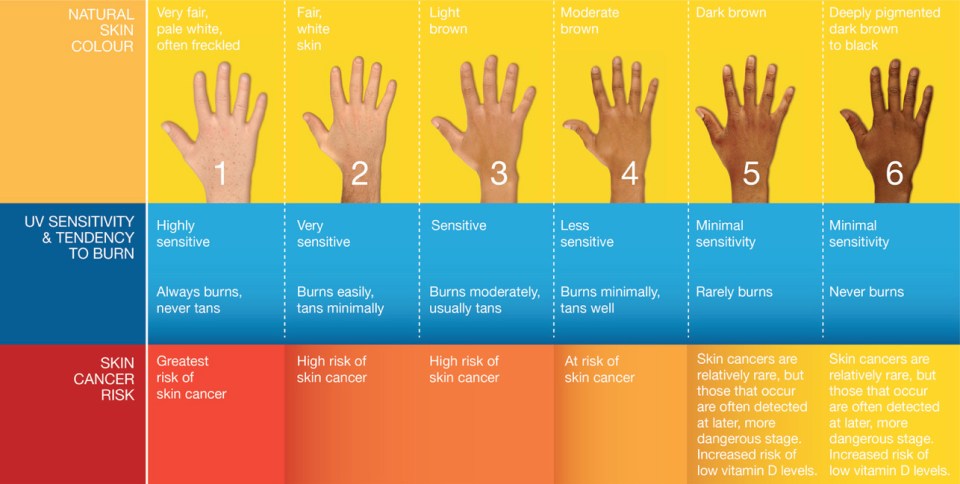

To avoid the influence of solar UVB exposure, this study was conducted in February. On day 0, both the black and white subjects were vitamin D deficient. The median levels of vitamin D in the white subjects was 11.9 ng/ml whereas for the black subjects it was 8.6 ng/ml—a non-statistically significant difference. On day two, however, concentrations of vitamin D in the blood rose from 11.9 to 13.3 ng/ml—a statistically significant difference. For the black cohort, however, there was no statistically significant difference in vitamin D levels. On day 6, levels in the white subjects rose from 11.6 to 14.3 ng/ml whereas for the black subjects it was 8.6 to 9.57 ng/ml. At the end of day 6, there was a statistically significant difference in circulating vitamin D levels between the white and black subjects (14.3 ng/ml compared to 9.57 ng/ml).
Different phototypes absorb different amounts of UV rays and, therefore, peoples with different skin color absorb different levels of vitamin D. Lighter-skinned people absorb more UV rays than darker-skinned people, showing that white skin’s primary cause is to synthesize vitamin D.
UVB exposure increases vitamin D production in white skin, but not in black skin. Pigmented skin, on the other hand, hinders the transformation of 7-dehydrocholesterol to vitamin D. This is why blacks have higher rates of prostate cancer—they are outside of their ancestral environment and what comes with being outside of one’s ancestral environment are evolutionary mismatches. We have now spread throughout the world, and people with certain skin colors may not be adapted for their current environment. This is what we see with black Americans as well as white Americans who spend too much time in climes that are not ancestral to them. Nevertheless, different-colored skin does synthesize vitamin D differently, and knowledge of this will increase the quality of life for everyone.
Even the great Darwin wrote about differences in human skin color. He didn’t touch human evolution in On the Origin of Species (Darwin, 1859), but he did in his book Descent of Man (Darwin, 1871). Darwin talks about the effects of climate on skin color and hair, writing:
It was formerly thought that the colour of the skin and the character of the hair were determined by light or heat; and although it can hardly be denied that some effect is thus produced, almost all observers now agree that the effect has been very small, even after exposure during many ages. (Darwin, 1871: 115-116)
Darwin, of course, championed sexual selection as the cause for human skin variation (Darwin, 1871: 241-250). Jared Diamond has the same view, believing that natural selection couldn’t account for hair loss, black skin and white skin weren’t products of natural selection, but female mate preference and sexual selection (Greaves, 2014).
Parental selection for white skin
Judith Rich Harris, author of the book The Nurture Assumption: Why Kids Turn Out the Way They Do (Harris, 2009), posits another hypothesis for the evolution of light skin for those living in northern latitudes—parental selection. This hypothesis may be controversial to some, as it states that dark skin is not beautiful and that white skin is.
Harris posits that selection for lighter skin was driven by sexual selection, but states that parental selection for lighter skin further helped the fixation of the alleles for white skin in northern populations. Neanderthals were a furry population, as they had no clothes, so, logic dictates that if they didn’t have clothes then they must have had some sort of protection against the cold Ice Age climate, therefore they must have had fur.
Harris states that since lighter skin is seen as more beautiful than darker skin, then if a woman birthed a darker/furrier babe than the mother would have committed infanticide. Women who birth at younger ages are more likely to commit infanticide, as they still have about twenty years to birth a babe. On the other hand, infanticide rates for mothers decrease as she gets older—because it’s harder to have children the older you get.
Harris states that Erectus may have been furry up until 2 mya, however, as I’ve shown, Erectus was furless and had the ability to thermoregulate—something that a hairy hominin was not able to do (Lieberman, 2015).
There is a preference for lighter-skinned females all throughout the world, in Africa (Coetzee et al, 2012); China and India (Naidoo et al, 2016; Dixson et al, 2007); and Latin America and the Philipines (Kiang and Takeuchi, 2009). Light skin is seen as attractive all throughout the world. Thus, since light skin allows better synthesize of vitamin D in colder climes with fewer UV rays, then there would have been a myriad of selective pressures to push that along—parental selection for lighter-skinned babes being one of them. This isn’t talked about often, but infanticide and rape have both driven our evolution (more on both in the future).
Harris’ parental selection hypothesis is plausible, and she does use the right dates for fur loss which coincides with the endurance running of Erectus and how he was able to thermoregulate body heat due to lack of fur and more sweat glands. This is when black skin began to evolve. So with migration into more northerly climes, lighter-skinned people would have more of an advantage than darker-skinned people. Infanticide is practiced all over the world, and is caused—partly—by a mother’s unconscious preferences.
Skin color and attractiveness
Lighter skin is seen as attractive all throughout the world. College-aged black women find lighter skin more attractive (Stephens and Thomas, 2012). It is no surprise that due to this, a lot of black women lighten their skin with chemicals.
In a sample of black men, lighter-skinned blacks were more likely to perceive discrimination than their darker-skinned counterparts (Uzogara et al, 2014). Further, in appraising skin color’s effect on in-group discrimination, medium-skinned black men perceived less discrimination than lighter- and darker-skinned black men. Lastly—as is the case with most studies—this effect was particularly pronounced for those in lower SES brackets. Speaking of SES, lighter-skinned blacks with higher income had lower blood pressure than darker-skinned blacks with higher income (Sweet et al, 2007). The authors conclude that a variety of psychosocial stress due to discrimination must be part of the reason why darker-skinned blacks with a high SES have worse blood pressure—but I think there is something else at work here. Darker skin on its own is associated with high blood pressure (Mosley et al, 2000). I don’t deny that (perceived) discrimination can and does heighten blood pressure—but the first thing that needs to be looked at is skin color.
Lighter-skinned women are seen as more attractive (Stephen et al, 2009). This is because it signals fertility, femininity, and youth. One more important thing it signals is the ability to carry a healthy child to term since lighter skin in women is associated with better vitamin D synthesis which is important for a growing babe.
Skin color and intelligence
There is a high negative correlation between skin color and intelligence, about –.92 (Templer and Arikawa, 2006). They used the data from Lynn and Vanhanen’s 2002 book IQ and the Wealth of Nations and found that there was an extremely strong negative correlation between skin color and IQ. However, data wasn’t collected for all countries tested and for half of the countries the IQs were ‘estimated’ from other surrounding countries’ IQs.
Jensen (2006) states that the main limitation in the study design of Arikawa and Templer (2006) is that “correlations obtained from this type of analysis are completely non-informative regarding any causal or functional connection between individual differences in skin pigmentation and individual differences in IQ, nor are they informative regarding the causal basis of the correlation, e.g., simple genetic association due to cross-assortative mating for skin color and IQ versus a pleiotropic correlation in which both of the phenotypically distinct but correlated traits are manifested by one and the same gene.”
Lynn (2002) purported to find a correlation of .14 in a representative sample of American blacks (n=430), concluding that the proportion of European genes in African Americans dictates how intelligent that individual black is. However, Hill (2002) showed that when controlling for childhood environmental factors such as SES, the correlation disappears and therefore, a genetic causality cannot be inferred from the data that Lynn (2002) used.
Since Lynn found a .14 correlation between skin color and IQ in black Americans, that means that only .0196 percent of the variation in IQ within black American adults can be explained by skin color. This is hardly anything to look at and keep in mind when thinking about racial differences in IQ.
However, other people have different ideas. Others may say that since animal studies find that lighter animals are less sexually active, are less aggressive, have a larger body mass, and greater stress resistance. So since this is seen in over 40 species of vertebrate, some fish species, and over 30 bird species (Rushton and Templer, 2012) that means that it should be a good predictor for human populations. Except it isn’t.
we know the genetic architecture of pigmentation. that is, we know all the genes (~10, usually less than 6 in pairwise between population comparisons). skin color varies via a small number of large effect trait loci. in contrast, I.Q. varies by a huge number of small effect loci. so logically the correlation is obviously just a correlation. to give you an example, SLC45A2 explains 25-40% of the variance between africans and europeans.
long story short: it’s stupid to keep repeating the correlation between skin color and I.Q. as if it’s a novel genetic story. it’s not. i hope don’t have to keep repeating this for too many years.
Finally, variation in skin color between human populations are primarily due to mutations on the genes MC1R, TYR, MATP (Graf, Hodgson, and Daal, 2005), and SLC24A5 (also see Lopez and Alonso, 2014 for a review of genes that account for skin color) so human populations aren’t “expected to consistently exhibit the associations between melanin-based coloration and the physiological and behavioural traits reported in our study” (Ducrest, Keller, and Roulin, 2008). Talking about just correlations is useless until causality is established (if it ever is).
Conclusion
The evolution of human skin variation is complex and is driven by more than one variable, but some are stronger than others. The evolution of black skin evolved—in part—due to skin cancer after we lost our fur. White skin evolved due to sexual selection (proximate cause) and to better absorb UV rays for vitamin D synthesis in colder climes (the true need for light skin in cold climates). Eurasians split around 40kya, and after this split both evolved light skin pigmentation independently. As I’ve shown, the alleles that code for skin color between blacks and whites don’t account for differences in aggression, nor do they account for differences in IQ. The genes that control skin color (about a dozen) pale in comparison to the genes that control intelligence (thousands of genes with small effects). Some other hypotheses for the evolution of white skin are on par with being as controversial as the hypothesis that skin color and intelligence co-evolved—mainly that mothers would kill darker-skinned babies because they weren’t seen as beautiful as lighter-skinned babies.
The evolution of human skin variation is extremely interesting with many competing hypotheses, however, to draw wild conclusions based on just correlations in regards to human skin color and intelligence and aggression, you’re going to need more evidence than just correlations.
References
Bang KM, Halder RM, White JE, Sampson CC, Wilson J. 1987. Skin cancer in black Americans: A review of 126 cases. J Natl Med Assoc 79:51–58
Beleza, S., Santos, A. M., Mcevoy, B., Alves, I., Martinho, C., Cameron, E., . . . Rocha, J. (2012). The Timing of Pigmentation Lightening in Europeans. Molecular Biology and Evolution,30(1), 24-35. doi:10.1093/molbev/mss207
Blum, H. F. (1961). Does the Melanin Pigment of Human Skin Have Adaptive Value?: An Essay in Human Ecology and the Evolution of Race. The Quarterly Review of Biology,36(1), 50-63. doi:10.1086/403275
Coetzee V, Faerber SJ, Greeff JM, Lefevre CE, Re DE, et al. (2012) African perceptions of female attractiveness. PLOS ONE 7: e48116.
Darwin, C. (1859). On the origin of species by means of natural selection, or, the preservation of favoured races in the struggle for life. London: J. Murray.
Darwin, C. (1871). The descent of man, and selection in relation to sex. London: John Murray, Albemarle Street.
Dixson, B. J., Dixson, A. F., Li, B., & Anderson, M. (2006). Studies of human physique and sexual attractiveness: Sexual preferences of men and women in China. American Journal of Human Biology,19(1), 88-95. doi:10.1002/ajhb.20584
Ducrest, A., Keller, L., & Roulin, A. (2008). Pleiotropy in the melanocortin system, coloration and behavioural syndromes. Trends in Ecology & Evolution,23(9), 502-510. doi:10.1016/j.tree.2008.06.001
Frost, P. (2007). Human skin-color sexual dimorphism: A test of the sexual selection hypothesis. American Journal of Physical Anthropology,133(1), 779-780. doi:10.1002/ajpa.20555
Graf, J., Hodgson, R., & Daal, A. V. (2005). Single nucleotide polymorphisms in theMATP gene are associated with normal human pigmentation variation. Human Mutation,25(3), 278-284. doi:10.1002/humu.20143
Greaves, M. (2014). Was skin cancer a selective force for black pigmentation in early hominin evolution? Proceedings of the Royal Society B: Biological Sciences,281(1781), 20132955-20132955. doi:10.1098/rspb.2013.2955
Gupta, D., Lammersfeld, C. A., Trukova, K., & Lis, C. G. (2009). Vitamin D and prostate cancer risk: a review of the epidemiological literature. Prostate Cancer and Prostatic Diseases,12(3), 215-226. doi:10.1038/pcan.2009.7
Guthrie RD. 1970. Evolution of human threat display organs. Evol Biol 4:257–302.
Harris, J. R. (2006). Parental selection: A third selection process in the evolution of human hairlessness and skin color. Medical Hypotheses,66(6), 1053-1059. doi:10.1016/j.mehy.2006.01.027
Harris, J. R. (2009). The nurture assumption: why children turn out the way they do. New York: Free Press.
Hill, Mark E. 2002. Skin color and intelligence in African Americans: A reanalysis of Lynn’s data. Population and Environment 24, no. 2:209–14
Holick, M. F. (2006). Resurrection of vitamin D deficiency and rickets. Journal of Clinical Investigation,116(8), 2062-2072. doi:10.1172/jci29449
Jablonski, N. G., & Chaplin, G. (2000). The evolution of human skin coloration. Journal of Human Evolution,39(1), 57-106. doi:10.1006/jhev.2000.0403
Jablonski, N. G., & Chaplin, G. (2014). Skin cancer was not a potent selective force in the evolution of protective pigmentation in early hominins. Proceedings of the Royal Society B: Biological Sciences,281(1789), 20140517-20140517. doi:10.1098/rspb.2014.0517
Jensen, A. R. (2006). Comments on correlations of IQ with skin color and geographic–demographic variables. Intelligence,34(2), 128-131. doi:10.1016/j.intell.2005.04.003
Kiang, L., & Takeuchi, D. T. (2009). Phenotypic Bias and Ethnic Identity in Filipino Americans. Social Science Quarterly,90(2), 428-445. doi:10.1111/j.1540-6237.2009.00625.x
Libon, F., Cavalier, E., & Nikkels, A. (2013). Skin Color Is Relevant to Vitamin D Synthesis. Dermatology,227(3), 250-254. doi:10.1159/000354750
Lieberman, D. E. (2015). Human Locomotion and Heat Loss: An Evolutionary Perspective. Comprehensive Physiology, 99-117. doi:10.1002/cphy.c140011
Lieberman, D. E., Raichlen, D. A., Pontzer, H., Bramble, D. M., & Cutright-Smith, E. (2006). The human gluteus maximus and its role in running. Journal of Experimental Biology,209(11), 2143-2155. doi:10.1242/jeb.02255
López, S., & Alonso, S. (2014). Evolution of Skin Pigmentation Differences in Humans. ELS. doi:10.1002/9780470015902.a0021001.pub2
Lynn, R. (2002). Skin color and intelligence in African Americans. Population and Environment, 23, 365–375.
Mosley, J. D., Appel, L. J., Ashour, Z., Coresh, J., Whelton, P. K., & Ibrahim, M. M. (2000). Relationship Between Skin Color and Blood Pressure in Egyptian Adults : Results From the National Hypertension Project. Hypertension,36(2), 296-302. doi:10.1161/01.hyp.36.2.296
Naidoo, L.; Khoza, N.; Dlova, N.C. A fairer face, a fairer tomorrow? A review of skin lighteners. Cosmetics 2016, 3, 33.
Norton, H. L., Kittles, R. A., Parra, E., Mckeigue, P., Mao, X., Cheng, K., . . . Shriver, M. D. (2006). Genetic Evidence for the Convergent Evolution of Light Skin in Europeans and East Asians. Molecular Biology and Evolution,24(3), 710-722. doi:10.1093/molbev/msl203
Rushton, J. P., & Templer, D. I. (2012). Do pigmentation and the melanocortin system modulate aggression and sexuality in humans as they do in other animals? Personality and Individual Differences,53(1), 4-8. doi:10.1016/j.paid.2012.02.015
Sankararaman, S., Mallick, S., Dannemann, M., Prüfer, K., Kelso, J., Pääbo, S., . . . Reich, D. (2014). The genomic landscape of Neanderthal ancestry in present-day humans. Nature,507(7492), 354-357. doi:10.1038/nature12961
Stephen, I. D., Smith, M. J., Stirrat, M. R., & Perrett, D. I. (2009). Facial Skin Coloration Affects Perceived Health of Human Faces. International Journal of Primatology,30(6), 845-857. doi:10.1007/s10764-009-9380-z
Stephens, D., & Thomas, T. L. (2012). The Influence of Skin Color on Heterosexual Black College Women’s Dating Beliefs. Journal of Feminist Family Therapy,24(4), 291-315. doi:10.1080/08952833.2012.710815
Sweet, E., Mcdade, T. W., Kiefe, C. I., & Liu, K. (2007). Relationships Between Skin Color, Income, and Blood Pressure Among African Americans in the CARDIA Study. American Journal of Public Health,97(12), 2253-2259. doi:10.2105/ajph.2006.088799
Tateno, Y., Komiyama, T., Katoh, T., Munkhbat, B., Oka, A., Haida, Y., . . . Inoko, H. (2014). Divergence of East Asians and Europeans Estimated Using Male- and Female-Specific Genetic Markers. Genome Biology and Evolution,6(3), 466-473. doi:10.1093/gbe/evu027
Templer, D. I., & Arikawa, H. (2006). Temperature, skin color, per capita income, and IQ: An international perspective. Intelligence,34(2), 121-139. doi:10.1016/j.intell.2005.04.002
Uzogara, E. E., Lee, H., Abdou, C. M., & Jackson, J. S. (2014). A comparison of skin tone discrimination among African American men: 1995 and 2003. Psychology of Men & Masculinity, 15(2), 201–212. http://doi.org/10.1037/a0033479
van den Berghe PL, Frost P. 1986. Skin color preference, sexual dimorphism and sexual selection: a case of gene-culture coevolution? Ethn Racial Stud 9:87–113.
