
Home » Posts tagged 'HBD' (Page 5)
Tag Archives: HBD
IQ Test Construction, IQ Test Validity, and Raven’s Progressive Matrices Biases
2050 words
There are a lot of conceptual problems with IQ tests that I never see talked about. The main ones are how the tests are constructed (to fit a normal curve, no less); to the fact that there is no construct validity to the tests (IQ tests aren’t calibrated against a biological model like breathalyzers are calibrated against a model of blood in the blood stream); and how the Raven’s Progressive Matrices test is actually biased despite being touted as the most culture-free test since all you’re doing is rotating abstract symbols to see what comes next in the sequence. These three assumptions have important implications for the ‘power’ of the IQ tests, the most important being the test construction and validity.
I) IQ test construction
IQ tests are constructed with the assumption that we know what IQ tests test (we don’t) and with the prior ‘knowledge’ of who is or is not intelligent. Test constructors construct the tests to reveal presumed differences between individuals.
It is assumed that 1) IQ scores lie on a normal distribution (they don’t) and 2) few natural bio functions conform to this curve. Another problem with IQ test construction is the assumption that it increases with age and levels off after puberty. Though this, like the other things, has been built into the test by choosing items that an increasing proportion of children pass. You can, of course, reverse this effect by choosing items that older people do well on and younger people don’t.
Further, they keep 50 percent of items that children get right while keeping a smaller proportion of items that children get right, which, in effect, presupposes who is or is not intelligent.
Though, you never see those who believe that IQ is a ‘good enough’ proxy for intelligence ever being this up. Why? This is very important for the validity of these tests. Because if how the tests are constructed is wrong and test scores are not to fit a normal distribution when no normal distribution actually exists for most human mental (including IQ scores) and physiological traits, then the assumptions and conclusions drawn from them are wrong. IQ tests are constructed with the prior idea of who is or is not ‘intelligent’ and this is done by how the items are chosen—50 percent of the items that people get right are kept while the smaller proportion of items people get right or wrong are kept. This is how this so-called ‘normal curve’ appears in IQ tests and is why the book The Bell Curve has the name it has. But bell curve don’t exist for a modicum of traits including IQ!!
Simon (1997: 204) writes (emphasis mine):
There is another, and completely irrefutable, reason why the bell-shaped curve proves nothing at all in the context of H-M’s book: The makers of IQ tests consciously force the test into such a form that it produces this curve, for ease of statistical analysis. The first versions of such tests invariably produce odd-shaped distributions. The test-makers then subtract and add questions to find those that discriminate well between more-successful and less-successful test-takers. For this reason alone the bell-shaped IQ curve must be considered an artifact rather than a fact, and therefore tells us nothing about human nature or human society.
The analysis and selection of items that go on the tests are biased since there is no cognitive theory on which the analysis and selection of items are based. Carpenter, Just and Shell (1990: 408) note how John Raven, the creator of the Raven’s Progressive Matrices, even discussed this in his personal notes, writing “He used his intuition and clinical experience to rank order the difficulty of the six problem types. Many years later, normative data from Forbes (1964), shown in Figure 3, became the basis for selecting problems for retention in newer versions of the test and for arranging the problems in order of increasing difficulty, without regard to any underlying processing theory.”
II) IQ test validity
Another problem with IQ tests are its validity. People attempt to ‘prove’ its validity with correlating job performance success with IQ scores, though there are huge flaws in the studies purporting to show a .5 correlation between IQ and job performance (Richardson, 2002; Richardson and Norgate, 2015). IQ tests are not like, say, breathalyzers (which are calibrated against a model of blood alcohol) or white blood cell count (which is a proxy for disease in the body). Those two measures have a solid theoretical basis and underpinning; as blood alcohol rises, the individual had increased alcohol consumption. The same is true for white blood cell count. The same is not true for IQ tests.
One of the biggest measures used in regards to job performance and IQ testing (people attempt to use job performance to attempt to validate IQ tests) is supervisor rating. However, supervisory ratings are hugely subjective and a lot of factors that would have a supervisor be said to be a ‘good worker’ are not variables that entail just that job.
The only ‘validity’ that IQ test have is correlations with other IQ tests and tests like the SAT. This is not validity. Say the breathalyzer wasn’t calibrated against a model of blood alcohol in the body, would breathalyzers still be a valid tool to test people’s blood/alcohol level? On that same note let’s say that white blood cells wasn’t construct valid. Would we be able to reliably use white blood cell count as a valid measure for disease in the body? These very same problems plague IQ tests and people accept them as ‘proxies’ for intelligence, they test ‘enough of intelligence’ to be able to say that one is smarter than another because they scored higher in a test and therefore tap into this mystical ‘g’ that they have more of which is like a ‘power’ or ‘energy’.
These tests, therefore, are constructed with the idea of who is or is not intelligent and you can see that by looking at how the items are chosen for the test. That’s not scientific. So a true test of ‘intelligence’ may not even exist since these tests have this type of construct bias already in them.
IQ tests have no validity like breathalyzers and white blood cell count, and the so-called ‘culture-free’ IQ test Raven’s Progressive Matrices is anything but.
III) Raven’s and culture bias
I specifically asked Dr. James Thompson about Raven’s being culture-fair. I said that I recall Linda Gottfredson saying that people say that Ravens is culture-fair only because Jensen said it:
Yes, Gottfredson made that remark, and I remember her doing it at an ISIR conference.
So that’s one thing about Ravens that crumbles. A quote from Ken Richardson’s book Genes, Brains, and Human Potential: The Science and Ideology of Intelligence:
It is well known that families and subcultures vary in their exposure to, and usage of, the tools of literacy, numeracy, and associated ways of thinking. Children will vary in these because of accidents of background. …that background experience with specific cultural tools like literacy and numeracy is reflected in changes in brain networks. This explains the importance of social class context to cognitive demands, but is says nothing about individual potential.
(This argument on social class is much more complex than ‘poor people are genetically predisposed to be dumb and poor’.
Consider a recent GCTA study by Plomin et al., who reported a SNP-based heritability estimate of 35% for “general cognitive ability” among UK 12 year olds (as compared to a twin heritability estimate of 46%) [8]. According to the Wellcome Trust “genetic map of Britain,” striking patterns of genetic clustering (i.e. population stratification) exist within different geographic regions of the UK, including distinct genetic clusterings comprised of the residents of the South, South-East and Midlands of England; Cumbria, Northumberland and the Scottish borders; Lancashire and Yorkshire; Cornwall; Devon; South Wales; the Welsh borders; Anglesey in North Wales; Scotland and Ireland; and the Orkney Islands [8]. Now consider the title of a study from the University and College Union: “Location, Location, Location – the widening education gap in Britain and how where you live determines your chances” [9]. This state of affairs (not at all unique to the UK), combined with widespread geographic population stratification, is fertile ground for spurious heritability estimates.
Still Chasing Ghosts: A New Genetic Methodology Will Not Find the “Missing Heritability”
I think this argument is interesting, and it throws a wrench into a lot of things, but more on that another day.)
Richardson continues:
In other words, items like those in the Raven contain hidden structure which makes them more, not less, culturally steeped than any other kind of intelligence testing items, like the Raven, as somehow not knowledge-based, when all are clearly learning dependent. Ironically, such cultural-dependency testing is sometimes tacitly admitted by test users. For example, when testing children in Kuwait on the Raven in 2006, Ahmed Abdel-Khalek and John Raven transposed the items “to read from left to right following the custom of Arabic writings.” (Richardson, 2017: 99)
Finally, we have this dissertation which shows that urban peoples score better than hunter-gatherers (relevant to this present article):
Reading was the greatest predictor of performance Raven’s, despite controlling for age and sex. Attendance was also strongly correlated with Raven’s performance. These findings suggest that reading, or pattern recognition, could be fundamentally affecting the way an individual problem solves or learns to learn, and is somehow tapping into ‘g’. Presumably the only way to learn to read is through schooling. It is, therefore, essential that children are exposed to formal education, have the motivation to go/stay in school, and are exposed to consistent, quality training in order to develop the skills associated with improved performance. (pg. 83)
This is telling: This means that there is no such thing as a ‘culture-free’ IQ test and there will always be something involved that makes it culture un-fair.
People may say ‘It’s only rotating pictures and shapes to get the final answer, how much schooling could you need??’, well as seen above with the Tsimane, schooling is very important to IQ tests since they test learned skills. I’ve seen some people claim that IQ tests don’t test learned ability and that it’s all native, unlearned ability. That’s a very incorrect statement.
So although the symbols in a test like the RPM are experience-free, the rules governing their changes across the matrix are certainly not, and they are more likely to be already represented in the minds of children from middle-class homes, less so in others. Performance on the Raven’s test, in other words, is a question not of inducing ‘rules’ from meaningless symbols, in a totally abstract fashion, but of recruiting ones that are already rooted in the activites of some cultures rather than others. Like so many problems in life, including fields as diverse as chess, science and mathematics (e.g. Chi & Glaser, 1985), each item on the Raven’s test is a recognition problem (matching the covariation structure in a stimulus array to ones in background knowledge) before it is a reasoning problem. The latter is rendered easy when the former has been achieved. Similar arguments can be made about other so-called ‘culture-free’ items like analogies and classifications (Richardson & Webster, 1996). (Richardson, 2002: pg 292-292)
Everyday life is also more complex than the hardest items on Raven’s Matrices, while the test is not complex in its demands compared to tasks undertaken in everyday life (Carpenter, Just, and Shell, 1990). They conclude that the cause is differences in working memory, but that is an ill-defined concept in psychology. They do say, though, that “The processes that distinguish among individuals are primarily the ability to induce abstract relations and the ability to dynamically manage a large set of problem-solving goals in working memory.” So item complexity doesn’t make Raven’s items more difficult for others, since everyday life is more complex.
I’ll end with a bit of physiology. What physiological process is does IQ mimic in the body? If it is a physiological process, surely you’re aware that physiological processes *are not* static. IQ is said to be stable at adulthood, what a strange physiological process. Let’s say for arguments’ sake that IQ really does test some intrinsic, biological process. Does it seem weird to you that a supposed real, stable, biological, bodily function of an individual would be different at different times?
Conclusion
There are a lot of assumptions about IQ tests that are never talked about. The most important being how the tests are constructed to fit a normal curve when most traits important for survival aren’t normally distributed. IQ tests are constructed with the assumption of who is or isn’t intelligent just on the knowledge of how the items are prepared for the test. When you look at how the tests are constructed you can see how they are constructed to fit the normal curve because most of their assumptions and conclusions rest on the reality of the normal curve. There is no construct validity to IQ tests, they’re not like breathalyzers for instance which are calibrated against a model of blood alcohol or white blood cell count as a proxy for disease in the body. Raven’s—despite what is commonly stated about the test—is not unbiased, it perhaps is the most biased IQ test of them all. This highlights the problems with IQ tests that are rarely ever spoken about, and should have you call into question the ‘power’ of the IQ test which assumes who is or isn’t intelligent ahead of time.
My Response to (Ir)RationalWiki
1100 words
I was alerted to an article on the website (Ir)”RationalWiki” which in their own words “critique[s] and challenge[s] pseudoscience and the anti-science movement, explore[s] authoritarianism and fundamentalism, and analyze[s] how these subjects are handled in the media.” Unfortunately, it seems like the one who wrote this article (and is still adding to it) just selectively read certain articles and quote mined them.
The article on this website about me is an unfair mischaracterization of my views. Quotes will follow from the article with my comments.
In the opening paragraph they write:
NotPoliticallyCorrect is an Alt-right blog that promotes racialist pseudoscience and white nationalism; the owner posts as RaceRealist using the euphemism “racial realist” coined by the white supremacist J. P. Rushton who is extensively quoted on the blog.
- I’m not alt-right nor am I a white nationalist.
- I don’t promote ‘racialist pseudoscience’ nor do I promote ‘white nationalism’.
- Correct, Rushton did coin the term ‘race realist’, but he was not a ‘white supremacist’.
They continue, quoting an article of mine that I wrote almost two years ago titled Non-Western People are Abnormal to Our Society. I still stand by everything that I wrote in that article.
They continue:
A racist crank obsessed with controversial topics such as race and IQ and eugenics, RaceRealist argues in a 2016 blog essay “Non-Western People are Abnormal to Our [Western] Societies”[1] and its comments[2] that “MENA” and “SSA’s” (i.e. people from the Middle-East, North Africa and Sub-Saharan Africa) as well as other non-Westerners are somehow abnormal to the US and Europe:
They then quote me:
MENA and SSA people are abnormal to Western societies. It’s clear that, on average, full-on acclimation is not possible.
One only needs to look at what is occurring in Western European countries to see that, on average, this is true.
They continue:
In the same essay, RaceRealist goes on to post crude racism, such as “Negros” are biologically inferior:
Quoting me writing:
The same can be said for Negros[sic] in America as well. They are deviant, dysfunctional, they cause distress in our country and finally, they pose a danger to us, our families and societies as a whole. Just like those immigrants we have come into our countries who cannot assimilate because it’s not in their biology.
Except everything I wrote here was logically sound (last sentence notwithstanding). Look at the 4 d’s of abnormal psychology (which is the next quote they provide):
The “4 d’s of abnormality” and how they relate to our culture and the current culture/biology of those non-Western immigrants coming into our country is extremely telling. It’s clear that those people cannot assimilate into our societies because of differing biology and differing locations in which they evolved in. We chose our environments based on our biology. Environment increasingly depends on their genes, rather than being the cause of their exogenous behavior.
The 4 d’s of abnormality are deviance, dysfunction, distress and danger. Everything I wrote and then provided examples for in regards to the 4 d’s of abnormality are sound.
You can read my article Diversity in the Social Context for more evidence for this argument.
They then quote my article The Evolution of Jewish Nepotism writing:
RaceRealist is an anti-Semite who dislikes Ashkenazi Jews, accusing them of “derogating other ethnicities”; when discussing Ashkenazi Jews, he bizarrely maintains their higher average IQ is partly a product of “breeding with beautiful Roman women a few thousand years ago”,[3] for which there exists no evidence.
I admit it is conjecture. Evidence exists for Jewish men migrating to Rome to mate with Roman women (Atzmon et al, 2010). I never stated that I ‘dislike Ashkenazi Jews’. In regards to the derogation, it’s true. Close-knit ethnic groups derogate the out-group (Sampasivam et al, 2016). Further, oxytocin promotes human ethnocentrism, which caused in-group favoritism and out-group derogation (Drew et al, 2010). In-groups derogate out-groups. Read the literature.
And the final thing the page shows is my tweet saying that “I finally made it on (Ir)”RationalWiki””:
to which they wrote:
Twitter contributor Race Realist Eighty frickin’ Eight wishes to make it absolutely clear to everyone that he does not in fact consider himself “altright” and certainly not a “white nationalist”.[4]
Just because I have the numbers “88” in my handle doesn’t make me “alt-right” nor does it make me a “white nationalist.” I thought about changing it, then I realized that it’s good to weed out the people who aren’t serious about discussion and just look for things to discredit people that are meaningless to the conversation at hand. It tells you a lot about someone when they bring up irrelevant things. I’m not a white nationalist, nor am I an alt-righter. Just because I write about politics rarely and use them as an example (like in my article The Rise of Ethnocentrism and the Alt-Right: The Rebirth of Selfish Genes which I also disavow now that I realize that ‘selfish genes’ are a metaphor; Noble, 2011; Noble, 2013; Noble et al, 2014).
Take a look at the tags it tagged the article with: “Alt-righters, Pseudoscience, Racists, Internet kooks, Psuedoscience promoters, Alt-right, Internet Hate Sites.” Not an alrighter, I don’t push psuedoscience, I’m not a ‘racist’ (whatever that means). If you don’t like what I write, respond to any article you disagree with and explain why with logical, rational arguments. This piece is garbage and mischaracterizes my views using selective quotations (which, even then, failed to prove their point. No, numbers after a username are not evidence).
All in all, this article is garbage. It says that Rushton is ‘extensively quoted’, which is true for what I wrote in the beginning of this blog’s history, but not so for the past, say, 18 months. Rushton has been the target of my attacks on penis size, testosterone, and my personal favorite, r/K selection theory. But sure, go and dig in the archives for old articles to quote mine. This article written about me is dumb, doesn’t characterize my views correctly (calls me a ‘white nationalist’ and ‘alt-righter’). Selectively quote certain articles, assert that Rushton is ‘extensively quoted’ when I hardly discuss him anymore and when I do it’s about testosterone/to rebut him. (Ir)RationalWiki should think about reading a bit of my blog before characterizing me as something I’m not.
For the record, I don’t care about politics. I am not alt-right. I am not a white nationalist. I’m not an anti-semite. This will be updated to cover whatever else they decide to write about me. Hopefully it’s at least a bit closer to reality next time, because this article sucks.
Black-White Differences in Physiology
2050 words
Black-white differences in physiology can tell a lot about how the two groups have evolved over time. On traits like resting metabolic rate (RMR), basal metabolic rate (BMR), adiposity, heart rate, Vo2 max, etc. These differences in physiological variables between groups, then, explain part of the reason why there are different outcomes in terms of life quality/mortality between the two groups.
Right away, by looking at the average black and average white, you can see that there are differences in somatype. So if there are differences in somatype, then there must be differences in physiological variables, and so, this may be a part of the cause of, say, differing obesity rates between black and white women (Albu et al, 1997) and even PCOS (Wang and Alvero, 2013).
Resting metabolic rate
Resting metabolic rate is your body’s metabolism at rest, and is the largest component of the daily energy budget in modern human societies (Speakman and Selman, 2003). So if two groups, on average, differ in RMR, then one with the lower RMR may have a higher risk of obesity than the group with the higher RMR. And this is what we see.
Black women do, without a shadow of a doubt, have a lower BMR, lower PAEE (physical activity energy expenditure) and TDEE (total daily expenditure) (Gannon, DiPietro, and Poehlman, 2000). Knowing this, then it is not surprising to learn that black women are also the most obese demographic in the United States. This could partly explain why black women have such a hard time losing weight. Metabolic differences between ethnic groups in America—despite living in similar environments—show that a genetic component is responsible for this.
There are even predictors of obesity in post-menopausal black and white women (Nicklas et al, 1999). They controlled for age, body weight and body composition (variables that would influence the results—no one tell me that “They shouldn’t have controlled for those because it’s a racial confound!”) and found that despite having a similar waist-to-hip ratio (WHR) and subcutaneous fat area, black women had lower visceral fat than white women, while fasting glucose, insulin levels, and resting blood pressure did not differ between the groups. White women also had a higher Vo2 max, which remained when lean mass was controlled for. White women could also oxidize fat at a higher rate than black women (15.4 g/day, which is 17% higher than black women). When this is expressed as percent of total kcal burned in a resting state, white women burned more fat than black women (50% vs 43%). I will cover the cause for this later in the article (one physiologic variable is a large cause of these differences).
We even see this in black American men with more African ancestry—they’re less likely to be obese (Klimentidis et al 2016). This, too, goes back to metabolic rate. Black American men have lower levels of body fat than white men (Vickery et al, 1988; Wagner and Heyward, 2000). All in all, there are specific genetic variants and physiologic effects, which cause West African men to have lower central (abdominal) adiposity than European men and black women who live in the same environment as black men—implying that genetic and physiologic differences between the sexes are the cause for this disparity. Whatever the case may be, it’s interesting and more studies need to be taken out so we can see how whatever gene variants are *identified* as protecting against central adiposity work in concert with the system to produce the protective effect. Black American men have lower body fat, therefore they would have, in theory, a higher metabolic rate and be less likely to be obese—while black women have the reverse compared to white women—a lower metabolic rate.
Skeletal muscle fiber
Skeletal muscle fibers are the how and why of black domination in explosive sports. This is something I’ve covered in depth. Type II fibers contract faster than type I. This has important implications for certain diseases that black men are more susceptible to. Though the continuous contraction of the fibers during physical activity leads to a higher disease susceptibility in black men—but not white men (Tanner et al, 2001). If you’re aware of fiber type differences between the races (Ama et al, 1986; Entine, 2000; Caeser and Henry, 2015); though see Kerr (2010’s) article The Myth of Racial Superiority in Sports for another view. That will be covered here in the future.
Nevertheless, fiber typing explains racial differences in sports, with somatype being another important variable in explaining racial disparities in sports. Two main variables that work in concert are the somatype (pretty much body measurements, length) and the fiber type. This explains why blacks dominate baseball and football; this explains why ‘white men can’t jump and black men can’t swim’. Physiological variables—not only ‘motivation’ or whatever else people who deny these innate differences say—largely explain why there are huge disparities in these sports. Physiology is important to our understanding of how and why certain groups dominate certain sports.
This is further compounded by differing African ethnies excelling in different running sports depending on where their ancestors evolved. Kenyans have an abundance of type I fibers whereas West Africans have an abundance of type II fibers. (Genetically speaking, ‘Jamaicans’ don’t exist; genetic testing shows them to come from a few different West African countries.) Lower body symmetry—knees and ankles—show that they’re more symmetrical than age-matched controls (Trivers et al, 2014). This also goes to show that you can’t teach speed (Lombardo and Deander, 2014). Though, of course, training and the will to want to do your best matter as well—you just cannot excel in these competitions without first and foremost having the right physiologic and genetic make-up.
Further, although it’s only one gene variant, ACTN3 and ACE explain a substantial percentage of sprint time variance, which could be the difference between breaking a world record and making a final (Papadimitriou et al, 2016). So, clearly, certain genetic variants matter more than others—and the two best studied are ACTN3 and ACE. Some authors, though, may deny the contribution of ACTN3 to elite athletic performance—like one researcher who has written numerous papers on ACTN3, Daniel MacArthur. However, elite sprinters are more likely to carry the RR ACTN3 genotype compared to the XX ACTN3 genotype, and the RR ACTN3 genotype—when combined with type II fibers and morphology—lead to increased athletic performance (Broos et al, 2016). It’s also worth noting that 2 percent of Jamaicans carry the XX ACTN3 genotype (Scott et al, 2010), so this is another well-studied variable that lends to superior running performance in Jamaicans.
In regards to Kenyans, of course when you are talking about genetic reasons for performance, some people don’t like it. Some may say that certain countries dominate in X, and that for instance, North Africa is starting to churn out elite athletes, should we begin looking for genetic advantages that they possess (Hamilton, 2000)? Though people like Hamilton are a minority view in this field, I have read a few papers that there is no evidence that Kenyans possess a pulmonary system that infers a physiologic advantage over whites (Larsen and Sheel, 2015).
People like these three authors, however, are in the minority here and there is a robust amount of research that attests to East African running dominance being genetic/physiologic in nature—though you can’t discredit SES and other motivating variables (Tucker, Onywera, and Santos-Concejero, 2015). Of course, a complex interaction between SES, genes, and environment are the cause of the success of the Kalenjin people of Kenya, because they live and train in such high altitudes (Larsen, 2003), though the venerable Bengt Saltin states that the higher Vo2 max in Kenyan boys is due to higher physical activity during childhood (Saltin et al, 1995).
Blood pressure
The last variable I will focus on (I will cover more in the future) is blood pressure. It’s well known that blacks have higher blood pressure than whites—with black women having a higher BP than all groups—which then leads to other health implications. Some reasons for the cause are high sodium intake in blacks (Jones and Hall, 2006); salt (Lackland, 2014; blacks had a similar sensitivity than whites, but had a higher blood pressure increase); while race and ethnicity was a single independent predictor of hypertension (Holmes et al, 2013). Put simply, when it comes to BP, ethnicity matters (Lane and Lip, 2001).
While genetic factors are important in showing how and why certain ethnies have higher BP than others, social factors are arguably more important (Williams, 1992). He cites stress, socioecologic stress, social support, coping patterns, health behavior, sodium, calcium, and potassium consumption, alcohol consumption, and obesity. SES factors, of course, lead to higher rates of obesity (Sobal and Stunkard, 1989; Franklin et al, 2015). So, of course, environmental/social factors have an effect on BP—no matter if the discrimination or whatnot is imagined by the one who is supposedly discriminated against, this still causes physiologic changes in the body which then lead to higher rates of BP in certain populations.
Poverty does affect a whole slew of variables, but what I’m worried about here is its effect on blood pressure. People who are in poverty can only afford certain foods, which would then cause certain physiologic variables to increase, exacerbating the problem (Gupta, de Wit, and McKeown, 2007). Whereas diets high in protein predicted lower BP in adults (Beundia et al, 2015). So this is good evidence that the diets of blacks in America do increase BP, since they eat high amounts of salt, low protein and high carb diets.
Still, others argue that differences in BP between blacks and whites may not be explained by ancestry, but by differences in education, rather than genetic factors (Non, Gravlee, and Mulligan, 2012). Their study suggests that educating black Americans on the dangers and preventative measures of high BP will reduce BP disparities between the races. This is in-line with Williams (1992) in that the social environment is the cause for the higher rates of BP. One hypothesis explored to explain why this effect with education was greater in blacks than whites was that BP-related factors, such as stress, poverty and racial discrimination (remember, even if no racial discrimination occurs, any so-called discrimination is in the eye of the beholder so that will contribute to a rise in physiologic variables) and maybe social isolation may be causes for this phenomenon. Future studies also must show how higher education causes lower BP, or if it only serves as other markers for the social environment. Nevertheless, this is an important study in our understanding of how and why the races differ in BP and it will go far to increase our understanding of this malady.
Conclusion
This is not an exhaustive list—I could continue writing about other variables—but these three are some of the most important as they are a cause for higher mortality rates in America. Understanding the hows and whys of these variables will have us better equipped to help those who suffer from diseases brought on by these differences in physiological factors.
The cause for some of these physiologic differences come down to evolution, but still others may come down to the immediate obesogenic environment (Lake and Townshend, 2006) which is compounded by lower SES. Since high carbs diets increase BP, this explains part of the reason why blacks have higher BP, along with social and genetic factors. Muscle fiber typing is set by the second trimester, and no change is seen after age 6 (Bell, 1980). Resting metabolic rate gap differences between black and white women can be closed, but not completely, if black women were to engage in exercise that use their higher amounts of type II muscle fibers (Tanner et al, 2001). This research is important to understand differences in racial mortality; because when we understand them then we can begin to theorize on how and why we see these disparities.
Physiologic differences between the races are interesting, they’re easily measurable and they explain both disparities in sports and mortality by different diseases. Once we study these variables more, we will be better able to help people with these variables—race be dammed. Race is a predictor here, only because race is correlated with other variables that lead to negative health outcomes. So once we understand how and why these differences occur, then we can help others with similar problems—no matter their race.
Differing Race Concepts and the Existence of Race: Biologically Scientific Definitions of Race
2700 words
Do you need to look at genetic differences between races to see if race is real? Some may argue that you do, and when you do you’ll see that genetic variation is too small to say that race exists. However, other arguments exist that do not look at genetic differences between races, but look at geographic ancestry, reproductive isolation between races, and morphologic differences. Those three variables are enough to prove the existence of race without looking at genetic differences between races. They do correspond to genetic differences between races. The four concepts I will briefly lay out are from Michael Hardimon, professor of philosophy at University of California, San Diego. The concepts are the racialist concept of race, minimalist concept of race concept, populationist concept of race, and the socialrace concept of race. One doesn’t need to look at the racialist concept of race to prove the existence of race, which I will prove below.
Michael Hardimon published Rethinking Race: The Case for Deflationary Realism earlier this year. In the book, he makes the case that race exists if minimalist race exists (I will get into what minimalist race entails below). Nevertheless, race deniers will say that even by looking at variables such as morphology, reproductive isolation, and geographic ancestry, race as a concept is scientifically invalid. This is patently false.
Concepts of race
The racialist concept of race
Hardimon’s first race concept is the racialist concept. The racialist concept (keep in mind, this is, as Hardimon writes on page 17 of his book Rethinking Race “the specific concept I have dubbed “the racialist concept” which “is hierarchal“) as defined by Hardimon holds that “racialist race is the idea of a fundamental division between groups and individuals” (Hardimon, 2017: 17). I think that Hardimon strawmans the racialist concept as he as defined it, but that’s for another day.
He also says that the racialist concept “is closely associated with racism” while the terms racialism and racism are “sometimes used interchangeably” (Hardimon, 2017: 17).
His argument against the racialist concept of race (as he defines it) is as follows (Hardimon, 2017: 21):
A third line of argument starts from the idea that in order for racialist races to exist, certain things must be true of human genetics, namely the following:
(a) The fraction of human genetic diversity between populations must exceed the fraction of diversity between them.
(b) The fraction of human genetic diversity within populations must be small.
(c) The fraction of diversity between populations must be large.
(d) Most genes must be highly differentiated by race.
(e) The variation in genes that underlie obvious physical differences must be typical of the genome in general.
(f) There must be several important genetic differences between races apart from the genetic differences that underlie obvious physical differences.
Note: (b) says that racialist races are genetically racially homogeneous groups; (c)-(f) say that racialist races are distinguised by major biological differences.
Call (a)-(f) the racialist concept of race’s genetic profile.
Now that his argument against the racialist concept (as he defines it) is laid out, you can see why I said that I think he strawmans the racialist concept. But I’ll get into that another day.
He then cites Lewontin’s (1972) analysis of blood groups by race as evidence against the racialist concept. Lewontin found that 85.4 percent of total human variation fell within populations. He also found that populations that populations classically defined as human races (Caucasians, Africans, Mongoloids, South Asian Aborigines, American Indians, and Oceanians) accounted for 8.3 percent of total human variation. Total variation between the classically defined races accounted for 6.3 percent of the variance.
It’s worth noting that the numbers given by Lewontin are true; where he goes wrong is assuming that there is no taxonomic significance for race based on the data he got from his analysis. “Call this Lewontin’s cleaver,” writes Hardimon on page 22.
Then in 2002, 31 years after Lewontin published his analysis, A.W.F. Edwards published his paper Human Genetic Diversity: Lewontin’s Fallacy. (Edwards, 2003). In the paper, Edwards argues that Lewontin’s conclusion is incorrect. Edwards (2003: 800-801) writes in his conclusion (emphasis mine):
There is nothing wrong with Lewontin’s statistical analysis of variation, only with the belief that it is relevant to classification. It is not true that ‘‘racial classification is … of virtually no genetic or taxonomic significance’’. It is not true, as Nature claimed, that ‘‘two random individuals from any one group are almost as different as any two random individuals from the entire world’’, and it is not true, as the New Scientist claimed, that ‘‘two individuals are different because they are individuals, not because they belong to different races’’ and that ‘‘you can’t predict someone’s race by their genes’’. Such statements might only be true if all the characters studied were independent, which they are not.
Of course, Lewontin’s conclusion is fallacious because small genetic differences do not entail that racial classification that race has no taxonomic significance (Richard Dawkins accepts the taxonomic existence of race). As you can see from the quote from Edwards, he does not object to Lewontin’s analysis of the races, he objects to his conclusion—namely that races do not exist based on the within-race variation being greater than between-race variation.
On page 22-23, Hardimon writes about Edwards’ objection to Lewontin’s conclusion:
Lewontin’s locus-by-locus analysis (which does not consider the possibility of a correlation between individual loci) does not preclude the possibility that individual loci might be correlated in such a way that people could be grouped into traditional racial categories. The underlying thought is that racial classification would have “taxonomic significance” were it possible to group people into traditional racial categories by making use of correlations between individual loci. However, Lewontin’s argument that there are no racialist races because the component of within-race genetic variation is larger than the component of between-race variation is untouched by Edwards’s objection.
In 2002, Rosenberg et al, in their paper Genetic Structure of Human Populations confirmed Lewontin’s analysis. They looked at 377 autosomal loci in 1,056 individuals from 52 populations and found that within-population differences between major groups (Africa, Europe, Asia, the Middle East, Central and South Asia, East Asia, Oceania, and America) accounted for 3-5 percent of genetic variation while genetic differences between individuals accounted for 93-95 percent of genetic variation. So Rosenberg et al (2002) confirmed Lewontin’s (1972) analysis—though do recall that Lewontin’s conclusion is incorrect. According to Hardimon’s interpretation of the racialist concept of race, both Lewontin’s and Rosenberg et al’s analysis disprove the racialist concept of race, but that doesn’t mean that there is no scientific basis for the biological reality of race (Hardimon, 2012).
The minimalist concept of race
The minimalist concept of race is similar to the racialist concept, though there are some stark differences. It does not say that there are intrinsic differences between races—call them essences if you will), but it does say that you can distinguish races by patterns of different physical features such as skin color, hair type, nose shape, morphology, etc, which then correspond to differences in geographic ancestry in geographically, genetically isolated breeding populations.
The minimalist concept of race further states that (i) races are distinguised from other races by patterns of visible physical features; (ii) the members are linked by a common ancestry which is peculiar to members of the group; and (iii) this group must originate from a distinct location.
The minimalist concept of race does not require: that the fraction of human genetic diversity between minimalist races is larger than the fraction of diversity within them; it is compatible with within-race diversity being large and between-race diversity being small; it does not require most genes to be highly differentiated by race; it does not require the existence of a lot genetic differences between races that underlie more than the phenotypic differences already noticed; the concept does not imply that there can be predictions made from yet unstudied characteristics; it finally does not require any genetic differences between races other than those found in the genes that underlie differences in physical appearance between race. This is called the minimalist concept of biological race (Hardimon, 2017: 66) and it survives all objections from Lewontin’s and Rosenberg et al’s analysis of between-race genetic variation.
This is my favorite race concept, personally, because it covers any and all objections from the race-denialist crowd—people who deny any genetic differences between races—because the only genetic differences it counts on are those physical traits that are already noticed.
Hardimon (2017: 29) writes:
Such readers should feel free to regard the minimalist concept of race, that is, as a concept that, though in many respects similar to the ordinary concept, is nonetheless distinct from it. What I would insist on is that minimalist races (groups satisfying the minimalist concept of race) are *races* (that is races so properly called)—either because the minimalist concept of race just is the ordinary concept of race or because it captures enough of the ordinary concept of race for minimalist races to be counted as races. My view is that if it can be shown that minimalist races exist, races exist. And if it can be shown that *minimalist race* is real, race is real.
The populatonist concept of race
The populationist concept of race is a nonessentialist, non-hierarchical concept of race that slightly differs from the minimalist concept of race. The populationist concept of race can be said to be a scientific concept of race (as can the minimalist concept) because it characterizes races as groups belonging to different groups of biological descent, they are distinguished by patterns of phenotypic differences, and these phenotypic differences trace back to geographically separated and genetically isolated founding populations.
The populationist concept of race also holds that “A race is a subdivision of Homo sapiens—a group or population that exhibits a distinctive pattern of genetically transmitted phenotypic characters that corresponds to the group’s geographical ancestry and belongs to a biological line of descent initiated by a geographically separated and reproductively isolated founding population” (Hardimon, 2017: 99). So with these criteria, you can see that even if you do not accept the racialist concept of race (as Hardimon defines it), you can still be a race realist. The populationist concept is likely to exist, and if the populationist concept of race exists then race is real.
Defining race as geographically and reproductively isolated breeding populations that share a common line of biological descent with similar phenotypic characters is as barebones a concept of race as you can get—and it is perfectly in line with how most people view races on the basis of phenotypic characterization. The populationist concept of race supposes that numerous concepts from the racialist concept of race are true—but do not presuppose any to-be-studied differences between those races. The strength of the populationist argument, as you can see, is very strong and it holds up to numerous lines of criticism very well. Although both the populationist and minimalist race concepts do not presupposed any to-be-studied differences between races, this still is not good enough for race deniers.
It is clear that without even looking at the brain and physiological differences between races, that race does indeed exist and it does—contrary to popular belief—have implications for people’s health of certain races.
The socialrace concept of race
Finally, the last concept of race laid out by Hardimon is the concept of socialrace. The concept of socialrace takes a race to be a racialist race, it refers to a position that is occupied by a social group that is a socialrace, and the socialrace concept refers to the system of social positions that are socialraces. This concept of race is, clearly, different from the minimalist and populationist race concepts but does indeed correlate with popular notions of race (and would correlate with the minimalist and populationist concept of race very well). The socialrace concept is, basically, what is believed to be racialist races.
The concept of socialrace is a concept of race as a social group (Hardimon, The Ontology of Race: 31)
The socialrace concept differs from the minimalist and populationist concept of race in that it looks at so-called social—not biological—correlates of race. Though, still, the socialrace concept can be said to show the reality of race since how one socially defines themselves correlates almost perfectly with geographic ancestry (which is a prerequisite for the existence of the minimalist concept of race and the populationist concept of race) (Tang et al, 2005). They showed that self-identified racial categories lined up almost perfectly with geographic ancestry (99.86 percent of the time). So, as you can see, the concept of socialrace also gives credence to the existence of the minimalist and populationist concepts of race.
This concept of race—as its name implies—does not talk race is a biological manner, but a social one, as its name implies. However, due to the extremely high chance that one’s self-identified race (their socialrace) lines up with the geographic ancestry of the classical races, we can see that the socialrace concept further buttresses the argument for the existence for the reality of the minimalist concept of race and the populationist concept of race.
The socialrace concept is kind of like Templeton (2014) defines race: that human races exist in a cultural sense, but not biologic sense. I have shown, though, that races exist in a cultural, social, and biological sense with the arguments presented in this article. Socialrace, culturalrace, whatever you want to call it, it is evidence for the existence of race.
Conclusion
Race exists whether or not the racialist position of race (as Hardimon defines it) is true or not. The minimalist concept of race and populationist concept of race show that race is real while the concept of socialrace further lends credence to the biological models of the minimalist and populationist concept of race. Even still, people who deny race because the genetic distance between races is too small for their to be any meaningful differences between them do not accept that three arguments above (sans the racialist concept) for the existence of race. They’ll still talk about the genetic differences between them and, say, morphology, but the minimalist concept of race and the populationist concept of race define race in enough of a way that genetic differences do not need to be looked at—we can only look at reproductive isolation, morphology, geographic ancestry and physical differences between minimalist and populationist races such as hair, nose, and skin color along with morphological differences.
Minimalist and populationist races exist and are a biological reality. We can take those two concepts to be a scientific basis for race. While we can take the concept of socialrace not as a biological concept, but as a social concept and we can then say that socialrace is socially real while being a significant social reality. That social reality is manifested by noticing different racial phenotypes, along with differences in SES, educational attainment, etc, and placing different races in different average social positions, which would correlate with the concepts of race mentioned above. This also correlated nearly perfectly with geographic ancestry. So, I’m saying it again, the existence of race as a social reality is real; the existence of socialrace buttresses the arguments for both the existence of the minimalist concept of race and the populationist concept of race—both of which are scientific concepts of race.
Minimalist races exist, and is a superficial biological reality, populations races may exist and if they exist, they are a relatively superficial biological reality. Socialraces exist and are a social reality which also lend credence to the minimalist and populationist concepts. I personally am privy to the minimalist race concept because it is shown to be real, so race is real.
In sum, race exists whether you look at genetic differences between races or not, morphology, geographic ancestry, reproductive and genetic isolation are all you need to prove the existence of race. There is a scientific concept of race, and the minimalist and populationist race concepts provide the existence for it, while the socialrace concept does as well. It is clear that for a scientific concept of race, you only need phenotypic variation, morphologic variation between races,
(Also read the American Rennaisance review for the book, A Tactical Retreat for Race Denial. I think it is balanced and fairly written, though a bit biased and doesn’t account for Hardimon’s views well enough in my opinion.)
Racial Differences in Physical Activity and Acquisition of Coronary Artery Calcification
1200 words
Last week a study was published stating that white men who exercised 3 times the recommendation of 1.5 hours (450 minutes, 7.5 hours) had a higher chance of getting coronary artery calcification (CAC), which is the accumulation of plaque and calcium in the arteries of the heart. You, of course see news headlines such as: “Physically active white men at high risk for plaque buildup in arteries“; “White Men Who Exercise Every Day Have 86 Per Cent Higher Risk of Heart Disease Than Black Men, Study Claims“; “Excessive Exercise May Harm The Heart, Study Suggests “; “Excessive exercise increases risk of arterial plaque buildup in white men“; (and my personal favorite headline about this study): “You can exercise yourself to death, says new study“. People just passing by and reading the title (like most do) may then conclude that “they’re saying not to exercise because of CAC.” No, this is not what they are saying at all.
The Coronary Artery Risk Development in Young Adults (CARDIA) study is one of the most important studies in the study of coronary heart disease that have been undertaken. It is a sample of men and women, about equal numbers of each race, from Birmingham, Alabama; Chicago, Illinois; Minneapolis, Minnesota; and Oakland, California. The study began in 1985-86 and there were follow-up examinations at “1987-1988 (Year 2), 1990-1991 (Year 5), 1992-1993 (Year 7), 1995-1996 (Year 10), 2000-2001 (Year 15), 2005-2006 (Year 20), 2010-2011 (Year 25), and 2015-2016 (Year 30).” The CARDIA website writes:
Data have also been collected on physical measurements such as weight and body composition as well as lifestyle factors such as dietary and exercise patterns, substance use (tobacco and alcohol), behavioral and psychological variables, medical and family history, and other chemistries (e.g., insulin).
So there is a goldmine of information to be gleaned from this data. The study that is getting press in the news uses data from this cohort.
The study
The study is titled 25-Year Physical Activity Trajectories and Development of Subclinical Coronary Artery Disease as Measured by Coronary Artery Calcium by Laddu et al (2017). They studied three cohorts by the amount of time they exercised per week: below requirement, at requirements, or above requirements. It is recommended to exercise at least 150 minutes per week.
There were 3,175 men and women who participated in the CARDIA study between 1985 and 2011 who had CAC data available for 25 years. About 47.4 percent of the sample was black, with 56.6 being women. The cohort “consisted of 18.9% black men, 24.6% white men, 28.6% black women, and 28.0% white women” (Laddu et al, 2017).
Of the three activity levels they studies (below 150 minutes, 150 minutes, and over 150 minutes), they observed that white men who exercised 3 times the weekly recommendation (150 minutes(3)= 450 minutes=7.5 hours) had a higher chance of developing CAC. It’s worth noting that exercise time was self-reported (which is the only way I can see how something like this would work, are you supposed to follow people with a camera every day to see how long they engage in physical activity?).
In regards to the physical activity measurement, Laddu et al (2017) write:
At each of the 8 examinations, self-reported leisure-time PA was ascertained by the interviewer-administered CARDIA Physical Activity History Questionnaire.17 Participants were asked about the frequency of participation in 13 specific categories (8 vigorous intensity and 5 moderate intensity) of recreational sports, exercise, home maintenance, and occupational activities during the previous 12 months. Intensity for each activity was expressed as metabolic equivalents (METs), in which 1 MET is defined as the energy expended at rest, which is approximately equivalent to an oxygen consumption of 3.5 mL per 1 kg of body weight per minute.18Vigorous activities (≥6 METs) included running or jogging; racquet sports; biking; swimming; exercise or dance class; job lifting, carrying, or digging; shoveling or lifting during leisure; and strenuous sports. Moderate-intensity activities (3-5 METs) included nonstrenuous sports, walking and hiking, golfing and bowling, home exercises or calisthenics, and home maintenance or gardening.19 Each activity was scored according to whether it was performed for 1 hour or longer during any 1 month during the past year, the number of months it was performed at that level, and the number of months the activity was performed frequently. Each activity was then assigned an intensity score, ranging from 3 to 8 METs, and a duration threshold (ranging from 2-5 hours per week), above which participation was considered to be frequent.20
This is a good metric; though I would like to see a study that looks at just gym-going activity and death, time spent in the gym strength training/moderate to intense cardio. Nevertheless, white men who reported more physical activity had a higher chance of acquiring CAC. Though I can see people’s recall being hazy, people over/under reporting, etc etc.
White men who exercised 7.5 hours per week were 27 percent more likely to get CAC, whereas blacks who exercised that much were at no greater risk to acquire CAC when compared to whites (7.5 hours of exercise compared to less than 2.5 hours per week). Black women who exercised less than the recommendations had a higher chance of acquiring CAC. The researchers couldn’t ascertain why white men who exercised three times the recommendations had such a higher chance of acquiring CAC by the time they reached middle age, but Dr. Jamal Rana says “however this plaque buildup may well be of the more stable kind, and thus less likely to rupture and causes heart attack, which was not evaluated in this study.” The head author, Dr. Deepika Laddu also reiterated: “it does not suggest that anyone should stop exercising.” So people who just read these click bait headlines who say “They’re telling whites not to exercise!”, you’re wrong and you should read papers and not news articles.
This is the perfect example of people reading click baity, fear-mongering headlines and running with it. I saw some people saying “They’re telling us not to exercise!” No. If you were to read the paper and any serious news articles on the matter, you’d see that they do not recommend that people do not exercise. Now the question is, why do whites who exercise more than 7.5 hours per week have a higher chance of acquiring heart disease? I can think of a few explanations (though they are not satisfactory): 1) genes: which genes? Why? How do they interact with the body over time to lead to arterial calcification?; 2) dietary habits: I’d like to know what their diet was like and see their macro composition, carbohydrates, not saturated fat, causes heart disease (Siri-Tirino et al, 2010; de Souza et al, 2015) so that may be a huge contributing factor.
Nevertheless, this is yet another physiological race difference. Oddly enough, black men are more likely than white men to have hypertension (Hicken et al, 2013).
Even though black men, on average, have higher rates of hypertension than white men, white men who are physically active for 7.5 had a higher chance of acquiring CAC than those who exercised less than 2.5 hours per week. This effect wasn’t seen in black men who had physical activity at that level, which, of course, implies that differences in genes and SES underlie this difference. I await more papers into this matter into the mechanisms of how and why this occurs and will ruminate on this myself in the future. No, this study does not tell white men not to exercise.
Evidence for Natural Selection in Humans: East Asians Have Higher Frequency of CASC5 Brain Size Regulating Gene
1500 words
Brain size is one physical difference that the races differ on. East Asians have bigger brains than Europeans who have bigger brains than Africans (Beals et al, 1984; Rushton, 1997). What caused these average differences and the ultimate causes for them have been subject to huge debate. Is it drift? Natural/sexual selection? Mutation? Gene flow? Epigenetic? One reason why brains would need to be large in colder climates is due to heat retention, while in tropical climates heads need to be smaller to dissipate heat. One of the biggest criticisms of HBD is that there is no/little evidence of recent natural selection between human races. Well, that has changed.
CASC5 “performs two crucial functions during mitosis, being required for correct attachment of chromosome centromeres to the microtubule apparatus, and also essential for spindle-assembly checkpoint (SAC) signaling” (Shi et al, 2016). The gene has been found to be important in recent human evolution along with neurogenesis.
Shi et al (2016) genotyped 278 Han Chinese (174 females and 104 males with a mean age of 36) who were free of maladies or genetic defects. They had the coding sequences of CASC5 for humans, chimpanzees, gorillas, baboons, gibbons, orangutans, tarsiers, Denisovans, and Neanderthals. They downloaded genotypes from the Human Genome Project for their analysis.
They compared CASC5 among three human species: humans, Neanderthals, and Denisovans. Using chimpanzees as an outgroup, they discovered 45 human-specific mutations, 48 Neanderthal-specific mutations, and 41 Neanderthal-specific mutations. Further, when one exon region was aligned among modern humans, non-human primates and other mammalian species, 12 amino acid sites showed divergence between modern humans, Neanderthals, and Denisovans with 8 occurring in modern humans. Of the 8 sites in humans, 6 are preserved which implies that they were important in our evolutionary history.
Shi et al (2016) write:
At the population level, among the 8 modern human amino acid changes, two (H159R and G1086S) are fixed in current human populations, and the other six are polymorphic Fig. 1). Surprisingly, 5 of the 6 amino acid polymorphic sites showed deep between-population divergence in allele frequencies. East Asians possess much higher frequencies of the derived alleles at four sites (T43R-rs7177192, A113T-rs12911738, S486A-rs2412541 and G936R-rs8040502) as compared to either Europeans or Africans (Fig. 1), while E1285K-rs17747633 is relatively enriched in Europeans (46%), and rare in East Asians (10%) and Africans (3%). No between-population divergence was observed for T598 M-rs11858113 (Fig. 1).
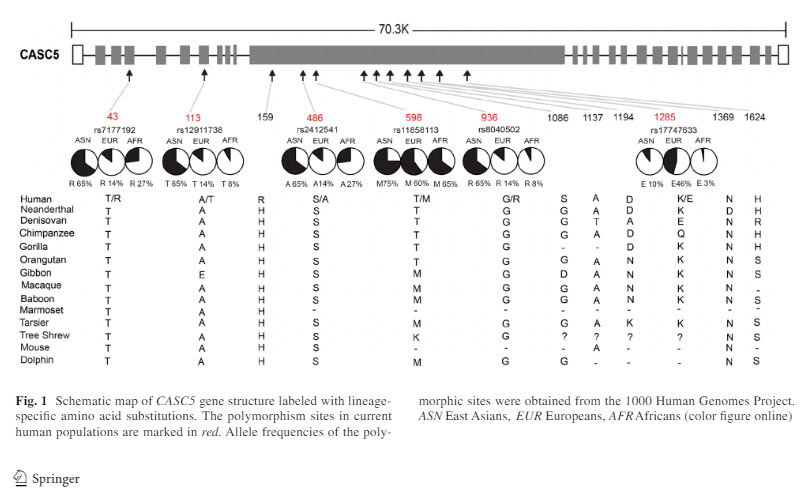
So East Asians have a much higher frequency of this derived trait. This is direct evidence for natural selection in recent human evolution in regards to the physical structure of the brain.
Since most of the amino acid polymorphic sites showed between-population divergence, they decided to analyze the three classical races using 1000 genomes. The variation between the races could be due to either genetic drift or natural selection. When they analyzed certain gene regions, they observed a signal of positive selection for East Asians but not Europeans or Africans. They further tested this selection signal using “the standardized integrated haplotype score (iHS) which is used for detecting recent positive selection with incomplete sweep (i.e. the selected allele is not yet fixed)” (Shi et al, 2016). Using this method, they discovered a few SNPs with large iHS values in Europeans (7 SNPs at 4.2 percent) and none in Africans.
They also conducted a genome-wide scan of Fst, iHS, and “XPCLR (searching for highly differentiated genomimc regions as targets of selective sweeps)” (Shi et al, 2016). Several SNPs had high Fst, iHS and XPCLR scores, which indicate that these alleles have been under positive selection in East Asians. Among the fixed amino acid sites in human populations, East Asians showed 5, Europeans showed 1, and Africans showed 0 which, the authors write, “[imply] that these amino acid changes may have functional effects” (Shi et al, 2016). Furthermore, using the HDGP, they obtained the frequency of the 6 amino acid sites in 53 populations. This analysis showed that 4 of the 6 amino acid sites are “regionally enriched in East Asia .. in line with the suggested signal of population-specific selection in this area” (Shi et al, 2016).
Then, since CASC5 is a brain size regulating gene, they looked for phenotypic effects. They recruited 167 Han Chinese (89 men, 178 women) who were free of maladies. They genotyped 11 SNPs and all of the frequencies followed Harvey-Weinberg Equilibrium (which states that allele and genotype frequencies will remain constant in a population from generation to generation in the absence of evolutionary pressures; Andrews, 2010). In the female sample, 5 regions were related to gray matter volume and four were on the amino acid polymorphic sites. Interestingly, the four alleles which showed such a stark difference between East Asians and Europeans and Africans showed more significant associations in Han Chinese females than males. Those carrying the derived alleles had larger brain volumes in comparison with those who had the ancestral alleles, implying recent natural selection in East Asia for brain size.
Shi et al (2015) also attempted two replications on this allele writing:
We further conducted a replication analysis of the five significant CAC5 SNPs in two other independent Han Chinese samples (Li et al. 2015; Xu et al. 2015). The results showed that three SNPs (rs 7177192, rs11858113 and rs8040502) remained significant in Replication-1 for total brain volume and gray matter volume (Xu et al. 2015), but no association was detected in Replication-2 (Li et al. 2015) (Table S4).
It is very plausible that the genes that have regulated brain growth in our species further aid differences in brain morphology within and between races. This effect is seen mostly in Han Chinese girls. Shi et al (2016) write in the Discussion:
If this finding is accurate and can be further verified, it suggests that that [sic] after modern humans migrated out of Africa less than 100,000 years ago, the brain size may still be subject to selection.
I do believe it is accurate. Of course, the brain size could still be subject to selection; there is no magic field shielding the brain against selection pressure. Evolution does not stop at the neck.
So Shi et al (2016) showed that there were brain genes under recent selection in East Asians. What could the cause be? There are a few:
- Climate: In colder climates you need a smaller body size and big brain to survive the cold to better thermoregulate. A smaller body means there is less surface area to cover, while a larger head retains heat. It, obviously, would have been advantageous for these populations to have large brains and thus get selected for them—whether by natural or sexual selection. This could also have to do with the fact that one needs bigger eyes in colder environments, which would cause an increase in the size of the brain for the larger eyes, as well as being sharper visio-spatially.
- Intelligence: East Asians in this study showed that they had high levels of gray matter in the skull. Further, large brains are favored by an intermediately challenging environment (Gonzalez-Forero, Faulwasser, and Lehmann, 2017).
- Expertise: I used Skoyle’s (1999) theory on expertise and human evolution and applied it to racial differences in brain size and relating it to the number of tools they had to use which differed based on climate. Now, of course, if one group uses more tools then, by effect, they would need more expertise with which to learn how to make those tools so large brains would be selected for expertise—especially in novel areas.
- Vision: Large brains mean large eyes, and people from cold climates have large eyes and large brains (Pearce and Dunbar, 2011). Decreasing light levels select for larger eye size and visual cortex size in order to “increase sensitivity and maintain acuity“. Large eyeballs mean enlarged visual cortices. Therefore, in low light, large brains and eyes get selected for so one can see better in a low light environment.
Of course, all four of the examples below could (and probably do) work in tandem. However, before jumping to conclusions I want to see more data on this and how the whole of the system interacts with these alleles and these amino acid polymorphic sites.
In sum, there is now evidence for natural selection on East Asians (and not Africans or Europeans) that favored large brains, particularly gray matter, in East Asians with considerable sexual dimorphism favoring women. Four of the genes tested (MCPH1, ASPM, CDK5RAP2, and WDR62) are regulated by estradiol and contribute to sexual dimorphism in human and non-human primates (Shi et al, 2016). Though it still needs to be tested if this holds true for CASC5.
This is some of the first evidence that I have come across for natural selection on genes that are implicated in brain evolution/structural development between and within populations. It does show the old “Rushton’s Rule of Three“, that is, Mongoloids on top, Caucasians in the middle, and Negroids on bottom, though Caucasians were significantly closer to Africans than Mongoloids in the frequency of these derived alleles. I can see a HBDer going “They must be related to IQ”, I doubt it. They don’t ‘have’ to be related to IQ. It just infers a survival advantage in low light, cold environments and therefore it gets selected for until it reaches a high frequency in that population due to its adaptive value—whether spreading by natural or sexual selection.
MAOA, Race, and Crime: A Simple Relationship?
2400 words
When I first got into HBD back in 2012, one of the first things I came across—along with the research on racial IQs from Rushton, Lynn, Jensen et al—was that the races differed in a gene called MAOA-L, which has a frequency in Caucasians at .1 percent (Beaver et al, 2013), 54 percent in Chinese people (Lu et al, 2013; 56 percent in Maoris (Lea and Chambers 2007) while about 60-65 percent of Japanese people have the low-frequency version of this gene (Way and Lieberman, 2007).
So if these ethnies have a higher rate of this polymorphism and it is true that this gene causes crime, then the Chinese and Japanese should have the highest rates of crime in the world, since even apparently the effect of MAOA and violence and antisocial behavior is seen even without child abuse (Ficks and Waldman, 2014). Except East Asian countries have lower rates of crime (Rushton, 1995; Rushton and Whytney, 2002). Though, Japan’s low crime rate is relatively recent, and when compared with other countries on certain measures “Japan fares the same or worse when compared to other nations” (Barberet 2009, 198). This goes against a lot of HBD theory, and I will save that for another day. (Japan has a 99 percent prosecution rate, which could be due to low prosecutorial budgets; Ramseyer and Rasmusen, 2001. I will cover this in the future.)
The media fervor—as usual—gave the MAOA gene the nickname “the warrior gene“, which is extremely simplistic (I will have much more to say on ‘genes for’ any trait towards the end of the article). I will show how this is a very simplistic view.
The MAOA gene was first discovered in 1993 in a Dutch family who had a history of extreme violence going as far back as the 1890s. Since the discovery of this gene, it has been invoked as an ultimate cause of crime. However, as some hereditarians do note, MAOA only ’causes’ violence if one has a specific MAOA genotype and if they have been abused as a child (Caspi et al, 2002; Cohen et al, 2006; Beaver et al, 2009; Ferguson et al, 2011; Cicchetti, Rogosch, Thibodeau, 2012;). People have invoked these gene variants as ultimate causes of crime—that is, people who have the low-expressing MAOA variants are more likely to commit more crime—but the relationship is not so simple.
Maoris are more four times more likely to have the low-expressing gene variant than Europeans, the same holding for African Americans and Europeans (Lea and Chambers, 2007).
There is, however, a protective effect that protects whites (and not non-whites in certain cases) against antisocial behavior/violent attitudes if one has a certain genotype (Widom and Brzustowicz, 2006), though the authors write on page 688: “For non-whites, the effect of child abuse and neglect on the juvenile VASB was not significant (beta .08, SE .11, t 1.19, ns), whereas the effect of child maltreatment on lifetime VASB composite approached significance (beta .13, SE .12, t 1.86, p .06). For non-whites (see Figure 2), neither gene (MAOA) environment (child abuse and neglect) interaction was significant: juvenile VASB (beta .06, SE .28, t .67, ns) and lifetime VASB (beta .01, SE .29, t .14, ns).” So as you can see, there are mixed results. Whites seem to be protected against the effect of antisocial behavior and violence but only if they have a certain genotype (which implies that if they have the other genotype, then if abused they will show violent and antisocial behavior). So, we can see that the relationship between MAOA and criminal behavior is not as simple as some would make it out to be.
MAOA, like other genetic variants, of course, has been linked to numerous other traits. Steven J. Heine, author of the book DNA is Not Destiny: The Remarkable and Completely Misunderstood Relationship Between You and Your Genes:
However, any labels like “the warrior gene” are highly problematic because they suggest that the this gene is specifically associated with violence. It’s not, just as alleles from other genes do not only have one outcome. Pleiotropy is the term for how a single genetic variant can influence multiple different phenotypes. MAOA is highly pleiotropic: the traits and conditions potientially connected to the MAOA gene invlude Alzheimer’s. anoerxia, autism, body mass index, bone mineral density, chronic fatigue syndrome, depression, extraversion, hypertension, individualism, insomnia, intelligence, memory, neuroticism, obesity, openness to experience, persistence, restless leg syndrome, schizophrenia, social phobia, sudden infant death syndrome, time perception and voting behavior. (59) Perhaps it would be more fitting to call MAOA “the everything but the kitchen sink gene. (Heine, 2017: 195)
Something that I have not seen brought up when discussions of race, crime, and MAOA come up is that Japanese people have the highest chance—even higher than blacks, Maoris, and whites—to have the low repeat MAOA variant (Way and Lieberman) yet have lower rates of crime. So MAOA cannot possibly be a ‘main cause’ of crime. It is way more complex than that. “However intuitively satisfying it may be to explain cultural differences in violence in terms of genes“, Heine writes, “as of yet there is no direct evidence for this” (Heine, 2017: 196).
Numerous people have used ‘their genes’ in an attempt to get out of criminal acts that they have committed. A judge even knocked off one year off of a murder’s sentence since he found the evidence for the MAOA gene’s link to violence “particularly compelling.” I find it “particularly ridiculous” that the man got less time in jail than someone who ‘had a choice’ in his actions to murder someone. Doesn’t it seem ridiculous to you that someone gets less time in jail than someone else, all because he may have the ‘crime/warrior gene’?
Aspinwall, Brown, and Tabery (2012) showed that when evidence of a ‘biomechanic’ cause of violence/psychopathy was shown to the judges (n=191), that they reduced their sentences by almost one year if they were reading a story in which the accused was found to have the low-repeat MAOA allele (13.93 to 12.83 years). So, as you can see, this can sway judges’ perception into giving one a lighter sentence since they believe that the evidence shows that one ‘can not control themselves’, which results in the judge giving assailants lighter sentences because ‘it’s in their genes’.
Further, people would be more lenient on sentences for criminals who are found to have these ‘criminal genes’ than those who were found to not have them (Cheung and Heine, 2015). Monterosso, Royzman, and Schwartz (2010) also write: “Physiologically explained behavior was more likely to be characterized as “automatic,” and willpower and character were less likely to be cited as relevant to the behavior. Physiological explanations of undesirable behavior may mitigate blame by inviting nonteleological causal attributions.” So, clearly, most college students would give a lighter sentence if the individual in question were found to have ‘criminal genes’. But, if these genes really did ’cause’ crime, shouldn’t they be given heavier sentences to keep them on the inside more so those with the ‘non-criminal genes’ don’t have to suffer from the ‘genetically induced’ crime?
Heine (2017: 198-199) also writes:
But is someone really less any responsible for their actions if his or her genes are implicated? A problem with this argument is that we would be hard-pressed to find any actions that we engage in where our genes are not involved—our behaviors do not occur in any gene-free zones. Or, consider this: there actually is a particular genetic variant that, if you possess it, makes you about 40 times more likely to engage in same-sex homicides than those who possess a different variant. (66) It’s known as the Y chromosome—that is, people who possess it are biologically male. Given this, should we infer that Y chromosomes cause murders, and thus give a reduced sentence to anyone who is the carrier of such a chromosome because he is really not responsible for his actions? The philosopher Stephen Morse calls the tendency to excuse a crime because of a biological basis the “fundamental psycholegal error.” (67) The problem with this tendency is that it involves separating yout genes from yourself. Saying “my genes made me do it” doesn’t make sense because there is no “I” that is independent of your genetic makeup. But curiously, once genes are implicaed, people see, to feel that the accused is no longer fully in control of his or her actions.
Further, in the case of a child pornographer, one named Gary Cossey, the court said:
The court predicted that some fifty years from now Cossey’s offense conduct would likely be discovered to be caused by “a gene you were born with. And it’s not a gene you can get rid of.” The court expressed its belief that although Cossey was in therapy, it “can only lead, in my view, to a sincere effort on your part to control, but you can’t get rid of it. You are what you’re born with. And that’s the only explanation for what I see here.”
However, this judge punished Cossey more severely due to the ‘possibility’ that scientists may find ‘genes for’ child pornography use in 50 years. Cossey was then given another, unbiased judge, and was given a ‘more lenient’ sentence than the genetic determinist judge did.
Sean Last over at The Alternative Hypothesis is also a big believer in this so-called MAOA-race difference that explains racial differences in crime. However, as reviewed above (and as he writes), MAOA can be called the “everything but the kitchen sink gene” (Heine, 2017: 195), as I will touch on briefly below, to attribute ’causes’ to genes is not the right way to look at them. It’s not so easy to say that since one ‘has the warrior gene’ that they’d automatically be violent. Last cites a study saying that even those who have the MAOA allele who were not abused showed higher rates of violent behavior (Ficks and Waldman, 2014). They write (pg. 429):
The frequency of the ‘‘risk’’ allele in nonclinical samples of European ancestry ranges from 0.3 to 0.4, although the frequency of this allele in individuals of Asian and African ancestry appears to be substantially higher (*0.6 in both groups; Sabol et al. 1998).
So, why don’t Asians have higher rates of crime—along with blacks—if MAOA on its own causes violent and antisocial behavior? Next I know that someone would claim that “AHA! TESTOSTERONE ALSO MEDIATES THIS RELATIONSHIP!!” However, as I’ve talked about countless times (until I’m blue in the face), blacks do not have/have lower levels of testosterone than whites (Richards et al, 1992; Gapstur et al, 2002; Rohrmann et al, 2007; Mazur, 2009; Lopez et al, 2013; Hu et al, 2014; Richard et al, 2014). Though young black males have higher levels of testosterone due to the environment (honor culture) (Mazur, 2016). So that canard cannot be trotted out.
All in all, these simplistic and reductionist approaches to ‘figuring out’ the ’causes’ of crime do not make any sense. To point at one gene and say that this is ‘the cause’ of that do not make sense.
One last point on ‘genes as causes’ for behavior. This is something that deserves a piece of its own, but I will just provide a quote from Eva Jablonska and Marion Lamb’s book Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life (Jablonska and Lamb, 2014: 17; read chapter one of the book here; I have the nook version so the page number may be different):
Although many psychiatrists, biochemists, and other scientists who are not geneticists (yet express themselves with remarkable facility on genetic issues) still use the language of genes as simple causal agents, and promise their audience rapid solutions to all sorts of problems, they are no more than propagandists whose knowledge or motives must be suspect. The geneticists themselves now think and talk (most of the time) in terms of genetic networks composed of tens or hundreds of genes and gene products, which interact with each other and together affect the development of a particular trait. They recognize that whether or not a trait (a sexual preference, for example) develops does not depend, in the majority of cases, on a difference in a single gene. It involves interactions among many genes, many proteins and other types of molecule, and the environment in which an individual develops.
So to say that those who have low-functioning MAOA variants have an ‘excuse’ as to why they commit crime is incorrect. I know that most people know this, but when you read some people’s writings on things like this it’s like they think that these singular genes/polymorphisms/etc cause these things on their own. In actuality, you need to look at how the whole system interacts with these things, and not reduce whole complex physiological systems to a sum of its parts. This is why implicating singular genes/polymorphisms as explanations for racial differences in crime does not make sense (as can be seen with the Japanese example).
To reduce behaviors simply to gene X and not look at the whole system does not make any sense. There are no ‘genes for’ anything, except a few Mendelian diseases (Ropers, 2010). Stating that certain genes ’cause’ X, as I have shown does not make sense and, wrongly, in my opinion, gives criminals less of a sentencing since judges find stuff like this ‘very compelling’. If that’s the case, why implicate any murderer? ‘Their genes made them do it’, right? Though, things are not that simple to implicate one gene as a cause for crime or any other complex behavior; in this sense—like for most things to do with the human body—holism makes way more sense and not reductionism. We need to look at how these genes that are ‘implicated’ in criminal behavior interact with the whole system. Only then can we understand the causes of criminal behavior. Looking at singular genes impedes us from figuring out the true underlying reasons why people commit crime.
Remember: we can’t blame “warrior genes” for violent crime. If someone does have a ‘genetic predisposition to crime’ from the MAOA gene, then wouldn’t it make more sense to give them more time? Though, the relationship is not so simple as I have covered. So to close, there is no ‘simple relationship’ between race, crime and MAOA. Not in the way that other hereditarians would like you to believe. Because if this relationship were so simple, then East Asians (Chinese, Japanese) would have the highest rates of crime, and they do not.
Responses to The Alternative Hypothesis and Robert Lindsay on Testosterone
2300 words
I enjoy reading what other bloggers write about testosterone and its supposed link to crime, aggression, and prostate cancer; I used to believe some of the things they did, since I didn’t have a good understanding of the hormone nor its production in the body. However, once you understand how its produced in the body, then what others say about it will seem like bullshit—because it is. I’ve recently read a few articles on testosterone from the HBD-blog-o-sphere and, of course, they have a lot of misconceptions in them—some even using studies I have used myself on this blog to prove my point that testosterone does not cause crime!! Now, I know that most people don’t read studies that are linked, so they would take what it says on face value because, why not, there’s a cite so what he’s saying must be true, right? Wrong. I will begin with reviewing an article by someone at The Alternative Hypothesis and then review one article from Robert Lindsay on testosterone.
The Alternative Hypothesis
Faulk has great stuff here, but the one who wrote this article, Testosterone, Race, and Crime, 1) doesn’t know what he’s talking about and 2) clearly didn’t read the papers he cited. Read this article, you’ll see him make bold claims using studies I have used for my own arguments that testosterone doesn’t cause crime! Let’s take a look.
One factor which explains part of why Blacks have higher than average crime rates is testosterone. Testosterone is known to cause aggression, and Blacks are known to at once have more of it and, for genetic reasons, to be more sensitive to its effects.
- No it doesn’t.
- “Testosterone is known to cause aggression“, but that’s the thing: it’s only known that it ’causes’ aggression, it really doesn’t.
- Evidence is mixed on blacks being “… for genetic reasons … more sensitive to its effects” (Update on Androgen Receptor gene—Race/History/Evolution Notes).
Testosterone activity has been linked many times to aggression and crime. Meta-analyses show that testosterone is correlated with aggression among humans and non human animals (Book, Starzyk, and Quinsey, 2001).
Why doesn’t he say what the correlation is? It’s .14 and this study, while Archer, Graham-Kevan and Davies, (2005) reanalyzed the studies used in the previous analysis and found the correlation to be .08. This is a dishonest statement.
Women who suffer from a disease known as congenital adrenal hyperplasia are exposed to abnormally high amounts of testosterone and are abnormally aggressive.
Abnormal levels of androgens in the womb for girls with CAH are associated with aggression, while boys with and without CAH are similar in aggression/activity level (Pasterski et al, 2008), yet black women, for instance, don’t have higher levels of testosterone than white women (Mazur, 2016). CAH is just girls showing masculinized behavior; testosterone doesn’t cause the aggression (See Archer, Graham-Kevan and Davies, 2005)
Artificially increasing the amount of testosterone in a person’s blood has been shown to lead to increases in their level of aggression (Burnham 2007; Kouri et al. 1995).
Actually, no. Supraphysiological levels of testosterone administered to men (200 and 600 mg weekly) did not increase aggression or anger (Batrinos, 2012).
Finally, people in prison have higher than average rates of testosterone (Dabbs et al., 2005).
Dabbs et al don’t untangle correlation from causation. Environmental factors can explain higher testosterone levels (Mazur, 2016) in inmates, and even then, some studies show socially dominant and aggressive men have the same levels of testosterone (Ehrenkraz, Bliss, and Sheard, 1974).
Thus, testosterone seems to cause both aggression and crime.
No, it doesn’t.
Why Testosterone Does Not Cause Crime
Testosterone and Aggressive Behavior
Furthermore, of the studies I could find on testosterone in Africans, they have lower levels than Western men (Campbell, O’Rourke, and Lipson, 2003; Lucas and Campbell, and Ellison, 2004; Campbell, Gray, and Ellison, 2006) so, along with the studies and articles cited on testosterone, aggression, and crime, that’s another huge blow to the testosterone/crime/aggression hypothesis.
Richard et al. (2014) meta-analyzed data from 14 separate studies and found that Blacks have higher levels of free floating testosterone in their blood than Whites do.
They showed that blacks had 2.5 to 4.9 percent higher testosterone than whites, which could not explain the higher prostate cancer incidence (which meta-analyses call in to question; Sridhar et al 2010; Zagars et al 1998). That moderate amount would not be enough to cause differences in aggression either.
Exacerbating this problem even further is the fact that Blacks are more likely than Whites to have low repeat versions of the androgen receptor gene. The androgen reception (AR) gene codes for a receptor by the same name which reacts to androgenic hormones such as testosterone. This receptor is a key part of the mechanism by which testosterone has its effects throughout the body and brain.
The rest of the article talks about CAG repeats and aggressive/criminal behavior, but it seems that whites have fewer CAG repeats than blacks.
Robert Lindsay
This one is much more basic, and tiring to rebut but I’ll do it anyway. Lindsay has a whole slew of articles on testosterone on his blog that show he doesn’t understand the hormone, but I’ll just talk about this one for now: Black Males and Testosterone: Evolution and Perspectives.
It was also confirmed by a recent British study (prostate cancer rates are somewhat lower in Black British men because a higher proportion of them have one White parent)
Jones and Chinegwundoh (2014) write: “Caution should be taken prior to the interpretation of these results due to a paucity of research in this area, limited accurate ethnicity data, and lack of age-specific standardisation for comparison. Cultural attitudes towards prostate cancer and health care in general may have a significant impact on these figures, combined with other clinico-pathological associations.”
This finding suggests that the factor(s) responsible for the difference in rates occurs, or first occurs, early in life. Black males are exposed to higher testosterone levels from the very start.
In a study of women in early pregnancy, Ross found that testosterone levels were 50% higher in Black women than in White women (MacIntosh 1997).
I used to believe this, but it’s much more nuanced than that. Black women don’t have higher levels of testosterone than white women (Mazur, 2016; and even then Lindsay fails to point out that this was pregnant women).
According to Ross, his findings are “very consistent with the role of androgens in prostate carcinogenesis and in explaining the racial/ethnic variations in risk” (MacIntosh 1997).
Testosterone has been hypothesized to play a role in the etiology of prostate cancer, because testosterone and its metabolite, dihydrotestosterone, are the principal trophic hormones that regulate growth and function of epithelial prostate tissue.
Testosterone doesn’t cause prostate cancer (Stattin et al, 2003; Michaud, Billups, and Partin, 2015). Diet explains any risk that may be there (Hayes et al, 1999; Gupta et al, 2009; Kheirandish and Chinegwundoh, 2011; Williams et al, 2012; Gathirua-Mingwai and Zhang, 2014). However in a small population-based study on blacks and whites from South Carolina, Sanderson et al (2017) “did not find marked differences in lifestyle factors associated with prostate cancer by race.”
Regular exercise, however, can decrease PCa incidence in black men (Moore et al, 2010). A lot of differences can be—albeit, not too largely— ameliorated by environmental interventions such as dieting and exercising.
Many studies have shown that young Black men have higher testosterone than young White men (Ellis & Nyborg 1992; Ross et al. 1992; Tsai et al. 2006).
Ellis and Nyborg (1992) found 3 percent difference. Ross et al (1992) have the same problem as Ross et al (1986), which used University students (~50) for their sample. They’re not representative of the population. Ross et al (1992) also write:
Samples were also collected between 1000 h and 1500 h to avoid confounding
by any diurnal variation in testosterone concentrations.
Testosterone levels should be measured near to 8 am. This has the same time variation too, so I don’t take this study seriously due to that confound. Assays were collected “between” the hours of 10 am and 3 pm, which means it was whenever convenient for the student. No controls on activities, nor attempting to assay at 8 am. People of any racial group could have gone at whatever time in that 5 hour time period and skew the results. Assaying “between” those times completely defeats the purpose of the study.
This advantage [the so-called testosterone advantage] then shrinks and eventually disappears at some point during the 30s (Gapstur et al., 2002).
Gapstur et al (2002) help my argument, not yours.
This makes it very difficult if not impossible to explain differing behavioral variables, including higher rates of crime and aggression, in Black males over the age of 33 on the basis of elevated testosterone levels.
See above where I talk about crime/testosterone/aggression.
Critics say that more recent studies done since the early 2000’s have shown no differences between Black and White testosterone levels. Perhaps they are referring to recent studies that show lower testosterone levels in adult Blacks than in adult Whites. This was the conclusion of one recent study (Alvergne et al. 2009) which found lower T levels in Senegalese men than in Western men. But these Senegalese men were 38.3 years old on average.
Alvergne, Fauri, and Raymond (2009) show that the differences are due to environmental factors:
This study investigated the relationship between mens’ salivary T and the trade-off between mating and parenting efforts in a polygynous population of agriculturists from rural Senegal. The men’s reproductive trade-offs were evaluated by recording (1) their pair-bonding/fatherhood status and (2) their behavioral profile in the allocation of parental care and their marital status (i.e. monogamously married; polygynously married).
They also controlled for age, so his statement “But these Senegalese men were 38.3 years old on average” is useless.
These critics may also be referring to various studies by Sabine Rohrmann which show no significance difference in T levels between Black and White Americans. Age is poorly controlled for in her studies.
That is one study out of many that I reference. Rohrmann et al (2007) controlled for age. I like how he literally only says “age is poorly controlled for in her studies“, because she did control for age.
That study found that more than 25% of the samples for adults between 30 and 39 years were positive for HSV-2. It is likely that those positive samples had been set aside, thus depleting the serum bank of male donors who were not only more polygamous but also more likely to have high T levels. This sample bias was probably worse for African American participants than for Euro-American participants.
Why would they use diseased samples? Do you even think?
Young Black males have higher levels of active testosterone than European and Asian males. Asian levels are about the same as Whites, but a study in Japan with young Japanese men suggested that the Japanese had lower activity of 5-alpha reductase than did U.S. Whites and Blacks (Ross et al 1992). This enzyme metabolizes testosterone into dihydrotestosterone, or DHT, which is at least eight to 10 times more potent than testosterone. So effectively, Asians have the lower testosterone levels than Blacks and Whites. In addition, androgen receptor sensitivity is highest in Black men, intermediate in Whites and lowest in Asians.
Wu et al (1995) show that Asians have the highest testosterone levels. Evidence is also mixed here as well. See above on AR sensitivity.
Ethnicmuse also showed that, contrary to popular belief, Asians have higher levels of testosterone than Africans who have higher levels of testosterone than Caucasians in his meta-analysis. (Here is his data.)
The Androgen Receptor and “masculinization”
Let us look at one study (Ross et al 1986) to see what the findings of a typical study looking for testosterone differences between races shows us. This study gives the results of assays of circulating steroid hormone levels in white and black college students in Los Angeles, CA. Mean testosterone levels in Blacks were 19% higher than in Whites, and free testosterone levels were 21% higher. Both these differences were statistically significant.
Assay times between 10 am and 3 pm, unrepresentative sample of college men, didn’t have control for waist circumference. Horribly study.
A 15% difference in circulating testosterone levels could readily explain a twofold difference in prostate cancer risk.
No, it wouldn’t (if it were true).
Higher testosterone levels are linked to violent behavior.
Causation not untangled.
Studies suggest that high testosterone lowers IQ (Ostatnikova et al 2007). Other findings suggest that increased androgen receptor sensitivity and higher sperm counts (markers for increased testosterone) are negatively correlated with intelligence when measured by speed of neuronal transmission and hence general intelligence (g) in a trade-off fashion (Manning 2007).
Who cares about correlations? Causes matter more. High testosterone doesn’t lower IQ. Racial differences in testosterone are tiring to talk about now, but there are still a few more articles I need to rebut.
Conclusion
Racial differences in testosterone don’t exist/are extremely small in magnitude (as I’ve covered countless times). The one article from TAH literally misrepresents studies/leaves out important figures in the testosterone differences between the two races to push a certain agenda. Though if you read the studies you see something completely different. It’s the same with Lindsay. He misunderstood a few studies to push his agenda about testosterone and crime and prostate cancer. They’re both wrong, though.
Why Testosterone Does Not Cause Crime
Testosterone and Aggressive Behavior
Race, Testosterone, and Prostate Cancer
Population variation in endocrine function—Race/History/Evolution Notes
Racial differences in testosterone are tiring to talk about now, but there are still a few more articles I need to rebut. People read and write about things they don’t understand, which is the cause of these misconceptions with the hormone, as well as, of course, misinterpreting studies. Learn about the hormone and you won’t fear it. It doesn’t cause crime, prostate cancer nor aggression; these people who write these articles have one idea in their head and they just go for it. They don’t understand the intricacies of the endocrine system and how sensitive it is to environmental influence. I will cover more articles that others have written on testosterone and aggression to point out what they got wrong.
HBD and Sports: Baseball and Reaction Time
2050 words
If you’ve ever played baseball, then you have first-hand experience on what it takes to play the game, one of the major abilities you need is a quick reaction time. Baseball players are in the upper echelons in regards to pitch recognition and ability to process information (Clark et al, 2012).
Some people, however, believe that there is an ‘IQ cutoff’ in regards to baseball; since general intelligence is supposedly correlated with reaction time (RT), then those with higher RTs must have higher intelligence and vice-versa. However, this trait—in a baseball context—is trainable to an extent. To those that would claim that IQ would be a meaningful metric in baseball I pose two question: would higher IQ teams, on average, beat lower IQ teams and would higher IQ people have better batting averages (BAs) than lower IQ people? This, I doubt, because as I will cover, these variables are trainable and therefore talking about reaction time in the MLB in regards to intelligence is useless.
Meden et al (2012) tested athlete and non-athlete college students on visual reaction time (VRT). They tested the athletes’ VRT once, while they tested the non-athletes VRT two times a week for a 3 week period totaling 6 tests. Men ended up having higher VRTs in comparison to women, and athletes had better VRTs than non-athletes. So therefore, this study proves that VRT is a trainable variable. If VRT can be improved with training, then hitting and fielding can also be trained as well.
Reaction time training is the communication between the brain, musculoskeletal system and spinal cord, which includes both physical and cognitive training. So since VRT can be trained, then it makes logical sense that Major League hitting and fielding can be trained as well.
David Epstein, author of The Sports Gene says that he has a faster reaction time than Albert Pujols:
One of the big surprises for me was that pro athletes, particularly in baseball, don’t have faster reflexes on average than normal people do. I tested faster than Albert Pujols on a visual reaction test. He only finished in the 66thpercentile compared to a bunch of college students.
It’s not a superior RT that baseball players have in comparison to the normal population, says Epstein, but “learned perceptual skills that the MLB players don’t know they learned.” Major League baseball players do have average reaction times (Epstein, 2013: 1) but a far superior visual acuity. Most pro-baseball players had visual acuity of 20/13, with some players having 20/11; the theoretical best visual acuity that is possible is 20/8 (Clark et al, 2012). Laby, Kirschen, and Abbatine show that 81 percent of the 1500 Major and Minor League Mets and Dodgers players had visual acuities of 20/15 or better, along with 2 percent of players having a visual acuity of 20/9.2. Baseball players average a 20/13 visual acuity with the best eyesight humanly possible being 20/8. (Laby et al, 1996).
So it’s not faster RT that baseball players have, but a better visual acuity—on average—in comparison to the general population. Visual reaction time is a highly trainable variable, and so since MLB players have countless hours of practice, they will, of course, be superior on that variable.
Clark et al (2012) showed that high-performance vision training can be performed at the beginning of the season and maintained throughout the season to improve batting parameters. They also state that visual training programs can help hitters, since the eyes account for 80 percent of the information taken into the brain. Reichow, Garchow, and Baird (2011) conclude that a “superior ability to recognize pitches presented via tachistoscope may correlate with a higher skill level in batting.” Clark et al (2012) posit that their training program will help batters to better recognize the spot of the ball and the pitcher’s finger position in order to better identify different pitches. Clark et al (2012) conclude:
The University of Cincinnati baseball team, coaches and vision performance team have concluded that our vision training program had positive benefits in the offensive game including batting and may be providing improved play on defense as well. Vision training is becoming part of out pre-season and in season conditioning program as well as for warmups.
Classe et al (1997) showed that VRT was related to batting, but not fielding or pitching skill. Further, there was no statistically significant difference observed between VRT and age, race or fielding. Therefore, we can say that VRT has no statistical difference on race and does not contribute to any racial differences in baseball.
Baseball and basketball athletes had faster RTs than non-athletes (Nakamoto and Mori, 2008). The Go/NoGo response that is typical of athletes is most certainly trainable. Kida et al (2005) showed that intensive practice improved the Go/NoGo reaction time, but not simple reaction time. Kida et al (2005: 263-264) conclude that simple reaction time is not an accurate indicator of experience, performance or success in sports; Go/NoGo can be improved by practice and is not innate (but simple reaction time was not altered) and the Go/NoGo reaction time can be “theoretically shortened toward a certain value determined by the simple reaction time proper to each individual.“
In baseball players in comparison to a control group, readiness potential was significantly shorter for the baseball players (Park, Fairweather, and Donaldson, 2015). Hand-eye coordination, however, had no effect on earned run average (ERA) or batting average in a sample of 410 Major and Minor League members of the LA Dodgers (Laby et al, 1997).
So now we know that VRT can be trained, VRT shows no significant racial differences, and that Go/NoGo RT can be improved by practice. Now a question I will tackle is: can RT tell us anything about success in baseball and is RT related to intelligence/IQ?
Khodadi et al (2014) conclude that “The relationship between reaction time and IQ is too complicated and revealing a significant correlation depends on various variables (e.g. methodology, data analysis, instrument etc.).” So since the relationship is too complicated between the two variables, mostly due to methodology and the instrument used, RT is not a good correlate of IQ. It can, furthermore, be trained (Dye, Green, and Bavelier, 2012).
In the book A Question of Intelligence, journalist Dan Seligman writes:
In response, Jensen made two points: (1) The skills I was describing involve a lot more than just reaction time, they also depended heavily on physcial coordination and endless practice. (2) It was, however, undoubtedly true that there was some IQ requirement-Jensen guessed it might be around 85- below which you could never recruit for major league baseball. (About one-sixth of Americans fall below 85).
I don’t know where Jensen grabbed the ‘IQ requirement’ for baseball, which he claims to be around 85 (which is at the black average in America). This quote, however, proves my point that there is way more than RT involved in hitting a baseball, especially a Major League fastball:
Hitting a baseball traveling at 100 mph is often considered one of the most difficult tasks in all of sports. After all, if you hit the ball only 30% of the time, baseball teams will pay you millions of dollars to play for them. Pitches traveling at 100 mph take just 400 ms to travel from the pitcher to the hitter. Since the typical reaction time is 200 ms, and it takes 100 ms to swing the bat, this leaves just 100 ms of observation time on which the hitter can base his swing.
This lends more credence to the claim that hitting a baseball is more than just quick reflexes; considerable training can be done to learn certain cues that certain pitchers use; for instance, like identifying different pitches a particular pitcher does with certain arm motions coming out of the stretch. This, as shown above in the Epstein quote, is most definitely a trainable variable.
Babe Ruth, for instance, had better hand-eye coordination than 98.8 percent of the population. Though that wasn’t why he was one of the greatest hitters of all time; it’s because he mastered all of the other variables in regards to hitting, which are learnable and not innate.
Witt and Proffitt (2005) showed that the apparent ball size is correlated with batting average, that is, the better batters fared at the plate, the bigger they perceived the ball to be so they had an easier time hitting it. Hitting has much less to do with reaction time and much more to do with prediction, as well as the pitching style of the pitcher, his pitching repertoire, and numerous other factors.
It takes a 90-95 mph fast ball about 400 milliseconds to reach home plate. It takes the brain 100 milliseconds to process the image that the eyes are taking in, 150 milliseconds to swing and 25 milliseconds for his brain to send a signal to his body to swing. This leaves the hitter with 125 milliseconds left to hit the incoming fastball. Clearly, there is more to hitting than reaction time, especially when all of these variables are in play. Players have .17 seconds to decide whether or not to hit a pitch and where to place their bat (Clark et al, 2012)
A so-called ‘IQ cutoff’ for baseball does exist, but only because IQs lower than 85 (once you begin to hit the 70s range, especially the lower levels) indicate developmental disorders. Further, the 85-115 IQ range encompasses 68 percent of the population. However, RT is not even one of the most important factors in hitting; numerous other (trainable) variables influence fastball hitting, and all of the best players in the world employ these strategies. People may assume that since intelligence and RT are (supposedly) linked, that baseball players, since they (supposedly) have quick RTs. Nevertheless, if quick RTs were correlated with baseball profienciency—namely, in hitting, then why are Asians 1.2 percent of the players in the MLB? Maybe because RT doesn’t really have anything to do with hitting proficiency and other variables have more to do with it.
People may assume that since intelligence and RT are (supposedly) linked, that baseball players, since they (supposedly) have quick RTs then they must be intelligent and therefore there must be an IQ cutoff because intelligence/g and RT supposedly correlate. However, I’ve shown 2 things: 1) RT isn’t too important to hitting at an elite level and 2) more important skills can be acquired in hitting fastballs, most notable, in my opinion, is pitch verification and the arm location of the pitcher. The Go/NoGo RT can also be trained and is, arguably, one of the most important training systems for elite hitting. Clearly, elite hitting is predicated on way more than just a quick RT; and most of the variables that are involved in elite hitting are most definitely trainable, as reviewed in this article.
People, clearly, make unfounded claims without having any experience in something. It’s easy to make claims about something when you’re just looking at numbers and attempting to draw conclusions based on data. But it’s a whole other ballgame (pun intended) when you’re up at the plate yourself or coaching someone on how to hit or play in the infield. These baseless claims would be avoided a lot more if only the people who make these claims had any actual athletic experience. If so, they would know of the constant repetition that goes into hitting and fielding, the monotonous drills you have to do everyday until your muscle memory is trained to flawlessly—without even thinking about it—throw a ball from shortstop to first base.
Practice, especially Major League practice, is pivotal to elite hitting; only with elite practice can a player learn how to spot the ball and the pitcher’s finger position to quickly identify the pitch type in order to decide if he wants to swing or not. In conclusion, a whole slew of cognitive/psychological abilities are involved in the upper echelons of elite baseball, however a good majority of the traits needed to succeed in baseball are trainable, and RT has little to do with elite hitting.
(When I get time I’m going to do a similar analysis like what I wrote about in the article on my possible retraction of my HBD and baseball article. Blacks dominate in all categories that matter, this holds for non-Hispanic whites and blacks as well as Hispanic blacks and whites, read more here. Nevertheless, I may look at the years 1997-2017 and see if anything has changed from the analysis done in the late 80s. Any commentary on that matter is more than welcome.)
Racial Differences in Testosterone…again
1200 words
Testosterone is a fascinating hormone—the most well-known hormone to the lay public. What isn’t well-known to the lay public is how the hormone is produced and the reasons why it gets elevated. I’ve covered racial differences in testosterone in regards to crime, penis size and Rushton’s overall misuse of r/K selection theory. In this article, I will talk about what raises and decreases testosterone, as well as speak about racial differences in testosterone again since it’s such a fun topic to cover.
JP Rushton writes, in his 1995 article titled Race and Crime: An International Dilemma:
One study, published in the 1993 issue of Criminology by Alan Booth and D. Wayne Osgood, showed clear evidence of a testosterone-crime link based on an analysis of 4,462 U.S. military personnel. Other studies have linked testosterone to an aggressive and impulsive personality, to a lack of empathy, and to sexual behavior.
Booth and Osgood (1993: 93) do state that “This pattern of results supports the conclusions that (I) testosterone is one of a larger constellation of factors contributing to a general latent propensity toward deviance and (2) the influence of testosterone on adult deviance is closely tied to social factors.” However, as I have extensively documented, the correlation between testosterone and aggression is extremely low (Archer, 1991; Book et al (2001); Archer, Graham-Kevan and Davies, 2005), and therefore cannot be the cause of crime.
Another reason why testosterone is not the cause of aggression/deviant behavior is due to what times most crimes are committed at. Therefore, testosterone cannot possibly be the cause of crime. I’ve also shown that, contrary to popular belief, blacks don’t have higher levels testosterone than whites, along with the fact that testosterone does not cause prostate cancer, that even if blacks did have these supposed higher levels of the hormone, that it would NOT explain higher rates of crime.
Wu et al (1995) show that Asian Americans had the highest testosterone levels, African Americans were intermediate and European Americans were last, after adjustments for BMI and age were made. Though, I’ve shown in larger samples that, if there is any difference at all (and a lot of studies show no difference), it is a small advantage favoring blacks. We then are faced with the conclusion that this would not explain disease prevalence nor higher rates of crime or aggression.
Testosterone, contrary to Rushton’s (1999) assertion, is not a ‘master switch’. Rushton, of course, cites Ross et al (1986) which I’ve tirelessly rebutted. Assay times were all over the place (between 10 am and 3 pm) with testosterone levels being highest at 8 am. The most important physiological variable in Rushton’s model is testosterone, and without his highly selected studies, his narrative falls apart. Testosterone doesn’t cause crime, aggression, nor prostate cancer.
The most important take away is this: Rushton’s r/K theory hinges on 1) blacks having higher levels of testosterone than whites and 2) that these higher levels of testosterone then influence higher levels of aggression which lead to crime and then prostate cancer. Even then, Sridhar et al (2010) meta-analyzed 17 articles which talk about racial differences in prostate cancer survival rates. They state in their conclusion that “there are no differences between African American and Whites in survival from prostate cancer.” Zagars et al (1998) show that there were no significant racial differences in serum testosterone. Furthermore, when matched for major prognostic factors “the outcome for clinically local–regional prostate cancer does not depend on race (6,7,14–19). Moreover there appear to be no racial differences in the response of advanced prostate cancer to androgen ablation (29,47). Our study provides further evidence that racial differences in disease outcome are absent for clinically localized prostate cancer” (Zagars et al, 1998: 521). So it seems that these two studies also provide further support that Rushton et al were wrong in regards to prostate cancer mortality as well.
Rushton (1997 185) writes:
In any case, socialization cannot account for the early onset of the traits, the speed of dental and other maturational variables, the size of the brain, the number of gametes produced, the physiological differences in testosterone, nor the evidence on cross-cultural consistency.
There are no racial differences in testosterone and if there were, social factors would explain the difference between the races. However, as I’ve noted in the past, testosterone levels are high in young black males with low educational attainment (Mazur, 2016). The higher levels of testosterone in blacks compared to whites (which, if you look at figure 1 the levels are not high at all) is accounted for by honor culture, a social variable. Furthermore, the effects of the environment are also more notable on testosterone than are genetics at 5 months of age (Carmaschi et al, 2010). Environmental factors greatly influence testosterone (Booth et al, 2006), so Rushton stating that “socialization cannot account for the early onset” of “physiological differences in testosterone” is clearly wrong since environmental influences can be seen in infants as well as adults. Testosterone is strongly mediated by the environment; this is not up for debate.
Testosterone is one of many important hormones in the body; the races do not differ in the variable. So, therefore, all of Rushton’s ‘r/K predictions’, which literally hinge on testosterone (Lynn, 1990) fall apart without this ‘master switch’ (Rushton, 1999) driving all of these behaviors. Any theories of crime that include testosterone as a main driver in crime need to be rethunk; numerous studies attest to the fact that testosterone does not cause crime. Racial differences in testosterone only appear in small studies and the studies that do show these differences get touted around all the while, all of the better, larger analyses don’t get talked about because it goes against a certain narrative.
Finally, there is no inevitability of a testosterone decrease in older men. So-called “age-related declines” in the hormone are largely explained by smoking, obesity, chronic disease, marital status, and depression (Shi et al, 2013), and even becoming a father explains lower levels of testosterone (Gray, Yang, and Pope, 2006). On top of that, marriage also reduces testosterone, with men who went from unmarried to married showing a sharp decline in testosterone over a ten-year period (Holmboe et al, 2017). This corroborates numerous other studies showing that marriage lowers testosterone levels in men (Mazur and Michalek, 1998; Nansunga et al, 2014) But some of this decrease may be lessened by frequent sexual intercourse (Gettler et al, 2013). So if you live a healthy lifestyle, the testosterone decrease that plagues most men won’t occur to you. The decreases are due to lifestyle changes; not explicitly tied to age.
People are afraid of higher levels of testosterone at a young age and equally as terrified of lowering testosterone levels at an old age. However, I’ve exhasutively shown that testosterone is not the boogeyman, nor the ‘master switch’ (Rushton, 1999) it’s made out to be. There are no ‘genes for’ testosterone; its production is indirect through DNA. Thusly, if you keep an active lifestyle, don’t become obese, and don’t become depressed, you can bypass the so-called testosterone decrease. Fear mongering on both sides of the ‘testosterone curve’ are seriously blown out of proportion. Testosterone doesn’t cause crime, aggression, nor prostate cancer (even then, large meta-analyses show no difference in PCa mortality between blacks and whites).
The fear of the hormone testosterone is due to ignornace of what it does in the body and how it is produced in the body. If people were to understand the hormone, they would not fear it.
