
Home » Posts tagged 'Evolution' (Page 2)
Tag Archives: Evolution
DNA is not a “Blueprint”
2200 words
Leading behavior geneticist Robert Plomin is publishing “Blueprint: How DNA Makes Us Who We Are” in October of 2018. I, of course, have not read the book yet. But if the main thesis of the book is that DNA is a “code”, “recipe”, or “blueprint”, then that is already wrong. This is because presuming that DNA is any of the three aforementioned things marries one to certain ideas, even if they themselves do not explicitly state them. Nevertheless, Robert Plomin is what one would term a “hereditarian”, meaning that he believes that genes—more than environment—shape an individual’s psychological and other traits. (That’s a false dichotomy, though.) In the preview for the book at MIT Press, they write:
In Blueprint, behavioral geneticist Robert Plomin describes how the DNA revolution has made DNA personal by giving us the power to predict our psychological strengths and weaknesses from birth. A century of genetic research shows that DNA differences inherited from our parents are the consistent life-long sources of our psychological individuality—the blueprint that makes us who we are. This, says Plomin, is a game-changer. It calls for a radical rethinking of what makes us who were are.
Genetics accounts for fifty percent of psychological differences—not just mental health and school achievement, but all psychological traits, from personality to intellectual abilities. Nature defeats nurture by a landslide.
Plomin explores the implications of this, drawing some provocative conclusions—among them that parenting styles don’t really affect children’s outcomes once genetics is taken into effect. Neither tiger mothers nor attachment parenting affects children’s ability to get into Harvard. After describing why DNA matters, Plomin explains what DNA does, offering readers a unique insider’s view of the exciting synergies that came from combining genetics and psychology.
I won’t get into most of these things today (I will wait until I read the book for that), but this will be just an article showing that DNA is, in fact, not a blueprint, and DNA is not a “code” or “recipe” for the organism.
It’s funny that the little blurb says that “Nature defeats nurture by a landslide“, because, as I have argued at length, nature vs nurture is a false dichotomy (See Oyama, 1985, 2000, 1999; Moore, 2002; Schneider, 2007; Moore, 2017). Nature vs nurture is the battleground that the false dichotomy of genes vs environment is fought on. However, it makes no sense to partition heritability estimates if it is indeed true that genes interact with environment—that is, if nature interacts with nurture.
DNA is also called “the book of life”. For example, in her book The Epigenetics Revolution: How Modern Biology Is Rewriting Our Understanding of Genetics, Disease, and Inheritance, Nessa Carey writes that “There’s no debate that the DNA blueprint is a starting point” (pg 16). This, though, can be contested. “But the promise of a peep into the ‘book of life’ leading to a cure for all diseases was a mistake” (Noble, 2017: 161).
Developmental psychologist and cognitive scientist David S. Moore concurs. In his book The Developing Genome: An Introduction to Behavioral Epigenetics, he writes (pg 45):
So, although I will talk about genes repeatedly in this book, it is only because there is no other convenient way to communicate about contemporary ideas in molecular biology. And when I refer to gebe, I will be talking about a segment or segments of DNA containing sequence information that is used to help construct a protein (or some other product that performs a biological function). But it is worth remembering that contemporary biologists do not mean any one thing when they talk about “genes”; the gene remains a fundementally hypothetical concept to this day. The common belief that there are things inside of us that constitute a set of instructions for building bodies and minds—things that are analogous to “blueprings” or “recipes”—is undoubedtly false. Instead, DNA segements often contain information that is ambiguous, and that must be edited or arranged in context-dependent ways before it can be used.
Still, other may use terms like “genes for” trait T. This, too, is incorrect. In his outstanding book Making Sense of Genes, Kostas Kamporakis writes (pg 19):
I also explain why the notion of “genes for,” in the vernacular sense, is not only misleading but also entirely inaccurate and scientifcally illegitamate.
[…]
First, I show that genes “operate” in the context of development only. This means that genes are impllicated in the development of characters but do not determine them. Second, I explain why single genes do not alone produce characters or disease but contribute to their variation. This means that genes can account for variation in characters but cannot alone explain their origin. Third, I show that genes are not the masters of the game but are subject to complex regulatory processes.
Genes can only be seen as passive templates, not ultimate causes (Noble, 2011), and they cannot explain the origin of different characters but can account for variation in physical characters. Genes only “do” something in the context of development; they are inert molecules and thusly cannot “cause” anything on their own.
Genes are not ‘for’ traits, but they are difference-makers for traits. Sterelny and Griffiths (1999: 102), in their book Sex and Death: An Introduction to Philosophy of Biology write:
Sterelny and Griffiths (1988) responded to the idea that genes are invisible to selection by treating genes as difference makers, and as visible to selection by virtue of the differences they make. In doing so, they provided a formal reconstruction of the “gene for” locution. The details are complex, but the basic intent of the reconstruction is simple. A certain allele in humans is an “allele for brown eyes” because, in standard environments, having that allele rather than alternatives typically available in the population means that your eyes will be brown rather than blue. This is the concpet of a gene as a difference maker. It is very important to note, however, that genes are context-sensitive difference makers. Their effects depend on the genetic, cellular, and other features of their environment.
(Genes can be difference makers for physical traits, but not for psychological traits because no psychophysical laws exist, but I’ll get to that in the future.)
Note how the terms “context-sensitive” and “context-dependent” continue to appear. The DNA-as-blueprint statement presumes that DNA is context-independent, but we cannot divorce genes—whatever they are—from their context, since genes and environment, nature and nurture, are intertwined. (And it is even questioned if ‘genes’ are truly units of inheritance, see Fogle, 1990. Fogle, 2000 also argues to dispense with the concept of “gene” and that biologists should be using terms like intron, promoter region, and exon. Nevertheless, there is a huge disconnect with the term “gene” in molecular biology and classical genetics. Keller 2000 argues that there are still uses for the term “gene” and that we should not dispense with the term. I believe we should dispense with it.)
Susan Oyama (2000: 77) writes in her book The Ontogeny of Information:
“Though a plan implies action, it does not itself act, so if the genes are a blueprint, something else is the constructor-construction worker. Though blueprints are usually contrasted with building materials, the genes are quite easily conceptualized as templates for building tools and materials; once so utilized, of course, they enter the developmental process and influence its course. The point of the blueprint analogy, though, does not seem to be to illuminate developmental processes, but rather to assume them and, in celebrating their regularity, to impute cognitive functions to genes. How these functions are exercised is left unclear in this type of metaphor, except that the genetic plan is seen in some peculiar way to carry itself out, generating all the necessary steps in the necessary sequence. No light is shed on multiple developmental possibilities, species-typical or atypical.“
The Modern Synthesis is one of the causes for the genes-as-blueprints thinking; the Modern Synthesis has causation in biology wrong. Genes are not active causes, but they are passive templates, as argued by many authors. They, thus, cannot “cause” anything on their own.
In his 2017 book Dance to the Tune of Life: Biological Relativity, Denis Noble writes (pg 157):
As we saw earlier in this chapter, these triplet sequences are formed from any combination of the four bases U, C, A and G in RNA and T, C, A and G in DNA. They are often described as a genetic ‘code’, but it is important to understand that this usage of the word ‘code’ carries overtones that can be confusing.
A code was originally an intentional encryption used by humans to communicate. The genetic ‘code’ is not intentional in that sense. The word ‘code’ has unfortunately reinforced the idea that genes are active and even complete causes, in much the same was as a computer is caused to follow the instructions of a computer program. The more nuetral word ‘template’ would be better. Templates are used only when required (activated); they are not themselves active causes. The active causes lie within the cells themselves since they determine the expression patterns for the different cell types and states. These patterns are comminicated to the DNA by transcrption factors, by methylation patterns and by binding to the tails of histones, all of which influence the pattern and speed of transcription of different parts of the genome. If the word ‘instruction’ is useful here at all, it is rather that the cell instructs the genome. As Barbara McClintock wrote in 1984 after receiving her Nobel Prize, the genome is an ‘organ of the cell’, not the other way around.
Realising that DNA is under the control of the system has been reinforced by the discovery that cells use different start, stop and splice sites for producing different messenger RNAs from a single DNA sequence. This enables the same sequence to code different proteins in different cell types and under different conditions [here’s where context-dependency comes into play again].
Representing the direction of causality in biology the wrong way round is therefore confusing and has far-reaching conseqeunces. The causality is circular, acting both ways: passive causality by DNA sequences acting as otherwise inert templates, and active causality by the functional networks of interactions that determine how the genome is activated.
This takes care of the idea that DNA is a ‘code’. But what about DNA being a ‘blueprint’, that all of the information is contained in the DNA of the organism before conception? DNA is clearly not a ‘program’, in the sense that all of the information to construct the organism exists already in DNA. The complete cell is also needed, and its “complex structures are inherited by self-templating” (Noble, 2017: 161). Thus, the “blueprint” is the whole cell, not just the genome itself (remember that the genome is an organ of the cell).
Lastly, GWA studies have been all the rage recently. However, there is only so much we can learn just from association studies, before we need to turn to the physiological sciences for functional analyses. Indeed, Denis Noble (2018) writes in a new editorial:
As with the results of GWAS (genome-wide association studies) generally, the associations at the genome sequence level are remarkably weak and, with the exception of certain rare genetic diseases, may even be meaningless (13, 21). The reason is that if you gather a sufficiently large data set, it is a mathematical necessity that you will find correlations, even if the data set was generated randomly so that the correlations must be spurious. The bigger the data set, the more spurious correlations will be found (3).
[…]
The results of GWAS do not reveal the secrets of life, nor have they delivered the many cures for complex diseases that society badly needs. The reason is that association studies do not reveal biological mechanisms. Physiology does. Worse still, “the more data, the more arbitrary, meaningless and useless (for future action) correlations will be found in them” is a necessary mathematical statement (3).
Nor does applying a highly restricted DNA sequence-based interpretation of evolutionary biology, and its latest manifestation in GWAS, to the social sciences augur well for society.
It is further worth noting that there is no privileged level of causation in biological systems (Noble, 2012)—a priori, there is no justification to privilege one system over another in regard to causation, so saying that one level of the organism is “higher” than another (for instance, saying that genes are, and should be, privileged over the environment or any other system in the organism regarding causation) is clearly false, since there is upwards and downwards causation, influencing all levels of the system.
In sum, it is highly misleading to refer to DNA as “blueprints”, a “code”, or a “recipe.” Referring to DNA in this way means that one presumes that DNA can be divorced from its context—that it does not work together with the environment. As I have argued in the past, association studies will not elucidate genetic mechanisms, nor will heritability estimates (Richardson, 2012). We need physiological testing for these functional analyses, and association studies like GWAS and even heritability estimates don’t tell us this type of information (Panofsky, 2014). So, it seems, that what Plomin et al are looking for that they assume are “in the genes”, are not there, because they use a false model of the gene (Burt, 2015; Richardson, 2017). Genes are resources—templates to be used by and for the system—not causes of traits and development. They can account for differences in variation, but cannot be said to be the origin of trait differences. Genes can be said to be difference makers, but knowing whether or not they are difference makers for behavior, in my opinion, cannot be known.
(For further information on genes and what they do, reach Chapters Four and Five of Ken Richardson’s book Genes, Brains, and Human Potential: The Science and Ideology of Intelligence. Plomin himself seems to be a reductionist, and Richardson took care of that paradigm in his book. Lickliter (2018) has a good review of the book, along with critiques of the reductionist paradigm that Plomin et al follow.)
Otzi Man’s Last Meal and the Diet of Neanderthals
1100 words
The debate on what type of diet in regard to macronutrient differences rages on. Should we eat high carb, low fat (HCLF)? Or low carb, high fat (LCHF) or something in between? The answer rests on, of course, the type of diets that our ancestors ate—both immediate and in the distant past. In the 1990s, a frozen human was discovered in the Otzal mountains, which gave him the name “Otzi man.” About 5,300 years ago, he was frozen in the mountains. The contents of his stomach have been analyzed in the 27 years since the discovery of Otzi, but an in-depth analysis was not possible until now.
A new paper was published recently, which analyzed the stomach contents of Otzi man (Maixner et al, 2018). There is one reason why it took so long to analyze the contents of his stomach: the authors state that, due to mummification, his stomach moved high up into his rib cage. The Iceman was “omnivorous, with a diet consisting both of wild animal and plant material” (Maixner et al, 2018: 2). They found that his stomach had a really high fat content, with “the presence of ibex and red deer” (pg 3). He also “consumed either fresh or dried wild meat“, while “a slow drying or smoking of the meat over the fire would explain the charcoal particles detected previously in the lower intestine content.“(pg 5).
The extreme alpine environment in which the Iceman lived and where he have been found (3,210 m above sea level) is particularly challenging for the human physiology and requires optimal nutrient supply to avoid rapid starvation and energy loss [31]. Therefore, the Iceman seemed to have been fully aware that fat displays an excellent energy source. On the other hand, the intake of animal adipose tissue fat has a strong correlation with increased risk of coronary artery disease [32]. A high saturated fats diet raises cholesterol levels in the blood, which in turn can lead to atherosclerosis. Importantly, computed tomography scans of the Iceman showed major calcifications in arteria and the aorta indicating an already advanced atherosclerotic disease state [33]. Both his high-fat diet and his genetic predisposition for cardiovascular disease [34] could have significantly contributed to the development of the arterial calcifications. Finally, we could show that the Iceman either consumed fresh or dried meat. Drying meat by smoking or in the open air are simple but highly effective methods for meat preservation that would have allowed the Iceman to store meat long term on journeys or in periods of food scarcity. In summary, the Iceman’s last meal was a well-balanced mix of carbohydrates, proteins, and lipids, perfectly adjusted to the energetic requirements of his high-altitude trekking. (Maixner et al, 2018: 5)
They claim that “the intake of animal adipose tissue fat has a strong correlation with increased risk of coronary artery disease“, of course, citing a paper that the AHA is involved in (Sacks et al, 2017) which says that “Randomized clinical trials showed that polyunsaturated fat from vegetable oils replacing saturated fats from dairy and meat lowers CVD.” This is nonsense, because dietary fat guidelines have no evidence (Harcombe et al, 2016; Harcombe, Baker, and Davies, 2016; Harcombe, 2017). Saturated fat consumption is not even associated with all-cause mortality, type II diabetes, ischemic stroke, CVD (cardiovascular disease) and CHD (coronary heart disease) (de Sousa et al, 2015).
Thus, if anything, what contributed to Otzi man’s arterial calcification seems to be grains/carbohydrates (see DiNicolantonio et al, 2017), not animal fat. Fats, at 9 kcal per gram, were better for Otzi to consume, as he got more kcal for his buck; eating a similar portion in carbohydrates, for example, would have meant that Otzi would have had to spend more time eating (since carbs have less than half the energy that animal fat does). Since his stomach had ibex (a type of goat) and red deer, it’s safe to say that many of his meals consisted mainly of animal fat, protein with some cereals and plants thrown in (he was an omnivore).
We can then contrast the findings of Otzi’s diet with that of Neanderthals. It has been estimated that, during glacial winters, Neanderthals would have consumed around 74-85 percent of their diet from animal fat when there were no carbohydrates around, with the rest coming from protein (Ben-Dor, Gopher, and Barkai, 2016). Furthermore, based on contemporary data from polar peoples, it is estimated that Neanderthals required around 3,360 to 4,480 kcal per day to winter foraging and cold resistance (Steegmann, Cerny, and Holliday, 2002). The upper-limit for protein intake for Homo sapiens is 4.0 g/bw/day while for erectus it is 3.9 g/bw/day (Ben-Dor et al, 2011), and so this shows that Neanderthals consumed a theoretical upper-maximum of protein due to their large body size. So we can assume that Neanderthals consumed somewhere near 3800 kcal per day. The average Neanderthal is said to have consumed about 292 grams of protein per day, or 1,170 kcal (with a lower end of 985 kcal and an upper end of 1,170 at the high end) (Ben-Dor, Gopher, and Barkai, 2016: 370).
Then if we further assume that Neanderthals consumed no carbohydrates during glacial winters, that leaves protein as the main source of energy, since the large game the Neanderthals hunted were not around. Thus, Neanderthals would have consumed between 2,812 and 3,230 kcal from animal fat with the rest coming from protein. We can also put this into perspective. The average American man consumes about 100 grams of protein per day, while consuming 2,195 kcal per day (Ford and Dietz, 2013). For these reasons, and more, I argued that Neanderthals were significantly stronger than Homo sapiens, and this does have implications for racial differences in athletic ability.
In sum, the last meal of Otzi man is now known. Of course, this is a case of n = 1, so we should not draw too large a conclusion from this, but it is interesting. I don’t see why the composition of the diets of any of Otzi’s relatives would have been any different (or that the contents of his normal diet would have been any different). He ate a diet high in animal fat like Neanderthals, but unlike Neanderthals, they ate a more cereal-based diet which may have contributed to Otzi’s CVD and arterial calcification. We can learn a lot about ourselves and our ancestors through the analysis of their stomach contents (if possible) and teeth (if possible), and maybe even genomes (Berens, Cooper, and Lachance, 2017) because if we learn what they ate then we can maybe begin to shift dietary advice to a more ‘natural’ way and avoid diseases of civilization. But, we have not had time to adapt to the new obesogenic environments we have constructed for ourselves. It’s due to this that we have an obesity epidemic, and by studying the diets of our ancestors, we can then begin to remedy our obesity and other health problems.
Mini-Review of “J. Phillipe Rushton: A Life History Perspective” by Edward Dutton
1500 words
JP Rushton was a highly controversial psychologist professor, teaching at the University of Western Ontario for his entire career. In the mid-1980s, he proposed that evolution was “progressive” and that there was a sort of “hierarchy” between the three races that he termed “Mongoloid, Caucasoid, and Negroid” (Rushton, 1985). His theory was then strongly criticized scientists from numerous disciplines (Lynn, 1989; Cain, 1990; Weizmann et al, 1990; Anderson, 1991; Graves, 2002). Rushton responded to these criticisms (Rushton, 1989; Rushton, 1991; Rushton, 1997; though it’s worth noting that Rushton never responded to Graves’ 2002 critiques). (Also see Rushton’s and Graves’ debate.) Copping, Campbell, and Muncer (2014) write that “high K scores were related to earlier sexual debut and unrelated to either pubertal onset or number of sexual partners. This suggests that the HKSS does not reflect an underlying “K dimension.”“, which directly contradicts Rushton’s racial r/K proposal.
There is a now a new critique of Rushton’s theory out now, by Edward Dutton, English anthropologist, with a doctorate in religious studies, just published at the end of last month (Dutton, 2018). I ordered the book the day after publication and it took three weeks to get to my residence since it came from the UK. I finally received it on Friday. It’s a small book, 143 pages sans acknowledgments, references and the index, and seems well-written and researched from what I’ve read so far.
Here is the plan of the book:
Accordingly, in this chapter [Chapter One], we will begin by getting to grips with the key concepts of intelligence and personality. This part is primarily aimed at non-specialist readers or those who are sceptical of the two concepts [it’s really barebones; I’m more than ‘sceptical’ and it did absolutely nothing for me]. In Chapter Two, we will explore Rushton’s theory in depth. Readers who are familiar with Life History Theory may wish to fast forward through to the section on the criticisms of Rushton’s model. I intend to be as fair to his theory as possible, in a way so few of the reviewers were when he presented it. I will respond to the many fallacious criticisms of it, all of which indicate non-scientific motives [what about Rushton? Did he have any non-scientific motives?]. However, I will show that Rushton is just as guilty of these kinds of techniques as his opponents. I will also highlight serious problems with his work, including cherry picking, confirmation bias, and simply misleading other researchers. In Chapter Three, we will explore the concept of ‘race’ and show that although Rushton’s critics were wrong to question the concept’s scientific validity, Rushton effectively misuses the concept, cherry-picking such that his concept works. In Chapter Four, we will explore the research that has verified Rushton’s model, including new measures which he didn’t examine. We will then, in Chapter Five, examine the concept of genius and look at how scientific geniuses tend to be highly intelligent r-strategists, though we will see that Rushton differed from accepted scientific geniuses in key ways.
In Chapter Six, we will find that Rushton’s theory itself is problematic, though not in the ways raised by his more prominent critics. It doesn’t work when it comes to a key measure of mental stability as well as to many other measures, specifically preference for oral sex, the desire to adopt non-related children, the desire to have pets, and positive attitudes to the genetically distant. It also doesn’t work if you try to extend it to other races, beyond the three large groups he examined [because more races exist than Rushton allows]. In Chapter Seven, with all the background, we will scrutinize Rushton’s life up until about the age of 30, while in Chapter Eight, we will follow Rushton from the age of 30 until his death. I will demonstrate the extent to which he was a highly intelligent r-strategist and a Narcissist and we will see that Rushton seemingly came from a line of highly intelligent r-strategists. In Chapter Nine, I will argue that for the good of civilization those who strongly disagree with Rushton must learn to tolerate people like Rushton. (Dutton, 2018: 12-13).
On the back of the book, he writes that Rushton had “two illegitimate children including one by a married black woman.” This is intriguing. Could this be part of Rushton’s motivation to formulate his theory (his theory has already been rebutted by numerous people, so speculating on motivations in lieu of new information seems apt)?
Some people, such as PumpkinPerson, may wonder why Dutton is attacking someone “on his team“, but he addresses people who would ask such questions, writing (pg. 15):
“But on this basis, it could be argued that my critique of Rushton simply gives ammunition to emotionally-driven scientists and their friends in the media. However, it could be countered that my critique only goes to show that it is those who are genuinely motivated by the understanding of the world — those who accept empirical evidence, such as with regard to intelligence and race — who are prepared to critique those regarded as being ‘on their side.’ And this is precisely because they are unbiased and thus do not think in terms of ‘teams.’”
Dutton argues that “many of the criticisms leveled against Rushton’s work by mainstream scientists were actually correct” (pg 13). This is a truism. One only need to read the replies to Rushton, especially Anderson (1991) to see that he completely mixed up the theory. He stated ‘Negroids’ were r-strategists and ‘Mongoloids’ were K-strategists, but this reasoning shows that he did not understand the theory—or, if anything, he knowingly attempted to obfuscate the theory in order to lend stronger credence to his own theory (and personal biases).
The fatal flaw for Rushton’s theory is that, if r/K selection theory did apply to human races, that ‘Mongoloids’ would be r-strategists while ‘Negroids’ would be K-strategists. This is because “Rushton’s own suggested agents of natural selection on African populations imply that African populations have had a strong history of K-selection, as well as the r-selection implied by “droughts”” (Anderson, 1991: 59). As for Mongoloids, “Rushton lists many traits of Mongoloid peoples that are thought to represent adaptation to cold. Cold weather acts in a density-independent fashion (adaptations to cold improve survival in cold weather regardless of population density); cold weather is normally an agent of r-selection” (Anderson, 1991: 59). Rushton’s own arguments imply that ‘Negroids’ would have had more time to approach their environmental carrying capacity and experience ‘K-selecting’ pressures.
Thus, Rushton’s claim about the empirical ordering of life history and behavioural traits in the racial groups exactly contradicts general predictions that follow from his own claims about their ancestral ecology and the r/K model (Boyce, 1984; MacArthur, 1972; MacArthur & Wilson, 1967; Pianka, 1970; Ricklefs, 1990, p. 577). (Specific predictions from the model could be made only about individual populations after careful study in their historical habitat, as I have pointed out above). (Anderson, 1991: 59) [And it is not possible, because the populations in question should be living in the environment that the selection is hypothesized to have occurred. That, of course, is not possible today.]
Though, near the end of the book, Dutton writes that (pg 148) that “Rushton was not a scientific genius. As we have discussed, unlike a scientific genius, his models had clear deficiencies, he cherry-picked data to fit his model, and he was biased in favor of his model. However, Rushton was a highly original scientist who developed an extremely original and daring theory: a kind of artistic-scientist genius combination.”
The final paragraph of the book, though, sums up the whole book up well. Dutton talks about when Jared Taylor introduces Rushton at one of his American Renaissance conferences (February 25th, 2006):
‘Well, thank you very much and . . . eh . . . and thank you Jared for . . . erm . . . putting on another wonderful conference.’ Rushton was reserved, yet friendly and avuncular. ‘Eh . . . it’s a great honor to be the after dinner speaker; to be elevated up like this.’ He was certainly elevated up. Taylor had even remarked that ‘in a sane and civilized world’ Rushton’s work would have ‘worldwide acclaim.’ Rushton’s audience admired him, trusted him . . . They weren’t familiar with him at all.
All in all, to conclude this little mini-review, I would recommend picking up this book as it’s a great look into Rushton’s life, the pitfalls of his theory (and for the new work and other variables that Dutton shows showed Rushton’s M>C>N ‘hierarchy’). Rushton’s work, while politically daring, did not hold up to scientific scrutiny, since the model was beginning to be abandoned in the late 70s (Graves, 2002), with most scientists completely dismissing the model in the early 90s. Commenting on r/K selection, Stearns (1992: 206) writes that “This explanation was suggestive and influential but incorrect” (quoted in Reznick et al, 2002), while Reznick et al (2002: 1518) write that “The r- and K-selection paradigm was replaced by new paradigm that focused on age-specific mortality (Stearns 1976, Charlesworth 1980).” Rushton’s model, while it ‘made sense with the data’, was highly flawed. And even then, it doesn’t matter that it ‘made sense’ with the data, since Rushton’s theory is one large just-so story (Gould and Lewontin, 1976; Lloyd, 1999; Richardson, 2007; Nielsen, 2009; see also Pigliucci and Kaplan, 2000 and Kaplan, 2002
Black-White Differences in Anatomy and Physiology: Black Athletic Superiority
3000 words
Due to evolving in different climates, the different races of Man have differing anatomy and physiology. This, then, leads to differences in sports performance—certain races do better than others in certain bouts of athletic prowess, and this is due to, in large part, heritable biological/physical differences between blacks and whites. Some of these differences are differences in somatotype, which bring a considerable advantage for, say, runners (an ecto-meso, for instance, would do very well in sprinting or distance running depending on fiber typing). This article will discuss differences in racial anatomy and physiology (again) and how it leads to disparities in certain sports performance.
Kerr (2010) argues that racial superiority in sport is a myth. (Read my rebuttal here.) In his article, Kerr (2010) attempts to rebut Entine’s (2000) book Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It. In a nutshell, Kerr (2010) argues that race is not a valid category; that other, nongenetic factors play a role other than genetics (I don’t know if anyone has ever argued if it was just genetics). Race is a legitimate biological category, contrary to Kerr’s assertions. Kerr, in my view, strawman’s Entine (2002) by saying he’s a “genetic determinist”, but while he does discuss biological/genetic factors more than environmental ones, Entine is in no way a genetic determinist (at least that’s what I get from my reading of his book, other opinions may differ). Average physical differences between races are enough to delineate racial categories and then it’s only logical to infer that these average physical/physiological differences between the races (that will be reviewed below) would infer an advantage in certain sports over others, while the ultimate cause was the environment that said race’s ancestors evolved in (causing differences in somatotype and physiology).
Black athletic superiority has been discussed for decades. The reasons are numerous and of course, this has even been noticed by the general public. In 1991, half of the respondents of a poll on black vs. whites in sports “agreed with the idea that “blacks have more natural physical ability,“” (Hoberman, 1997: 207). Hoberman (1997) of course denies that there is any evidence that blacks have an advantage over whites in certain sports that come down to heritable biological factors (which he spends the whole book arguing). However, many blacks and whites do, in fact, believe in black athletic superiority and that physiologic and anatomic differences between the races do indeed cause racial differences in sporting performance (Wiggins, 1989). Though Wiggins (1989: 184) writes:
The anthropometric differences found between racial groups are usually nothing more than central tendencies and, in addition, do not take into account wide variations within these groups or the overlap among members of different races. This fact not only negates any reliable physiological comparisons of athletes along racial lines, but makes the whole notion of racially distinctive physiological abilities a moot point.
This is horribly wrong, as will be seen throughout this article.
The different races have, on average, differing somatotypes which means that they have different anatomic proportions (Malina, 1969):
| Data from Malina, (1969: 438) | n | Mesomorph | Ectomorph | Endomorph |
| Blacks | 65 | 5.14 | 2.99 | 2.92 |
| Whites | 199 | 4.29 | 2.89 | 3.86 |
| Data from Malina (1969: 438) | Blacks | Whites |
| Thin-build body type | 8.93 | 5.90 |
| Submedium fatty development | 48.31 | 29.39 |
| Medium fleshiness | 33.69 | 43.63 |
| Fat and very fat categories | 9.09 | 21.06 |
This was in blacks and whites aged 6 to 11. Even at these young ages, it is clear that there are considerable anatomic differences between blacks and whites which then lead to differences in sports performance, contra Wiggins (1989). A basic understanding of anatomy and how the human body works is needed in order to understand how and why blacks dominate certain sports over whites (and vice versa). Somatotype is, of course, predicated on lean mass, fat mass, bone density, stature, etc, which are heritable biological traits, thus, contrary to popular belief that somatotyping holds no explanatory power in sports today (see Hilliard, 2012).
One variable that makes up somatotype is fat-free body mass. There are, of course, racial differences in fat mass, too (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000). Lower fat mass would, of course, impede black excellence in swimming, and this is what we see (Rushton, 1997; Entine, 2000). Wagner and Heyward (2000) write:
Our review unequivocally shows that the FFB of blacks and whites differs significantly. It has been shown from cadaver and in vivo analyses that blacks have a greater BMC and BMD than do whites. These racial differences could substantially affect measures of body density and %BF. According to Lohman (63), a 2% change in the BMC of the body at a given body density could, theoretically, result in an 8% error in the estimation of %BF. Thus, the BMC and BMD of blacks must be considered when %BF is estimated.
While Vickery, Cureton, and Collins (1988) found that blacks had thinner skin folds than whites, however, in this sample, somatotype did not explain racial differences in bone density, like other studies (Malina, 1969), Vickery, Cureton, and Collins (1988) found that blacks were also more likely to be mesomorphic (which would then express itself in racial differences in sports).
Hallinan (1994) surveyed 32 sports science, exercise physiology, biomechanics, motor development, motor learning, and measurement evaluation textbooks to see what they said racial differences in sporting performance and how they explained them. Out of these 32 textbooks, according to Wikipedia, these “textbooks found that seven [textbooks] suggested that there are biophysical differences due to race that might explain differences in sports performance, one [textbook] expressed caution with the idea, and the other 24 [textbooks] did not mention the issue.” Furthermore, Strklaj and Solyali (2010), in their paper “Human Biological Variation in Anatomy Textbooks: The Role of Ancestry” write that their “results suggest that this type of human variation is either not accounted for or approached only superficially and in an outdated manner.”
It’s patently ridiculous that most textbooks on the anatomy and physiology of the human body do not talk about the anatomic and physiologic differences between racial and ethnic groups. Hoberman (1997) also argues the same, that there is no evidence to confirm the existence of black athletic superiority. Of course, many hypotheses have been proposed to explain how and why blacks are at an inherent advantage in sport. Hoberman (1997: 269) discusses one, writing (quoting world record Olympian in the 400-meter dash, Lee Evans):
“We were bred for it [athletic dominance] … Certainly the black people who survived in the slave ships must have contained the highest proportion of the strongest. Then, on the plantations, a strong black man was mated with a strong black woman. We were simply bred for physical qualities.”
While Hoberman (1997: 270-1) also notes:
Finally, by arguing for a cultural rather than a biological interpretation of “race,” Edwards proposed that black athletic superiority results from “a complex of societal conditions” that channels a disproporitionate number of talented blacks into athletic careers.
The fact that blacks were “bred for” athletic dominance is something that gets brought up often but has little (if any) empirical support (aside from just-so stories about white slavemasters breeding their best, biggest and strongest black slaves). The notion that “a complex of societal conditions” (Edwards, 1971: 39) explains black dominance in sports, while it has some explanatory power in regard to how well blacks do in sporting competition, it, of course, does not tell the whole story. Edwards (1978: 39) argues that these complex societal conditions “instill a heightened motivation among black male youths to achieve success in sports; thus, they channel a proportionately greater number of talented black people than whites into sports participation.” While this may, in fact, be true, this does nothing to rebut the point that differences in anatomic and physiologic factors are a driving force in racial differences in sporting performance. However, while these types of environmental/sociological arguments do show us why blacks are over-represented in some sports (because of course motivation to do well in the sport of choice does matter), they do not even discuss differences in anatomy or physiology which would also be affecting the relationship.
For example, one can have all of the athletic gifts in the world, one can be endowed with the best body type and physiology to do well in any type of sport you can imagine. However, if he does not have a strong mind, he will not succeed in the sport. Lippi, Favaloro, and Guidi (2008) write:
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.
Any athlete—no matter their race—needs a strong mental background, for if they don’t, they can have all of the physical gifts in the world, they will not become top-tier athletes in the sport of their choice; advantageous physical factors are imperative for success in differing sports, though myriad variables work in concert to produce the desired effect so you cannot have one without the other. On the other side, one can have a strong mental background and not have the requisite anatomy or physiology needed to succeed in the sport in question, but if he has a stronger mind than the individual with the requisite morphology, then he probably will win in a head-to-head competition. Either way, a strong mind is needed for strong performance in anything we do in life, and sport is no different.
Echoing what Hoberman (1997) writes, that “racist” thoughts of black superiority in part cause their success in sport, Sheldon, Jayaratne, and Petty (2007) predicted that white Americans’ beliefs in black athletic superiority would coincide with prejudice and negative stereotyping of black’s “intelligence” and work ethic. They studied 600 white men and women to ascertain their beliefs on black athletic superiority and the causes for it. Sheldon, Jayaratne, and Petty (2007: 45) discuss how it was believed by many, that there is a “ perceived inverse relationship between athleticism and intelligence (and hard work).” (JP Rushton was a big proponent of this hypothesis; see Rushton, 1997. It should also be noted that both Rushton, 1997 and Entine, 2000 believe that blacks’ higher rate of testosterone—3 to 15 percent— [Ross et al, 1986; Ellis and Nyborg, 1992; see rebuttal of both papers] causes their superior athletic performance, I have convincingly shown that they do not have higher levels of testosterone than other races, and if they do the difference is negligible.) However, in his book The Sports Gene: Inside the Science of Extraordinary Athletic Performance, Epstein (2014) writes:
With that stigma in mind [that there is an inverse relationship between “intelligence” and athletic performance], perhaps the most important writing Cooper did in Black Superman was his methodological evisceration of any supposed inverse link between physical and mental prowess. “The concept that physical superiority could somehow be a symptom of intellectual superiority became associated with African Americans … That association did not begin until about 1936.”
What Cooper (2004) implied is that there was no “inverse relationship” with intelligence and athletic ability until Jesse Owens blew away the competition at the 1936 Olympics in Berlin, Germany. In fact, the relationship between “intelligence” and athletic ability is positive (Heppe et al, 2016). Cooper is also a co-author of a paper Some Bio-Medical Mechanisms in Athletic Prowess with Morrison (Morrison and Cooper, 2006) where they argue—convincingly—that the “mutation appears to have triggered a series of physiological adjustments, which have had favourable athletic consequences.”
Thus, the hypothesis claims that differences in glucose conversion rates between West African blacks and her descendants began, but did not end with the sickling of the hemoglobin molecule, where valine is substituted for glutamic acid, which is the sixth amino acid of the beta chain of the hemoglobin molecule. Marlin et al (2007: 624) showed that male athletes who were inflicted with the sickle cell trait (SCT) “are able to perform sprints and brief exercises at the highest levels.” This is more evidence for Morrison and Cooper’s (2006) hypothesis on the evolution of muscle fiber typing in West African blacks.
Bejan, Jones, and Charles (2010) explain that the phenomenon of whites being faster swimmers in comparison to blacks being faster runners can be accounted for by physics. Since locomotion is a “falling-forward cycle“, body mass falls forward and then rises again, so mass that falls from a higher altitude falls faster and forward. The altitude is set by the position of center of mass above the ground for running, while for swimming it is set by the body rising out of the water. Blacks have a center of gravity that is about 3 percent higher than whites, which implies that blacks have a 1.5 percent speed advantage in running whereas whites have a 1.5 percent speed advantage in swimming. In the case of Asians, when all races were matched for height, Asians fared even better, than whites in swimming, but they do not set world records because they are not as tall as whites (Bejan, Jones, and Charles, 2010).
It has been proposed that stereotype threat is part of the reasons for East African running success (Baker and Horton, 2003). They state that many theories have been proposed to explain black African running success—from genetic theories to environmental determinism (the notion that physiologic adaptations to climate, too, drive differences in sporting competition). Baker and Horton (2003) note that “that young athletes have internalised these stereotypes and are choosing sport participation accordingly. He speculates that this is the reason why white running times in certain events have actually decreased over the past few years; whites are opting out of some sports based on perceived genetic inferiority.” While this may be true, this wouldn’t matter, as people gravitate toward what they are naturally good at—and what dictates that is their mind, anatomy, and physiology. They pretty much argue that stereotype threat is a cause of East African running performance on the basis of two assertions: (1) that East African runners are so good that it’s pointless to attempt to win if you are not East African and (2) since East Africans are so good, fewer people will try out and will continue the illusion that East Africans would dominate in middle- and long-distance running. However, while this view is plausible, there is little data to back the arguments.
To explain African running success, we must do it through a systems view—not one of reductionism (i.e., gene-finding). We need to see how the systems in question interact with every part. So while Jamaicans, Kenyans, and Ethiopians (and American blacks) do dominate in running competitions, attempting to “find genes” that account for success n these sports seems like a moot point—since the whole system is what matters, not what we can reduce the system in question to.
However, there are some competitions that blacks do not do so well in, and it is hardly discussed—if at all—by any author that I have read on this matter. Blacks are highly under-represented in strength sports and strongman competitions. Why? My explanation is simple: the causes for their superiority in sprinting and distance running (along with what makes them successful at baseball, football, and basketball) impedes them from doing well in strength and strongman competitions. It’s worth noting that no black man has ever won the World’s Strongest Man competition (indeed the only African country to even place—Rhodesia—was won by a white man) and the causes for these disparities come down to racial differences in anatomy and physiology.
I discussed racial differences in the big four lifts and how racial differences in anatomy and physiology would contribute to how well said race performed on the lift in question. I concluded that Europeans and Asians had more of an advantage over blacks in these lifts, and the reasons were due to inherent differences in anatomy and physiology. One major cause is also the differing muscle fiber typing distribution between the races (Alma et al, 1986; Tanner et al, 2002; Caesar and Henry, 2015 while blacks’ fiber typing helps them in short-distance sprinting (Zierath and Hawley, 2003). Muscle fiber typing is a huge cause of black athletic dominance (and non-dominance). Blacks are not stronger than whites, contrary to popular belief.
I also argued that Neanderthals were stronger than Homo sapiens, which then had implications for racial differences in strength (and sports). Neanderthals had a wider pelvis than our species since they evolved in colder climes (at the time) (Gruss and Schmidt, 2016). With a wider pelvis and shorter body than Homo sapiens, they were able to generate more power. I then implied that the current differences in strength and running we see between blacks and whites can be used for Neanderthals and Homo sapiens, thusly, evolution in differing climates lead to differences in somatotype, which eventually then lead to differences in sporting competition (what Baker and Horton, 2003 term “environmental determinism” which I will discuss in the context of racial differences in sports in the future).
Finally, blacks dominate the sport of bodybuilding, with Phil Heath dominating the competition for the past 7 years. Blacks dominate bodybuilding because, as noted above, blacks have thinner skin folds than whites, so their striations in their muscles would be more prevalent, on average, at the same exact %BF. Bodybuilders and weightlifters were similar in mesomorphy, but the bodybuilders showed more musculature than the bodybuilders whereas the weightlifters showed higher levels of body fat with a significant difference observed between bodybuilders and weightlifters in regard to endomorphy and ectomorphy (weightlifters skewing endo, bodybuilders skewing ecto, as I have argued in the past; Imran et al, 2011).
To conclude, blacks do dominate American sporting competition, and while much ink has been spilled arguing that cultural and social—not genetic or biologic—factors can explain black athletic superiority, they clearly work in concert with a strong mind to produce the athletic phenotype, no one factor has prominence over the other; though, above all, if one does not have the right mindset for the sport in question, they will not succeed. A complex array of factors is the cause of black athletic dominance, including muscle fibers, the type of mindset, anatomy, overall physiology and fat mass (among other variables) explain the hows and whys of black athletic superiority. Cultural and social explanations—on their own—do not tell the whole story, just as genetic/biologic explanations on their own would not either. Every aspect—including the historical—needs to be looked at when discussing the dominance (or lack thereof) in certain sports along with genetic and nongenetic factors to see how and why certain races and ethnies excel in certain sports.
Vitamin D, Physiology, and the Cold
1200 words
I’ve been chronicling the VDH recently since it has great explanatory—and predictive—power. Light skin is a clear adaptation to low UVR, while dark skin is a clear adaptation to high UVR. Dark, highly melanized skin confers advantages in high UVR environments, such as protection against DNA damage, and also absorbs sufficient UV for vitamin D production while also protecting against folate depletion. However, when our ancestors migrated out of Africa, dark skin would not cut it in temperate environments with highly variable UV rays. This is where our highly adaptive physiology came into play, ensuring that we survived in highly variable environments. Light skin was important in low UVR environments in order to synthesize ample vitamin D, however, that synthesized vitamin D then conferred numerous other physiological advantages to the cold.
Eighty to ninety percent of the vitamin D required for humans comes from the sun, whereas ten to twenty percent comes from the diet, such as fatty fish, eggs, and dairy products (fortified with vitamin D, of course) (Ajabshir, Asif, and Nayer, 2014). Humans need to rely on high amounts of UV rays for vitamin D synthesis (Carlberg, 2014) other than Arctic peoples. Since dark skin does not synthesize vitamin D as well as light skin, skin gradually lightened as our ancestors migrated out of Africa (Juzeniene et al, 2009). This was then imperative to the physiologic adaptations that then occurred as our physiology had to adapt to novel, colder environments with fewer UV rays.
Sufficient amounts of vitamin D are highly important for the human musculoskeletal system (Wintermeyer et al, 2016), which is extremely important for birthing mothers. Along with the increased vitamin D synthesis in low UV environments, the heightened production of vitamin D conferred numerous other physiologic benefits which then helped humans adapt to colder environments with more varying UVR.
Vasoconstriction occurs when the blood vessels constrict which leads to heightened blood pressure, whereas vasodilation is the dilation of blood vessels which decreases blood pressure. So evolutionarily speaking, we had to have adaptive physiology in order to be able to “switch” back and forth between vasoconstriction and vasodilation, depending on what the current environment needed. Vasodilation, though, most likely had no advantage in high UV environments, and thus must have been an advantage in low UV environments, where it was more likely to be colder and so, when the blood vessels constrict, blood pressure increases and thus, heat loss could be considerably slowed in these environments due to these physiologic adaptations.
The races also differ, along with many other physiologic abilities, in nitric oxide-mediated vasodilation. Vasodilation is the dilation of blood vessels, which increases blood pressure. Mata-Greenwood and Chen (2008) reviewed the relevant literature regarding black/white differences in nitric oxide-dependent vasorelaxation and concluded that nitric oxide vasodilation is reduced in darker-skinned populations. Thus, we can infer that in lighter-skinned populations nitric oxide vasodilation is increased in lighter-skinned populations, which would have conferred a great physiological advantage when it came to colonizing environments with lower UV rays.
VDR and vitamin D metabolizing enzymes are present in adipose tissue. Tetrahydrobiopterin; which acts as a cofactor in the synthesis of nitric oxide and its primary function is as a vasodilator in the blood vessels (meaning that blood pressure is increased, to keep more heat in the cold) (Chalupsky and Cai, 2005). Since vasodilation is the body’s primary response to heat stress, blood flow increases which allows heat to leave the body. Therefore, the human body’s ability regarding vasodilation and vasoconstriction mechanisms were important in surviving areas with varying UVR.
One function of our adipose tissue is the storage of vitamin D, while vitamin D metabolizing enzymes and VDR are also expressed in the adipocyte (Abbas, 2017). With these known actions of vitamin D on adipose tissue, we can speculate that since vitamin D and the VDR are expressed in adipose tissue, it may have exerted a role in the adipose tissue which may have been important for surviving in cold, low UV environments (see below).
Furthermore, since these mechanisms are brought on by short-term changes, we can infer that it would hardly be of any use in high UVR environments and would be critical in temperate environments. So, vasodilation and vasoconstriction have little to no benefit in high UVR environments but seem to be imperative in temperate environments where UVR varies. It’s also likely that vitamin D influences vasodilation by influential nitric oxide synthesis (see Andrukhova et al, 2014) and vasoconstriction by influencing the renin-angiotensin system (Ajabshir, Asig, and Nayer, 2014).
This would have conferred great benefit to our ancestors as they migrated into more temperate and colder climates. You can read this for information on how adaptive our physiology is and why it’s like that. Because we went into numerous new environments and natural selection couldn’t act quickly enough, therefore the human body’s physiology is extremely adaptive.
What this suggests is that as skin lightened and adapted to low UV, the increased synthesis in vitamin D influenced vasodilation by a strong influence on nitric oxide synthase, along with vasoconstriction, implies that it would have been easier to survive in novel environments due to adaptive physiology and skin color, along with body fat reserves and the physiologic effects of vitamin D on adipose tissue. These physiologic adaptations would have been of no to little use in Africa. Thus, they must have been useful after we migrated out of Africa and experienced wildly varying environments—the whole reason why our physiology evolved (Richardson, 2017: chapter 5).
When the human body is exposed to cold, a few things occur: cutaneous vasoconstriction, shivering (Castellani and Young, 2016), “behavioral thermoregulation” (Young, Sawka, and Pandolf, 1996), while the human body can adapt physiologically to the cold (Young, 1994). The physiologic functions that vitamin D and folate in regard to vasodilation and vasoconstriction, there is a great chance that these effects were important in maintaining energy homeostasis in colder climates.
In sum, the evolution of light skin conferred a great survival advantage to our ancestors. This then upped the production of vitamin D synthesis in the body, which where then of utmost importance in regard to the adaptation of the human physiology to colder, lower-UV environments. Without our adaptive physiological systems, we would not have been able to leave Africa into novel environments. We need both behavioral thermoregulation as well as adaptive physiology to be able to survive in novel environments. Thus, the importance of skin lightening in our evolution becomes clearer:
As humans migrated out of Africa, lighter skin was needed to synthesize vitamin D. This was especially important to women, who needed higher amounts of vitamin D, in order to produce enough calcium for lactation and pregnancy—so the babe had enough calcium to grow its skeleton in the womb. With the uptake in vitamin D synthesis, this then allowed more adaptive physiologic changes that occurred due to the cold, and along with vasodilation and vasoconstriction, along with shivering and adapting behaviorally to the new environments, were our ancestors able to survive. Dark skin cannot synthesize vitamin D as well as light skin in low UV environments; this also can be seen with the lowered production of nitric oxide-dependent vasodilation in dark-skinned populations. Thus, vasoconstriction conferred no physiologic benefit in high UV environments, but almost certainly conferred a physiologic benefit in low UV environments.
Why Are Women Lighter than Men? Skin Color and Sexual Selection
1550 words
Skin color differences between the sexes are always discussed in terms of women being lighter than men, but never men being darker than women. This is seen in numerous animal studies (some reviewed by Rushton and Templer, 2012; read rebuttal here; also see Ducrest, Keller, and Roulin, 2008). Though, the colors that evolved on the animal’s fur due to whatever mate choices are irrelevant to the survival capabilities that the fur, feathers etc give to the organism in question. So, when we look at humans, we lost our protective body hair millions of years ago (Lieberman, 2015), and with that, we could then sweat. So since furlessness evolved in the lineage Homo, there was little flexibility in what could occur due to environmental pressures on skin color in Africa. It should be further noted that, as Nina Jablonski writes in her book Living Color: The Biological and Social Meaning of Skin Color (2012, pg 74)
No researchers, by the way, have explored the opposite possibility, that women deliberately selected darker men!
One hypothesis proposes that lighter skin in women first arose as a byproduct due to the actions of differing levels of hormones in the sexes—with men obviously having higher levels of testosterone, making them darker them women. So according to this hypothesis, light-skinned women evolved since men could tell high-quality from low-quality mates as well as measure hormonal status and childbearing potential, which was much easier to do with lighter- than darker-skinned women.
Another hypothesis put forth is that further from the equator, sexual competition between women would have increased for mates since mates were depleted, and so light skin evolved since men found it more beautiful. Thus, women living at higher latitudes were lighter than women living at lower latitudes because men had to go further to hunt which meant they were more likely to die which caused even greater competition between females, lightening their skin even more. And another, related, argument, proposed that light skin in women evolved due to a complex of childlike traits which includes a higher voice, smoother skin and childlike facial features, which then reduced male competition and aggressiveness. But women did not stay around waiting to be provisioned and they got out and gathered, and hunted sometimes, too.
Harris (2005) proposes that light skin evolved due to parental selection—mothers choosing the lightest daughters to survive, killing off the darker ones. All babies are born pale—or at least lacking the amount of pigment they have later in life. So how would parental—mostly maternal—selection have caused selection for lighter skin in girls as Harris (2005) proposes? It’d be a pretty large guessing game.
The role of sexual selection in regard to human skin color, though, has been tested and falsified. Madrigal and Kelly (2007a) tested the hypothesis that skin reflectance should be positively correlated with distance from the equator. It was proposed by other authors that as our ancestors migrated out of Africa, environmental selection relaxed and sexual selection took over. Their data did not lend credence to the hypothesis and falsified it.
Madrigal and Kelly (2007a: 475) write (emphasis mine):
We tested the hypothesis that human sexual dimorphism in skin color should be positively correlated with distance from the equator, a proposal generated by the sexual selection hypothesis. We found no support for that proposition. Before this paper was written, the sexual selection hypothesis was based on stated male preference data in a number of human groups. Here, we focused on the actual pattern of sexual dimorphism. We report that the distribution of human sexual dimorphism in relation to latitude is not that which is predicted by the sexual selection hypothesis. According to that hypothesis, in areas of low solar radiation, there should be greater sexual dimorphism, because sexual selection for lighter females is not counterbalanced by natural selection for dark skin. Our data analysis does not support this prediction.
Though Frost (2007) replied, stating that Madrigal and Kelly (2007a) presumed that sexual selection was equal in all areas. Madrigal and Kelly (2007b) responded, stating that they tested one specific hypothesis regarding sexual selection and found it to be false. Frost (2007) proposed two hypotheses in order to test his version, but, again, no one has proposed that women select darker men, which could be a cause of lighter-skinned women (though sexual selection does not—and cannot—explain the observed gradation in skin color between men and women).
Skin color differences between men and women first arose to ensure women enough calcium for lactation and pregnancies. Since skin pigmentation protects against UVR but also must generate vitamin D, it must be light or dark enough to ensure ample vitamin D production in that certain climate, along with protecting against the UVR in that climate. So women needed sufficient vitamin D, which meant they needed sufficient calcium to ensure a strong skeleton for the fetus, for breastfeeding and for the mother’s own overall health.
However, breastfeeding new babes is demanding on the mother’s body (calcium reserves are depleted four times quicker), and the calcium the babe needs to grow its skeleton comes directly from the mother’s bones. Even a mother deficient in vitamin D will still give calcium to the babe at the expense of her own health. But she then needs to increase her reserves of calcium in order to ensure future pregnancies aren’t fatal for her or her offspring.
Though, at the moment to the best of my knowledge, there are no studies on calcium absorption, vitamin D levels and the recovery of the female skeleton after breastfeeding. (Though n3 fatty acids are paramount as well, and so a mother must have sufficient fat stores; see Lassek and Gaulin, 2008.) Thus, light-skinned women are most likely at an advantage when it comes to vitamin D production: The lighter they are, the more vitamin D and calcium they can produce for more pregnancies. Since light skin synthesizes vitamin D more efficiently, the body could then synthesize and use calcium more efficiently. The body cannot use and absorb calcium unless vitamin D is present. Since the fetus takes calcium from the mother’s skeleton, ample amounts of vitamin D must be present. For ample amounts of vitamin D to be present, the skin must be light enough to ensure vitamin D synthesis which would be needed for calcium absorption (Cashman, 2007; Gallagher, Yalamanchili, and Smith, 2012; Aloia et al, 2013).
Nina Jablonski writes in her book (2012, 77):
Women who are chronically deficient in vitamin D because of successive pregnancies and periods of breastfeeding experience a form of bone degeneration called osteomalacia. This has serious consequences for infants born of later pregnancies and for mothers themselves, who are at greater risk of breaking bones. It makes sense that protection of female health during the reproductive years would be a top evolutionary priority, so we are now investigating whether, in fact, slightly lighter skin in women might be a fairly simple way of ensuring that women get enough vitamin D after pregnancy and breastfeeding to enable their bodies to recover quickly. The need for maintaining strong female skeletons through multiple pregnancies may have been the ultimate evolutionary reason for the origin of differences in skin color between men and women.
While Jablonski and Chaplin (2000: 78) write:
We suggest that lighter pigmentation in human females began as a trait directly tied to increased fitness and was subsequently reinforced and enhanced in many human populations by sexual selection.
It is obvious that skin color in women represents a complex balancing act between giving the body the ability to synthesize ample vitamin D and protect from UVR. Skin coloration in humans is very clearly highly adaptive to UVR, and so, with differing average levels of UVR in certain geographic locales, skin color would have evolved to accommodate the human body to whichever climate it found itself in—because human physiology is perhaps the ultimate adaptation.
Sexual selection for skin color played a secondary, not primary role (Jablonski, 2004: 609) in the evolution of skin color differences between men and women. There is a delicate balancing act between skin color, vitamin D synthesis, and UVR protection. Women need to produce enough vitamin D in order to ensure enough calcium and its absorption to the baby and then ensure there are ample amounts to replace what the baby took while in the womb in order for future pregnancies to be successful. Sexual selection cannot explain the observed gradation in skin color between the races and ethnies of the human race. In my opinion, the only explanation for the observed explanation is the fact that skin color evolved due to climatic demands, while independent justification exists for the hypothesis as a whole (Jablonski and Chaplin, 2010).
I don’t see any way that sexual selection can explain the observed gradation in skin color around the world. Skin color is very clearly an adaptation to climate, though of course, cultural customs could widen the skin color differences between the sexes, and make women lighter over time. Nevertheless, what explains the observed skin gradation is adaptation to climate to ensure vitamin D synthesis among a slew of other factors (Jones et al, 2018). Sexual selection, while it may explain small differences between the sexes, cannot explain the differences noted between the native human races.
The Vitamin D Receptor and the Updated VDH
2850 words
The relationship between vitamin D and the vitamin D receptor (VDR) has been found to be of recent importance in explaining the modulation of gene expression. The VDR helps us adapt to the climate, is epistatic with skin color genes, and so on. Due to the importance of the VDR, vitamin D, and another nutrient I’ve discussed in the past—folate—this drives the argument that the need to produce vitamin D was an important factor in the evolution of skin colors around the world as migrations out of Africa took place. It is also important to note that other competing hypotheses are not necessarily alternative hypotheses to the VDH (which is short for vitamin D-folate hypothesis), since there is significant overlap between them due to what we now know about the roles of vitamin D, folate (especially due to what we know now about how vitamin D, folate and the VDR regulate gene expression),the VDR, and skin color genes. Thus, the theories have been integrated and the updated hypothesis takes into account the other theories which has significant overlap with the VDH.
Jones et al (2018) is the most recent review of the VDH; in the review, they integrate new findings of folate, vitamin D, the VDR, and skin color genes with other supposedly competing hypotheses into a new and improved VDH model which will be discussed at the end of this article.
Vitamin D
Vitamin D is an important hormone (since it is a steroid, not a vitamin), which is the only one that is produced exogenously (from UV rays). Vitamin D is responsible for many physiologic functions including: regulating calcium levels by increasing calcium absorption, stimulates intestinal absorption of phosphate, stimulates osteoblasts which then produce receptor activator nuclear factor (RANKL) which then stimulates osteoclastogenesis which then activates osteoclasts for bone reabsorption (DeLuca, 2004). It has been further noted that around 5 percent of the human genome is under the influence of vitamin D (Jones et al, 2018).
Folate
Folate is an important water-soluble B vitamin. Since vitamin D and folate are linked by their sensitivities to UVR, then we must look at them independently and see what they do. In the case of folate, UVR causes folate degradation through the absorption of UVRs or, on the other hand, when folate oxidizes through free radicals after UVR exposure (Jones et al, 2018). So the hypothesis proposes that skin color in high UV areas evolved due to the need for protection of folate levels due to UVR degradation. On the other hand, depigmentation occurred in order for the body to produce adequate vitamin D in low UV areas.
Folate is needed to synthesize and repair DNA (Mahmood, 2014). Though common rebuttals to the VDH include supposed lack of evidence for the VDH, Jones et al (2018) write:
The potential impacts of a deficiency of these nutrients on natural selection is an ongoing debate and is a common argument raised against the vitamin D–folate hypothesis. However, these arguments often do not consider that the benefits of an adequate vitamin D and folate status on reproductive success extend far beyond their roles in maintaining reproductive health.
Vitamin D receptor
In recent years, it has been found that the VDR has had a profound influence on our adaptation to local climates our ancestors found themselves in after the trek out of Africa. Most cells and organs of the body have a vitamin D receptor (Wacker and Holick, 2013), so the importance of the VDR and certain genes involved in the production of skin color, vitamin D, and folate can be seen. Thus, evidence for the hypothesis would be differential expression of certain genes that are related to the VDR. Jones et al (2018) report on a few common VDR variants and ethnicity: FOK1 which has a lower frequency in African than European and East Asian populations, and Cdx2 which was highest in Africans and lowest in Europeans. Tiosano et al (2016) reported that multiple loci which are involved with the VDR gene display strong latitudinal clines, which is evidence for the hypothesis.
The VDR helps humans adapt to changes in UV radiation, it is “part of an evolutionary complex that adapts humans to changing UV radiation” (Hochberg and Templeton, 2010: 310). This is further corroborated by the fact that the VDR promoter and skin color genes are epistatic (Popsiech et al, 2014; Tiosano et al, 2016). Skin pigmentation levels, furthermore, determine plasma vitamin D levels and VDR autoregulation (Saccone, Asani, and Bornman, 2015).
The VDR works in concert with retinoic acid receptors (Schrader et al, 1993) which then bind to nucleotide base pairs called the vitamin D-responsive elements (VDRE) which then exert their effects on gene expression (Kato, 2000; Pike and Meyer, 2010; Janik et al, 2017).
Gene expression
Vitamin D elicits numerous functions on gene expression through the VDR, by binding elements of vitamin D to the target genes. Since the VDR works together with other receptors that bind to the VDRE, they can have strong effects on gene expression. Now, we know that vitamin D and folate are important for humans. We know that the VDR gene appears to be under strong selection, though only in the context of other genes (Tiosano et al, 2016). Thus, the VDR—along with folate and vitamin D—are extremely important for gene expression and the adaptation of the human body to differing climates.
Competing hypotheses
Skin barrier hypothesis
The skin barrier hypothesis (SBH) proposes that dark skin color arose to protect against environmental damage. This hypothesis is based on the fact that darker-pigmented peoples posess an enchanced barrier function in comparison to ligher-pigmented people, which is mainly due to the role of melanin in the scattering of UVR across the skin (Jones et al, 2018). Jones et al state that this hypothesis is “proposed as a discrete theory to the vitamin D-folate hypothesis“, but since both vitamin D and folate both have other responsibilities in the human body such as the development of skin structure, and the development of defense mechanisms that protect against UV radiation including heat and microbial stressors.
Folate may also have another important role in the human body: regulating the production, and stabilizing tetrahydrobiopterin. Melanin supports folate from UVR degradation, which then supports folate’s influence on melanin. But, as Jones et al write, tetrahydrobiopterin also acts as a cofactor in the synthesis of nitric oxide which is important in regard to vasoconstriction (blood vessel constriction). Vasoconstriction is related to increased heat flow since blood vessels are constricted, along with an increase in heart rate. As I have noted in the past, shivering revs the body’s metabolism in cold clmates in order to produce ample heat. Jones et al (2018) write:
From an evolutionary perspective, our ability to maintain vasodilation/vasoconstriction mechanisms would have been important in surviving varying UVR environments. As these mechanisms may been seen as relatively short-term responses to temperature changes, they are likely to be of greater importance in temperate UVR environments rather than environments of high UVR. This is supported by nitric oxide dependent vasodilation shown to be reduced in darkly skinned populations [59]. This suggests that vasodilation processes offer no advantage in extreme UVR environments but may be important in temperate UVR environments, where seasonal and daily temperature fluctuations are seen.
Thus, since there would be no advantage for this mechanism in equatorial climates, it must be for more colder, Arctic climates which further lends credence to the VDH. (Since vitamin D and folate play many roles in regard to human physiologic adaptation to climate, along with the VDR.)
Metabolic conservation hypothesis
This hypothesis proposes that our ancestors became depigmented after the migrations out of Africa since there was a need to draw energetic resources away from melanin production and move that energy that would have been for melanin production for other metabolic processes that a population would need in a colder environment. Thus, it is argued that the lighter skin of European and East Asian populations can be explained by the need energetic resources being moved away from pigmenting the skin to other, more important, metabolic processes that the ancestors of Europeans and East Asians experienced. But this hypothesis has numerous premises of the VDH, including the main premise: that human skin depigmented as we migrated into areas with fewer UV rays (Jones et al, 2018). Thus, vitamin D was extremely important in driving the effects of vasodilation/vasoconstriction.
Clearly, the role of vitamin D in the adipose tissue was important for human adaptation to colder climates. Since lighter skin can produce more vitamin D in low UV climates, this was another factor that helped when we left Africa: skin lightened for better vitamin D synthesis. Since vitamin D synthesis is related to gene expression and expression of about 5 percent of our genomes, the production of more vitamin D was beneficial. So depigmentation, while being primarily due to low UV radiation, can also be seen to allow for more efficient physiologic responses and adaptations to the newer, colder climates.
Skin mutagenesis hypothesis
The last competing theory is the skin mutagenesis hypothesis. This hypothesis proposes that skin pigmentation arose as a mechanism to protect against various skin cancers. The hypothesis is based on the fact that darker-pigmented individuals are at lower risk of developing skin cancers since their skin pigmentation can fight off UV radiation. Of course, knowing what we know about vitamin D and folate, these two agents would be involved regarding this hypothesis, since both agents have photoprotective effects. Vitamin D is extremely important to DNA repair (Graziano et al, 2016), as vitamin D reduces cell and DNA damage.
Though many authors dispute the claims of this hypothesis since the effects of skin cancer would occur after the reproductive years and would thusly not have an effect on natural selection for skin color. Though those who argue for the validity of the hypothesis propose that it would help in hunter-gatherer peoples whose old train their young their ways of life.
Since these interactions have between these variables have been verified at the molecular genetic level, this lends even more credence to the VDH. (The findings inclue the frequency of common VDR variants between different ethnic groups, to UVR and folate metabolism genes which were found to be significantly associated with the frequency of 16 common folate variants and skin pigmentation in a genomic analysis of 30,000 people which were novel relationships; Jones et al, 2018a). These findings discussed by Jones et al (2018b) “indicate the existence of interactions between UVR, skin type, and vitamin D and folate genes, and they provide supporting molecular evidence for the vitamin D–folate hypothesis.”
Sexual selection
Madrigal and Kelly (2007a) tested a sexual selection hypothesis proposed by a few proponents of the sexual selection hypithesis. Madrigal and Kelly (2007a) tested the hypothesis that skin color reflectance should be positively correlated with distance from the equator. They, however, showed that the pattern in skin color dimorphism seen around the globe was not consistent with the sexual selection hypothesis, and thus their data did not lend credence to the sexual selection hypothesis. The hypothesis states that in areas with low UV radiation, environmental selection for skin color should be relaxed and there should be a higher rate of sexual dimorphism in peoples from northerly climates due to sexual selection for lighter-skinned women. Nevertheless, the data compiled by Madrigal and Kelly (2007a) do not lend credence to the hypothesis.
Frost (2007) responded that Madrigal and Kelly (2007a) presumed that sexual selection was equal in all areas, but was constrained by natural selection for dark skin. Frost (2007) also states that sexual dimorphism in human skin color may not be able to be expressed in lighter-skinned populations at higher latitudes. Frost’s objections stem from the fact that Madrigal and Kelly tested a specific hypothesis proposed by proponents of the sexual selection hypothesis, though Madrigal and Kelly hope that Frost can test his hypotheses. However, I think it’s a moot issue. Sexual selection for women occurred after selection for light skin due to vitamin D synthesis which ensured more calcium for pregnancy and lactation.
Thusly, sexual selection for lighter skin would continue to ensure ample vitamins for women and their pregnancies and lactation to feed their babies. This would further be butressed by the fact that vitamin D exerts effects on the adipocites which lends even more credence to the claim that light skin evolved first for vitamin D synthesis. Vitamin D then exerted effects on the adipocite since more vitamin D could be produced in the absence of high levels of UV, which then aided in human physiologic adaptations to climate.
Integration of current skin color theories
As can be seen from the competing theories, they are not necessarily explaining different things, and each supposed competing theory has an aspect from the VDH in it. Thusly, it is possible to integrate the so-called competing theories into a larger explanatory framework.
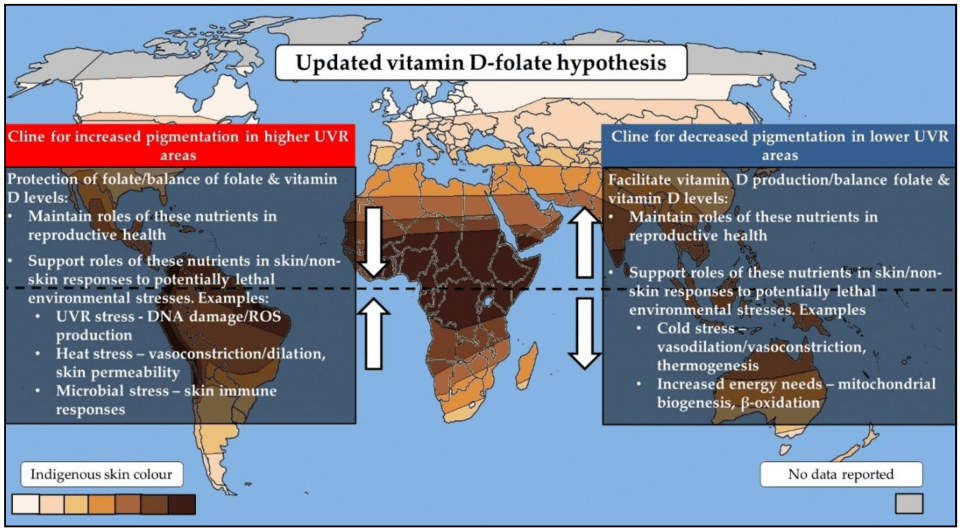
Jones et al (2018b) update the VDH by integrating the other theories into it, since they are similar and do not contradict the VDH (since aspects of each one can be used to explain different aspects of the VDH). The updated hypothesis is thus:
Vitamin D and folate have differing sensitivities to UVR. Vitamin D can be synthesized following UVR exposure, folate may be degraded. So the VDH proposes that the two differing skin colors (light and dark) evolved at differing latitudes as a “balancing mechanism” to maintain adequate levels of the two agents vitamin D and folate. Since adequate levels of vitamin D and folate were maintained, there would be no ill health effects after migrating into colder climates. Vitamin D and folate both act as photoprotectors of the skin and can decrease environmental stressors. Vitamin D also exerts important effects on adipocites—both types—which then further aid in human physiologic adaptations to the cold. Perhaps most importantly, the VDR and skin color genes are epistatic—the VDR is imperative in the human body’s adaptation to new climates.
The latest research (reviewed by Jones et al, 2018b) show strong support for the interaction between genes and folate/vitamin D processes with skin pigmentation and UVR.
Health Disparities
Health disparities due to vitamin D deficiency are well-noted in the literature. Human migrations over the recent centuries and decades have caused environmental mismatches between a population’s adapted skin type and current UV level in the location the population migrated to. Many darkly-pigmented people now live in areas with low UVR, and thusly suffer from health consequences. This leads to them either not having an adequate vitamin D-folate balance along with the risk of not having the adequate skin protections for a given environment, since UV rays influence folate and vitamin D production and so, a mismatched skin color to UV environment would cause problems for skin protection since the environment is not ancestral to that certain skin color.
As I have previously noted, it has been argued that blacks are not vitamin D deficient, and thusly not vitamin D deficient. Though, these claims rest on a slew of false arguments that have since been rebutted. It has been argued that since blacks are deficient in vitamin D, which begins in the womb, and vitamin D deficiencies cause changes in large and small arteries and arterials, that vitamin D deficiency could be the cause of higher rates of hypertension in black Americans (Rostand, 2010).
Conclusion
The role of vitamin D, folate, the VDR, and certain genes is under further invesitgation. This group of agents exert powerful effects on human physiology which then help with the adaptation of humans to differing climates. Folate and the vitamin D receptor play a crucial role in protecting the skin from environmental and microbial stressors. Vitamin D and the VDR are expressed in the adipose tissue, while vitamin D regulates adipogenesis and adipocite apoptosis (Abbas, 2017). Further evidence shows that there are multiple loci that are involved in the VDR that show latitudinal clines (Tiosano et al, 2016). One of the most things that lends credence to the VDH is the fact that the VDR and skin color genes are epistatic and help humans adapt to climates.
The VDH is in great shape, contrary to popular belief (Elias, Williams, and Bikle, 2016). The VDH is one of the only games in town to explain the skin color gradient noticed around the world, with vitamin D being the only agent that accounts for skin color differences. The VDH explains how and why human skin color is vastly different, and the main reason is adaptation to UV rays—or lack thereof.
Grant (2018) concludes that:
The UVB–vitamin D–cancer hypothesis has considerable supporting scientific evidence from a variety of study types: geographical ecological, observational, and laboratory studies of mechanisms, as well as several clinical trials.
Clearly, the VDH explains the incidence of the observed skin gradiation around the world the best out of the so-called competing hypotheses (which are similar enough to the VDH to where they can be absorbed into the VDH). Most importantly, the VDH predicted a novel fact—that molecular genetic evidence would show that light skin evolved independently numerous times in our lineage (Jablonksi and Chaplin, 2009).
Happy Darwin Day: The Modern Synthesis Has Causation in Biology Wrong
2800 words
The first Darwin Day I started writing just for this day, I wrote about (and defended Darwin’s words) how both Creationists and evolutionists who are themselves evolutionary progressionists twist Darwin’s words for their own gain. Darwin never wrote in The Descent of Man that the ‘higher races’ would take out ‘the lower races’, but that doesn’t stop Creationists and evolutionists—who I presume have not read one sentence in Darwin’s words from one of his books—from taking what Darwin meant out of context and attributing to him beliefs he does not hold. This year, though, I am going in a different direction. The Modern Synthesis (MS) has causation in biology wrong. The MS upholds the ‘gene’ as one of the highest seats in evolutionary biology, with a sort of ‘power’ to direct. Though, as I will show, genes do nothing unless transcribed by the system. Since the MS has causation in biology wrong, then we either need to extend or replace the MS.
To begin, Darwin, without knowledge of genes or other hypothesized units of inheritance, had a theory of inheritance in which things called ‘gemmules’ (what Darwin called heritable molecules) were transmitted to offspring (Choi and Mango, 2014). It’s ironic, because Darwin’s theory of inheritance was one of the more Lamarckian theories of inheritance in his day, and Darwin himself sympathized with the Lamarckian view of evolution—he most definitely did not discard it like modern-day Darwinists do. Darwin suggested that these gemmules circulated in the body and that some were used for the regeneration of some bodily tissues, but most aggregated in the reproductive organs (Jablonka and Lamb, 2015: 23). Further, according to Darwin, gemmules were not always immediately used but could reappear later in life or even be used in future generations. Darwin even said that “inheritance must be looked at as a form of growth” (Darwin, 1883, vol 2, p. 398; quoted by Jablonka and Lamb, 2015: 24).
The crux of the MS is the selfish gene theory of Dawkins (1976). Dawkins (1976, 2006) writing “They are in you and me; they created us, body and mind; and their preservation is the ultimate rationale for our existence.” “They”, of course, being genes. The gene has been given a sort of power that it does not have, but has been placed on it by overzealous people, quick to jump to conclusions while we still have yet to understand what ‘genes’ do. The MS—with the selfish gene theory—is at the forefront of the neo-Darwinist revolution, that evolution is gene-centered, with genes playing the starring role in the evolutionary story.
Though, numerous researchers are against such simplistic and reductionist viewpoints of evolution, mainly the gene-centered view of evolution pushed by the MS. There is no privileged level of causation in biology (though I will state later in this article that I think ATP comes close to it) (Noble, 2016).
Neo-Darwinists, like Richard Dawkins, overstate natural selection’s importance regarding evolution. They elevate the gene’s overall importance. In the quote from Dawkins above, where he stated that “they” (genes) “created us, body and mind”, he is implying that genes are a sort of ‘blueprint’, like a ‘plan’ or ‘recipe’ for the form of the organism. But this was taken care of by Susan Oyama in her 1985 book The Ontogeny of Information where she writes on pages 77:
“Though a plan implies action, it does not itself act, so if the genes are a blueprint, something else is the constructor-construction worker. Though blueprints are usually contrasted with building materials, the genes are quite easily conceptualized as templates for building tools and materials; once so utilized, of course, they enter the developmental process and influence its course. The point of the blueprint analogy, though, does not seem to be to illuminate developmental processes, but rather to assume them and, in celebrating their regularity, to impute cognitive functions to genes. How these functions are exercised is left unclear in this type of metaphor, except that the genetic plan is seen in some peculiar way to carry itself out, generating all the necessary steps in the necessary sequence. No light is shed on multiple developmental possibilities, species-typical or atypical.“
The genes-as-blueprints canard is one that is heavily used by proponents of the MS. Oyama also writes on page 53 “Just as traditional thought placed biological forms in the mind of God, so modern thought finds many ways of endowing the genes with ultimate formative power, a power bestowed by Nature over countless millennia.” This same sentiment from Oyama is also echoed by developmental systems theorist and psychologist David Moore in his book The Dependent Gene: The Fallacy of “Nature VS. nurture”, where he writes:
Such contextual dependence renders untenable the simplistic belief that there are coherent, long-lived entities called “genes” that dictate instructions to cellular machinery that merely constructs the body accordingly. The common belief that genes contain context-independent “information”—and so are analogous to “blueprints” or “recipes”—is simply false. (p. 81) (Quoted from Schneider, 2007)
Environmental factors are imperative in determining which protein-coding exons get read from a cistron, when and how often. So the very concept of a gene depends on the environment and environmental inputs, and thusly gene ABC does not code for trait T on its own.
When it comes to epigenetics (defined here as inherited changes in gene expression with no genetic change to the genome), this completely changes how we view evolution.
The underlying nucleotide sequence stays the same but differences are inherited due to environmental stressors. I’ve stated in the past that these inherited marks on the genome (through histone modification, DNA methylation, which then alter the chromatin structure of the DNA. Further, this would show up on heritability estimates as ‘genetic’ when the ’cause’ was ‘environmental’ in nature (which is also yet another reason that heritability estimates are inflated).
DNA methylation, histone modification and noncoding RNA all can affect the structure of chromatin. As of now, the mechanisms of mitotic inheritance aren’t too well known, but advances in the field are coming.
If you want to talk the P and F1 generations regarding transgenerational epigenetics, then you must realize that these changes do not occur on the genome, the genome remains the same, just certain genes are expressed differently (as I’m sure you know). Though mi-MRNA signals can change the DNA methylation patterns in the F2 sperm which then is replicated in meiotic and mitotic cycles (Trerotola et al, 2015).
For another similar process on how DNA methylation persists, this (semiconservative) replication of DNA methylation occurs on both strands of the DNA which then become hemimethylated DNA which can then become fully methylated by methylase maintenance. So chromatin structure affects the genetic expression of the eukaryotic genome which then becomes the basis for epigenetic effects. Xist RNA also mediates the X-chromosome deactivation. This doesn’t even get into how and why the microbiome can also affect gene expression (which has also been called ‘the second genome’ (Zhu, Wang, and Li, 2010) with other authors calling it an ‘organ’ (Clarke et al, 2014; Brown and Hazen, 2015) this can also affect gene expression and heritable variation that becomes the target of selection (along with the other modes of selection) (Maurice, Haiser, and Turnbaug, 2014; Byrd and Segre, 2015). This shows that gene expression in the F2 and F3 generations is not so simple, and that other factors such as our gut microbiota can also affect gene expression and stressors experienced by parents and grandparents can also be passed to future generations, and may have a chance of becoming part of heritable variation that natural selection then acts on (Jablonka and Lamb, 2015).
The point of the debate with neo-Darwinists is over causation: do genes hold this ‘ultimate formative power’ as people like Dawkins contest? Or are genes nothing but ‘slaves’, passive, not active, causes as Denis Noble writes in his 2016 book Dance to the Tune of Life. (Noble, 2008 discusses genes and causation, again showing that there is no true causation, but getting technical, ATP is up there in the ‘chain’, if you want to get literal. The point is that genes do not have the ‘power’ that the neo-Darwinists think they do, they’re just slaves for the intelligent physiological system.)
When discovering the structure of DNA, Francis Crick famously announced to his drinking companions in a Cambridge tavern that he had discovered ‘the secret of life’. The director of his Institute, Max Perutz, was rather more careful than Crick when he said that DNA was the ‘score of life’. That is more correct since a musical score does nothing until it is played, DNA does nothing until activated to do so.
[…]
Recent experimental work in biological science has deconstructed the idea of a gene, and an important message of this book is that it has thereby drthroned the gene as a uniquely privileged level of causation. As we will see, genes, defined as DNA sequences, are indeed essential, but not in the way in which they are often portrayed. (Noble, 2016: 53)
A 2017 paper titled Was the Watchmaker Blind? Or Was She One-Eyed?, Noble and Noble (2017) write that organisms and their interacting populations have evolved mechanisms so that they can harness blind stochasticity, thereby generating functional changes to the phenotype as to better respond to environmental challenges. They put forth a good argument, though it really makes me think because I’ve been such a staunch critic against evolution having a ‘direction’ and against the ‘teleological view’ of evolution: “If organisms have agency and, within obvious limits, can choose their lifestyles, and if these lifestyles result in inheritable epigenetic changes, then it follows that organisms can at least partially make choices that can have long-term evolutionary impact.”
Noble and Noble (2017) argue (using Dawkins’ analogy of the Blind Watchmaker) that humans are the only Watchmakers that we know of. Humans evolved from other organisms. The ability to become a Watchmaker has evolved. Ergo, there is no surprise that there is directed agency for other organisms that directs their evolution too. There are several processes, they conclude, that could account for directed evolutionary change which are “targeted mutation, gene transposition, epigenetics, cultural change, niche construction and adaptation” (Noble and Noble, 2017). Niche construction, for instance, is heavily pushed by Kevin Laland, author of the book Darwin’s Unfinished Symphony: How Culture Made the Human Mind who has a few papers and featured it heavily in his new book. Either way, these ways in which organisms can in a way direct their own evolution are not covered by the MS.
Though I couldn’t end this article without, of course, discussing Jerry Coyne who goes absolutely crazy at people pushing to either extend or replace the MS. His most recent article is about Kevin Laland and how he is “at it again” touting “a radically different view of evolution”. It seems as Coyne has made up his mind and that the MS is all there is—he believes it is no problem for our current understanding of evolutionary theory to absorb things such as niche construction, epigenetic inheritance, stochasticity, and even (way more controversially) directed mutations. Coyne has also criticized Noble’s attacks on the MS, though Noble came back and responded to Coyne during a video presentation.
Lastly, Portin and Wilkins (2017) review the history of the gene, and go through different definitions it has been given over the decades. They conclude in this paper that they “will propose a definition that we believe comes closer to doing justice to the idea of the “gene,”
in light of current knowledge. It makes no reference to “the unit of heredity”—the long-standing sense of the term—because we feel that it is now clear that no such generic universal unit exists.” Writing on page 1361-1362:
A gene is a DNA sequence (whose component segments do not necessarily need to be physically contiguous) that specifies one or more sequence-related RNAs/proteins that are both evoked by GRNs and participate as elements in GRNs, often with indirect effects, or as outputs of GRNs, the latter yielding more direct phenotypic effects. [GRNs are genetic regulatory networks]
This is similar to what Jablonka and Lamb (2015: 17) write:
Although many psychiatrists, biochemists, and other scientists who are not geneticists (yet express themselves with remarkable facility on genetic issues) still use the language of genes as simple causal agents, and promise their audience rapid solutions to all sorts of problems, they are no more than propagandists whose knowledge or motives must be suspect. The geneticists themselves now think and talk (most of the time) in terms of genetic networks composed of tens or hundreds of genes and gene products, which interact with each other and together affect the development of a particular trait. They recognize that whether or not a trait (a sexual preference, for example) develops does not depend, in the majority of cases, on a difference in a single gene. It involves interactions among many genes, many proteins and other types of molecule, and the environment in which an individual develops.
The gene as an active causal actor has been definitively refuted. Genes on their own do nothing at all, until they are transcribed by the intelligent physiological system. Noble likens genes as slaves that are used by the system to carry out processes by and for the system. So genes are caused to give their information by and to the system that activates them (Noble, 2011). Noble’s slave metaphor makes much more sense than Dawkins’ selfish metaphor, since genes are used like slaves by the system, the genes are then caused to give their information by and to the system that activates them, which shows how they are a passive, not active, cause, completely upending the MS and how it views causation in biology. Indeed, Jablonka and Lamb state that one of their problems with Dawkins is that “Dawkins assumes that the gene is the only biological (noncultural) hereditary unit. This simply is not true. There are additional biological inheritance systems, which he does not consider, and these have properties different from those we see in the genetic system. In these systems his distinction between replicator and vehicle is not valid.”
So, both Gould and Dawkins overlooked the inheritance of acquired characters, as Jablonka and Lamb write in their book. They argue that inherited variation had a large effect on the evolution of species, but admit that evidence for the view is scant. They write on page 145 “If you accept that heritable epigenetic variation is possible, self-evidently some of the variants will have an advantage relative to other variants. Even if all epigenetic variations were blind, this would happen, and it’s very much more likely if we accept that a lot of them are induced and directed.” Not everything that is inherited is genetic.
DNA is found in the cell, and what powers the cell? ATP (adenosine triphosphate). Cells use and store ATP to carry out their functions (Khakh and Burnstock, 2016). Cells produce ATP from ADP and Pi. Cells use exergonic reactions to provide the energy needed to synthesize ATP from ADP and Pi. The hydrolysis of ATP provides the energy needed to drive endergonic actions.So the cells continuously produced more ATP from ADP and Pi to then carry out diverse functionings across the body. So, in a way, you can argue that one of the ultimate causes is ATP since it has to power the cell, then you can look at all of the other reactions that occur before ATP is created and privilege that part of the chain, but there will never be some ultimate causation, since, as Noble argues in his book Dance to the Tune of Life, there is no privileged causation in biology.
In conclusion, evolution, development, and life, in general, is extremely complex. Paradigms like the selfish gene—a largely reductionist paradigm—do not account for numerous other factors that drive the evolution of species, such as targeted mutation, niche construction etc. An extended evolutionary synthesis that integrates these phenomena will better be able to describe what occurs to drive the evolution of species, and if the directed mutation idea has any weight, then it will be interesting to see how and why certain organisms have evolved this ability. It’s ironic how the MS is being defended as if it is infallible—like it can do no wrong and that it does not need to be added to/extended or replaced by something else that incorporates the phenomena brought up in this article.
Either way, a revolution in modern biology is coming, and Darwin would have it no other way. The Modern Synthesis has causation in biology wrong: the gene is not an active agent in evolution, it only does what it is told by the intelligent physiological system, and so we must look at whole organisms and not reduce organisms down to genes, but we must look at the whole organism—a holistic view of the organism, not one that is reduced down to just ‘the genes’, because there is no privileged level of causation in biology (Noble, 2016).
Race and Nutrition
2600 words
What we eat is important. What we eat can increase or decrease our lifespan. But do different races digest and metabolize different macro and micronutrients differently? On a racial level in terms of individual diet, would one individual benefit from adopting the diet of their ancestors over another diet? Many claims have been made like this in the past few years, such as Europeans evolving to eat plants and grains. This, some people would presume, implies that if you have a certain ancestry then you must eat a certain diet or take different steps in regard to nutrition. I will show this is wrong and that, at least in regard to health and nutrition, individual variation matters more than racial variation (don’t call Lewontin’s fallacy on me. This is not a fallacy).
Different genetically isolated breeding populations evolved eating different diets based on what they had in their environment. Over time, humans eventually developed agriculture and then changed the course of human evolution forever (Cochran and Harpending, 2009). This then leads to large changes in how our genes are expressed and how our microbiome metabolizes nutrients and food we ingest. The advent of farming was, obviously, pivotal to human evolution (Cochran and Harpending, 2009). This then lead to heritable changes in the genome brought on by new foods the farmers ate. This also started the environmental mismatches we now have in our modern world, which is the cause for rising obesity rates and a large part of the cause of so-called diseases of civilization (for a discussion of these matters, see Taubes, 2008, chapter 5; see also page 8 in this summary of his book on diseases of civilization and also see Burkitt, 1973; Cordain, Eades, and Eades, 2003; Sharma and Majumdar, 2009; Sikter, Rihmer, and Guevara, 2017. For an outstanding review on the subject, read Daniel Lieberman’s 2013 book The Story of the Human Body: Evolution, Health, and Disease for in-depth discussions on this point and more in regard to nutrition and our evolutionary history).
Studies come out all the time saying that X population evolved eating Y food therefore Z. Then, people not privy to nutrition science, jump to large sweeping conclusions (mostly laymen and journalists, who are also laymen). These assumptions imply that people’s metabolic systems aren’t, first and foremost, based on an individual level with individual variation in physiologic and metabolic traits. This, I will show, is the reason why these studies don’t mean you should change your diet to what your ancestors supposedly ate based on these studies (though as I have argued in the past, high consumption of processed foods lead to obesity, insulin resistance, diabetes etc which is the cause of a lot of the modern-day maladies currently present in our population today). This assumption is wrong on numerous levels.
Buckley et al (2017), using data from the 1000 Genomes Project (see also Via, Gignoux, and Burchard, 2010), identified novel potential selections in the FADs region. The 1000 Genomes Project tested the genomes of 101 Bronze Age Europeans. They show that SNPs which are associated with arachidonic acid and eicosapentaenoic acid has been favored in Europeans since the Bronze Age (the selection for arachidonic acid being due to milk consumption which is a form of niche construction; see Laland, Odling-Smee, and Feldman, 1999; Laland, Odling-Smee, and Feldman, 2001; Laland and Brown, 2006; Rendell, Fogarty, and Laland, 2011; Laland, et al, 2016; but see Gupta et al, 2017 for a different view which will be covered in the future). They also hypothesize that differences in the selection of these regions is different in different population, implying different epigenetic changes brought on by diet (more on this later).
The FADS1 gene codes for an enzyme called fatty acid desaturase 1 which desaturates n3 and n6 which then catalyzes eicosapentaenoic and Arachidonic acid (Park et al, 2009). These genes code for enzymes that then aid in the breakdown of fatty acids. So, by testing Bronze Age Europeans and comparing their genomes with modern-day Europeans, researchers can see how the expression of genes changed and then work backward and hypothesize how and why the differing gene expression occurred.
The regions selected for are involved in processing n3 and n6 fatty acids. We need a certain ratio of them, and if either is thrown out of whack then deleterious effects occur. This, of course, can be seen by comparing our ratio of n3 to n6 fatty acid consumption with our ancestors’, who ate a 1:1 ratio of n3 to n6 (Kris-Etherson et al, 2000) which you can then compare to our n3 to n6 ratio, which is 14 to 25 times higher than it should be. The authors state that n6 is important, but it’s only important to have the correct ratio, having too much n6 is not a good thing (as I have covered here).
Twenty percent of the dry weight of the brain is made up of long-chain polyunsaturated fatty acids (Lassek and Gaulin, 2009). Therefore it is pivotal we get the correct amount of n3 fatty acids for brain development both in vitro and during infancy, the best bet being to breastfeed the babe as the mother packs on fat during pregnancy so the babe can have PUfAs during its time on the womb as well as during infancy through breastfeeding.
About 85kya selective sweeping occurred in Africa on the FADs genes. Buckley et al (2017) write: “Humans migrating out of Africa putatively carried mostly the ancestral haplotype, which remained in high frequency in non-African populations, while the derived haplotype came close to fixation in Africa. It is unclear why positive selection for the derived haplotype appears to be restricted to Africa. Mathias et al. (2012) suggested that the emergence of regular hunting of large animals, dated to ∼50 kya, might have diminished the pressure for humans to endogenously synthesize LC-PUFAs.” This is true. There is a wealth of important fatty acids in the fatty and muscle tissue of animals, which we need for proper brain functioning and development.
They also write about a study on the Inuit that proves that certain alleles have been selected for that have to do with fatty acid metabolism, which I have also covered in the past in a response to Steve Sailer. Nevertheless, on a population level, this is worth it, but individual variation in metabolism matters more than population. In the article, Sailer implied—with a quote from New York Times science editor Carl Zimmer—that the Inuit have certain gene variants that influence fatty acid metabolism in that population. Sailer goes on to write “So maybe you should try different diets and see if one works better for you.” Of course, you should. However individual variation is more important than racial variation. (It’s also interesting to note that these genes that are expressed on the Inuit are also related to height.)
Nevertheless, it is true that selection occurred on these parts of the genome in these populations studied by Buckley et al (2017), but to claim that all populations wouldn’t benefit from a low carb, high fat diet is not true. I do agree with Sailer on, in the future, the scanning of individual genomes to see which diet would have a better effect. Though I would insist that most, if not all, humans should eat a higher fat lower carb diet.
Buckley et al (2017) cite a study (Mathieson et al, 2015) which “provides strong evidence of selection in the FADSregion in Europe over the past 4,000 years, in addition to the patterns of selection already reported in Africans, South Asians, and the Inuit.” Buckley et al (2017) also cite a study (Pan et al, 2017) which shows an SNP, rs174557, regulates FADS1.
In their analysis, they showed that “this variation is largely attributable to high differentiation between two haplotype clusters: a cluster widespread in Africa, largely containing derived alleles and possibly subject to a selective sweep (Mathias et al. 2011,, 2012), and an ancestral cluster, which is present across Eurasia.” They also showed that Neanderthal genomes cluster with the derived cluster, which is present in Africans, while Denisovans cluster with the ancestral cluster, which Eurasians also have.
Buckley et al (2017) write: “Thus the derived alleles appear to promote expression of FADS1 while simultaneously abating the expression of FADS2.” This is important to keep in mind for the end of this article when I talk about nutrition and how it affects the epigenome which can then become heritable in a certain population.
Buckley et al (2017) also confirm the results of the European sample using the Nurses Health Study and the Health Professionals follow-up study GWASs: “These results reinforce the associations with cholesterol from the GLGC GWAS. This confirms the hypothesized phenotypic effect of the selected variants in terms of increased EPA and ARA levels of the putatively positively selected variants in the European population.”
Selective (dietary) pressures on the three populations tested (Africans, Europeans and South Asians) have “have driven allele frequency changes in different FADS SNPs that are only in weak LD with each other [LD is linkage disequilibrium which is the nonrandom associations of alleles at different loci in a given population]” (Buckley et al, 2017). Further, the alleles (FADS1 and FADS2) that were under selection in Europeans were strongly associated with lipid metabolism, specifically reduced linoleic acid levels. An opposite pattern was noticed in the Inuit, where selection acted to “decrease conversion of SC-PUFAs to LC-PUFAs to compensate for the relative high dietary intake of LC-PUFAs.” The allele under selection was associated with a decrease in linoleic acid levels and an increase in eicosapentaenoic acid, which may possibly be due to improved metabolism in converting LC-PUFAs from SC-PUFAs.
Buckley et al (2017) hypothesize that the cause is eating a more plant-based diet which is rich in fatty acids (n6 and n3) while a subsequent decrease in fatty animal meats occurred. Of course, relative to hunter-gatherer populations, the increased plant consumption brought on by agriculture caused different methylation on the genome which then eventually became part of the heritable variation. So, of course, farmers would have eaten more plants and the like, which one then select for the production of SC-PUFAs to LC-PUFAs. This of course began at the dawn of agriculture (Cochran and Harpending, 2009).
Of course, this can help guide individual diets as we better map the human genome. These studies, for instance, can be used as guides for individual diets based on ancestral evolution. More studies, of course, are needed.
Also, in an email with correspondence with Arstechnica, one of the authors, Nelson Rasmussen, stated: “Of course, within the last century there have been drastic changes in the diets in many areas of Europe. Diets have typically become more caloric with a higher intake of simple sugars, and perhaps also more rich in proteins and fat from animals. So selection is unlikely to be working in exactly the same way now.”
Though the strong claim from Arstechnica that “This is another nail in the coffin for the scientific validity of paleo diets” is a strong claim which needs much more evidence because low carb high-fat diets are mostly best for people since their insulin levels aren’t spiked too much which then leads to obesity, diabetes and along with it hyperinsulinemia.
Now I need to talk about how epigenetics is involved here. Nutrition can alter the genome and epigenome (Niculescu and Lupu, 2011; Niculescu, 2012; Anderson, Sant, and Dolinoy, 2012) and cause permanent heritable variation in a population if a certain allele reaches fixation, since there is evidence that maternal and paternal dietary changes possibly affecting multiple generations (Rosenfeld, 2017; though see Burggren, 2016 for the view that there is no evidence for heritable epigenetic phenotype in the genome. I will return to this in the future; see also the Dutch Famine Study showing heritable epigenetic change from famine; Lumey et al, 1993; Heijmans, 2008; Stein et al, 2009; Tobi et al, 2009; Schulz, 2010; Lumey, Stein, and Susser, 2011; Hajj et al, 2014; Jang and Serra, 2014; Tobi et al, 2014). Of course, based on what a population eats (or does not eat), epigenetic changes can and will occur. This not only affects the expression of genes in the body, but also the trillions of gut microbiota in our microbiome that partly drive our metabolic functions. Diet can change the composition of the microbiome, diet can change the epigenome and gene expression, and the microbiome can also up- and down-regulate genes (Hullar and Fu, 2014) Lipid metabolism is also related to developmental epigenetic programming (Marchlewicz et al, 2016). They showed that circulating fatty lipids in the mother during pregnancy are associated with DNA methylation in the genomes of the child. This can also, of course, contribute to health and disease risk in the future for the affected infant. FADS1 is also involved here.
Nutritional factors also come into play in regards to epigenetic inheritance (Alam et al, 2015). The n3 PUFAs also affect gene expression and DNA methylation (Hussey, Lindley, and Mastana, 2017). Further, DNA methylation is also associated with FADS1 and, to a lesser extent, FADS2 (Howard et al, 2014). This is strong evidence that, of course, that what was reviewed above in regards to selection for certain alleles for fatty acid metabolism in certain populations was strongly driven by the consumption of certain foods. Epigenetic changes that occur both in the womb and previous generations like the grandparents’, for instance, also have an effect in regard to which genes are expressed in the baby in vitro as well as consequences for future generations. The study of epigenetics, along with transgenerational epigenetic inheritance, of course, will be very important for our future understanding of both the evolution of humans and the evolution of the human diet.
Finally, I need to touch on why this doesn’t really matter in terms of individual diet choice. The fact of the matter is, anatomic, physiologic, and metabolic variation within race trumps variation between it. Two different randomly selected individuals will have different anatomy, along with different organs missing (Saladin, 2010). This implies that the individual differences in these traits trump whatever racial selection occurred since the dawn of agriculture 10kya. This is why, in my opinion, one should not look to just their ancestry when choosing a diet and should always choose a diet based that’s good for them, individually. Now, I’m not saying that this research is useless in regards to healthy diets, however, increased consumption of processed foods is the cause of obesity since processed foods (high in carbs) spike insulin which lead to obesity and diabetes (insulin causes weight gain). So, obviously, full-on plant-based diets will lead to these maladies. Contrary to the Alternative Hypothesis’ thesis on race and nutrition, this doesn’t really matter, not at the individual level, anyway. This could have small implications in regard to the population as a whole, but as an effect on the diet of individuals? No. Individual variation in traits matters much more than racial variation here (again, don’t call Lewontin’s fallacy because I’ve explained my reasoning which is logically sound).
In sum, the SNPs associated with the increased expression of FADs1 and increased the production of eicosapentaenoic and Arachidonic acid in Europeans occurred around 5kya. These studies are interesting to see how diet and how we construct our niches leads to changes in the genome based on those changes that we enact ourselves. However, laypersons who read these popular science articles on the evolution of diet in human populations will then assume that since they have X ancestry then they must eat how their immediate ancestors ate. The Arstechnica article makes some strong claims that Buckley et al (2017) prove that the paleo diet is not a viable solution for diseases of civilization. Do not make sweeping claims about eating X and Y because your ancestors evolved in Z environment, because individual variation in metabolic and physiologic functioning is greater and matters way more than racial variation
[Note: Diet changes under Doctor’s supervision only.]
Small Brain, Normal IQ
1650 words
Emil Kirkegaard left a short commentary on John Skoyles’ 1999 paper Human Evolution Expanded Brains to Increase Expertise Capacity, not IQ, in which Emil writes in his article Evolution and imperfect mediators:
If we condense the argument, it becomes a little clearer:
John Skoyles (1999) [Condensed argument from Emil; paragraph 2] Brain expansion causes problems. Thus, whatever selected for increased brain size must have offered compensating benefits. People can have below average size brains yet exhibit normal intelligence. Thus, the compensating benefit offered by large brains is unlikely to be intelligence. Why should evolution have increased brain size with its associated problems for something smaller sized brains could have without expansion?
I merely edited out the unnecessary parts. Now try substituting some other trait, say fighting ability and some mediator of it.
Muscle size increases causes problems. Thus, whatever selected for increased muscle size must have offered compensating benefits. People can have below average size muscles yet exhibit normal fighting ability. Thus, the compensating benefit offered by large muscles is unlikely to be fighting ability. Why should evolution have increased muscle size with its associated problems for something smaller sized muscles could have without increase?
See the issue? This argument works for any imperfect physical underpinning of a trait, which is to say, basically all of them. Longer legs didn’t evolve for running well for some people with short legs run well. Bigger/stronger hears didn’t evolve for better cardio, because some people smaller/weaker hearts have good cardio. Longer arms didn’t evolve for fighting because some short armed people fight well. Darker skin didn’t evolve as a protection against sun exposure for some relative light skinned people don’t get skin cancer or sunburns. Larger eyes didn’t evolve for seeing better for some people with smaller eyes see well. Bigger ears… Bigger noses… Stronger hands… …
I don’t agree. Our brains sap about 20 percent of our daily energy needs while being 2 percent of our overall body mass whereas, in other primates, their brains cost about 9 percent of their daily energy needs (Fonseca-Azevedo and Herculano-Houzel, 2012).
In regards to Emil’s counterarguments, I’ll address them one by one:
Long legs: People with longer legs were better runners and could escape from predators and chase prey. People with shorter legs were killed.
Bigger/stronger hearts: Those with a larger heart (sans cardiomegaly) could run for longer distance (remember, we are distance runners; Carrier, 1984; Skoyles and Sagan, 2002; Bramble and Lieberman, 2004; Mattson, 2012) and so long legs and bigger/stronger hearts tie in with each other.
Long arms: This, again, goes back to our morphology in Africa. Long limbs are more conducive to heat dissipation (Lieberman, 2015). So those who had the right body plan for distance running could survive better during our evolutionary history.
Dark skin: A light-skinned person who spends enough time without protection in a tropical climate will develop skin cancer. (It is hypothesized that skin cancer is what caused the evolution of dark skin; Greaves, 2014, though this was contested by Jablonksi and Chaplin, 2014.)
Large eyes: Bigger eyes don’t mean better eyesight in comparison to smaller ones.
All in all, the brain size argument is 100 percent different from these arguments: large brains come with large problems. Further, there is evidence (which will be reviewed below) that people can live long, normal lives with half of their brain missing
The brain-size/IQ puzzle
The oft-repeated wisdom is that our brains evolved to such a large size so we could become more intelligent. And looking at when our brains began to increase (starting with erectus, which had to do with the advent of cooking/fire use), we can see that that’s when our modern body plan appeared. We can ascertain this by looking at Nariokotome boy, an erectus that lived about 1.6 mya.
Further, in regards to brain size, there was a man named Daniel Lyon. What was so extraordinary about this man is that, at the time of his death, had a brain that weighed 1.5 pounds (see Wilder, 1911)! Skoyles and Sagan (2002: 239) write:
Upon examination, anatomists could find no difference between it [Lyon’s brain] and other human brains apart from its size with one exception: The part of his brain attached to the brainstem, the cerebellum, was near normal size. Thus, the total size of Lyon’s cerebral hemisphere was smaller than would be suggested by a total brain weight of 1.5 lb. We do not know how bright he was—being a watchman is not particularly intellectually demanding—but he clearly was not retarded. A pound and a half brain may not be enough to manage a career as an attorney, a professor of theology, or a composer, but it was sufficient to let Lyon survive for 20 years in New York City.
Skoyles and Sagan (2002) review numerous lines of evidence of individuals with small brains/people with severe TBI living full lives, even having IQs in the average/above average range. They write (pg 238):
You would think that cutting out one-half of people’s brains would kill them, or at least leave them vegetables needing care for the rest of their lives. But it does not. Consider this striking story. A boy starts having seizures at 10 years of age when his right cerebral hemisphere atrophies. By the time he is 12, the left side of his body is paralyzed. When he is 19, surgeons decide to operate and remove the right side of his brain, as it is causing gits in his intact left one. You might think this would lower his IQ or leave him severely retarded, but no. His IQ shoots up 14 points, to 142! The mystery is not so great when you realize that the operation has gotten rid of the source of his fits, which had previously hampered his intelligence. When doctors saw him 15 years later, they described him as “having obtained a university doploma . . . [and now holding] a responsible administrative position with a local authority.” (18)
They also write about the story of an Argentinian boy who had a right hemispherectomy when he was 3-years-old who was notable for “the richness of his vocabulary and syntax” and also “attends English classes at school, in which he attains a high level of success (20; quote from Skoyles and Sagan, 2002: 238).
It is also a “medical myth that microcephaly (having a head smaller than two standard deviations (SD) below average circumference) is invariably linked to retardation.” (Skoyles and Sagan, 2002: 239).
There are some important things to be noted in regards to the study of Nariokotome boy’s skeleton and skull size. Skoyles and Sagan (2002: 240) write (emphasis mine):
So how well equipped was Homo erectus? To throw some figures at you (calculations shown in the notes), easily well enough. Of Nariokotome boy’s 673 cc of cortex, 164 cc would have been prefrontal cortex, roughly the same as half-brained people. Nariokotome boy did not need the mental competence required by cotemporary hunter-gatherers. … Compared to that of our distant ancestors, Upper Paleolithic technology is high tech. And the organizational skills used in hunts greatly improved 400,000 years ago to 20,000 years ago. These skills, in terms of our species, are recent, occurring by some estimates in less than the last 1 percent of our 2.5 million year existence as people. Before then, hunting skills would have required less brain power, as they were less mentally demanding. If you do not make detailed forward plans, then you do not need as much mental planning abilities as those who do. This suggests that the brains of Homo erectus did not arise for reasons of survival. For what they did, they could have gotten away with much smaller, Daniel Lyon-sized brains.
Lastly, I will touch on the fact that since we are running apes, that we need a narrow pelvis. As I stated above, our modern body plan came to be around 1.6 mya with the advent of erectus, which could be inferred from footprints (Steudel-Numbers, 2006; Bennett et al, 2009). Now the picture is beginning to become clearer: if people with brains the size of erectus could have intelligence in the modern range, and if our modern body plans evolved 1.6 mya (which is when our brains began to really increase in size due to metabolic constraints being unlocked due to erectus’ cooking ability), then you can see that it’d be perfectly possible for modern Homo sapiens to have brains the size of erectus while still having an IQ in the normal range.
Lastly, Skoyles and Sagan (2002: 245) write (emphasis mine):
Kanzi seems to do remarkably well with a chimp-sized brain. And while we tend to link retardation with small brains, we have seen that people can live completely normal lives while missing pieces of their brains. Brain size may enhance intelligence, but it seems we can get away without 3 pounders. Kanzi shows there is much potential in even 13 oz.
So Skoyles and Sagan do concede that brain size may enhance intelligence, however, as they have argued (and as Skoyles does in his 1999 paper), it is perfectly possible to live a normal life with half a brain, as well as have an average/above average IQ (as reviewed in Skoyles, 1999). So if people with erectus-sized brains can have IQs in the normal range and live normal lives, then brains must have increased for another reason, which Skoyles has argued is expertise capacity.
Large brains are, clearly, not needed for high IQs.
(Also search for this paper: Reiss, A. L., Abrams, M. T., Singer, H. S., Ross, J. L. & Denckla, M. B. (1996). Brain development, gender and IQ in children: A volumetric imaging study. Brain, 119, 1763-1774. where they show that there is a plateau, and a decrease in IQ in the largest brains; see table 2. I also reviewed some studies on TBI and IQ and how even those with severe TBI can have IQs in the normal range (Bigler, 1995; Wood and Rutterford, 2006; Crowe et al, 2012). Yet more evidence that people with half of their brains missing can lead normal lives and have IQs in the modern range.)
