
Home » Posts tagged 'anatomy'
Tag Archives: anatomy
A Systems View of Kenyan Success in Distance Running
1550 words
The causes of sporting success are multi-factorial, with no cause being more important than the other since the whole system needs to work in concert to produce the athletic phenotype–call this “causal parity” of athletic success determinants. For a refresher, take what Shenk (2010: 107):
As the search for athletic genes continues, therefore, the overwhelming evidence suggests that researchers will instead locate genes prone to certain types of interactions: gene variant A in combination with gene variant B, provoked into expression by X amount of training + Y altitude + Z will to win + a hundred other life variables (coaching, injuries, etc.), will produce some specific result R. What this means, of course, What this means, of course, is that we need to dispense rhetorically with thick firewall between biology (nature) and training (nurture). The reality of GxE assures that each person’s genes interacts with his climate, altitude, culture, meals, language, customs and spirituality—everything—to produce unique lifestyle trajectories. Genes play a critical role, but as dynamic instruments, not a fixed blueprint. A seven- or fourteen- or twenty-eight-year-old is not that way merely because of genetic instruction. (Shenk, 2010: 107) [Also read my article Explaining African Running Success Through a Systems View.]
This is how athletic success needs to be looked at; not reducing it to genes or a group of genes that ’cause’ athletic success. Since to be successful in the sport of the athlete’s choice takes more than being born with “the right” genes.
Recently, a Kenyan woman—Joyciline Jepkosgei—won the NYC marathon in here debut (November 3rd, 2019), while Eliud Kipchoge—another Kenyan—became the first human ever to complete a marathon (26.2 miles) in under 2 hours. I recall in the spring reading that he said he would break the 2-hour mark in October. He also attempted to break it in 2017 in Italy but, of course, he failed. His official time in Italy was 2:00:25! While he set the world record in Berlin at 2:01:39. Kipchoge’s official time was 1:59:40—twenty seconds shy of 2 hours—that means his average mile pace was about 4 minutes and 34 seconds. That is insane. (But the IAAF does not accept the time as a new world record since it was not in an open competition—Kipchoge had a slew of Olympic pacesetters following him; an electric car drove just ahead of him and pointed lasers at the ground showing him where to run; so he shaved 2 minutes off his time—2 crucial minutes—according to sport scientist Ross Tucker; and . So he did not set a world record. His feat, though, is still impressive.)
Now, Kipchoge is Kenyan—but what’s his ethnicity? Surprise surprise! He is of the Nandi tribe, more specifically, of the Talai subgroup, born in Kapsisiywa in the Nandi county. Jepkosgei, too, is Nandi, from Cheptil in Nandi county. (Jepkosgei also set the record for the half marathon in 2017. Also, see her regular training regimen and what she does throughout the day. This, of course, is how she is able to be so elite—without hard training, even without “the right genetic makeup”, one will not become an elite athlete.) What a strange coincidence that these two individuals who won recent marathons—and one who set the best time ever in the 26.2 mile race—are both Kenyan, specifically Nandi?
Both of these runners are from the same county in Kenya. Nandi county is elevated about 6,716 ft above sea level. Being born and living at a high elevation means that they have different kinds of physiological adaptations due to being born at such a higher elevation. Living and training at such high elevations means that they have greater lung capacities since they are breathing in thinner air. Those born in highlands like Kipchoge and Jepkosgei have larger lungs and thorax volumes, while oxygen intake is enhanced by increases in lung compliance, pulmonary diffusion, and ventilation (Meer, Heymans, and Zijlstra, 1995).
Those exposed to such elevation develop what is known as “high-altitude hypoxia.” Humans born at high altitudes are able to cope with such a lack of oxygen, since our physiological systems are dynamic—not static—and can respond to environmental changes within seconds of them occurring. Babes born at higher elevations have increased ventilation, and a rise in the alveolar and the pressure of arterial oxygen (Meer, Heymans, and Zjilstra, 1995).
Kenyans have 5 percent longer legs and 12 percent lighter muscles than Scandinavians (Suchy and Waic, 2017). Mooses et al (2014) notes that “upper leg length, total leg length and total leg length to body height ratio were correlated with running performance.” Kong and de Heer (2008) note that:
The slim limbs of Kenyan distance runners may positively contribute to performance by having a low moment of inertia and thus requiring less muscular effort in leg swing. The short ground contact time observed may be related to good running economy since there is less time for the braking force to decelerate forward motion of the body.
An abundance of type I muscle fibers is conducive to success in distance running (Zierath and Hawley, 2004), though Kenyans and Caucasians have no difference in type I muscle fibers (Saltin et al, 1995; Larsen and Sheel, 2015). That, then, throws a wrench in the claim that a whole slew of anatomic and physiologic variables conducive to running success is the cause for Kenyan running success—specifically the type I fibers—right? Wrong. Recall that the appearance of the athletic phenotype is due to nature and nurture—genes and environment—working together in concert. Kenyans are more likely to have slim, long limbs with lower body fat while they lived and trained over 6000 ft high. Their will to win to better themselves and their families’ socioeconomic status, too, plays a part. As I have argued in-depth for years—we cannot understand athletic success and elite athleticism without understanding individual histories, how they grew up, and what they did as a child.
For example, Wilbur and Pitsiladis (2012) espouse a systems view of Kenyan marathon success, writing:
In general, it appears that Kenyan and Ethiopian distance-running success is not based on a unique genetic or physiological characteristic. Rather, it appears to be the result of favorable somatotypical characteristics lending to exceptional biomechanical and metabolic economy/efficiency; chronic exposure to altitude in combination with moderate-volume, high-intensity training (live high + train high), and a strong psychological motivation to succeed athletically for the purpose of economic and social advancement.
Becoming a successful runner in Kenya can lead to economic opportunities not afforded to those who do not do well in running. This, too, is a factor in Kenyan running success. So, for the ignorant people who would—pushing a false dichotomy of genes and environment—state that Kenyan running success is due to “socioeconomic status”—they are right, to a point (even if they are mocking it and making their genetic determinism seem more palatable). See figure 6 for their hypothetical model:

This is one of the best models I have come across explaining the success of these people. One can see that it is not reductonist; note that there is no appeal to genes (just variables that genes are implicated IN! Which is not the same as reductionism). It’s not as if one can have an endomorphic somatotype with Kenyan training and their psychological reasons for becoming runners. The ecto-dominant somatotype is a necessary factor for success; but all four of these—biomechanical & physiological, training, and psychological—factors explain the success of the running Kenyans and, in turn, the success of Kipchoge and Jepkosgei. African dominance in distance running is, also, dominated by the Nandi subtribe (Tucker, Onywera, and Santos-Concejero, 2015). Knechtle et al (2016) also note that male and female Kenyan and Ethiopian runners are the youngest and fast at the half and full marathons.
The actual environment—climate—on the day of the race, too plays a factor. El Helou et al (2012) note that “Air temperature is the most important factor influencing marathon running performance for runners of all levels.” Nikolaidis et al (2019) note that “race times in the Boston Marathon are influenced by temperature, pressure, precipitations, WBGT, wind coming from the West and wind speed.”
The success of Kenyans—and other groups—shows how the dictum “Athleticism is irreducible to biology” (St. Louis, 2004) is true. How does it make any sense to attempt to reduce athletic success down to one variable and say that that explains the overrepresentation of, say, Kenyans in distance running? A whole slew of factors needs to occur to an individual, along with actually wanting to do something, in order for them to succeed at distance running.
So, what makes Kenyans like Kipchoge and Jepkosgei so good at distance running? It’s due to an interaction with genes and environment, since we take a systems and not a reductionist view of sport success. Even though Kipchoge’s time does not count as an official world record, what he did was still impressive (though not as impressive if he would have done so without all of the help he had). Looking at the system, and not trying to reduce the system to its parts, is how we will explain why some groups are better than others. Genes, of course, play a role in the ontogeny of the athletic phenotype, but they are not the be-all-end-all that genetic reductionists seem to make it out to be. The systems view for Kenyan running success shown here is how and why Kenyans—Kipchoge and Jepkosgei—dominate distance running.
HBD and Sports: Basketball
1600 words
In the past, I have written on the subject of HBD and sports (it is a main subject of this blog). I have covered baseball, football, running, bodybuilding, and strength over many articles. Though, I have not covered basketball yet. Black Americans comprised 74.4 percent of the NBA, compared to 19.1 percent of whites (TIDES, 2017). Why do blacks dominate the racial composition of baskeball? Height is strongly related to success in basketball, though whites and blacks are around the same height, with blacks being slightly shorter (blacks being 69.4 inches compared to whites who were 69.8 inches; CDC, 2012). So, why do blacks dominate basketball?
Basketball success isn’t predicated so much on height, rather, limb length plays more of a factor in basketball success. Blacks have longer limbs than whites (Wagner and Heyward, 2000; Bejan, Jones, and Charles, 2010). The average adult man has an arm span about 2.1 inches greater than his height (Nwosu and Lee, 2008), while Monson, Brasil, and Hlusko (2018) state that taller basketball players had a greater height-to-wingspan ratio and they were, therefore, more successful. The Bleacher Report reports that:
The average NBA Player’s wingspan differential came out at 4.3 percent, so anything above that is going to be reasonably advantageous.
So, more successful basketball players have a longer arm span compared to their height, which makes them more successful in the sport. Blacks have longer limbs than whites, even though they are on average the same height. Thus, one reason why blacks are more successful than whites at basketball is due to their somatotype—their long limbs, specifically,
David Epstein (2014: 129) writes in The Sports Gene:
Based on data from the NBA and NBA predraft combines (using only true, shoes-off measurements of players), the Census Bureaum abd the Centers for Disease Control’s National Center for Health Statistics, there is such a premium on extra height for NBA that the probability of an American man between the ages of twenty and forty being a current NBA player rises nearly a full order of magnitude with every two-inch increase in height starting at six feet. For a man between six feet and 6’2”, the chance of his currently being in the NBA is five in a million. At 6’2” to 6’4”, that increases to twenty in a million. For a man between 6’10” and seven feet tall, it rises to thirty-two thousand in a million, or 3.2 percent. An American man who is seven feet tall is such a rarity that the CDC does not even list a height percentile at that stature. But the NBA measurements combined with the curve formed by the CDC’s data suggest that of American men ages twenty to forty who stand seven feet tall, a startling 17 percent of them are in the NBA right now.* Find six honest seven-footers, and one will be in the NBA.
* Many of the men who NBA rosters claim are seven feet tall prove to be an inch or even two inches shorter when measured at the combine with their shows off. Shaquille O’Neal, however, is a true 7’1” with his shoes off.
And on page 132 he writes:
The average arm-span-to-height ratio of an NBA player is 1.063. (For medical context, a ratio greater than 1.05 is one of the traditonal diagnostic criteria for Marfan syndrome, the disorder of the body’s connective tissues that results in elongated limbs.) An average-height NBA player, one who is about 6’7”, has a wingspan of seven feet.
So we can clearly see that NBA players, on average, are freaks of nature when it comes to limb length, having freakish arm length proportions which is conducive to success in basketball.
Why are long limbs so conducive to basketball success? I can think of a few reasons.
(1) The taller one is and the longer one’s limbs are the less likely they are to have a blocked shot.
(2) The taller one is and the longer one’s limbs are is advantageous when performing a lay-up.
(3) The taller one is and the longer one’s limbs are means they can battle for rebounds at better than a shorter man with shorter limbs.
Epstein (2014: 136) also states that the predraft data shows that the average white NBA player is 6’7.5” with a wingspan of 6’10” while the average black NBA player is 6’5.5” with an average wingspan of 6’11”—meaning that blacks were shorter but “longer.” What this means is that blacks don’t play at “their height”—they play as if they were taller due to their wingspan.
Such limb length differences are a function of climate. Shorter, stockier bodies (i.e., an endomorphic somatotype) is conducive to life in colder climes, whereas longer, more narrowbodies (ecto-meso) are conducive to life in the tropics. Endomorphic somas are conducive to life in colder climes because there is less surface area to keep warm—and this is seen by looking at those whose ancestors evolved in cold climes (Asians, Inuits)—shorter, more compact bodies retain more heat. Conversely, ecto-meso somas are conducive to life in hotter, more tropical climes since this type of body dissipates heat more efficiently than endo somas (Lieberman, 2015). So, blacks are more likely to have the soma conducive to basketball success due to where their ancestors evolved.
So, now we have discussed the facts that height and limb length are conducive to success in basketball. Although blacks and whites in America are the same height, they have vastly different average limb lengths, as numerous studies attest to. These average differences in limb length are how and why blacks succeed far better than whites in the NBA.
Athleticism is irreducible to biology (Lewis, 2004), as has been argued in the past. However, that does not mean that there are NOT traits that are conducive to success in basketball and other sports. Both height and limb length are related: more than likely, the taller one is, the longer their limbs are relative to their height. This is what we see in elite NBA players. Height, will, altitude, and myriad other factors combine to create the elite NBA phenotype; height seems to be a necessary—not sufficient—condition for basketball success (since one can be successful at basketball without the freakish heights of the average player). Though, as Epstein wrote in his book, both height and limb length are conducive to success in basketball, and it just so happens that blacks have longer limbs than whites which of course translates over to their domination in basketball.
Contrary to popular belief, though, players coming from broken homes and an impoverished life are not the norm. As Dubrow and Adams (2010) write:
We find that, after accounting for methodological problems common in newspaper data, most NBA players come from relatively advantaged social origins and African Americans from disadvantaged social origins have lower odds of being in the NBA than African American and white players from relatively advantaged origins.
Sports writer Peter Keating writes that:
[Dubrow and Adams] found that among African-Americans, a child from a low-income family has 37 percent lower odds of making the NBA than a child from a middle- or upper-income family. Poor white athletes are 75 percent less likely to become NBA players than middle-class or well-off whites. Further, a black athlete from a family without two parents is 18 percent less likely to play in the NBA than a black athlete raised by two parents, while a white athlete from a non-two-parent family has 33 percent lower odds of making the pros. As Dubrow and Adams put it, “The intersection of race, class and family structure background presents unequal pathways into the league.”
(McSweeney, 2008 also has a nice review of the matter.)
Turner et al (2015) state that black males were more likely to play basketball than whites males. Higher-income boys were more likely to play baseball, whereas lower-income boys were more likely to play basketball. Though, it seems that when it comes to elite basketball success, players seem to come from higher-income homes.
Therefore, to succeed in basketball, one needs height and long limbs to succeed in basketball. Contrary to popular belief, it is less likely for an NBA player to come from a low-income family—they come from middle-class families the most. Indeed, those who come from lower-income families, even if they have the skill, most likely won’t have the money to develop the talent they have. Though there are some analyses which point to basketball being played by lower-income children—and I have no reason to disagree with them—when it comes to professional play, both blacks and whites are less likely to become NBA players if they grew up in poverty.
The limb length differences between blacks and whites which are conducive to sport success are a function of the climate that their ancestors evolved in. Now, although athleticism is irreducible to biology (because biological and cultural factors interact to create the elite athletic phenotype), that does not mean that there are no traits conducive to sporting success. Quite the opposite: A taller player would more often than not beat a shorter player; when it comes to players with the same height and different limb lengths, the one with the longer limbs will stand a better chance at beating the one with shorter limbs. Blacks and whites have different limb lengths, and this explains how and why blacks are more successful at basketball than whites. Cultural and biological factors combine in order to cause what one is good at.
Basketball is huge in the black community (due in part to people gravitating toward what they are good at), and due to this, since blacks have an advantage right out of the gate, they will gravitate more toward the sport and, therefore, height and limb length is a huge reason why black dominate at this sport.
Usain Bolt, Michael Phelps, and Caster Semenya: Should Semenya Take Drugs to Decrease Testosterone Levels?
1300 words
In the past week in the world of sport, all the rage has been over mid- to long-distance runner Caster Semenya. Semenya has won the 800 m in 1:56.72 and setting world records in the 400, 800, and 1500 m with times of 50.74, 1:58.45 and 4:10.93 respectively. In 2012 and 2016, Semenya won the gold for the 800 m with times of 1:57.23 and 1:55.28 respectively. I won’t really discuss the anatomic and physiologic advantages today. What I will discuss, though, is the fact that Semenya has been told that she has to take drugs to decrease her testosterone levels or she cannot compete anymore. Semenya was told to decrease her testosterone levels or she could face a ban in the 800 m. The new rules state that:
Female athletes affected must take medication for six months before they can compete, and then maintain a lower testosterone level.
If a female athlete does not want to take medication, then they can compete in:
- International competitions in any discipline other than track events between 400m and a mile
- Any competition that is not an international competition
- The male classification at any competition, at any level, in any discipline
- Any intersex, or similar, classification
But Semenya has declined taking these drugs—so her future is up in the air. So, if Semenya—or any other athlete—has to take drugs to decrease their levels since it gives an unfair advantage, then, in my opinion, this may lead to changes in other sports as well.
Look at Michael Phelps. Michael Phelps has won a record 28, winning 23 medals at Rio in 2016. Phelps has a long, thin torso which decreases drag in the water. Phelps’ wing span is 6’7” while he is 6’4”—which is disproportionate to his height. He has the torso of a 6’8” person, which gives him a greater reach per stroke. His lower body is 5’10” which lowers the resistance against the water. He has large hands and feet (with flexible ankles), which help with paddling capacity (size 14 shoe; yours truly wears a size 13).
There is one more incredible thing about Phelps: He produces around 50 percent lower lactic acid. Think of the last time that you have run for some distance. The burning you feel in your legs is a build up of lactic acid. Lactic acid causes fatigue and also slows muscle contractions—this occurs through lactic acid passing through the bloodstream, becoming lactate. (Note that it does not necessarily cause fatigue; Brooks, 2001.) Phelps does not produce normal levels of lactic acid, and so he is ready to go again shortly after a bout of swimming.
Phelps said “In between the 200m free and the fly heats I have probably had in total about 10 minutes to myself.” A normal person’s muscles would be too fatigued and cramped. I would also assume that Phelps has an abundance of type I muscle fibers as well.
Now take Usain Bolt. The 100 m dash is, mostly, an anaerobic race. What this means is that mitochondrial respiration has minimal effect on the type of energy used during the event (Majumdar and Robergs, 2011). So during anaerobic events, there is no free oxygen to drive energy—the energy stored in the muscle is used to perform movement through a process called glycolysis. Sprinting is an intense exercise—fuel choice during exercise is determined by the intensity of said exercise. “A 100-meter sprint is powered by stored ATP, creatine phosphate, and anaerobic glycolysis of muscle glycogen.”
Now we can look at the physical advantages they have. Swimmers and runners, on average, have different centers of mass (Bejan, Jones, and Charles, 2010). In all actuality, Phelps and Bolt are the perfect example of this phenomenon. Winning runners have a West-African origin and winning runners are more likely to be white. These somatotypic differences between the two races influence why they excel in these two different sports.
Usain Bolt is 6’5”. Since he is that height, and he has long legs, he necessarily has a longer stride—Bolt is the perfect example of Bejan, Jones, and Charles’ (2010) paper. So take the average white sprinter of the same height as Bolt. Ceteris paribus, Bolt will have a higher center of mass than the white athlete due to his longer limbs and and smaller circumference. Krogman (1970) found that, in black and white youths of the same height, blacks had shorter trunks and longer limbs, which lends credence to the hypothesis.
Phelps is 6’4”. As noted above, he has a long torso and long limbs. Long torsos are conducive to a lower center of mass—what whites and Asians have, on average. So long torsos mean that one will have taller sitting heights than those with short torsos. This means that whites and Asians have taller sitting heights than blacks, who have shorter torsos. This average taller sitting height is conducive to the longer torsos which is why whites excel in swimming. Bejan, Jones, and Charles (2010) also note that, if it were not for the short stature of Asians, they would be better swimmers than whites.
In any case, the different centers of mass on average between blacks and whites are conducive to faster times in the sports they excel at. For whites, the three percent increase in center of mass means that there would be a 1.5 percent increase in winning speed and a 1.5 percent decrease in winning time in the case of swimming. The same holds for blacks, but in the case of running: their higher center of mass is conducive to a 1.5 percent increase in winning speed and also a 1.5 percent decrease in winning time, which would be a .15 second decrease, or from 10 s to 9.85 s—which is a large differential when it comes to sprinting. (Note that this phenomenon also holds for black women and white women—black women are better sprinters and white women are better swimmers. Asian women excel in the 100 m freestyle, but not Asian men for reasons discussed above.)
Now put this all together. If Phelps and Bolt have such advantages over their competition and they—supposedly—win due to them, then if Semenya has to decrease her T levels, why shouldn’t Phelps and Bolt decrease X, Y, or Z since they have physiologic/anatomic advantages as well? Why does no one talk about Semenya’s anatomic advantages over, say, white women and why only bring up Semenya’s testosterone levels? Forcing Semenya to decrease T levels will set a bad precedent in sport. What would stop a losing competitor from complaining that the winner—who keeps winning—has an “unfair” physiologic/anatomic advantage and must do X to change it? (Or say that the anatomic advantage they possess is “unfair” and they should be barred from competition?)
Here’s the thing: Watching sport, we want to see the best-of-the-best compete. Wouldn’t that logically imply that we want to see Semenya compete and not rid herself of her advantage? If Semenya’s physiologic advantage(s) is being discussed, why not Semenya’s anatomic advantages? It does not make sense to focus on one variable—as all variables interact to produce the athletic phenotype (Louis, 2004). Phelps and Bolt perfectly embody the results of Bejan, Jones, and Charles (2010)—they have, what I hope are—well-known advantages, and these advantages, on average, are stratified between race due to anatomic differences (see Gerace et al, 1994; Wagner and Heyward, 2000).
Phelps and Bolt have anatomic and physiologic advantages over their competition, just as Semenya does, just like any elite athlete, especially the winners compared to their competition. If Semenya is forced to decrease her testosterone levels, then this will set a horrible precedent for sport, and people may then clamor for Phelps and Bolt to do X, Y, and Z due to their physical advantages. For this reason, Semenya should not decrease her testosterone levels and should be allowed to compete in mid-distance running.
The White-Black Swimming/Drowning Gap and Disparities in Elite Running and Swimming Competitions
1600 words
Blacks are better sprinters and whites are better swimmers. Why is this? A whole slew of factors influence this—social, physiological, anatomic. However, there is a stereotype about blacks that has been repeated since I was a child: that blacks can’t swim. How true is this? If it is true, what explains it? It is my opinion that it is true, and that social, cultural, and anatomic and physiologic factors account for this. The same for whites and running. Black children drown at a rate of about 3 times higher than white children. About 70 percent of black children cannot swim, compared to 60 percent of “Hispanic” children and 40 percent of white children. Why is that? Well, one of the most telling answers why is anatomic. Irwin et al (2011) note in their study that blacks are more likely to be “aquaphobes”—having a fear of water—compared to whites.
Almost three years ago, I wrote White Men Can’t Jump? That’s OK, Black Men Can’t Swim. In the article, I explain how and why blacks have a harder time swimming than whites. One anatomic reason is their chest cavity. Compared to whites, blacks have a narrower chest cavity. They have denser, shallower chests. This is a burden while swimming, since those who have a wider chest can take longer strides with their arms while swimming. Blacks have denser bones than whites (Ettinger et al, 1997), Swimmers have lower bone density than non-swimmers (Gomez-Bruton et al, 2013), and so, high bone density is not conducive to swimming success, either.
The first black man to make the swim team for America in the Olympics was Anthony Ervin in 2000. (Funny story. In a class I took a few years ago, racial differences in sports came up. I brought up race differences in swimming. A black guy behind me said “My grandfather was the first to qualify for the Olympics.” I said “Yea? Your grandfather is Anthony Ervin?” He didn’t say anything, it seemed like he got mad at me for calling him out.) That it took this long for a black man to qualify for the US in swimming is telling, and anatomy and physiology, in my opinion, are how we can explain the observed disparity,
So, blacks have lower body fat (on average), and narrower chest cavities. These two things play a role in why blacks are not good swimmers. Yet another role-player, could be, the fact that black women don’t want their hair to get wet and so never taught their children how to swim, parental encouragement, to “swimming is something that white people do” (Wiltse, 2014). Who knows? Maybe in the coming years, blacks could match whites at swimming. Though, with what we know about anatomy and physiology of elite swimmers, this is highly unlikely. It’s like saying “Who knows? Maybe in the coming years, whites could match blacks at running.” Our knowledge of anatomy and physiology throws a wrench in claims like that.
The phenomenon of fast black sprinters and fast white swimmers is predictable through physics (Bejan, Jones, and Charles, 2010). The finalists of running competitions are continuously black, whereas in the swimming competitions they are continuously white. What accounts for this? Well, other than the factors discussed above, there is one more: center of mass.
It is well-known that different races have different anatomic measurements. Blacks have longer limbs than whites (Gerace et al, 1994; Wagner and Heyward, 2000) and longer legs and smaller muscle circumferences (e.g., calves, arms), then they have a higher center of mass than an individual of the same height. So since Asians and whites have long torsos (i.e., since they are endomorphic), they have a lower centers of mass. Asians have the tallest sitting heights, matched with people of the same height, and so we would expect them to be exceptional swimmers. However, since they are not as tall as whites, they do not set records. Blacks, on the other hand, have a lower sitting height when matched with someone of the same height—3 cm shorter. Whites’ sitting height was lower than Asians, but whites are taller so whites dominate swimming compared to Asians because of their average center of mass. See Table 3 from Bejan, Jones, and Charles (2010).

So the difference between blacks’ and whites’ center of mass is 3 percent. This 3 percent difference can account for why the two races excel in running and swimming. When it comes to the runners (blacks), the 3 percent increase in center of mass translates to a 1.5 percent increase in winning speed for the 100 m dash, and a 1.5 percent decrease in winning time, from 10 s to 9.85 s, for example. So the 3 percent difference in running is a huge advantage for blacks.
When it comes to whites, the same holds, except for swimming. So the 3 percent increase in correct length for whites translates over to a 1.5 increase in winning speed and a 1.5 decrease in winning time.
So for taller athletes, mass that falls from a higher altitude falls faster, down and forward; speed increases with larger physiques. So since blacks have larger physiques than whites, then, at the extremes of elite sports (running), their mass allows them to fall down forward, faster and since they have larger physiques, they are faster. So world records are set by athletes with different centers of mass: black athletes in running and white athletes in swimming.
Shifting away from physics, we will now discuss the cultural/social component. The fact that many blacks do not know how to swim became apparent after the Red River drownings of 2010 (Wiltse, 2014). Wiltse (2014) notes three reasons why blacks may be bad swimmers compared to whites: (1) white swimmers denied blacks access to pools; (2) cities provided few pools to black communities and the pools they did provide were small; and (3) the cities closed many public pools after desegregation occurred. White parents taught their children how to swim, but black parents hardly ever did. As this occurred as swimming became popular in American culture, this could be one reason why blacks aren’t as represented in swimming when compared to whites.
Wiltse’s (2014) argument is that past discrimination to blacks from whites when it came to swimming explains the drowning disparity between the races. Whites passed down their swimming knowledge, whereas blacks had little to no chance to pass theirs down—if they even knew how to swim, that is. This type of cultural transmission could explain most—if not all—of the disparity in drowning between the races. It is simple: to address the disparity, the claim that swimming is “what white people do” needs to be addressed. I would assume that this claim grew from the 60s and desegregation from when blacks were barred from swimming pools, as Wiltse (2014) notes. While the swimming and drowning gap can be closed, the elite sports (running and swimming) gap cannot be—as most of what drives the relationship between race and those sports are anatomic and physiologic in nature, combined with numerous other irreducible variables.
However, pointing to these types of cultural/social causes can be reversed. We can say that since white parents don’t teach their children how to sprint and thus they have not taught their children how to sprint for successive generations, then if white parents did just that then whites would begin to close the gap when it comes to sprinting. While I do not deny that we would have more black swimmers had these types of discriminatory acts had not occurred, it is ridiculous to claim that the two races can and will become equal if this were to occur. It’d be like saying that if we train this person from birth to become an elite sprinter then they would be. Though the right analogy would be that since there are fewer whites than blacks in elite running sports, then what explains the disparity is that they are not trained that way from pretty much conception. However this betrays the systems view of athleticism, and while there are necessary factors in regard to running success, the whole system must be looked at when assessing what makes an elite athlete.
In conclusion, there are many anatomic and physiologic reasons why blacks and whites differ in running and swimming sports. Anatomic differences, such as center of mass, explain the disparities in swimming and running. Blacks’ morphology—long limbs and short torso—is conducive to running success. They can take longer strides and take fewer strides a race compared to someone of the same size that does not have the same limb length. When it comes to white swimmers, where the altitude is set by the body rising out of the water, whites hold a 1.5 percent speed advantage in swimming.
Though there are these anatomic differences that lend themselves to differences in elite sporting competitions, these differences do not lend themselves do the swimming and drowning gap in regard to blacks and whites. What explains those gaps is generational access to swimming pools; blacks were barred access to swimming pools just as they started to become popular in America, after the 60s when the country was desegregated. This led to swimming being looked at as “something that white people do”, and so, fewer and fewer black parents taught their children how to swim. Further cultural and social factors explain this, too. While I would assume that some of these aforementioned factors would then play a role in the black-white swimming/drowning gaps, I doubt that it would count for a super-majority of it. Thus, the gap can be closed by ridding the stigma that swimming is “something that white people do”.
The elite sporting gap in running and swimming, however, cannot be closed.
Athleticism is Irreducible to Biology: A Systems View of Athleticism
1550 words
Reductionists would claim that athletic success comes down to the molecular level. I disagree. Though, of course, understanding the molecular pathways and how and why certain athletes excel in certain sports can and will increase our understanding of elite athleticism, reductionist accounts do not tell the full story. A reductionist (which I used to be, especially in regard to sports; see my article Racial Differences in Muscle Fiber Typing Cause Differences in Elite Sporting Competition) would claim that, as can be seen in my article, the cause for elite athletic success comes down to the molecular level. Now, that I no longer hold such reductionist views in this area does not mean that I deny that there are certain things that make an elite athlete. However, I was wrong to attempt to reduce a complex bio-system and attempt to pinpoint one variable as “the cause” of elite athletic success.
In the book The Genius of All of Us: New Insights into Genetics, Talent, and IQ, David Shenk dispatches with reductionist accounts of athletic success in the 5th chapter of the book. He writes:
2. GENES DON’T DIRECTLY CAUSE TRAITS; THEY ONLY INFLUENCE THE SYSTEM.
Consistent with other lessons of GxE [Genes x Environment], the surprising finding of the $3 billion Human Genome Project is that only in rare instances do specific gene variants directly cause specific traits or diseases. …
As the search for athletic genes continues, therefore, the overwhelming evidence suggests that researchers will instead locate genes prone to certain types of interactions: gene variant A in combination with gene variant B, provoked into expression by X amount of training + Y altitude + Z will to win + a hundred other life variables (coaching, injuries, etc.), will produce some specific result R. What this means, of course, What this means, of course, is that we need to dispense rhetorically with thick firewall between biology (nature) and training (nurture). The reality of GxE assures that each persons genes interacts with his climate, altitude, culture, meals, language, customs and spirituality—everything—to produce unique lifestyle trajectories. Genes play a critical role, but as dynamic instruments, not a fixed blueprint. A seven- or fourteen- or twenty-eight-year-old is not that way merely because of genetic instruction. (Shenk, 2010: 107) [Also read my article Explaining African Running Success Through a Systems View.]
This is looking at the whole entire system: genes, to training, to altitude, to will to win, to numerous other variables that are conducive to athletic success. You can’t pinpoint one variable in the entire system and say that that is the cause: each variable works together in concert to produce the athletic phenotype. One can invoke Noble’s (2012) argument that there is no privileged level of causation in the production of an athletic phenotype. There are just too many factors that go into the production of an elite athlete, and attempting to reduce it to one or a few factors and attempt to look for those factors in regard to elite athleticism is a fool’s errand. So we can say that there is no privileged level of causation in regard to the athletic phenotype.
In his paper Sport and common-sense racial science, Louis (2004: 41) writes:
The analysis and explanation of racial athleticism is therefore irreducible to
biological or socio-cultural determinants and requires a ‘biocultural approach’
(Malina, 1988; Burfoot, 1999; Entine, 2000) or must account for environmental
factors (Himes, 1988; Samson and Yerl`es, 1988).
Reducing anything, sports included, to environmental/socio-cultural determinants and biology doesn’t make sense; I agree with Louis that we need a ‘biocultural approach’, since biology and socio-cultural determinants are linked. This, of course, upends the nature vs. nurture debate; neither “nature” nor “nurture” has won, they causally depend on one another to produce the elite athletic phenotype.
Louis (2004) further writes:
In support of this biocultural approach, Entine (2001) argues that athleticism is
irreducible to biology because it results from the interaction between population-based genetic differences and culture that, in turn, critiques the Cartesian dualism
‘which sees environment and genes as polar-opposite forces’ (p. 305). This
critique draws on the centrality of complexity, plurality and fluidity to social
description and analysis that is significant within multicultural common sense. By
pointing to the biocultural interactivity of racial formation, Entine suggests that
race is irreducible to a single core determinant. This asserts its fundamental
complexity that must be understood as produced through the process of
articulation across social, cultural and biological categories.
Of course, race is irreducible to a single core determinant; but it is a genuine kind in biology, and so, we must understand the social, cultural, and biological causes and how they interact with each other to produce the athletic phenotype. We can look at athlete A and see that he’s black and then look at his somatotype and ascertain that the reason why athlete A is a good athlete is conducive to his biology. Indeed, it is. One needs a requisite morphology in order to succeed in a certain sport, though it is quite clearly not the only variable needed to produce the athletic phenotype.
One prevalent example here is the Kalenjin (see my article Why Do Jamaicans, Kenyans, and Ethiopians Dominate Running Competitions?). There is no core determinant of Kalenjin running success; even one study I cited in my article shows that Germans had a higher level of a physiological variable conducive to long-distance running success compared to the Kalenjin. This is irrelevant due to the systems view of athleticism. Low Kenyan BMI (the lowest in the world), combined with altitude training (they live in higher altitudes and presumably compete in lower altitudes), a meso-ecto somatotype, the will to train, and even running to and from where they have to go all combine to show how and why this small tribe of Kenyans excel so much in these types of long-distance running competitions.
Sure, we can say that what we know about anatomy and physiology that a certain parameter may be “better” or “worse” in the context of the sport in question, no one denies that. What is denied is the claim that athleticism reduces to biology, and it does not reduce to biology because biology, society, and culture all interact and the interaction itself is irreducible; it does not make sense to attempt to partition biology, society, and culture into percentage points in order to say that one variable has primacy over another. This is because each level of the system interacts with every other level. Genes, anatomy and physiology, the individual, the overarching society, cultural norms, peers, and a whole slew of other factors explain athletic success not only in the Kalenjin but in all athletes.
Broos et al (2016) showed that in those with the RR genotype, coupled with the right morphology and fast twitch muscle fibers, this would lead to more explosive contractions. Broos et al (2016) write:
In conclusion, this study shows that a-actinin-3 deficiency decreases the contraction velocity of isolated type IIa muscle fibers. The decreased cross-sectional area of type IIa and IIx fibers may explain the increased muscle volume in RR genotypes. Thus, our results suggest that, rather than fiber force, combined effects of morphological and contractile properties of individual fast muscle fibers attribute to the enhanced performance observed in RR genotypes during explosive contractions.
This shows the interaction between the genotype, morphology, fast twitch fibers (which blacks have more of; Caeser and Henry, 2015), and, of course, the grueling training these elite athletes go through. All of these factors interact. This further buttresses the argument that I am making that different levels of the system causally interact with each other to produce the athletic phenotype.
Pro-athletes also have “extraordinary skills for rapidly learning complex and neutral dynamic visual scenes” (Faubert, 2013). This is yet another part of the system, along with other physical variables, that an elite athlete needs to have. Indeed, as Lippi, Favalaro, and Guidi (2008) write:
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.
So now we have: (1) strong mental background; (2) genes; (3) morphology; (4) Vo2 max; (5) altitude; (6) will to win; (7) training; (8) coaching; (9) injuries; (10) peer/familial support; (11) fiber typing; (12) heart strength etc. There are of course myriad other variables that are conducive to athletic success but are irreducible since we need to look at it in the whole context of the system we are observing.
In conclusion, athleticism is irreducible to biology. Since athleticism is irreducible to biology, then to explain athleticism, we need to look at the whole entire system, from the individual all the way to the society that individual is in (and everything in between) to explain how and why athletic phenotypes develop. There is no logical reason to attempt to reduce athleticism to biology since all of these factors interact. Therefore, the systems view of athleticism is the way we should view the development of athletic phenotypes.
(i) Nature and Nurture interact.
(ii) Since nature and nurture interact, it makes no sense to attempt to reduce anything to one or the other.
(iii) Since it makes no sense to attempt to reduce anything to nature or nurture since nature and nurture interact, then we must dispense with the idea that reductionism can causally explain differences in athleticism between individuals.
Muscle Fibers, Obesity, Cardiometabolic Disorders, and Race
2650 words
The association between muscle fiber typing obesity and race is striking. It is well-established that blacks have a higher proportion of type II skeletal muscle fibers than whites and these higher proportions of these specific types of muscle fibers lead to physiological differences between the two races which then lead to differing health outcomes between them—along with differences in athletic competition. Racial differences in health are no doubt complex, but there are certain differences between the races that we can look at and say that there is a relationship here that warrants further scrutiny.
Why is there an association between negative health outcomes and muscle phsyiology? The answer is very simple if one knows the basics of muscle physiology and how and why muscles contract (it is worth noting that out of a slew of anatomic and phsyiologic factors, movement is the only thing we can consciously control, compare to menstration and other similar physiologic processes which are beyond our control). In this article, I will describe what muscles do, how they are controlled, muscle physiology, the differences in fiber typing between the races and what it means for health outcomes between them.
Muscle anatomy and physiology
Muscle fiber number is determined by the second trimester. Bell (1980) noted that skeletal muscle fiber in 6 year olds is not different from normal adult tissue, and so, we can say that between the time in the womb and age 6, muscle fiber type is set and cannot be changed (though training can change how certain fibers respond, see below).
Muscle anatomy and physiology is interesting because it shows us how and why we move the way we do. Tendons attach muscle to bone. Attached to the tendon is the muscle belly. The muscle belly is made up of facsicles and the fascicles are made up of muscle fibers. Muscle fibers are made up of myofibrils and myofibrils are made up of myofilaments. Finally, myofilaments are made up of proteins—specifically actin and myosin, this is what makes up our muscles.

(Image from here.)
Muscle fibers are encased by sarcolemma which contains cell components such as sarcoplasm, nuclei, and mitochondria. They also have other cells called myofibrils which contain myofilaments which are then made up of actin (thin filaments) and mysoin (thick filaments). These two types of filaments form numerous repeating sections within a myofibril and each repeating section is known as a sarcomere. Sarcomeres are the “functional” unit of the muscle, like the neuron is for the nervous system. Each ‘z-line’ denotes another sarcomere across a myofibril (Franzini-Armstrong, 1973; Luther, 2009).
Other than actin and myosin, there are two more proteins important for muscle contraction: tropomyosin and troponin. Tropomyosin is found on the actin filament and it blocks myosin binding sites which are located on the actin filament, and so it keeps myosin from attaching to muscle while it is in a relaxed state. On the other hand, troponin is also located on the actin filament but troponin’s job is to provide binding sites for calcium and tropomyosin when a muscle needs to contract.
So the structure of skeletal muscle can be broken down like so: epymyseum > muscle belly > perimyseum > fascicle > endomyseum > muscle fibers > myofibrils > myofilaments > myosin and actin. Note diagram (C) from above; the sarcomere is the smallest contractile unit in the myofibril. According to sliding filament theory (see Cook, 2004 for a review), a sarcomere shortens as a result of the ‘z-lines’ moving closer together. The reason these ‘z-lines’ converge is because myosin heads attach to the actin filament which asynchronistically pulls the actin filament across the myosin, which then results in the shortening of the muscle fiber. Sarcomeres are the basic unit controlling changes in muscle length, so the faster or slower they fire depends on the majority type of fiber in that specific area.
But the skeletal muscle will not contract unless the skeletal muscles are stimulated. The nervous system and the muscular system communicate, which is called neural activiation—defined as the contraction of muscle generated by neural stimulation. We have what are called “motor neurons”—neurons located in the CNS (central nervous system) which can send impulses to muscles to move them. This is done through a special synapse called the neuromuscular junction. A motor neuron that connects with muscle fibers is called a motor unit and the point where the muscle fiber and motor unit meet is callled the neuromuscular junction. It is a small gap between the nerve and muscle fiber called a synapse. Action potentials (electrical impulses) are sent down the axon of the motor neuron from the CNS and when the action potential reaches the end of the axon, hormones called neurotransmitters are then released. Neurotransmitters transport the electrical signal from the nerve to the muscle.
Muscle fiber types
The two main categories of muscle fiber are type I and type II—‘slow’ and ‘fast’ twitch, respectively. Type I fibers contain more blood cappilaries, higher levels of mitochondria (which transforms food into ATP) and myoglobin which allows for an improved delivery of oxygen. Since myoglobin is similar to hemoglobin (the red pigment which is found in red blood cells), type I fibers are also known as ‘red fibers.’ Type I fibers are also smaller in diameter and slower to produce maximal tension, but are also the most fatigue-resistant type of fiber.
Type II fibers have two subdivisions—IIa and IIx—based on their mechanical and chemical properties. Type II fibers are in many ways the opposite of type I fibers—they contain far fewer blood cappilaries, mitochondria and myoglobin. Since they have less myoglobin, they are not red, but white, which is why they are known as ‘white fibers.’ IIx fibers have a lower oxidative capacity and thusly tire out quicker. IIa, on the other hand, have a higher oxidative capacity and fatigue slower than IIx fibers (Herbison, Jaweed, and Ditunno, 1982; Tellis et al, 2012). IIa fibers are also known as intermediate fast twitch fibers since they can use both anarobic and aerobic metabolism equally to produce energy. So IIx fibers are a combo of I and II fibers. Type II fibers are bigger, quicker to produce maximal tension, and tire out quicker.
Now, when it comes to fiber typing between the races, blacks have a higher proportion of type II fibers compared to whites who have a higher proportion of type I fibers (Ama et al, 1986; Ceaser and Hunter, 2015; see Entine, 2000 and Epstein, 2014 for reviews). Higher proportions of type I fibers are associated with lower chance of cardiovascular events, whereas type II fibers are associated with a higher risk. Thus, “Skeletal muscle fibre composition may be a mediator of the protective effects of exercise against cardiovascular disease” (Andersen et al, 2015).
Now that the basics of muscle anatomy and physiology are apparent, hopefully the hows and whys of muscle contraction and what different muscle fibers do are becoming clear, because these different fibers are distributed between the races in uneven frequencies, which then leads to differences in sporting performance but also differents in health outcomes.
Muscle fibers and health outcomes
We now know the physiology and anatomy of muscle and muscle fiber typing. We also know the differences between each type of skeletal muscle fiber. Since the two races do indeed differ in the percentage of skeletal muscle fiber possessed on average, we then should find stark differences in health outcomes, part of the reason being these differences in muscle fiber typing.
While blacks on average have a higher proportion of type II muscle fibers, whites have a higher proportion of type I muscle fibers. Noting what I wrote above about the differences between the fiber types, and knowing what we know about racial differences in disease outcomes, we can draw some inferences on how differences in muscle fiber typing between races/individuals can then affect disease seriousness/acquisition.
In their review of black-white differences in muscle fiber typing, Ceaser and Hunter (2015) write that “The longitudinal data regarding the rise in obesity indicates obesity rates have been highest among non-Hispanic Black women and Hispanic women.” And so, knowing what we know about fiber type differences between races and how these fibers act when they fire, we can see how muscle fiber typing would contribute to differences in disease acquisition between groups.
Tanner et al (2001) studied 53 women (n=28, lean women; and n=25, obese women) who were undergoing an elective abdominal surgery (either a hysterectomy or gastric bypass). Their physiologic/anatomic measures were taken and they were divided into races: blacks and whites, along with their obesity status. Tanner et al found that the lean subjects had a higher proportion of type I fibers and a lower proportion of type IIx fibers whereas those who were obese were more likely to have a higher proportion of type IIb muscle fibers.
Like other analyses on this matter, Tanner et al (2001) showed that the black subjects had a higher proportion of type II fibers in comparison to whites who had a higher proportion of type I fibers (adiposity was not taken into account). Fifty-one percent of the fiber typing from whites was type I whereas for blacks it was 43.7 pervent. Blacks had a higher proportion of type IIx fibers than whites (16.3 percent for whites and 23.4 for blacks). Lean blacks and lean whites, though, had a similar percentage of type IIx fibers (13.8 percent for whites and 15 percent for blacks). It is interesting to note that there was no difference in type I fibers between lean whites and blacks (55.1 percent for whites and 54.1 percent for blacks), though muscle fibers from obese blacks contained far fewer type I fibers compared to their white counterparts (48.6 percent for whites and 34.5 for blacks). Obese blacks’ muscle fiber had a higher proportion of type IIx fibers than obese whites’ fiber typing (19.2 percent for whites and 31 percent for blacks). Lean blacks and lean whites had a higher proportion of type I fibers than obese blacks and obese whites. Obese whites and obese blacks had more type IIx fibers than lean whites and lean blacks.
So, since type II fibers are insulin resistant (Jensen et al, 2007), then they should be related to glucose intloerance—type II diabetes—and blacks with ancestry from West Africa should be most affected. Fung (2016, 2018) shows that obesity is a disease of insulin resistance, and so, we can bring that same rationale to racial differences in obesity. Indeed, Nielsen and Christensen (2011) hypothesize that the higher prevalence of glucose intolerance in blacks is related to their lower percentage of type I fibers and their higher percentage of type II fibers.
Nielsen and Christensen (2011) hypothesize that since blacks have a lower percentage of type I fibers (the oxidative type), this explains the lower fat oxidation along with lower resting metabolic rate, sleeping metabolic rate, resting energy expenditure and Vo2 max in comparison to whites. Since type I fibers are more oxidative over the glycolitic type II fibers, the lower oxidative capacity in these fibers “may cause a higher fat storage at lower levels of energy intake than in individuals with a higher oxidative capacity” (Nielsen and Christensen, 2011: 611). Though the ratio of IIx and IIa fibers are extremely plastic and affected by lifestyle, Nielsen and Christensen do note that individuals with different fiber typings had similar oxidative capacity if they engaged in physical activity. Recall back to Caesar and Hunter (2015) who note that blacks have a lower maximal aerobic capacity and higher proportion of type II fibers. They note that lack of physical activity exacerbates the negative effects that a majority type II fibers has over majority type I. And so, some of these differences can be ameliorated between these two racial groups.
The point is, individuals/groups with a higher percentage of type II fibers who do not engage in physical activity have an even higher risk of lower oxidative capacity. Furthermore, a higher proportion of type II fibers implies a higher percentage of IIx fibers, “which are the least oxidative fibres and are positively associated with T2D and obesity” (Nielsen and Christensen, 2011: 612). They also note that this may explain the rural-urban difference in diabetes prevalance, with urban populations having a higher proportion of type II diabetics. They also note that this may explain the difference in type II diabetes in US blacks and West African natives—but the reverse is true for West Africans in the US. There is a higher rate of modernization and, with that, a higher chance to be less physically active and if the individual in question is less physically active and has a higher proportion of type II fibers then they will have a higher chance of acquiring metabolic diseases (obesity is also a metabolic disease). Since whites have a higher proportion of type I fibers, they can increase their fat intake—and with it, their fat oxidation—but this does not hold for blacks who “may not adjust well to changes in fat intake” (Nielsen and Christensen, 2011: 612).
Nielsen and Christensen end their paper writing:
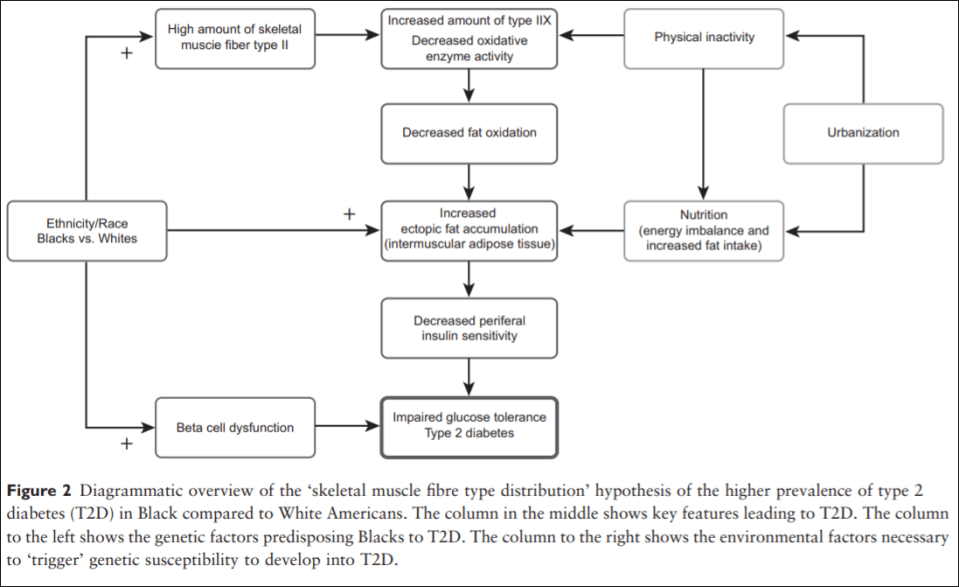
Thus, Blacks of West African ancestry might be genetically predisposed to T2D because of an inherited lower amount of skeletal muscle fibre type I, whereby the oxidative capacity and fat oxidation is reduced, causing increased muscular tissue fat accumulation. This might induce skeletal muscle insulin resistance followed by an induced stress on the insulin-producing beta cells. Together with higher beta-cell dysfunction in the West African Diaspora compared to Whites, this will eventually lead to T2D (an overview of the ‘skeletal muscle distribution hypothesis’ can be seen in Figure 2).
Lambernd et al (2012) show that muscle contractions eliminated insuin resistance by blocking pro-inflammatory signalling pathways: this is the mechanism by which physical activity decreases glucose intolerance and thusly improves health outcomes—especially for those with a higher proportion of type II fibers. Thus, it is important for individuals with type II fibers to exercise, since sedentariness is associated with an age-related insulin resistance due to impaired GLUT4 utilization (Bunprajun et al, 2013).
(Also see Morrison and Cooper’s (2006) hypothesis that “reduced oxygen-carrying capacity induced a shift to more explosive muscle properties” (Epstein, 2014: 179). Epstein notes that the only science there is on this hypothesis is one mouse and rat study showing that low hemoglobin can “induce a switch to more explosive muscle fibers” (Epstein, 2014: 178), but this has not been tested on humans to see if it would hold. If this is tested on humans and if it does hold, then that would lend credence to Morrison’s and Cooper’s (2006) hypothesis.)
Conclusion
Knowing what we know about muscle anatomy and physiology and how muscles act we can understand the influence the different muscle types have on disease and how they contribute to disease variation between race, sex and the individual level. Especially knowing how type II fibers act when the individual in question is insulin resistant is extremely important—though it has been noted that individuals who participate in aerobic exercise decrease their risk for cardiometabolic diseases and can change the fiber distribution difference between IIx and IIa fibers, lowering their risk for acquiring cardiometabolic diseases (Ceaser and Hunter, 2015).
Thinking back to sarcomeres (the smallest contractile unit in the muscle) and how they would act in type II fibers: they would obviously contract much faster in type II muscles over type I muscles; they would then obviously tear faster than type I muscles; since type II muscles are more likely to be insulin resistant, then those with a higher proportion of type II fibers need to focus more on aerobic activity to “balance out” type IIx and IIa fibers and decrease the risk of cardiometabolic disease due to more muscle contractions (Lambernd et al, 2012). Since blacks have a higher proportion of type II fibers and are more likely to be sedentary than whites, and since those who have a higher proportion of type II fibers are more likely to be obese, then it is clear that exercise can and will ameliorate some of the disparity in cardiometabolic diseases between blacks and whites.
Black-White Differences in Anatomy and Physiology: Black Athletic Superiority
3000 words
Due to evolving in different climates, the different races of Man have differing anatomy and physiology. This, then, leads to differences in sports performance—certain races do better than others in certain bouts of athletic prowess, and this is due to, in large part, heritable biological/physical differences between blacks and whites. Some of these differences are differences in somatotype, which bring a considerable advantage for, say, runners (an ecto-meso, for instance, would do very well in sprinting or distance running depending on fiber typing). This article will discuss differences in racial anatomy and physiology (again) and how it leads to disparities in certain sports performance.
Kerr (2010) argues that racial superiority in sport is a myth. (Read my rebuttal here.) In his article, Kerr (2010) attempts to rebut Entine’s (2000) book Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It. In a nutshell, Kerr (2010) argues that race is not a valid category; that other, nongenetic factors play a role other than genetics (I don’t know if anyone has ever argued if it was just genetics). Race is a legitimate biological category, contrary to Kerr’s assertions. Kerr, in my view, strawman’s Entine (2002) by saying he’s a “genetic determinist”, but while he does discuss biological/genetic factors more than environmental ones, Entine is in no way a genetic determinist (at least that’s what I get from my reading of his book, other opinions may differ). Average physical differences between races are enough to delineate racial categories and then it’s only logical to infer that these average physical/physiological differences between the races (that will be reviewed below) would infer an advantage in certain sports over others, while the ultimate cause was the environment that said race’s ancestors evolved in (causing differences in somatotype and physiology).
Black athletic superiority has been discussed for decades. The reasons are numerous and of course, this has even been noticed by the general public. In 1991, half of the respondents of a poll on black vs. whites in sports “agreed with the idea that “blacks have more natural physical ability,“” (Hoberman, 1997: 207). Hoberman (1997) of course denies that there is any evidence that blacks have an advantage over whites in certain sports that come down to heritable biological factors (which he spends the whole book arguing). However, many blacks and whites do, in fact, believe in black athletic superiority and that physiologic and anatomic differences between the races do indeed cause racial differences in sporting performance (Wiggins, 1989). Though Wiggins (1989: 184) writes:
The anthropometric differences found between racial groups are usually nothing more than central tendencies and, in addition, do not take into account wide variations within these groups or the overlap among members of different races. This fact not only negates any reliable physiological comparisons of athletes along racial lines, but makes the whole notion of racially distinctive physiological abilities a moot point.
This is horribly wrong, as will be seen throughout this article.
The different races have, on average, differing somatotypes which means that they have different anatomic proportions (Malina, 1969):
| Data from Malina, (1969: 438) | n | Mesomorph | Ectomorph | Endomorph |
| Blacks | 65 | 5.14 | 2.99 | 2.92 |
| Whites | 199 | 4.29 | 2.89 | 3.86 |
| Data from Malina (1969: 438) | Blacks | Whites |
| Thin-build body type | 8.93 | 5.90 |
| Submedium fatty development | 48.31 | 29.39 |
| Medium fleshiness | 33.69 | 43.63 |
| Fat and very fat categories | 9.09 | 21.06 |
This was in blacks and whites aged 6 to 11. Even at these young ages, it is clear that there are considerable anatomic differences between blacks and whites which then lead to differences in sports performance, contra Wiggins (1989). A basic understanding of anatomy and how the human body works is needed in order to understand how and why blacks dominate certain sports over whites (and vice versa). Somatotype is, of course, predicated on lean mass, fat mass, bone density, stature, etc, which are heritable biological traits, thus, contrary to popular belief that somatotyping holds no explanatory power in sports today (see Hilliard, 2012).
One variable that makes up somatotype is fat-free body mass. There are, of course, racial differences in fat mass, too (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000). Lower fat mass would, of course, impede black excellence in swimming, and this is what we see (Rushton, 1997; Entine, 2000). Wagner and Heyward (2000) write:
Our review unequivocally shows that the FFB of blacks and whites differs significantly. It has been shown from cadaver and in vivo analyses that blacks have a greater BMC and BMD than do whites. These racial differences could substantially affect measures of body density and %BF. According to Lohman (63), a 2% change in the BMC of the body at a given body density could, theoretically, result in an 8% error in the estimation of %BF. Thus, the BMC and BMD of blacks must be considered when %BF is estimated.
While Vickery, Cureton, and Collins (1988) found that blacks had thinner skin folds than whites, however, in this sample, somatotype did not explain racial differences in bone density, like other studies (Malina, 1969), Vickery, Cureton, and Collins (1988) found that blacks were also more likely to be mesomorphic (which would then express itself in racial differences in sports).
Hallinan (1994) surveyed 32 sports science, exercise physiology, biomechanics, motor development, motor learning, and measurement evaluation textbooks to see what they said racial differences in sporting performance and how they explained them. Out of these 32 textbooks, according to Wikipedia, these “textbooks found that seven [textbooks] suggested that there are biophysical differences due to race that might explain differences in sports performance, one [textbook] expressed caution with the idea, and the other 24 [textbooks] did not mention the issue.” Furthermore, Strklaj and Solyali (2010), in their paper “Human Biological Variation in Anatomy Textbooks: The Role of Ancestry” write that their “results suggest that this type of human variation is either not accounted for or approached only superficially and in an outdated manner.”
It’s patently ridiculous that most textbooks on the anatomy and physiology of the human body do not talk about the anatomic and physiologic differences between racial and ethnic groups. Hoberman (1997) also argues the same, that there is no evidence to confirm the existence of black athletic superiority. Of course, many hypotheses have been proposed to explain how and why blacks are at an inherent advantage in sport. Hoberman (1997: 269) discusses one, writing (quoting world record Olympian in the 400-meter dash, Lee Evans):
“We were bred for it [athletic dominance] … Certainly the black people who survived in the slave ships must have contained the highest proportion of the strongest. Then, on the plantations, a strong black man was mated with a strong black woman. We were simply bred for physical qualities.”
While Hoberman (1997: 270-1) also notes:
Finally, by arguing for a cultural rather than a biological interpretation of “race,” Edwards proposed that black athletic superiority results from “a complex of societal conditions” that channels a disproporitionate number of talented blacks into athletic careers.
The fact that blacks were “bred for” athletic dominance is something that gets brought up often but has little (if any) empirical support (aside from just-so stories about white slavemasters breeding their best, biggest and strongest black slaves). The notion that “a complex of societal conditions” (Edwards, 1971: 39) explains black dominance in sports, while it has some explanatory power in regard to how well blacks do in sporting competition, it, of course, does not tell the whole story. Edwards (1978: 39) argues that these complex societal conditions “instill a heightened motivation among black male youths to achieve success in sports; thus, they channel a proportionately greater number of talented black people than whites into sports participation.” While this may, in fact, be true, this does nothing to rebut the point that differences in anatomic and physiologic factors are a driving force in racial differences in sporting performance. However, while these types of environmental/sociological arguments do show us why blacks are over-represented in some sports (because of course motivation to do well in the sport of choice does matter), they do not even discuss differences in anatomy or physiology which would also be affecting the relationship.
For example, one can have all of the athletic gifts in the world, one can be endowed with the best body type and physiology to do well in any type of sport you can imagine. However, if he does not have a strong mind, he will not succeed in the sport. Lippi, Favaloro, and Guidi (2008) write:
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.
Any athlete—no matter their race—needs a strong mental background, for if they don’t, they can have all of the physical gifts in the world, they will not become top-tier athletes in the sport of their choice; advantageous physical factors are imperative for success in differing sports, though myriad variables work in concert to produce the desired effect so you cannot have one without the other. On the other side, one can have a strong mental background and not have the requisite anatomy or physiology needed to succeed in the sport in question, but if he has a stronger mind than the individual with the requisite morphology, then he probably will win in a head-to-head competition. Either way, a strong mind is needed for strong performance in anything we do in life, and sport is no different.
Echoing what Hoberman (1997) writes, that “racist” thoughts of black superiority in part cause their success in sport, Sheldon, Jayaratne, and Petty (2007) predicted that white Americans’ beliefs in black athletic superiority would coincide with prejudice and negative stereotyping of black’s “intelligence” and work ethic. They studied 600 white men and women to ascertain their beliefs on black athletic superiority and the causes for it. Sheldon, Jayaratne, and Petty (2007: 45) discuss how it was believed by many, that there is a “ perceived inverse relationship between athleticism and intelligence (and hard work).” (JP Rushton was a big proponent of this hypothesis; see Rushton, 1997. It should also be noted that both Rushton, 1997 and Entine, 2000 believe that blacks’ higher rate of testosterone—3 to 15 percent— [Ross et al, 1986; Ellis and Nyborg, 1992; see rebuttal of both papers] causes their superior athletic performance, I have convincingly shown that they do not have higher levels of testosterone than other races, and if they do the difference is negligible.) However, in his book The Sports Gene: Inside the Science of Extraordinary Athletic Performance, Epstein (2014) writes:
With that stigma in mind [that there is an inverse relationship between “intelligence” and athletic performance], perhaps the most important writing Cooper did in Black Superman was his methodological evisceration of any supposed inverse link between physical and mental prowess. “The concept that physical superiority could somehow be a symptom of intellectual superiority became associated with African Americans … That association did not begin until about 1936.”
What Cooper (2004) implied is that there was no “inverse relationship” with intelligence and athletic ability until Jesse Owens blew away the competition at the 1936 Olympics in Berlin, Germany. In fact, the relationship between “intelligence” and athletic ability is positive (Heppe et al, 2016). Cooper is also a co-author of a paper Some Bio-Medical Mechanisms in Athletic Prowess with Morrison (Morrison and Cooper, 2006) where they argue—convincingly—that the “mutation appears to have triggered a series of physiological adjustments, which have had favourable athletic consequences.”
Thus, the hypothesis claims that differences in glucose conversion rates between West African blacks and her descendants began, but did not end with the sickling of the hemoglobin molecule, where valine is substituted for glutamic acid, which is the sixth amino acid of the beta chain of the hemoglobin molecule. Marlin et al (2007: 624) showed that male athletes who were inflicted with the sickle cell trait (SCT) “are able to perform sprints and brief exercises at the highest levels.” This is more evidence for Morrison and Cooper’s (2006) hypothesis on the evolution of muscle fiber typing in West African blacks.
Bejan, Jones, and Charles (2010) explain that the phenomenon of whites being faster swimmers in comparison to blacks being faster runners can be accounted for by physics. Since locomotion is a “falling-forward cycle“, body mass falls forward and then rises again, so mass that falls from a higher altitude falls faster and forward. The altitude is set by the position of center of mass above the ground for running, while for swimming it is set by the body rising out of the water. Blacks have a center of gravity that is about 3 percent higher than whites, which implies that blacks have a 1.5 percent speed advantage in running whereas whites have a 1.5 percent speed advantage in swimming. In the case of Asians, when all races were matched for height, Asians fared even better, than whites in swimming, but they do not set world records because they are not as tall as whites (Bejan, Jones, and Charles, 2010).
It has been proposed that stereotype threat is part of the reasons for East African running success (Baker and Horton, 2003). They state that many theories have been proposed to explain black African running success—from genetic theories to environmental determinism (the notion that physiologic adaptations to climate, too, drive differences in sporting competition). Baker and Horton (2003) note that “that young athletes have internalised these stereotypes and are choosing sport participation accordingly. He speculates that this is the reason why white running times in certain events have actually decreased over the past few years; whites are opting out of some sports based on perceived genetic inferiority.” While this may be true, this wouldn’t matter, as people gravitate toward what they are naturally good at—and what dictates that is their mind, anatomy, and physiology. They pretty much argue that stereotype threat is a cause of East African running performance on the basis of two assertions: (1) that East African runners are so good that it’s pointless to attempt to win if you are not East African and (2) since East Africans are so good, fewer people will try out and will continue the illusion that East Africans would dominate in middle- and long-distance running. However, while this view is plausible, there is little data to back the arguments.
To explain African running success, we must do it through a systems view—not one of reductionism (i.e., gene-finding). We need to see how the systems in question interact with every part. So while Jamaicans, Kenyans, and Ethiopians (and American blacks) do dominate in running competitions, attempting to “find genes” that account for success n these sports seems like a moot point—since the whole system is what matters, not what we can reduce the system in question to.
However, there are some competitions that blacks do not do so well in, and it is hardly discussed—if at all—by any author that I have read on this matter. Blacks are highly under-represented in strength sports and strongman competitions. Why? My explanation is simple: the causes for their superiority in sprinting and distance running (along with what makes them successful at baseball, football, and basketball) impedes them from doing well in strength and strongman competitions. It’s worth noting that no black man has ever won the World’s Strongest Man competition (indeed the only African country to even place—Rhodesia—was won by a white man) and the causes for these disparities come down to racial differences in anatomy and physiology.
I discussed racial differences in the big four lifts and how racial differences in anatomy and physiology would contribute to how well said race performed on the lift in question. I concluded that Europeans and Asians had more of an advantage over blacks in these lifts, and the reasons were due to inherent differences in anatomy and physiology. One major cause is also the differing muscle fiber typing distribution between the races (Alma et al, 1986; Tanner et al, 2002; Caesar and Henry, 2015 while blacks’ fiber typing helps them in short-distance sprinting (Zierath and Hawley, 2003). Muscle fiber typing is a huge cause of black athletic dominance (and non-dominance). Blacks are not stronger than whites, contrary to popular belief.
I also argued that Neanderthals were stronger than Homo sapiens, which then had implications for racial differences in strength (and sports). Neanderthals had a wider pelvis than our species since they evolved in colder climes (at the time) (Gruss and Schmidt, 2016). With a wider pelvis and shorter body than Homo sapiens, they were able to generate more power. I then implied that the current differences in strength and running we see between blacks and whites can be used for Neanderthals and Homo sapiens, thusly, evolution in differing climates lead to differences in somatotype, which eventually then lead to differences in sporting competition (what Baker and Horton, 2003 term “environmental determinism” which I will discuss in the context of racial differences in sports in the future).
Finally, blacks dominate the sport of bodybuilding, with Phil Heath dominating the competition for the past 7 years. Blacks dominate bodybuilding because, as noted above, blacks have thinner skin folds than whites, so their striations in their muscles would be more prevalent, on average, at the same exact %BF. Bodybuilders and weightlifters were similar in mesomorphy, but the bodybuilders showed more musculature than the bodybuilders whereas the weightlifters showed higher levels of body fat with a significant difference observed between bodybuilders and weightlifters in regard to endomorphy and ectomorphy (weightlifters skewing endo, bodybuilders skewing ecto, as I have argued in the past; Imran et al, 2011).
To conclude, blacks do dominate American sporting competition, and while much ink has been spilled arguing that cultural and social—not genetic or biologic—factors can explain black athletic superiority, they clearly work in concert with a strong mind to produce the athletic phenotype, no one factor has prominence over the other; though, above all, if one does not have the right mindset for the sport in question, they will not succeed. A complex array of factors is the cause of black athletic dominance, including muscle fibers, the type of mindset, anatomy, overall physiology and fat mass (among other variables) explain the hows and whys of black athletic superiority. Cultural and social explanations—on their own—do not tell the whole story, just as genetic/biologic explanations on their own would not either. Every aspect—including the historical—needs to be looked at when discussing the dominance (or lack thereof) in certain sports along with genetic and nongenetic factors to see how and why certain races and ethnies excel in certain sports.
The Evolution of Racial Pelvis Variation
1850 words
It’s well-known that blacks have narrower hips than whites (Rushton, 1997; Handa et al, 2008). These pelvic differences then account for part of the variation in elite sporting events such as sprinting and jumping (Entine, 2000). These pelvic differences are the result of climatic variation and sexual selection.
The evolution of the pelvis is due to bipedalism. We are bipeds because of our S-shaped spine, which helps us to cope with differing loads. The human pelvis had to evolve in two ways—to make birthing babies easier and to become more efficient for bipedal walking. Termed the ‘obstetric dilemma’, it has implications for osteoarthritis in both men and women (Hogervorst, Heinse, and de Vos, 2009). Having a more efficient bipedal gait meant the body could allocate energy to other parts of the body—mainly our growing brains/neuronal count. Over time, the brain grew while the pelvis had to shrink for more efficient bipedalism. The pelvis also got narrower in our evolution, being wider in Australopithicenes, while becoming more narrow when erectus appeared—which is the first instance of a humanlike pelvis in the fossil record—which increased how far we could travel as well as reduce our energy expenditure (Lieberman, et al, 2006). Further discussion can be found in my article Man the Athlete.
So we began evolving a narrower pelvis in comparison to our ancestors because it was more efficient for heat dissipation. Smaller trunks are more efficient for heat dissipation (Lieberman, 2015), whereas wider trunks are more efficient for thermoregulation in colder climes (Weaver and Hublin, 2008; Weaver, 2009; Gruss and Schmidt, 2015). Now, simply applying this logic to Eurasians and Africans (I am grouping East Asians and Europeans together since they were a single breeding population up until about 23,000-6,500ya), we can see one reason why that population has wider pelves than Africans.
When anatomically modern humans (AMH) left Africa between 50-100kya, human skeletal morphology was just like modern-day Africans’ today. When Man migrated into northerly climes, however, a wider pelvis was needed to retain heat in colder climes (Gruss and Schmidt, 2015). So, along with a wider pelvis evolving due to climatic demands on the body, as we migrated north the human brain expanded due to the climate of the area, along with expanding the pelvis to better thermoregulate (which a bigger brain also does in northerly climes). I did argue two months back (and added to Skoyles’ (1999) theory) that brain size increased for expertise capacity and not IQ since Arctic people needed more tools, as well as tools that were more complex, in comparison to peoples who evolved in a hotter climate. So selection then occurred for larger brains and pelvis due to the demand for thermoregulation and bigger brains—which then led to earlier births and more helpless babes, which higher levels of intelligence were then needed to care for them (Piantadosi and Kidd, 2016). The helplessness of infants predicts the intelligence of adults in the primate genera (Piantadosi and Kidd, 2016), so I will assume that this holds within primate species as well (I am not able to locate a citation that this doesn’t hold within the primate genera; if I am in error, please provide a citation). Since African children are born earlier and more mature than Eurasian children who are born slightly later and more helpless/less developed, this is one reason why Eurasians have higher levels of intelligence than Africans (which is independent of any direct effects of climate I may add!).
So since Eurasians needed a larger brains to make more tools in the Arctic/colder climes, their brains needed to expand in size for increased expertise capacity, which would then have further selected for wider pelves in Eurasian women. Climatic variation caused the wider hips/bigger brains in Eurasians, which then allowed the evolution of larger brains in comparison to those who remained in Africa.
Finally, the obstetric dilemma has been recently called into question; there is evidence that a wider pelvis does not increase locomotor costs in humans (Warrener et al, 2015), a treadmill tracked their gait, as well as the motion of their pelvis. This study is used as evidence that the obstetric dilemma is wrong—they argue that there is no trade-off between narrower hips in men and wider hips in women. However, as the authors point out, all subjects in the study walked/ran at the same speed. Let’s say that the speed was heightened; do you think the women/men with wider pelves would have had the same locomotor costs as the men/women with narrower pelves? The answer is, obviously, no.
The pelvis of all of the races of Man has evolved the way they are due to environmental/climatic demands. A wider pelvis is better for thermoregulation in colder climates, while a narrower pelvis/body is more efficient for heat loss (Gruss and Schmidt, 2015).
Thus, we can look at the evolution of brain size/pelvic size in a few ways: 1) The amount of tools/complexity of the tools in the area that led to a need for an increase in brain size for more ‘chunks’ (Gobet and Simon, 1998), which then—along with colder climates—selected for larger brains and a wider body/pelvis which made birthing babes with large heads/brains easier along with helping to conserve heat due to the wider body (Gruss and Schmidt, 2015); 2) Since people in higher altitudes needed a high amount of expertise to survive, further selection for bigger brains, wider pelves occurred because of this; 3) Africans have smaller pelves in comparison to Eurasians because they evolved in hotter climes and didn’t have the amount of tools that peoples in more northerly climes did—which also increased brain size; 4) putting this all together, we can say that because Africans live in hotter climates, they need narrow pelves in order to lose body heat; Eurasians, after they migrated into more northerly climes, needed a wider body/pelvis in order to retain heat. When Man migrated north, he needed the ability to become an expert in, say, tool-making and thus needed a bigger brain for more informational chunks (Simon and Gobet, 1998; Skoyles, 1999). Due to this, Eurasians have wider pelves since they needed larger brains for a higher expertise capacity (Skoyles, 1999).
When Man migrated north, he needed the ability to become an expert in, say, tool-making and thus needed a bigger brain for more informational chunks (Simon and Gobet, 1998; Skoyles, 1999). Due to this, Eurasians have wider pelves than Africans; so they can birth larger-brained children. The width of the female pelvis, too, was shaped by sexual selection (Lassek and Gaulin, 2009). Therefore, the evolution of the modern pelvis in human populations comes down to climatic variation, which, in turn, affects how large of a brain the babe is able to have. Climate constrains brain size in either ‘direction’, big or small. We don’t even need to look at the variation within modern Homo sapiens to see the pattern in pelvic size we do today; because the pelvic differences noted among Man definitely were in effect millions of years ago, with hominids in colder climates having wider pelves while hominids in warmer climates had narrower pelves.
Along with everything above, the evolution of the human pelvis has a few implications for the human races today. Some recent studies have shown that there is no obstetric dilemma at all, with birth complications being caused by babies with higher weights than in our ancestral past, due to environmental mismatches causing higher-weight babies (Warrener et al, 2015; Betti, 2017), which was also beneficial for the evolution of our large brains (Cunnane and Crawford, 2003) with the largest amount of cortical neurons in the animal kingdom. However, marked differences in locomotion would be seen in people who had wide pelves compared to narrow pelves; which is what we see in elite running competitions: the elite runners have narrower pelves. So wider pelves don’t impede normal bipedal walking, but it does impede being able to efficiently run, as evidenced in participants of elite sprinting and marathon competitions. Looking at champion athletes and studying their locomotion (along with other traits as I’ve covered here) you can see that those with narrower pelves win more competitions than those with wider pelves (and happen to have different muscle fiber competition, fat distribution/percent, and morphology).
Racial differences in the pelvis explain the reasons behind why a certain race dominates in certain elite competitions; it largely comes down to skeletal morphology. These skeletal differences have evolutionary underpinnings, with the same pelvic differences seen in hominins that evolved in colder/warmer climates in the past. These pelvic differences (along with body fat percentage/distribution, musculoskeletal morphology, muscle fiber type, lean mass percentage, lower Vo2 max, poorer running economy, a larger Q-angle [4.6 degrees greater than men], etc) are why women are less efficient runners. People with wider hips are more likely to have be endomorphic while people with narrower hips are more likely to be ecto and meso. Not surprisingly, people from northerly climes consistently win WSM competitions whereas East and West Africans dominate bodybuilding and sprinting/marathons due to having a narrower pelvis and other advantageous morphological traits that lead to success in the sport. Nevertheless, pelvic differences between the races largely come down to differences in climate, which was also seen in ancient hominins. These pelvic differences further lead to racial differences in elite sporting competition.
References
Betti, L. (2017). Human Variation in Pelvic Shape and the Effects of Climate and Past Population History. The Anatomical Record,300(4), 687-697. doi:10.1002/ar.23542
Cunnane, S. C., & Crawford, M. A. (2003). Survival of the fattest: fat babies were the key to evolution of the large human brain. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology,136(1), 17-26. doi:10.1016/s1095-6433(03)00048-5
Dr. John R. Skoyles (1999) HUMAN EVOLUTION EXPANDED BRAINS TO INCREASE EXPERTISE CAPACITY, NOT IQ. Psycoloquy: 10(002) brain expertise
Entine, J. (2000). Taboo: why Black athletes dominate sports and why we are afraid to talk about it. New York: PublicAffairs.
Gobet, F., & Simon, H. A. (1998). Expert Chess Memory: Revisiting the Chunking Hypothesis. Memory,6(3), 225-255. doi:10.1080/741942359
Gruss, L. T., & Schmitt, D. (2015). The evolution of the human pelvis: changing adaptations to bipedalism, obstetrics and thermoregulation. Philosophical Transactions of the Royal Society B: Biological Sciences,370(1663), 20140063-20140063. doi:10.1098/rstb.2014.0063
Hogervorst, T., Heinse W.B., & de Vos J., (2009) Evolution of the hip and pelvis. Acta Orthopaedica, 80:sup336, 1-39, DOI: 10.1080/17453690610046620
Lieberman, D. E., Raichlen, D. A., Pontzer, H., Bramble, D. M., & Cutright-Smith, E. (2006). The human gluteus maximus and its role in running. Journal of Experimental Biology,209(11), 2143-2155. doi:10.1242/jeb.02255
Lieberman, D. E. (2015). Human Locomotion and Heat Loss: An Evolutionary Perspective. Comprehensive Physiology, 99-117. doi:10.1002/cphy.c140011
Piantadosi, S. T., & Kidd, C. (2016). Extraordinary intelligence and the care of infants. Proceedings of the National Academy of Sciences,113(25), 6874-6879. doi:10.1073/pnas.1506752113
Rushton J P (1997). Race, Evolution, and Behavior. A Life History Perspective (Transaction, New Brunswick, London).
Handa, V. L., Lockhart, M. E., Fielding, J. R., Bradley, C. S., Brubakery, L., Cundiffy, G. W., … Richter, H. E. (2008). Racial Differences in Pelvic Anatomy by Magnetic Resonance Imaging. Obstetrics and Gynecology, 111(4), 914–920.
Warrener, A. G., Lewton, K. L., Pontzer, H., & Lieberman, D. E. (2015). A Wider Pelvis Does Not Increase Locomotor Cost in Humans, with Implications for the Evolution of Childbirth. PLoS ONE, 10(3), e0118903.
Weaver, T. D., & Hublin, J. (2009). Neandertal birth canal shape and the evolution of human childbirth. Proceedings of the National Academy of Sciences,106(20), 8151-8156. doi:10.1073/pnas.0812554106
Weaver, T. D. (2009). The meaning of Neandertal skeletal morphology. Proceedings of the National Academy of Sciences,106(38), 16028-16033. doi:10.1073/pnas.0903864106
Expounding On My Theory for Racial Differences In Sports
2300 words
In the past, I’ve talked about why the races differ—at the extremes—(and the general population, but the extremes put the picture into focus) in terms of what sports they compete and do best in. These differences come down to morphology somatype, physiology. People readily admit racial differences in sports and—rightly say—that these differences are largely genetic in nature. Why is it easier for people to accept racial differences in sports and not accept other truisms, like racial IQ differences?
I’ve muscle fiber typing and how the variances in fiber typing dictate which race/ethny performs best at which sport. I’ve also further evidence that blacks have type II fibers (responsible for explosive power), which leads to a reduced Vo2 max. This lends yet more credence to my theory of racial differences in sports—black Americans (West African descendants) have the fiber typing that is associated with explosive power, less so with endurance activities. Since I’ve documented evidence on the differences in sports such as baseball, football, swimming and jumping, bodybuilding, and finally strength sports, tonight I will talk about the evolutionary reasons for muscle fiber and somatype differences that will have us better understand the evolutionary conditions in which these traits evolved and why they got selected for.
Most WSM winners are from Nordic countries or have Nordic ancestry. There’s a higher amount of slow twitch fibers in Nordics and East Asians (and Kenyans) which is more conducive to strength and less conducive to ‘explosive’ sports. West Africans and their descendants dominate in sprinting competitions. Yea yea yea white guy won in 1960. So they will be less likely to be in strength comps and more likely to win BBing and physique comps. This is what we see.
Only three African countries have placed in the top ten in the WSM (Kenya, Namibia, and Nigeria, however, one competitor from Namibia I was able to find has European ancestry). Here is a video of a Nigerian Strongest Man competition (notice how his chest isn’t up and his hips rise before the bar reaches his knees, horrible form). Fadesere Oluwatofunmi is Nigeria’s Strongest Man, competing in the prestigious Arnold Classic, representing Nigeria. However, men such as Fadesere Olutaofunmi are outliers.
Now for a brief primer on muscle fibers and which pathways they fire through. Understanding this aspect of the human body is paramount to understanding how and why the races differ in elite competition.
Life history and muscle fiber typing
Slow twitch fibers fire through aerobic pathways. Breaking down fats and proteins takes longer through aerobic respiration. Moreover, in cold temperatures, the body switches from burning fat to burning carbohydrates for energy, which will be broken down slower due to the aerobic perspiration. Slow twitch (Type I) fibers fire through aerobic pathways and don’t tire nearly as quickly as type II fibers. Also, CHO reserves will be used more in cold weather. The body’s ability to use oxygen decreases in cold weather as well, so having slow twitch fibers is ideal (think about evolution thousands of years ago). Type I fibers fire off slower so they’ll be able to be more active for a longer period of time (studies have shown that Africans with Type II fibers hit a ‘wall’ after 40 seconds of explosive activity, which is why there are so few West-African descended marathon runners, powerlifters, Strongmen, etc). Those who possess these traits will have a higher chance to survive in these environments. Those with slow twitch fibers also have to use more oxygen. They have larger blood vessels, more mitochondria and higher concentrations of myoglobin which gives the muscles its reddish color.
Each fiber fires off through different pathways, whether they be anaerobic or aerobic. The body uses two types of energy systems, aerobic or anaerobic, which then generate Adenosine Triphosphate, better known as ATP, which causes the muscles to contract or relax. Depending on the type of fibers an individual has dictates which pathway muscles use to contract which then, ultimately, dictate if there is high muscular endurance or if the fibers will fire off faster for more speed.
Type I fibers lead to more strength and muscular endurance as they are slow to fire off, while Type II fibers fire quicker and tire faster. Slow twitch fibers use oxygen more efficiently, while fast twitch fibers do not burn oxygen to create energy. Slow twitch muscles delay firing which is why the endurance is so high in individuals with these fibers whereas for those with fast twitch fibers have their muscles fire more explosively. Slow twitch fibers don’t tire as easily while fast twitch fibers tire quickly. This is why West Africans and their descendants dominate in sprinting and other competitions where fast twitch muscle fibers dominate in comparison to slow twitch.
Paleolithic Europeans who had more stamina spread more of their genes to the next generation as their genotype was conducive to reproductive success in Ice Age Europe. Conversely in Africa, those who could get away from predators and could hunt prey more efficiently survived. Over time, frequencies of genes related to what needed to be done to survive in those environments increased, along with the frequencies of muscle fibers in the races.
Racial differences in anatomy and physiology
Along with muscle fiber differences, blacks and whites also have differences in fat-free body mass, bone density, distribution of subcutaneous fat, length of limbs relative to the trunk, and body protein contents (Wagner and Heyward, 2000). These differences are noticed and talked about in the scientific literature, even in University biology and anatomy textbooks. However, in terms of University textbooks, authors who recognize the concept of race do so in spite of what other authors write (Hallinan, 1994). Furthermore, Strkalj and Solyali (2010) analyzed 18 English textbooks on anatomy and concluded that discussion of race was ‘superficial’ and the content ‘outdated’, i.e., using the ‘Mongoloid, Caucasoid, Negroid terminology (I still do out of habit). They conclude that most mentions of race are either not mentioned in anatomy textbooks or are only ‘superficially accounted for in textbooks’. Clearly, though they are outdated, some textbooks do talk about human biological diversity, though the information needs to be updated (especially now). The center of mass in blacks is 3 percent higher than in whites, meaning whites have a 1.5 percent speed advantage in swimming while blacks have a 1.5 percent speed difference in sprinting. East Asians that are the same height as whites are even more favored in swimming, however, they are shorter on average so that’s why they do not set world records (Bejan, Jones, and Charles, 2010).
For another hand grip strength (HGP) test, see Leong et al (2016). Most studies on HGS are done on Caucasian populations with little data for non-Caucasoid populations. They found that HGS values were higher for North America and Europe, intermediate in China, South America and the Middle East, and the lowest in South Asia, Southeast Asia, and Africa. These are, most likely, average Joes and not elite BBers or strength trainers. This is one of the best papers I’ve come across on this matter (though I would kill to have a nice study done on the three classical races and their performance on the Big 3 lifts: squat, bench press and deadlift; I’m positive it would be Asian/white and blacks as the tail end).
Among other physical differences is brain size and hip width. Blacks have narrower hips than whites who have narrower hips than Asians (Rushton, 1997: 163). Bigger-brained babes are more than likely born to women who have wider hips. If you think about athletic competition, one with wide hips will get decimated by one with narrower hips as he will be better able to move. People with big crania, in turn, have the hip structure to birth big brains. This causes further division between racial groupings in sports competition.
Some people may dispute a genetic causation and attribute the success of, say, Kenyan marathoners (the Kalenjin people) and attribute the effects to the immediate environment (not ancestral), training and willpower (see here for discussion). This Kenyan subpopulation also has the morphology conducive to success in marathons (tall and lengthy), as well as type II muscle fibers (which is why Kenya placed in the WSM).
I would also like to see a study of men in their prime (age 21 to 24) from European, Africans, and East Asian backgrounds with a high n (for a study like this it’d be 20 to 30 participants for each race), with good controls. You’d see, in my opinion, East Asians slightly nudge out whites who destroy blacks. The opposite would occur in sports that use type II fibers. West Africans also have the gene variant ACTN3 which is associated with explosive sports.
For a better (less ethical study) we can use a thought experiment.
We take two babes fresh out of the womb (one European, the other West African) and keep them locked in a metabolic chamber for their whole entire lives. We keep them in the chamber until they die and monitor them, feeding them the same exact diet and giving them the same amount of attention. They start training at age 15. Who will be stronger in their prime (the European man)? Who will have more explosive power (the West African man)? A simple thought experiment can have one thinking about intrinsic racial differences in things the average American watches in their everyday lives. The subject of racial differences in sports is a non-taboo subject, however, the subject of racial differences in intelligence is a big no-no.
Think about that for a second. People obviously accept racial differences in sports, yet they have some kind of emotional attachment to the blank slate narrative. We don’t hear that you can nurture athletic success. We do, however, hear that ‘we can succeed at anything we put out minds to’. That’s not in dispute; that’s a fact. But it’s twisted in a way that genetics and ancestry has no bearing on it, when it explains a lot of the variance. People accept racial differences when they’re cheering on their favorite football team or basketball team. For instance, NFL announcer Gus Johnson said during a broadcast of a Titans and Jaguars game “He’s [Chris Johnson] got gettin’ away from the cops speed!”
Pro-sports announcers, as well as college recruiters, know what the average person doesn’t who is not exposed to these differences daily for decades on end. People in these types of professions, especially collegiate sports recruiters, must get the low-down on average racial differences and then use what they know to make their choices on who to draft for their team.
For more (anecdotal) evidence, you can look up the race/ethnicity of the winners in competitions where peoples from all over the world compete in. More West African descendants place higher in physique, BBing comps, etc; more Caucasians and East Asians (and Kenyans) place higher in strength comps. A white man has won the WSM every year since its inception. West African descended blacks dominate BBing and physique comps. Eurasians (and Kenyans) dominate in marathon running.
All of this talk of racial differences in sports (which largely has to do with whites vs. blacks, though Asians are included in my overall analysis), I’ve hardly cited anything on East Asians directly. In regards to sports that take extreme dexterity or flexibility (and high reaction), East Asians shine. They shine in diving, ping-pong, skating and gymnastics events. They usually have long torsos and small limbs. I theorize that this was an evolutionary adaptation for the East Asians, as shorter people have less surface area to keep warm. Taller people would have died out quicker than one who’s smaller and can cover up and get warmer faster. They also have quicker reaction times (Rushton and Jensen, 2005) and it has been hypothesized that this is why they dominate in ping pong.
Conclusion
We don’t need any tests to show that there are racial differences in sports; the test is the competition itself. On average, A white will be stronger than an Asian who will be stronger than a black. Conversely, a Kenyan will be a better marathoner than a West African, European or Asian. West Africans will be more likely to beat all three groups in a sprint. These differences come down to morphology, but they start inside the muscles with the muscle fibers. Some anatomy textbooks acknowledge the existence of race, however, they have old and outdated information. It’s a good thing that anatomy textbooks talk about racial differences in physiology and anatomy, now we need to start doing heavy research into racial differences in the brain. The races evolved their fiber typing depending on what they had to do to survive along with their immediate environment, i.e., high elevation like the Kalenjin people.
The evolution of differing muscle fiber types in different races is easily explainable: Europeans have slow twitch fibers. In cold temperatures, the body switches from burning fat to burning carbs for energy. Furthermore, the average person would need to have a higher lung capacity and not tire out during long hunts on megafauna. Over time, selection occurred on the individuals with more type I fibers. Conversely, West Africans and their descendants have the ACTN3 gene, which is associated with elite human athletic performance (Yang et al, 2003). Africans who could get away from predators survived and passed on the genes and fiber typing for elite athletic performance.
In sum, the races differ in terms of entrants to elite athletic competition. These differences are largely genetic in nature. Evolutionary processes explain racial differences in sports. These same selection processes that explain racial differences in elite sports competitions also explain racial differences in intelligence. I await the day that we can freely speak about racial differences in intelligence just like we speak about racial differences in sports. Denying human variation makes no sense, especially in today’s world where we have state of the art testing.
