
Home » Posts tagged 'physiology'
Tag Archives: physiology
Racism, Stress, and Physiology
1800 words
The term ‘racism’ has many definitions. What does it mean for a person to be a ‘racist’? What does it mean for a person to have ‘racist beliefs’? What does the term ‘racism’ refer to? The answers to these questions then will inform the next part—what does racism have to do with stress and physiology?
What is ‘racism’?
Racism has many definitions, so many—and so many for uses in different contexts—that it has been argued, for example by those in the far-right, that it is, therefore, a meaningless term. However, just because there are many definitions of the term, it does not then mean that there is no referent for the term we use. A referent is a thing that is signified. In this instance, what is the referent for racism? I will provide a few on-hand definitions and then discuss them.
In Part VI of The Oxford Handbook of Race and Philosophy (edited by Naomi Zack, 2016) titled Racisms and Neo-Racisms, Zack writes (pg 469; my emphasis):
Logically, it would seem as though ideas about race would have to precede racism. But the subject of racism is more broad and complicated than the subject of race, for at least these two historical reasons. First, the kind of prejudice (prejudged cognitions and negative emotions) and discrimination (treating people differently on the grounds of group identities) that constitute racism have a longer history than the modern idea of race, for instance in European anti-Semitism. And second, insofar as modern ideas of race have been in the service of the dominant interests in international and internal interactions, these ideas of race are ideologies that have devalued non-white groups. That is, ideas of race are themselves already inherently racist.
In philosophy, racism has been treated as attitudes and actions of individuals that affect nonwhites unjustly and social structures and institutions that advantage whites and disadvantage nonwhites. The first is hearts-and-minds or classic racism, for instance the use of stereotypes and harmful actions by whites against people of color, as well as negative feelings about them. The second is structural racism, for instance the use of stereotypes or institutional racism, for instance, the facts of how American blacks and Hispanics are, compared to whites, worse off on major measures of human well-being, such as education, income, family wealth, health, family stability, longevity, and rates of incarceration.
John Lovchik in his book Racism: Reality Built on a Myth (2018: 12) notes that “racism is a system of ranking human beings for the purpose of gaining and justifying an unequal distribution of political and economic power.” Note that using this definition, “hereditarianism” (the theory that individual differences between groups and individuals can be reduced to genes; I will give conceptual reasons why hereditarianism is false as what I hope is my final word on the debate) is a racist theory as it attempts to justify the current social hierarchy. (The reason why IQ tests were first brought to America and created by Binet and Simon; see The History and Construction of IQ Tests and The Frivolousness of the Hereditarian-Environmentalist IQ Debate: Gould, Binet, and the Utility of IQ Testing.) This is why hereditarianism saw its resurgence with Jensen’s infamous 1969 paper. Indeed, many prominent hereditarians have held racist beliefs, and were even eugenicists espousing eugenic ideas.
Headley (2000) notes a few definitions of racism—motivational, behavioral, and cognitive racism. Motivational racism is “the infliction of unequal consideration, motivated by the desire to dominate, based on race alone“; behavioral racism is “failure to give equal consideration, based on the fact of race alone”; and cognitive racism is “unequal consideration, out of a belief in the inferiority of another race.”
I have presented six definitions of racism—though there are many more. Now, for the purposes of this article, I will present my own: the ‘inferiorization’ of a racialized group which is then used to explain disparities in things like IQ test scores, social class/SES, education differences, personality, etc. Now, knowing what we know about physiological systems and how they react to the environment around them—the immediate environment and the social environment—how does this then relate to stress and physiology?
Racism, stress, and physiology
Now that we know what racism is, having had a rundown of certain definitions of ‘racism’, I will now discuss the physiological effects such stances could have on groups racialized as ‘races’ (note that I am using socialraces in this article; recall that social constructivists about race need to be realists about race).
The term ‘weathering’ refers to the body’s breaking down due to stress over time. Such stressors can come from one’s immediate environment (i.e., pollution, foodstuffs, etc) or their social environment (a demanding job, how one perceives themselves and how people react to them). So as the body experiences more and more stress it becomes more and more ‘weathered’ which then leads to heightened risk for disease in stressed individuals/populations.
Allostatic states “refer to altered and sustained activity levels of the primary mediators (e.g., glucocorticosteroids) that integrate energetic and associated behaviours in response to changing environments and challenges such as social interactions, weather, disease, predators and pollution” (McEwen, 2005). Examples of allostatic overload such as acceleration of atherosclerosis, hypertension (HTN), stroke, and abdominal obesity (McEwen, 2005) are more likely to be found in the group we racialize as ‘black’ in America—particularly women (Gillum, 1987; Gillum and Hyattsville, 1996; Barnes, Alexander, and Staggers, 1997; Worral et al, 2002; Kataoka et al, 2013).
Geronimus et al (2006) set to find out whether or not the heightened rate of stressors (e.g., racism, environmental pollution, etc) can explain why black bodies are more ‘weathered’ than white bodies. They found that such differences were not explained by poverty, indicating that it even affects well-off blacks. Allostatic load refers to heightened hormonal production in response to stressors. We know that physiological is homeodynamic and therefore changes based on the immediate environment and social environment (for example, when you feel like you’re about to get into a fight, your heart rate increases and you get ready to ‘fight or flight’).
Experiencing racism (environmental stimuli; real or imagined, the outcome is the same) is associated with increased blood pressure (HTN). So if one experiences racism they will them experience an increase in blood pressure, as BP is a physiological variable (Armstead et al, 1987; McNeilly et al, 1995; see Doleszar et al, 2018 for a review). The concept of weathering, then, shows that racial health disparities are, in fact, racist health disparities (Sullivan, 2015: 106). Racism, then, contributes to higher levels of allostasis and, along with it, higher levels of certain hormones associated with higher allostasis.
One way to measure biological age is by measuring the length of telomeres. Telomeres are found at the ends of chromosomes. Since telomere lengths shorten with age (Shammas, 2012), those with shorter telomeres are ‘biologically older’ than those of the same age with longer telomeres. Geronimus et al (2011) showed that black women had shorter telomeres than white women, which was due to subjective and objective stressors (i.e., racism). Black women in the age group 49-55 were 7.5 years ‘older’ than white women. Thus, they had an older physiological age compared to their chronological age. It is known that direct contact with discriminatory events is associated with poor health outcomes. Harrell, Hall, and Taliaferro (2003) note that:
“…physiological set points and the mechanisms governing them are not fixed. External stressors can permanently alter physiological functioning. Racism increases the volume of stress one experiences and may contribute directly to the physiological arousal that is a marker of stress-related diseases.
Social factors can, indeed, influence physiology and there is a wealth of information on how the social becomes biological and how environmental (social) factors influence physiological systems. Forrester et al (2019) replicated Geronimus’ findings, showing that blacks have a higher ‘biological age’ than whites and that psychosocial factors affect blacks more than whites. Simons et al (2020) also replicated Geronimus’ findings, showing that persistent exposure to racism was associated with higher rates of inflammation in blacks which then predicted higher rates of disease in blacks compared to whites. Such discrimination can help to explain differences in birth outcomes (e.g., Jasienska, 2009), stress, inflammation, obesity, stroke rates, etc in blacks compared to whites (Molnar, 2015).
But what is the mechanism by which higher allostatic load scores contribute to negative outcomes and shorter telomeres indicating a higher biological age? When one feels that they are being discriminated against, the sympathetic nervous system activates due to chronic stress and along with it HPA dysfunction. What this means is that there is a loss of the anti-inflammatory effects of cortisol—it becomes blunted. This then increases oxidative stress and inflammation. Thus, the inflammatory processes result in cardiovascular disease and immune and metabolic dysfunction. The HPA axis monitors and responds to stress—allostatic load. When stress hormones are released, the adrenal gland is targeted. When it receives a signal from the pituitary gland, it pumps epinephrine and norepinephrine into the body, causing our hearts to beat faster, causing us to breathe more deeply—what is known as ‘fight or flight.’ Cortisol is also released and is known as a stress hormone, but when the stressful event is over, all three hormones return to baseline. Thus, the higher amount of stress hormones in circulation indicates higher levels of allostatic load—higher levels of stress in the individual in question. We know that blacks have higher levels of allostatic load (i.e., stress-related hormones) than whites (Duru et al, 2012). Barr (2014: 71-72) writes:
Imagine, though, that before the allostatic load has a chance to return to its baseline level, another stressor is sensed by the hypothalamus. The allostatic load will once again increase to the plateau level. Should the perception of stressors be ongoing, the allostatic load will not have the chance to ever fully recharge, and the adrenal gland will be producing an ongoing stream of stress response hormones. The body will experience chronic elevation in its allostatic load. […] A person experiencing repeated stressors, without the opportunity for intervals that are relatively stress-free, will experience a chronically elevated allostatic load, with higher than normal levels of circulating stress response hormones.
Conclusion
What these studies show, then, is that race is a cause of health inequalities, but it’s not inherent in biology but due to social factors that influence the physiology of the individual in question. The term ‘racism’ has many referents, and using one of them identifies ‘hereditarianism’ as a racist ideology (it is inherently ideological). These overviews of studies show that racial health inequalities are due, in part, to perceived discrimination (racism) thus they are racist health disparities. We know that physiology is a dynamic system that can respond to what occurs in the immediate environment—even the social environment (Williams, 1992). Thus, what explains part of the health inequalities between races is perceived discrimination—racism—and how it affects the body’s physiological systems (HPA axis, HTN, etc) and telomeres.
Follow the Leader? Selfish Genes, Evolution, and Nationalism
1750 words
Yet we get tremendously increased phenotypic variation … because the form and variation of cells, what they produce, whether to grow, to move, or what kind of cell to become, is under control of a whole dynamic system, not the genes. (Richardson, 2017: 125)
In 1976 Richard Dawkins published his groundbreaking book The Selfish Gene (Dawkins, 1976). In the book, Dawkins argues that selection occurs at the level of the gene—“the main theme of his book is a metaphorical account of competition between genes …” (Midgley, 2010: 45). Others then took note of the new theory and attempted to integrate it into their thinking. But is it as simple as Dawkins makes it out to be? Are we selfish due to the genes we carry? Is the theory testable? Can it be distinguished from other competing theories? Can it be used to justify certain behaviors?
Rushton, selfish genes, nationalism and politics
JP Rushton is a serious scholar, perhaps most well-known for attempting to use r/K selection theory to explain human behavior (Anderson, 1991). perhaps has the most controversial use of Dawkins’ theory. The main axiom of the theory is that an organism is just a gene’s way of ensuring the survival of other genes (Rushton, 1997). Thus, Rushton’s formulated genetic similarity theory posits that those who are more genetically similar—who share more genes—will be more altruistic toward those with more similar genes even if they are not related and will therefore show negative attitudes to less genetically similar individuals. This is the gene’s “way” of propagating themselves through evolutionary time. Richardson (2017: 9-11) tells us of all of the different ways in which genes are invoked to attempt to justify X.
In the beginning of his career, Rushton was a social learning theorist studying altruism, even publishing a book on the matter—Altruism, Socialization and Society (Rushton, 1980). Rushton reviews the sociobiological literature and concludes that altruism is a learned behavior. Though, Rushton seems to have made the shift from a social learning perspective to a genetic determinist perspective in the years between the publication of Altruism, Socialization and Society and 1984 when he published his genetic similarity theory. So, attempting to explain altruism through genes, while not part of Rushton’s original research programme, seems, to me, to be a natural evolution in his thought (however flawed it may be).
Dawkins responded to the uses of his theory to attempt to justify nationalism and patriotism through an evolutionary lens during an interview with Frank Miele for Skeptic:
Skeptic: How do you evaluate the work of Irena”us Eibl-Eibesfeldt, J.P. Rushton, and Pierre van den Berghe, all of whom have argued that kin selection theory does help explain nationalism and patriotism?
Dawkins: One could invoke a kind “misfiring” of kin selection if you wanted to in such cases. Misfirings are common enough in evolution. For example, when a cuckoo host feeds a baby cuckoo, that is a misfiring of behavior which is naturally selected to be towards the host’s own young. There are plenty of opportunities for misfirings. I could imagine that racist feeling could be a misfiring, not of kin selection but of reproductive isolation mechanisms. At some point in our history there may have been two species of humans who were capable of mating together but who might have produced sterile hybrids (such as mules). If that were true, then there could have been selection in favor of a “horror” of mating with the other species. Now that could misfire in the same sort of way that the cuckoo host’s parental impulse misfires. The rule of thumb for that hypothetical avoiding of miscegenation could be “Avoid mating with anybody of a different color (or appearance) from you.”
I’m happy for people to make speculations along those lines as long as they don’t again jump that is-ought divide and start saying, “therefore racism is a good thing.” I don’t think racism is a good thing. I think it’s a very bad thing. That is my moral position. I don’t see any justification in evolution either for or against racism. The study of evolution is not in the business of providing justifications for anything.
This is similar to his reaction when Bret Weinstein remarked that the Nazi’s “behaviors” during the Holocaust “were completely comprehensible at the level of fitness”—at the level of the gene.” To which Dawkins replied “I think nationalism may be an even greater evil than religion. And I’m not sure that it’s actually helpful to speak of it in Darwinian terms.” This is what I like to call “rampant adaptationism.”
This is important because Rushton (1998) invokes Dawkins’ theory as justification for his genetic similarity theory (GST; Rushton, 1997), attempting to justify ethno-nationalism from a gene’s-eye view. Rushton did what Dawkins warned against: using the theory to justify nationalism/patriotism. Rushton (1998: 486) states that “Genetic Similarity Theory explains why” ethnic nationalism has come back into the picture. Kin selection theory (which, like with selfish gene theory, Rushton invoked) has numerous misunderstandings attached to it, and of course, Rushton, too, was an offender (Park, 2007).
Dawkins (1981), in Selfish genes in race or politics stated that “It is annoying to find this elegant and important theory being dragged down to the ephemeral level of human politics, and parochial British politics at that.” Rushton (2005: 494), responded, stating that “feeling a moral obligation to condemn racism, some evolutionists minimised the theoretical possibility of a biological underpinning to ethnic or national favouritism.“
Testability?
The main premise of Dawkins’ theory is that evolution is gene-centered and that selection occurs at the level of the gene—genes that propagate fitness will be selected for while genes that are less fit are selected against. This “genes’-eye view” of evolution states “that adaptive evolution occurs through differential survival of competing genes, increasing the allele frequency of those alleles whose phenotypic trait effects successfully promote their own propagation, with gene defined as “not just one single physical bit of DNA [but] all replicas of a particular bit of DNA distributed throughout the world.“
Noble (2018) discusses “two fatal difficulties in the selfish gene version of neo-Darwinism“:
The first is that, from a physiological viewpoint, it does’t lead to a testable prediction. The only problem is that the central definition of selfish gene theory is not independent of the only experimental test of the theory, which is whether genes, defined as DNA sequences, are in fact selfish, i.e., whether their frequency in the gene pool increases (18). The second difficulty is that DNA can’t be regarded as a replicator separate from the cell (11, 17). The cell, and specifically its living physiological functionality, is what makes DNA be replicated faithfully, as I will explain later.
Noble (2017: 156) further elaborates in Dance to the Tune of Life: Biological Relativity:
Could this problem be avoided by attaching a meaning to ‘selfish’ as applied to DNA sequences that is independent of meanings in terms of phenotype? For example. we could say that a DNA sequence is ‘selfish’ to the extent which its frequency in subsequent generations is increased. This at least would be an objective definition that could be measured in terms of population genetics. But wait a minute! The whole point of the characterisation of a gene as selfish is precisely that this property leads to its success in reproducing itself. We cannot make the prediction of a theory be the basis of the definition of the central element of the theory. If we do that, the theory is empty from the viewpoint of empirical science.
Dawkins’ theory is, therefore “not a physiologically testable hypothesis” (Noble, 2011). Dawkins’ theory posits that the gene is the unit of selection, whereas the organism is only used to propagate the selfish genes. But “Just as Special Relativity and General Relativity can be succintly phrased by saying that there is no global (privileged) frame of reference, Biological Relativity can be phrased as saying that there is no global frame of causality in organisms” (Noble, 2017: 172). Dawkins’ theory privileges the gene as the unit of selection, when there is no direct unit of selection in multi-level biological systems (Noble, 2012).
In The Solitary Self: Darwin and the Selfish Gene, Midgley (2010) states “The choice of the word “selfish” is actually quite a strange one. This word is not really a suitable one for what Dawkins wanted to say about genetics because genes do not act alone.” As Dawkins later noted, “the cooperative gene” would have been a better description, while The Immortal Gene would have been a better title for the book. Midgley (2010: 16) states that Dawkins and Wilson (in The Selfish Gene and Sociobiology, respectively) “use a very simple concept of selfishness derived not from Darwin but from a wider background of Hobbesian social atomism, and give it a general explanation of all behaviour, including that of humans.” Dawkins and others claim that “the thing actually being selected was the genes” (Midgley, 2010: 47).
Conclusion
Developmental systems theory (DST) explains and predicts more than the neo-Darwinian Modern Synthesis (Laland et al, 2015). Dawkins’ theory is not testable. Indeed, the neo-Darwinian Modern Synthesis (and along with it Dawkins’ selfish gene theory) is dead, an extended synthesis explains evolution. As Fodor and Piattelli-Palmarini (2010a, b) and Fodor (2008) state in What Darwin Got Wrong, natural selection is not mechanistic and therefore cannot select-for genes or traits (also see Midgley’s 2010: chapter 6 discussion of Fodor and Piattelli-Palmarini). (Okasha, 2018 also discusses ‘selection-for- genes—and, specifically, Dawkins’ selfish gene theory.)
Dawkins’ theory was repurposed, used to attempt to argue for ethno-nationalism and patriotism—even though Dawkins himself is against such uses. Of course, theories can be repurposed from their original uses, though the use of the theory is itself erroneous, as is the case with regard to Rushton, Russel and Wells (1984) and Rushton (1997, 1998). Since the theory is itself not testable (Noble, 2011, 2017), it should therefore—along with all other theories that use it as its basis—be dropped. While Rushton’s change from social learning to genetic causation regarding altruism is not out of character for his former research (he began his career as a social learning theorist studying altruism; Rushton, 1980), his use of the theory to attempt to explain why individuals and groups prefer those more similar to themselves ultimately fails since it is “logically flawed” (Mealey, 1984: 571).
Genes ‘do’ what the physiological system ‘tells’ them to do; they are just inert, passive templates. What is active is the cell—the genome is an organ of the cell and is what is ‘immortal.’ Genes don’t “control” anything; they are used by and for the physiological system to carry out certain processes (Noble, 2017; Richardson, 2017: chapter 4, 5). There are new views of what ‘genes’ really are (Portin and Wilkins, 2017), what they are and were—are—used for.
Development is dynamic and not determined by genes. Genes (DNA sequences) are followers, not leaders. The leader is the physiological system.
A Systems View of Kenyan Success in Distance Running
1550 words
The causes of sporting success are multi-factorial, with no cause being more important than the other since the whole system needs to work in concert to produce the athletic phenotype–call this “causal parity” of athletic success determinants. For a refresher, take what Shenk (2010: 107):
As the search for athletic genes continues, therefore, the overwhelming evidence suggests that researchers will instead locate genes prone to certain types of interactions: gene variant A in combination with gene variant B, provoked into expression by X amount of training + Y altitude + Z will to win + a hundred other life variables (coaching, injuries, etc.), will produce some specific result R. What this means, of course, What this means, of course, is that we need to dispense rhetorically with thick firewall between biology (nature) and training (nurture). The reality of GxE assures that each person’s genes interacts with his climate, altitude, culture, meals, language, customs and spirituality—everything—to produce unique lifestyle trajectories. Genes play a critical role, but as dynamic instruments, not a fixed blueprint. A seven- or fourteen- or twenty-eight-year-old is not that way merely because of genetic instruction. (Shenk, 2010: 107) [Also read my article Explaining African Running Success Through a Systems View.]
This is how athletic success needs to be looked at; not reducing it to genes or a group of genes that ’cause’ athletic success. Since to be successful in the sport of the athlete’s choice takes more than being born with “the right” genes.
Recently, a Kenyan woman—Joyciline Jepkosgei—won the NYC marathon in here debut (November 3rd, 2019), while Eliud Kipchoge—another Kenyan—became the first human ever to complete a marathon (26.2 miles) in under 2 hours. I recall in the spring reading that he said he would break the 2-hour mark in October. He also attempted to break it in 2017 in Italy but, of course, he failed. His official time in Italy was 2:00:25! While he set the world record in Berlin at 2:01:39. Kipchoge’s official time was 1:59:40—twenty seconds shy of 2 hours—that means his average mile pace was about 4 minutes and 34 seconds. That is insane. (But the IAAF does not accept the time as a new world record since it was not in an open competition—Kipchoge had a slew of Olympic pacesetters following him; an electric car drove just ahead of him and pointed lasers at the ground showing him where to run; so he shaved 2 minutes off his time—2 crucial minutes—according to sport scientist Ross Tucker; and . So he did not set a world record. His feat, though, is still impressive.)
Now, Kipchoge is Kenyan—but what’s his ethnicity? Surprise surprise! He is of the Nandi tribe, more specifically, of the Talai subgroup, born in Kapsisiywa in the Nandi county. Jepkosgei, too, is Nandi, from Cheptil in Nandi county. (Jepkosgei also set the record for the half marathon in 2017. Also, see her regular training regimen and what she does throughout the day. This, of course, is how she is able to be so elite—without hard training, even without “the right genetic makeup”, one will not become an elite athlete.) What a strange coincidence that these two individuals who won recent marathons—and one who set the best time ever in the 26.2 mile race—are both Kenyan, specifically Nandi?
Both of these runners are from the same county in Kenya. Nandi county is elevated about 6,716 ft above sea level. Being born and living at a high elevation means that they have different kinds of physiological adaptations due to being born at such a higher elevation. Living and training at such high elevations means that they have greater lung capacities since they are breathing in thinner air. Those born in highlands like Kipchoge and Jepkosgei have larger lungs and thorax volumes, while oxygen intake is enhanced by increases in lung compliance, pulmonary diffusion, and ventilation (Meer, Heymans, and Zijlstra, 1995).
Those exposed to such elevation develop what is known as “high-altitude hypoxia.” Humans born at high altitudes are able to cope with such a lack of oxygen, since our physiological systems are dynamic—not static—and can respond to environmental changes within seconds of them occurring. Babes born at higher elevations have increased ventilation, and a rise in the alveolar and the pressure of arterial oxygen (Meer, Heymans, and Zjilstra, 1995).
Kenyans have 5 percent longer legs and 12 percent lighter muscles than Scandinavians (Suchy and Waic, 2017). Mooses et al (2014) notes that “upper leg length, total leg length and total leg length to body height ratio were correlated with running performance.” Kong and de Heer (2008) note that:
The slim limbs of Kenyan distance runners may positively contribute to performance by having a low moment of inertia and thus requiring less muscular effort in leg swing. The short ground contact time observed may be related to good running economy since there is less time for the braking force to decelerate forward motion of the body.
An abundance of type I muscle fibers is conducive to success in distance running (Zierath and Hawley, 2004), though Kenyans and Caucasians have no difference in type I muscle fibers (Saltin et al, 1995; Larsen and Sheel, 2015). That, then, throws a wrench in the claim that a whole slew of anatomic and physiologic variables conducive to running success is the cause for Kenyan running success—specifically the type I fibers—right? Wrong. Recall that the appearance of the athletic phenotype is due to nature and nurture—genes and environment—working together in concert. Kenyans are more likely to have slim, long limbs with lower body fat while they lived and trained over 6000 ft high. Their will to win to better themselves and their families’ socioeconomic status, too, plays a part. As I have argued in-depth for years—we cannot understand athletic success and elite athleticism without understanding individual histories, how they grew up, and what they did as a child.
For example, Wilbur and Pitsiladis (2012) espouse a systems view of Kenyan marathon success, writing:
In general, it appears that Kenyan and Ethiopian distance-running success is not based on a unique genetic or physiological characteristic. Rather, it appears to be the result of favorable somatotypical characteristics lending to exceptional biomechanical and metabolic economy/efficiency; chronic exposure to altitude in combination with moderate-volume, high-intensity training (live high + train high), and a strong psychological motivation to succeed athletically for the purpose of economic and social advancement.
Becoming a successful runner in Kenya can lead to economic opportunities not afforded to those who do not do well in running. This, too, is a factor in Kenyan running success. So, for the ignorant people who would—pushing a false dichotomy of genes and environment—state that Kenyan running success is due to “socioeconomic status”—they are right, to a point (even if they are mocking it and making their genetic determinism seem more palatable). See figure 6 for their hypothetical model:

This is one of the best models I have come across explaining the success of these people. One can see that it is not reductonist; note that there is no appeal to genes (just variables that genes are implicated IN! Which is not the same as reductionism). It’s not as if one can have an endomorphic somatotype with Kenyan training and their psychological reasons for becoming runners. The ecto-dominant somatotype is a necessary factor for success; but all four of these—biomechanical & physiological, training, and psychological—factors explain the success of the running Kenyans and, in turn, the success of Kipchoge and Jepkosgei. African dominance in distance running is, also, dominated by the Nandi subtribe (Tucker, Onywera, and Santos-Concejero, 2015). Knechtle et al (2016) also note that male and female Kenyan and Ethiopian runners are the youngest and fast at the half and full marathons.
The actual environment—climate—on the day of the race, too plays a factor. El Helou et al (2012) note that “Air temperature is the most important factor influencing marathon running performance for runners of all levels.” Nikolaidis et al (2019) note that “race times in the Boston Marathon are influenced by temperature, pressure, precipitations, WBGT, wind coming from the West and wind speed.”
The success of Kenyans—and other groups—shows how the dictum “Athleticism is irreducible to biology” (St. Louis, 2004) is true. How does it make any sense to attempt to reduce athletic success down to one variable and say that that explains the overrepresentation of, say, Kenyans in distance running? A whole slew of factors needs to occur to an individual, along with actually wanting to do something, in order for them to succeed at distance running.
So, what makes Kenyans like Kipchoge and Jepkosgei so good at distance running? It’s due to an interaction with genes and environment, since we take a systems and not a reductionist view of sport success. Even though Kipchoge’s time does not count as an official world record, what he did was still impressive (though not as impressive if he would have done so without all of the help he had). Looking at the system, and not trying to reduce the system to its parts, is how we will explain why some groups are better than others. Genes, of course, play a role in the ontogeny of the athletic phenotype, but they are not the be-all-end-all that genetic reductionists seem to make it out to be. The systems view for Kenyan running success shown here is how and why Kenyans—Kipchoge and Jepkosgei—dominate distance running.
Usain Bolt, Michael Phelps, and Caster Semenya: Should Semenya Take Drugs to Decrease Testosterone Levels?
1300 words
In the past week in the world of sport, all the rage has been over mid- to long-distance runner Caster Semenya. Semenya has won the 800 m in 1:56.72 and setting world records in the 400, 800, and 1500 m with times of 50.74, 1:58.45 and 4:10.93 respectively. In 2012 and 2016, Semenya won the gold for the 800 m with times of 1:57.23 and 1:55.28 respectively. I won’t really discuss the anatomic and physiologic advantages today. What I will discuss, though, is the fact that Semenya has been told that she has to take drugs to decrease her testosterone levels or she cannot compete anymore. Semenya was told to decrease her testosterone levels or she could face a ban in the 800 m. The new rules state that:
Female athletes affected must take medication for six months before they can compete, and then maintain a lower testosterone level.
If a female athlete does not want to take medication, then they can compete in:
- International competitions in any discipline other than track events between 400m and a mile
- Any competition that is not an international competition
- The male classification at any competition, at any level, in any discipline
- Any intersex, or similar, classification
But Semenya has declined taking these drugs—so her future is up in the air. So, if Semenya—or any other athlete—has to take drugs to decrease their levels since it gives an unfair advantage, then, in my opinion, this may lead to changes in other sports as well.
Look at Michael Phelps. Michael Phelps has won a record 28, winning 23 medals at Rio in 2016. Phelps has a long, thin torso which decreases drag in the water. Phelps’ wing span is 6’7” while he is 6’4”—which is disproportionate to his height. He has the torso of a 6’8” person, which gives him a greater reach per stroke. His lower body is 5’10” which lowers the resistance against the water. He has large hands and feet (with flexible ankles), which help with paddling capacity (size 14 shoe; yours truly wears a size 13).
There is one more incredible thing about Phelps: He produces around 50 percent lower lactic acid. Think of the last time that you have run for some distance. The burning you feel in your legs is a build up of lactic acid. Lactic acid causes fatigue and also slows muscle contractions—this occurs through lactic acid passing through the bloodstream, becoming lactate. (Note that it does not necessarily cause fatigue; Brooks, 2001.) Phelps does not produce normal levels of lactic acid, and so he is ready to go again shortly after a bout of swimming.
Phelps said “In between the 200m free and the fly heats I have probably had in total about 10 minutes to myself.” A normal person’s muscles would be too fatigued and cramped. I would also assume that Phelps has an abundance of type I muscle fibers as well.
Now take Usain Bolt. The 100 m dash is, mostly, an anaerobic race. What this means is that mitochondrial respiration has minimal effect on the type of energy used during the event (Majumdar and Robergs, 2011). So during anaerobic events, there is no free oxygen to drive energy—the energy stored in the muscle is used to perform movement through a process called glycolysis. Sprinting is an intense exercise—fuel choice during exercise is determined by the intensity of said exercise. “A 100-meter sprint is powered by stored ATP, creatine phosphate, and anaerobic glycolysis of muscle glycogen.”
Now we can look at the physical advantages they have. Swimmers and runners, on average, have different centers of mass (Bejan, Jones, and Charles, 2010). In all actuality, Phelps and Bolt are the perfect example of this phenomenon. Winning runners have a West-African origin and winning runners are more likely to be white. These somatotypic differences between the two races influence why they excel in these two different sports.
Usain Bolt is 6’5”. Since he is that height, and he has long legs, he necessarily has a longer stride—Bolt is the perfect example of Bejan, Jones, and Charles’ (2010) paper. So take the average white sprinter of the same height as Bolt. Ceteris paribus, Bolt will have a higher center of mass than the white athlete due to his longer limbs and and smaller circumference. Krogman (1970) found that, in black and white youths of the same height, blacks had shorter trunks and longer limbs, which lends credence to the hypothesis.
Phelps is 6’4”. As noted above, he has a long torso and long limbs. Long torsos are conducive to a lower center of mass—what whites and Asians have, on average. So long torsos mean that one will have taller sitting heights than those with short torsos. This means that whites and Asians have taller sitting heights than blacks, who have shorter torsos. This average taller sitting height is conducive to the longer torsos which is why whites excel in swimming. Bejan, Jones, and Charles (2010) also note that, if it were not for the short stature of Asians, they would be better swimmers than whites.
In any case, the different centers of mass on average between blacks and whites are conducive to faster times in the sports they excel at. For whites, the three percent increase in center of mass means that there would be a 1.5 percent increase in winning speed and a 1.5 percent decrease in winning time in the case of swimming. The same holds for blacks, but in the case of running: their higher center of mass is conducive to a 1.5 percent increase in winning speed and also a 1.5 percent decrease in winning time, which would be a .15 second decrease, or from 10 s to 9.85 s—which is a large differential when it comes to sprinting. (Note that this phenomenon also holds for black women and white women—black women are better sprinters and white women are better swimmers. Asian women excel in the 100 m freestyle, but not Asian men for reasons discussed above.)
Now put this all together. If Phelps and Bolt have such advantages over their competition and they—supposedly—win due to them, then if Semenya has to decrease her T levels, why shouldn’t Phelps and Bolt decrease X, Y, or Z since they have physiologic/anatomic advantages as well? Why does no one talk about Semenya’s anatomic advantages over, say, white women and why only bring up Semenya’s testosterone levels? Forcing Semenya to decrease T levels will set a bad precedent in sport. What would stop a losing competitor from complaining that the winner—who keeps winning—has an “unfair” physiologic/anatomic advantage and must do X to change it? (Or say that the anatomic advantage they possess is “unfair” and they should be barred from competition?)
Here’s the thing: Watching sport, we want to see the best-of-the-best compete. Wouldn’t that logically imply that we want to see Semenya compete and not rid herself of her advantage? If Semenya’s physiologic advantage(s) is being discussed, why not Semenya’s anatomic advantages? It does not make sense to focus on one variable—as all variables interact to produce the athletic phenotype (Louis, 2004). Phelps and Bolt perfectly embody the results of Bejan, Jones, and Charles (2010)—they have, what I hope are—well-known advantages, and these advantages, on average, are stratified between race due to anatomic differences (see Gerace et al, 1994; Wagner and Heyward, 2000).
Phelps and Bolt have anatomic and physiologic advantages over their competition, just as Semenya does, just like any elite athlete, especially the winners compared to their competition. If Semenya is forced to decrease her testosterone levels, then this will set a horrible precedent for sport, and people may then clamor for Phelps and Bolt to do X, Y, and Z due to their physical advantages. For this reason, Semenya should not decrease her testosterone levels and should be allowed to compete in mid-distance running.
Athleticism is Irreducible to Biology: A Systems View of Athleticism
1550 words
Reductionists would claim that athletic success comes down to the molecular level. I disagree. Though, of course, understanding the molecular pathways and how and why certain athletes excel in certain sports can and will increase our understanding of elite athleticism, reductionist accounts do not tell the full story. A reductionist (which I used to be, especially in regard to sports; see my article Racial Differences in Muscle Fiber Typing Cause Differences in Elite Sporting Competition) would claim that, as can be seen in my article, the cause for elite athletic success comes down to the molecular level. Now, that I no longer hold such reductionist views in this area does not mean that I deny that there are certain things that make an elite athlete. However, I was wrong to attempt to reduce a complex bio-system and attempt to pinpoint one variable as “the cause” of elite athletic success.
In the book The Genius of All of Us: New Insights into Genetics, Talent, and IQ, David Shenk dispatches with reductionist accounts of athletic success in the 5th chapter of the book. He writes:
2. GENES DON’T DIRECTLY CAUSE TRAITS; THEY ONLY INFLUENCE THE SYSTEM.
Consistent with other lessons of GxE [Genes x Environment], the surprising finding of the $3 billion Human Genome Project is that only in rare instances do specific gene variants directly cause specific traits or diseases. …
As the search for athletic genes continues, therefore, the overwhelming evidence suggests that researchers will instead locate genes prone to certain types of interactions: gene variant A in combination with gene variant B, provoked into expression by X amount of training + Y altitude + Z will to win + a hundred other life variables (coaching, injuries, etc.), will produce some specific result R. What this means, of course, What this means, of course, is that we need to dispense rhetorically with thick firewall between biology (nature) and training (nurture). The reality of GxE assures that each persons genes interacts with his climate, altitude, culture, meals, language, customs and spirituality—everything—to produce unique lifestyle trajectories. Genes play a critical role, but as dynamic instruments, not a fixed blueprint. A seven- or fourteen- or twenty-eight-year-old is not that way merely because of genetic instruction. (Shenk, 2010: 107) [Also read my article Explaining African Running Success Through a Systems View.]
This is looking at the whole entire system: genes, to training, to altitude, to will to win, to numerous other variables that are conducive to athletic success. You can’t pinpoint one variable in the entire system and say that that is the cause: each variable works together in concert to produce the athletic phenotype. One can invoke Noble’s (2012) argument that there is no privileged level of causation in the production of an athletic phenotype. There are just too many factors that go into the production of an elite athlete, and attempting to reduce it to one or a few factors and attempt to look for those factors in regard to elite athleticism is a fool’s errand. So we can say that there is no privileged level of causation in regard to the athletic phenotype.
In his paper Sport and common-sense racial science, Louis (2004: 41) writes:
The analysis and explanation of racial athleticism is therefore irreducible to
biological or socio-cultural determinants and requires a ‘biocultural approach’
(Malina, 1988; Burfoot, 1999; Entine, 2000) or must account for environmental
factors (Himes, 1988; Samson and Yerl`es, 1988).
Reducing anything, sports included, to environmental/socio-cultural determinants and biology doesn’t make sense; I agree with Louis that we need a ‘biocultural approach’, since biology and socio-cultural determinants are linked. This, of course, upends the nature vs. nurture debate; neither “nature” nor “nurture” has won, they causally depend on one another to produce the elite athletic phenotype.
Louis (2004) further writes:
In support of this biocultural approach, Entine (2001) argues that athleticism is
irreducible to biology because it results from the interaction between population-based genetic differences and culture that, in turn, critiques the Cartesian dualism
‘which sees environment and genes as polar-opposite forces’ (p. 305). This
critique draws on the centrality of complexity, plurality and fluidity to social
description and analysis that is significant within multicultural common sense. By
pointing to the biocultural interactivity of racial formation, Entine suggests that
race is irreducible to a single core determinant. This asserts its fundamental
complexity that must be understood as produced through the process of
articulation across social, cultural and biological categories.
Of course, race is irreducible to a single core determinant; but it is a genuine kind in biology, and so, we must understand the social, cultural, and biological causes and how they interact with each other to produce the athletic phenotype. We can look at athlete A and see that he’s black and then look at his somatotype and ascertain that the reason why athlete A is a good athlete is conducive to his biology. Indeed, it is. One needs a requisite morphology in order to succeed in a certain sport, though it is quite clearly not the only variable needed to produce the athletic phenotype.
One prevalent example here is the Kalenjin (see my article Why Do Jamaicans, Kenyans, and Ethiopians Dominate Running Competitions?). There is no core determinant of Kalenjin running success; even one study I cited in my article shows that Germans had a higher level of a physiological variable conducive to long-distance running success compared to the Kalenjin. This is irrelevant due to the systems view of athleticism. Low Kenyan BMI (the lowest in the world), combined with altitude training (they live in higher altitudes and presumably compete in lower altitudes), a meso-ecto somatotype, the will to train, and even running to and from where they have to go all combine to show how and why this small tribe of Kenyans excel so much in these types of long-distance running competitions.
Sure, we can say that what we know about anatomy and physiology that a certain parameter may be “better” or “worse” in the context of the sport in question, no one denies that. What is denied is the claim that athleticism reduces to biology, and it does not reduce to biology because biology, society, and culture all interact and the interaction itself is irreducible; it does not make sense to attempt to partition biology, society, and culture into percentage points in order to say that one variable has primacy over another. This is because each level of the system interacts with every other level. Genes, anatomy and physiology, the individual, the overarching society, cultural norms, peers, and a whole slew of other factors explain athletic success not only in the Kalenjin but in all athletes.
Broos et al (2016) showed that in those with the RR genotype, coupled with the right morphology and fast twitch muscle fibers, this would lead to more explosive contractions. Broos et al (2016) write:
In conclusion, this study shows that a-actinin-3 deficiency decreases the contraction velocity of isolated type IIa muscle fibers. The decreased cross-sectional area of type IIa and IIx fibers may explain the increased muscle volume in RR genotypes. Thus, our results suggest that, rather than fiber force, combined effects of morphological and contractile properties of individual fast muscle fibers attribute to the enhanced performance observed in RR genotypes during explosive contractions.
This shows the interaction between the genotype, morphology, fast twitch fibers (which blacks have more of; Caeser and Henry, 2015), and, of course, the grueling training these elite athletes go through. All of these factors interact. This further buttresses the argument that I am making that different levels of the system causally interact with each other to produce the athletic phenotype.
Pro-athletes also have “extraordinary skills for rapidly learning complex and neutral dynamic visual scenes” (Faubert, 2013). This is yet another part of the system, along with other physical variables, that an elite athlete needs to have. Indeed, as Lippi, Favalaro, and Guidi (2008) write:
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.
So now we have: (1) strong mental background; (2) genes; (3) morphology; (4) Vo2 max; (5) altitude; (6) will to win; (7) training; (8) coaching; (9) injuries; (10) peer/familial support; (11) fiber typing; (12) heart strength etc. There are of course myriad other variables that are conducive to athletic success but are irreducible since we need to look at it in the whole context of the system we are observing.
In conclusion, athleticism is irreducible to biology. Since athleticism is irreducible to biology, then to explain athleticism, we need to look at the whole entire system, from the individual all the way to the society that individual is in (and everything in between) to explain how and why athletic phenotypes develop. There is no logical reason to attempt to reduce athleticism to biology since all of these factors interact. Therefore, the systems view of athleticism is the way we should view the development of athletic phenotypes.
(i) Nature and Nurture interact.
(ii) Since nature and nurture interact, it makes no sense to attempt to reduce anything to one or the other.
(iii) Since it makes no sense to attempt to reduce anything to nature or nurture since nature and nurture interact, then we must dispense with the idea that reductionism can causally explain differences in athleticism between individuals.
Muscle Fibers, Obesity, Cardiometabolic Disorders, and Race
2650 words
The association between muscle fiber typing obesity and race is striking. It is well-established that blacks have a higher proportion of type II skeletal muscle fibers than whites and these higher proportions of these specific types of muscle fibers lead to physiological differences between the two races which then lead to differing health outcomes between them—along with differences in athletic competition. Racial differences in health are no doubt complex, but there are certain differences between the races that we can look at and say that there is a relationship here that warrants further scrutiny.
Why is there an association between negative health outcomes and muscle phsyiology? The answer is very simple if one knows the basics of muscle physiology and how and why muscles contract (it is worth noting that out of a slew of anatomic and phsyiologic factors, movement is the only thing we can consciously control, compare to menstration and other similar physiologic processes which are beyond our control). In this article, I will describe what muscles do, how they are controlled, muscle physiology, the differences in fiber typing between the races and what it means for health outcomes between them.
Muscle anatomy and physiology
Muscle fiber number is determined by the second trimester. Bell (1980) noted that skeletal muscle fiber in 6 year olds is not different from normal adult tissue, and so, we can say that between the time in the womb and age 6, muscle fiber type is set and cannot be changed (though training can change how certain fibers respond, see below).
Muscle anatomy and physiology is interesting because it shows us how and why we move the way we do. Tendons attach muscle to bone. Attached to the tendon is the muscle belly. The muscle belly is made up of facsicles and the fascicles are made up of muscle fibers. Muscle fibers are made up of myofibrils and myofibrils are made up of myofilaments. Finally, myofilaments are made up of proteins—specifically actin and myosin, this is what makes up our muscles.

(Image from here.)
Muscle fibers are encased by sarcolemma which contains cell components such as sarcoplasm, nuclei, and mitochondria. They also have other cells called myofibrils which contain myofilaments which are then made up of actin (thin filaments) and mysoin (thick filaments). These two types of filaments form numerous repeating sections within a myofibril and each repeating section is known as a sarcomere. Sarcomeres are the “functional” unit of the muscle, like the neuron is for the nervous system. Each ‘z-line’ denotes another sarcomere across a myofibril (Franzini-Armstrong, 1973; Luther, 2009).
Other than actin and myosin, there are two more proteins important for muscle contraction: tropomyosin and troponin. Tropomyosin is found on the actin filament and it blocks myosin binding sites which are located on the actin filament, and so it keeps myosin from attaching to muscle while it is in a relaxed state. On the other hand, troponin is also located on the actin filament but troponin’s job is to provide binding sites for calcium and tropomyosin when a muscle needs to contract.
So the structure of skeletal muscle can be broken down like so: epymyseum > muscle belly > perimyseum > fascicle > endomyseum > muscle fibers > myofibrils > myofilaments > myosin and actin. Note diagram (C) from above; the sarcomere is the smallest contractile unit in the myofibril. According to sliding filament theory (see Cook, 2004 for a review), a sarcomere shortens as a result of the ‘z-lines’ moving closer together. The reason these ‘z-lines’ converge is because myosin heads attach to the actin filament which asynchronistically pulls the actin filament across the myosin, which then results in the shortening of the muscle fiber. Sarcomeres are the basic unit controlling changes in muscle length, so the faster or slower they fire depends on the majority type of fiber in that specific area.
But the skeletal muscle will not contract unless the skeletal muscles are stimulated. The nervous system and the muscular system communicate, which is called neural activiation—defined as the contraction of muscle generated by neural stimulation. We have what are called “motor neurons”—neurons located in the CNS (central nervous system) which can send impulses to muscles to move them. This is done through a special synapse called the neuromuscular junction. A motor neuron that connects with muscle fibers is called a motor unit and the point where the muscle fiber and motor unit meet is callled the neuromuscular junction. It is a small gap between the nerve and muscle fiber called a synapse. Action potentials (electrical impulses) are sent down the axon of the motor neuron from the CNS and when the action potential reaches the end of the axon, hormones called neurotransmitters are then released. Neurotransmitters transport the electrical signal from the nerve to the muscle.
Muscle fiber types
The two main categories of muscle fiber are type I and type II—‘slow’ and ‘fast’ twitch, respectively. Type I fibers contain more blood cappilaries, higher levels of mitochondria (which transforms food into ATP) and myoglobin which allows for an improved delivery of oxygen. Since myoglobin is similar to hemoglobin (the red pigment which is found in red blood cells), type I fibers are also known as ‘red fibers.’ Type I fibers are also smaller in diameter and slower to produce maximal tension, but are also the most fatigue-resistant type of fiber.
Type II fibers have two subdivisions—IIa and IIx—based on their mechanical and chemical properties. Type II fibers are in many ways the opposite of type I fibers—they contain far fewer blood cappilaries, mitochondria and myoglobin. Since they have less myoglobin, they are not red, but white, which is why they are known as ‘white fibers.’ IIx fibers have a lower oxidative capacity and thusly tire out quicker. IIa, on the other hand, have a higher oxidative capacity and fatigue slower than IIx fibers (Herbison, Jaweed, and Ditunno, 1982; Tellis et al, 2012). IIa fibers are also known as intermediate fast twitch fibers since they can use both anarobic and aerobic metabolism equally to produce energy. So IIx fibers are a combo of I and II fibers. Type II fibers are bigger, quicker to produce maximal tension, and tire out quicker.
Now, when it comes to fiber typing between the races, blacks have a higher proportion of type II fibers compared to whites who have a higher proportion of type I fibers (Ama et al, 1986; Ceaser and Hunter, 2015; see Entine, 2000 and Epstein, 2014 for reviews). Higher proportions of type I fibers are associated with lower chance of cardiovascular events, whereas type II fibers are associated with a higher risk. Thus, “Skeletal muscle fibre composition may be a mediator of the protective effects of exercise against cardiovascular disease” (Andersen et al, 2015).
Now that the basics of muscle anatomy and physiology are apparent, hopefully the hows and whys of muscle contraction and what different muscle fibers do are becoming clear, because these different fibers are distributed between the races in uneven frequencies, which then leads to differences in sporting performance but also differents in health outcomes.
Muscle fibers and health outcomes
We now know the physiology and anatomy of muscle and muscle fiber typing. We also know the differences between each type of skeletal muscle fiber. Since the two races do indeed differ in the percentage of skeletal muscle fiber possessed on average, we then should find stark differences in health outcomes, part of the reason being these differences in muscle fiber typing.
While blacks on average have a higher proportion of type II muscle fibers, whites have a higher proportion of type I muscle fibers. Noting what I wrote above about the differences between the fiber types, and knowing what we know about racial differences in disease outcomes, we can draw some inferences on how differences in muscle fiber typing between races/individuals can then affect disease seriousness/acquisition.
In their review of black-white differences in muscle fiber typing, Ceaser and Hunter (2015) write that “The longitudinal data regarding the rise in obesity indicates obesity rates have been highest among non-Hispanic Black women and Hispanic women.” And so, knowing what we know about fiber type differences between races and how these fibers act when they fire, we can see how muscle fiber typing would contribute to differences in disease acquisition between groups.
Tanner et al (2001) studied 53 women (n=28, lean women; and n=25, obese women) who were undergoing an elective abdominal surgery (either a hysterectomy or gastric bypass). Their physiologic/anatomic measures were taken and they were divided into races: blacks and whites, along with their obesity status. Tanner et al found that the lean subjects had a higher proportion of type I fibers and a lower proportion of type IIx fibers whereas those who were obese were more likely to have a higher proportion of type IIb muscle fibers.
Like other analyses on this matter, Tanner et al (2001) showed that the black subjects had a higher proportion of type II fibers in comparison to whites who had a higher proportion of type I fibers (adiposity was not taken into account). Fifty-one percent of the fiber typing from whites was type I whereas for blacks it was 43.7 pervent. Blacks had a higher proportion of type IIx fibers than whites (16.3 percent for whites and 23.4 for blacks). Lean blacks and lean whites, though, had a similar percentage of type IIx fibers (13.8 percent for whites and 15 percent for blacks). It is interesting to note that there was no difference in type I fibers between lean whites and blacks (55.1 percent for whites and 54.1 percent for blacks), though muscle fibers from obese blacks contained far fewer type I fibers compared to their white counterparts (48.6 percent for whites and 34.5 for blacks). Obese blacks’ muscle fiber had a higher proportion of type IIx fibers than obese whites’ fiber typing (19.2 percent for whites and 31 percent for blacks). Lean blacks and lean whites had a higher proportion of type I fibers than obese blacks and obese whites. Obese whites and obese blacks had more type IIx fibers than lean whites and lean blacks.
So, since type II fibers are insulin resistant (Jensen et al, 2007), then they should be related to glucose intloerance—type II diabetes—and blacks with ancestry from West Africa should be most affected. Fung (2016, 2018) shows that obesity is a disease of insulin resistance, and so, we can bring that same rationale to racial differences in obesity. Indeed, Nielsen and Christensen (2011) hypothesize that the higher prevalence of glucose intolerance in blacks is related to their lower percentage of type I fibers and their higher percentage of type II fibers.
Nielsen and Christensen (2011) hypothesize that since blacks have a lower percentage of type I fibers (the oxidative type), this explains the lower fat oxidation along with lower resting metabolic rate, sleeping metabolic rate, resting energy expenditure and Vo2 max in comparison to whites. Since type I fibers are more oxidative over the glycolitic type II fibers, the lower oxidative capacity in these fibers “may cause a higher fat storage at lower levels of energy intake than in individuals with a higher oxidative capacity” (Nielsen and Christensen, 2011: 611). Though the ratio of IIx and IIa fibers are extremely plastic and affected by lifestyle, Nielsen and Christensen do note that individuals with different fiber typings had similar oxidative capacity if they engaged in physical activity. Recall back to Caesar and Hunter (2015) who note that blacks have a lower maximal aerobic capacity and higher proportion of type II fibers. They note that lack of physical activity exacerbates the negative effects that a majority type II fibers has over majority type I. And so, some of these differences can be ameliorated between these two racial groups.
The point is, individuals/groups with a higher percentage of type II fibers who do not engage in physical activity have an even higher risk of lower oxidative capacity. Furthermore, a higher proportion of type II fibers implies a higher percentage of IIx fibers, “which are the least oxidative fibres and are positively associated with T2D and obesity” (Nielsen and Christensen, 2011: 612). They also note that this may explain the rural-urban difference in diabetes prevalance, with urban populations having a higher proportion of type II diabetics. They also note that this may explain the difference in type II diabetes in US blacks and West African natives—but the reverse is true for West Africans in the US. There is a higher rate of modernization and, with that, a higher chance to be less physically active and if the individual in question is less physically active and has a higher proportion of type II fibers then they will have a higher chance of acquiring metabolic diseases (obesity is also a metabolic disease). Since whites have a higher proportion of type I fibers, they can increase their fat intake—and with it, their fat oxidation—but this does not hold for blacks who “may not adjust well to changes in fat intake” (Nielsen and Christensen, 2011: 612).
Nielsen and Christensen end their paper writing:
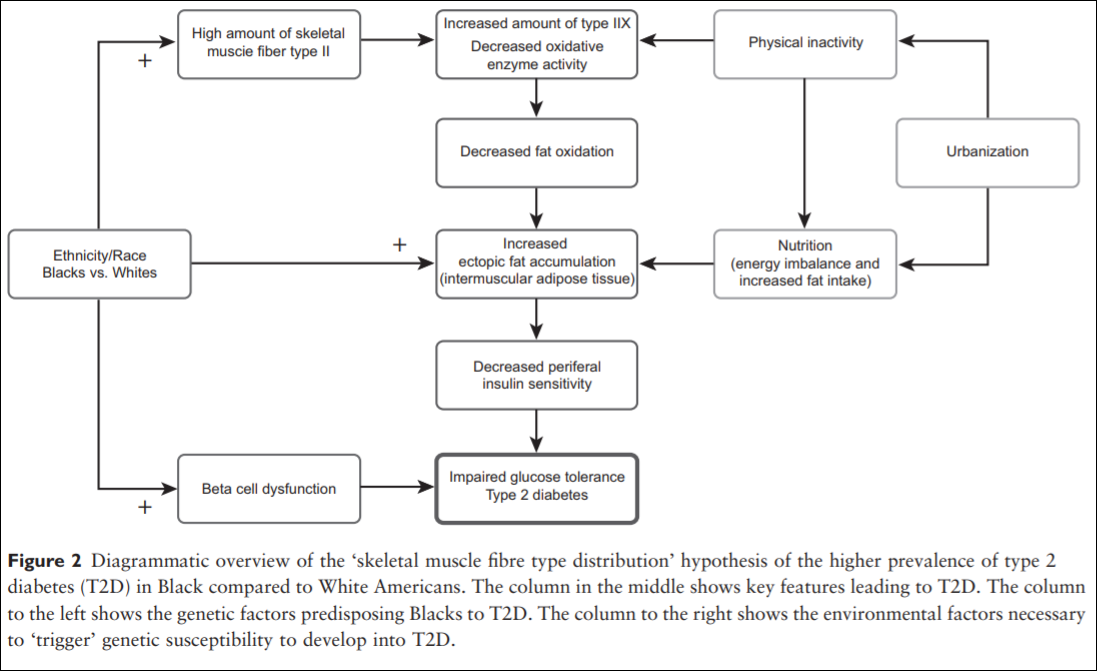
Thus, Blacks of West African ancestry might be genetically predisposed to T2D because of an inherited lower amount of skeletal muscle fibre type I, whereby the oxidative capacity and fat oxidation is reduced, causing increased muscular tissue fat accumulation. This might induce skeletal muscle insulin resistance followed by an induced stress on the insulin-producing beta cells. Together with higher beta-cell dysfunction in the West African Diaspora compared to Whites, this will eventually lead to T2D (an overview of the ‘skeletal muscle distribution hypothesis’ can be seen in Figure 2).
Lambernd et al (2012) show that muscle contractions eliminated insuin resistance by blocking pro-inflammatory signalling pathways: this is the mechanism by which physical activity decreases glucose intolerance and thusly improves health outcomes—especially for those with a higher proportion of type II fibers. Thus, it is important for individuals with type II fibers to exercise, since sedentariness is associated with an age-related insulin resistance due to impaired GLUT4 utilization (Bunprajun et al, 2013).
(Also see Morrison and Cooper’s (2006) hypothesis that “reduced oxygen-carrying capacity induced a shift to more explosive muscle properties” (Epstein, 2014: 179). Epstein notes that the only science there is on this hypothesis is one mouse and rat study showing that low hemoglobin can “induce a switch to more explosive muscle fibers” (Epstein, 2014: 178), but this has not been tested on humans to see if it would hold. If this is tested on humans and if it does hold, then that would lend credence to Morrison’s and Cooper’s (2006) hypothesis.)
Conclusion
Knowing what we know about muscle anatomy and physiology and how muscles act we can understand the influence the different muscle types have on disease and how they contribute to disease variation between race, sex and the individual level. Especially knowing how type II fibers act when the individual in question is insulin resistant is extremely important—though it has been noted that individuals who participate in aerobic exercise decrease their risk for cardiometabolic diseases and can change the fiber distribution difference between IIx and IIa fibers, lowering their risk for acquiring cardiometabolic diseases (Ceaser and Hunter, 2015).
Thinking back to sarcomeres (the smallest contractile unit in the muscle) and how they would act in type II fibers: they would obviously contract much faster in type II muscles over type I muscles; they would then obviously tear faster than type I muscles; since type II muscles are more likely to be insulin resistant, then those with a higher proportion of type II fibers need to focus more on aerobic activity to “balance out” type IIx and IIa fibers and decrease the risk of cardiometabolic disease due to more muscle contractions (Lambernd et al, 2012). Since blacks have a higher proportion of type II fibers and are more likely to be sedentary than whites, and since those who have a higher proportion of type II fibers are more likely to be obese, then it is clear that exercise can and will ameliorate some of the disparity in cardiometabolic diseases between blacks and whites.
Race, Body Fat, and Skin Folds
1250 words
Racial differences in body fat are clear to the naked eye: black women are more likely to carry more body fat than white women; Mexican American women are more likely to carry more body fat than white women, too. Different races/ethnies/genders of these races/ethnies have different formulas to assess body fat through the use of skin-folds. The sites to grasp the skin is different based on gender and race.
Body mass index (BMI) and waist circumference is overestimated in blacks, which means that they need different formulas to assess their BMI and adiposity/lean mass. Race-specific formulas/methods are needed to assess body fat and, along with it, disease risk, since blacks are more likely to be obese (black women, at least, it’s different with black American men with more African ancestry, see below). The fact of the matter is, when matched on a slew of variables, blacks had lower total and abdominal fat mass than whites.
This is even noted in Asian, black and white prepubertal children. He et al (2002) show that sex differences in body fat distribution are present in children who have yet to reach puberty and the differences in body fat in Asians is different than that from blacks and whites which also varies by sex. Asian girls had greater gynoid fat by DXA scan only, with girls having greater gynoid fat than boys. Asian girls had lower adjusted extremity fat and gynoid fat compared to white and black girls. Though, Asian boys had a lower adjusted extremity by fat as shown by DXA (a gold standard in body fat measurement) when compared to whites, but greater gynoid fat than whites and blacks.
Vickery, Cureton, and Collins, (1988), Wagner and Heyward (2000), and Robson, Bazin, and Soderstrom (1971) show that there are considerable body composition differences between blacks and whites. These differences in body composition come down to diet, of course, but there is also a genetic/physiologic component there as well. Combining the known fact that skin-fold testing is not conducive to a good estimate, black American men with more African ancestry are less likely to be obese.
Vickery, Cureton, and Collins (1988) argue that, if accurate estimates of body fat percentages are to be obtained, race-specific formulas need to be developed and used as independent variables to assess racial differences in body fat percentage. Differences in muscularity don’t seem to account for these skinfold differences, nor does greater mesomorphy. One possible explanation for differences in skinfold thickness is that blacks may store most of their body fat subcutaneously. (See Wagner and Heyward, 2000 for a review on fat patterning and body composition in blacks and whites.)
The often-used Durnin-Womersley formula which is used to predict body fat just from skin folds. However, “The 1974 DW equations did not predict %BF(DXA) uniformly in all races or ethnicities” (Davidson et al, 2011). Truesdale et al (2016) even show that numerous formulas used to estimate percent body fat are flawed, even some formulas used on different races. Most of the equations tested showed starkly different conclusions. But, this is based on NHANES data and the only data they provide regarding skin-folds is the tricep and subscapular skinfold so there may still be more problems with all of the equations used to assess body fat percentage between races. (Also see Cooper, 2010.)
Klimentidis et al (2016) show that black men—but not black women—seem to be protected against obesity and central adiposity (fat gain around the midsection) and that race negatively correlated with adiposity. The combo of male gender and West African ancestry predicted low levels of adiposity compared to black Americans with less African ancestry. Furthermore, since black men and women have—theoretically—the same SES, then cultural/social factors would not play as large a role as genetic factors in explaining the differences in adiposity between black men and black women. Black men with more African ancestry had a lower WHR and less central adiposity than black men with less African ancestry. If we assume that they had similar levels of SES and lived in similar neighborhoods, there is only one reason why this would be the case.
Klimentidis et al (2016) write:
One interpretation is that AAs are exposed to environmental and/or cultural factors that predispose them to greater obesity than EAs. Possibly, some of the genes that are inherited as part of their West-African ancestry are protective against obesity, thereby “canceling out” the obesifying effects of environment/culture, but only in men. Another interpretation is that genetic protection is afforded to all individuals of African descent, but this protection is overwhelmed by cultural and/or other factors in women.
Black men do, as is popularly believed, prefer bigger women over smaller women. For example, Freedman et al (2004) showed that black American men were more likely to prefer bigger women. Black American men “are more willing to idealize a woman
of a heavier body size, with more curves, than do their White American counterparts” (Freedman et al, 2004: 197). It is then hypothesized that black American men find these figures attractive (figures with “more curves” (Freedman et al, 2004: 197)) to protect against eating pathologies, such as anorexia and bulimia. So, it has been established that black men have thinner skin folds than whites which leads to skewed lean mass/body fat readings and black men with more African ancestry are less likely to be obese. These average differences between races, of course, contribute to differing disease acquisition.
I have covered differences in body fat in a few Asian ethnies and have come to the obvious conclusion: Asians, at the same height, weight etc as whites and blacks, will have more adipose tissue on their bodies. They, too, like blacks and whites, have different areas that need to be assessed for skin folds to estimate body fat.
Henriques (2016: 29) has a table on the equations for calculating estimated body density from skin fold measures from various populations. Of interest are the ones on blacks or ‘Hispanics‘, blacks or athletes and blacks and whites. (The table is provided from NSCA, 2008 so the references are not in the back of the text.)
For black and ‘Hispanic’ women aged 18-55 years, the sites to use for skin-folds are the chest, abdomen, triceps, subscapular, suprailiac, midaxillary, and the thigh. For blacks or athletes aged 18-61 years, the sites to use are the same as before (but a different equation is used for body fat estimation). For white women or anorexic women aged 18-55, the sites used are just triceps, suprailiac and the thigh. For black and white boys aged 6-17, only the triceps and the calf is used. It is the same for black and white girls, but, again, a different formula is used to assess body fat (Henriques, 2016: 29).
Morrison et al (2012) showed that white girls had a higher percent body fat when compared to black girls at ages 9-12 but every age after, black girls had higher percent body fat (which is related to earlier menarche in black girls since they have higher levels of body fat which means earlier puberty; Kaplowitz, 2008). Black girls, though, had higher levels of fat in their subscapular skin folds than white girls at all ages.
So, it seems, there are population-/race-specific formulas that need to be created to better assess body fat percentage in different races/ethnies and not assume that one formula/way of assessing body fat should be used for all racial/ethnic groups. According to the literature (some reviewed here and in Wagner and Heyward, 2000), these types of formulas are sorely needed to better assess health markers in certain populations. These differences in body fat percentage and distribution then have real health consequences for the races/ethnies in question.
DNA is not a “Blueprint”
2200 words
Leading behavior geneticist Robert Plomin is publishing “Blueprint: How DNA Makes Us Who We Are” in October of 2018. I, of course, have not read the book yet. But if the main thesis of the book is that DNA is a “code”, “recipe”, or “blueprint”, then that is already wrong. This is because presuming that DNA is any of the three aforementioned things marries one to certain ideas, even if they themselves do not explicitly state them. Nevertheless, Robert Plomin is what one would term a “hereditarian”, meaning that he believes that genes—more than environment—shape an individual’s psychological and other traits. (That’s a false dichotomy, though.) In the preview for the book at MIT Press, they write:
In Blueprint, behavioral geneticist Robert Plomin describes how the DNA revolution has made DNA personal by giving us the power to predict our psychological strengths and weaknesses from birth. A century of genetic research shows that DNA differences inherited from our parents are the consistent life-long sources of our psychological individuality—the blueprint that makes us who we are. This, says Plomin, is a game-changer. It calls for a radical rethinking of what makes us who were are.
Genetics accounts for fifty percent of psychological differences—not just mental health and school achievement, but all psychological traits, from personality to intellectual abilities. Nature defeats nurture by a landslide.
Plomin explores the implications of this, drawing some provocative conclusions—among them that parenting styles don’t really affect children’s outcomes once genetics is taken into effect. Neither tiger mothers nor attachment parenting affects children’s ability to get into Harvard. After describing why DNA matters, Plomin explains what DNA does, offering readers a unique insider’s view of the exciting synergies that came from combining genetics and psychology.
I won’t get into most of these things today (I will wait until I read the book for that), but this will be just an article showing that DNA is, in fact, not a blueprint, and DNA is not a “code” or “recipe” for the organism.
It’s funny that the little blurb says that “Nature defeats nurture by a landslide“, because, as I have argued at length, nature vs nurture is a false dichotomy (See Oyama, 1985, 2000, 1999; Moore, 2002; Schneider, 2007; Moore, 2017). Nature vs nurture is the battleground that the false dichotomy of genes vs environment is fought on. However, it makes no sense to partition heritability estimates if it is indeed true that genes interact with environment—that is, if nature interacts with nurture.
DNA is also called “the book of life”. For example, in her book The Epigenetics Revolution: How Modern Biology Is Rewriting Our Understanding of Genetics, Disease, and Inheritance, Nessa Carey writes that “There’s no debate that the DNA blueprint is a starting point” (pg 16). This, though, can be contested. “But the promise of a peep into the ‘book of life’ leading to a cure for all diseases was a mistake” (Noble, 2017: 161).
Developmental psychologist and cognitive scientist David S. Moore concurs. In his book The Developing Genome: An Introduction to Behavioral Epigenetics, he writes (pg 45):
So, although I will talk about genes repeatedly in this book, it is only because there is no other convenient way to communicate about contemporary ideas in molecular biology. And when I refer to gebe, I will be talking about a segment or segments of DNA containing sequence information that is used to help construct a protein (or some other product that performs a biological function). But it is worth remembering that contemporary biologists do not mean any one thing when they talk about “genes”; the gene remains a fundementally hypothetical concept to this day. The common belief that there are things inside of us that constitute a set of instructions for building bodies and minds—things that are analogous to “blueprings” or “recipes”—is undoubedtly false. Instead, DNA segements often contain information that is ambiguous, and that must be edited or arranged in context-dependent ways before it can be used.
Still, other may use terms like “genes for” trait T. This, too, is incorrect. In his outstanding book Making Sense of Genes, Kostas Kamporakis writes (pg 19):
I also explain why the notion of “genes for,” in the vernacular sense, is not only misleading but also entirely inaccurate and scientifcally illegitamate.
[…]
First, I show that genes “operate” in the context of development only. This means that genes are impllicated in the development of characters but do not determine them. Second, I explain why single genes do not alone produce characters or disease but contribute to their variation. This means that genes can account for variation in characters but cannot alone explain their origin. Third, I show that genes are not the masters of the game but are subject to complex regulatory processes.
Genes can only be seen as passive templates, not ultimate causes (Noble, 2011), and they cannot explain the origin of different characters but can account for variation in physical characters. Genes only “do” something in the context of development; they are inert molecules and thusly cannot “cause” anything on their own.
Genes are not ‘for’ traits, but they are difference-makers for traits. Sterelny and Griffiths (1999: 102), in their book Sex and Death: An Introduction to Philosophy of Biology write:
Sterelny and Griffiths (1988) responded to the idea that genes are invisible to selection by treating genes as difference makers, and as visible to selection by virtue of the differences they make. In doing so, they provided a formal reconstruction of the “gene for” locution. The details are complex, but the basic intent of the reconstruction is simple. A certain allele in humans is an “allele for brown eyes” because, in standard environments, having that allele rather than alternatives typically available in the population means that your eyes will be brown rather than blue. This is the concpet of a gene as a difference maker. It is very important to note, however, that genes are context-sensitive difference makers. Their effects depend on the genetic, cellular, and other features of their environment.
(Genes can be difference makers for physical traits, but not for psychological traits because no psychophysical laws exist, but I’ll get to that in the future.)
Note how the terms “context-sensitive” and “context-dependent” continue to appear. The DNA-as-blueprint statement presumes that DNA is context-independent, but we cannot divorce genes—whatever they are—from their context, since genes and environment, nature and nurture, are intertwined. (And it is even questioned if ‘genes’ are truly units of inheritance, see Fogle, 1990. Fogle, 2000 also argues to dispense with the concept of “gene” and that biologists should be using terms like intron, promoter region, and exon. Nevertheless, there is a huge disconnect with the term “gene” in molecular biology and classical genetics. Keller 2000 argues that there are still uses for the term “gene” and that we should not dispense with the term. I believe we should dispense with it.)
Susan Oyama (2000: 77) writes in her book The Ontogeny of Information:
“Though a plan implies action, it does not itself act, so if the genes are a blueprint, something else is the constructor-construction worker. Though blueprints are usually contrasted with building materials, the genes are quite easily conceptualized as templates for building tools and materials; once so utilized, of course, they enter the developmental process and influence its course. The point of the blueprint analogy, though, does not seem to be to illuminate developmental processes, but rather to assume them and, in celebrating their regularity, to impute cognitive functions to genes. How these functions are exercised is left unclear in this type of metaphor, except that the genetic plan is seen in some peculiar way to carry itself out, generating all the necessary steps in the necessary sequence. No light is shed on multiple developmental possibilities, species-typical or atypical.“
The Modern Synthesis is one of the causes for the genes-as-blueprints thinking; the Modern Synthesis has causation in biology wrong. Genes are not active causes, but they are passive templates, as argued by many authors. They, thus, cannot “cause” anything on their own.
In his 2017 book Dance to the Tune of Life: Biological Relativity, Denis Noble writes (pg 157):
As we saw earlier in this chapter, these triplet sequences are formed from any combination of the four bases U, C, A and G in RNA and T, C, A and G in DNA. They are often described as a genetic ‘code’, but it is important to understand that this usage of the word ‘code’ carries overtones that can be confusing.
A code was originally an intentional encryption used by humans to communicate. The genetic ‘code’ is not intentional in that sense. The word ‘code’ has unfortunately reinforced the idea that genes are active and even complete causes, in much the same was as a computer is caused to follow the instructions of a computer program. The more nuetral word ‘template’ would be better. Templates are used only when required (activated); they are not themselves active causes. The active causes lie within the cells themselves since they determine the expression patterns for the different cell types and states. These patterns are comminicated to the DNA by transcrption factors, by methylation patterns and by binding to the tails of histones, all of which influence the pattern and speed of transcription of different parts of the genome. If the word ‘instruction’ is useful here at all, it is rather that the cell instructs the genome. As Barbara McClintock wrote in 1984 after receiving her Nobel Prize, the genome is an ‘organ of the cell’, not the other way around.
Realising that DNA is under the control of the system has been reinforced by the discovery that cells use different start, stop and splice sites for producing different messenger RNAs from a single DNA sequence. This enables the same sequence to code different proteins in different cell types and under different conditions [here’s where context-dependency comes into play again].
Representing the direction of causality in biology the wrong way round is therefore confusing and has far-reaching conseqeunces. The causality is circular, acting both ways: passive causality by DNA sequences acting as otherwise inert templates, and active causality by the functional networks of interactions that determine how the genome is activated.
This takes care of the idea that DNA is a ‘code’. But what about DNA being a ‘blueprint’, that all of the information is contained in the DNA of the organism before conception? DNA is clearly not a ‘program’, in the sense that all of the information to construct the organism exists already in DNA. The complete cell is also needed, and its “complex structures are inherited by self-templating” (Noble, 2017: 161). Thus, the “blueprint” is the whole cell, not just the genome itself (remember that the genome is an organ of the cell).
Lastly, GWA studies have been all the rage recently. However, there is only so much we can learn just from association studies, before we need to turn to the physiological sciences for functional analyses. Indeed, Denis Noble (2018) writes in a new editorial:
As with the results of GWAS (genome-wide association studies) generally, the associations at the genome sequence level are remarkably weak and, with the exception of certain rare genetic diseases, may even be meaningless (13, 21). The reason is that if you gather a sufficiently large data set, it is a mathematical necessity that you will find correlations, even if the data set was generated randomly so that the correlations must be spurious. The bigger the data set, the more spurious correlations will be found (3).
[…]
The results of GWAS do not reveal the secrets of life, nor have they delivered the many cures for complex diseases that society badly needs. The reason is that association studies do not reveal biological mechanisms. Physiology does. Worse still, “the more data, the more arbitrary, meaningless and useless (for future action) correlations will be found in them” is a necessary mathematical statement (3).
Nor does applying a highly restricted DNA sequence-based interpretation of evolutionary biology, and its latest manifestation in GWAS, to the social sciences augur well for society.
It is further worth noting that there is no privileged level of causation in biological systems (Noble, 2012)—a priori, there is no justification to privilege one system over another in regard to causation, so saying that one level of the organism is “higher” than another (for instance, saying that genes are, and should be, privileged over the environment or any other system in the organism regarding causation) is clearly false, since there is upwards and downwards causation, influencing all levels of the system.
In sum, it is highly misleading to refer to DNA as “blueprints”, a “code”, or a “recipe.” Referring to DNA in this way means that one presumes that DNA can be divorced from its context—that it does not work together with the environment. As I have argued in the past, association studies will not elucidate genetic mechanisms, nor will heritability estimates (Richardson, 2012). We need physiological testing for these functional analyses, and association studies like GWAS and even heritability estimates don’t tell us this type of information (Panofsky, 2014). So, it seems, that what Plomin et al are looking for that they assume are “in the genes”, are not there, because they use a false model of the gene (Burt, 2015; Richardson, 2017). Genes are resources—templates to be used by and for the system—not causes of traits and development. They can account for differences in variation, but cannot be said to be the origin of trait differences. Genes can be said to be difference makers, but knowing whether or not they are difference makers for behavior, in my opinion, cannot be known.
(For further information on genes and what they do, reach Chapters Four and Five of Ken Richardson’s book Genes, Brains, and Human Potential: The Science and Ideology of Intelligence. Plomin himself seems to be a reductionist, and Richardson took care of that paradigm in his book. Lickliter (2018) has a good review of the book, along with critiques of the reductionist paradigm that Plomin et al follow.)
Genotypes, Athletic Performance, and Race
2050 words
Everyone wants to know the keys to athletic success, however, as I have argued in the past, to understand elite athletic performance, we must understand how the system works in concert with everything—especially in the environments the biological system finds itself in. To reduce factors down to genes, or training, or X or Y does not make sense; to look at what makes an elite athlete, the method of reductionism, while it does allow us to identify certain differences between athletes, it does not allow us to appreciate the full-range of how and why elite athletes differ in their sport of choice. One large meta-analysis has been done on the effects of a few genotypes on elite athletic performance, and it shows us what we already know (blacks are more likely to have the genotype associated with power performance—so why are there no black Strongmen or any competitors in the World’s Strongest Man?). A few studies and one meta-analysis exist, attempting to get to the bottom of the genetics of elite athletic performance and, while it of course plays a factor, as I have argued in the past, we must take a systems view of the matter.
One 2013 study found that a functional polymorphism in the angiotensinogen (ATG) region was 2 to 3 times more common in elite power athletes than in (non-athlete) controls and elite endurance athletes (Zarebska et al, 2013). This sample tested was Polish, n = 223, 156 males, 67 females, and then they further broke down their athletic sample into tiers. They tested 100 power athletes (29 100-400 m runners; 22 powerlifters; 20 weightlifters; 14 throwers and 15 jumpers) and 123 endurance athletes (4 tri-athletes; 6 race walkers; 14 road cyclists; 6 15 to 50 m cross-country skiers; 12 marathon runners; 53 rowers; 17 3 to 10 km runners; and 11 800 to 1500 m swimmers).
Zarebska et al (2013) attempted to replicate previous associations found in other studies (Buxens et al, 2009) most notably the association with the M235T polymorphism in the AGT (angiotensinogen) gene. Zarebska et al’s (2013) main finding was that there was a higher representation of elite power athletes with the CC and C alleles of the M235T polymorphism compared with endurance athletes and controls, which suggests that the C allele of the M235T gene “may be associated with a predisposition to power-oriented
events” (Zarebska et al, 2013: 2901).
Elite power athletes were more likely to possess the CC genotype; 40 percent of power athletes had the genotype whereas 13 percent of endurance had it and 18 percent of non-athletes had it. So power athletes were more than three times as likely to have the CC genotype, compared to endurance athletes and twice as likely to have it compared to non-athletes. On the other hand, one copy of the C allele was found in 55 percent of the power athletes whereas, for the endurance athletes and non-athletes, the C allele was found in about 40 percent of individuals. (Further, in the elite anaerobic athlete, explosive power was consistently found to be a difference maker in predicting elite sporting performance; Lorenz et al, 2013.)
Now we come to the more interesting parts: ethnic differences in the M235T polymorphism. Zarebska et al (2013: 2901-2902) write:
The M235T allele distribution varies widely according to the subject’s ethnic origin: the T235 allele is by far the most frequent in Africans (;0.90) and in African-Americans (;0.80). It is also high in the Japanese population (0.65–0.75). The T235 (C4027) allele distribution of the control participants in our study was lower (0.40) but was similar to that reported among Spanish Caucasians (0.41), as were the sports specialties of both the power athletes (throwers, sprinters, and jumpers) and endurance athletes (marathon runners, 3- to 10-km runners, and road cyclists), thus mirroring the aforementioned studies.
Zarebska et al (2013: 2902) conclude that their study—along with the study they replicated—supports the hypothesis that the C allele of the M235T polymorphism in the AGT gene may confer a competitive advantage in power-oriented sports, which is partly mediated through ANGII production in the skeletal muscles. Mechanisms can explain the mediation of ANGII production in skeletal muscles, such as a direct skeletal muscle hypertrophic effect, along with the redistribution of between muscle blood flow between type I (slow twitch) and II fibers (fast twitch), which would then augment power and speed. However, it is interesting to note that Zarebska et al (2013) did not find any differences between “top-elite” level athletes who had won medals in international competitions compared to elite-level athletes who were not medalists.
The big deal about this gene is that the AGT gene is part of the renin-angiotensin system which is partly responsible for blood pressure and body salt regulation (Hall, 1991; Schweda, 2014). There seems to be an ethnic difference in this polymorphism, and, according to Zarebska et al (2013), African Americans and Africans are more likely to have the polymorphisms that are associated with elite power performance.
There is also a meta-analysis on genotyping and elite power athlete performance (Weyerstrab et al, 2017). Weyerstrab et al (2017) meta-analyzed 36 studies which attempted to find associations between genotype and athletic ability. One of the polymorphisms studied was the famous ACTN3. It has been noted that, when conditions are right (i.e., the right morphology), the combined effects of morphology along with the contractile properties of the individual muscle fibers contribute to the enhanced performance of those with the RR ACTN3 genotype (Broos et al, 2016), while Ma et al (2013) also lend credence to the idea that genetics influences sporting performance. This is, in fact, the most-replicated association in regard to elite sporting performance: we know the mechanism behind how muscle fibers contract; we know how the fibers contract and the morphology needed to maximize the effectiveness of said fast twitch fibers (type II fibers). (Blacks have a higher proportion of type II fibers [see Caeser and Henry, 2015 for a review].)
Weyerstrab et al (2017) meta-analyzed 35 articles, finding significant associations with genotype and elite power performance. They found that ten polymorphisms were significantly associated with power athlete states. Their most interesting findings, though, were on race. Weyerstrab et al (2017: 6) write:
Results of this meta-analysis show that US African American carriers of the ACE AG genotype (rs4363) were more than two times more likely to become a power athlete compared to carriers of the ACE preferential genotype for power athlete status (AA) in this population.
“Power athlete” does not necessarily have to mean “strength athlete” as in powerlifters or weightlifters (more on weightlifters below).
Lastly, the AGT M235T polymorphism, while associated with other power movements, was not associated with elite weightlifting performance (Ben-Zaken et al, 2018). As noted above, this polymorphism was observed in other power athletes, and since these movements are largely similar (short, explosive movements), one would rightly reason that this association should hold for weightlifters, too. However, this is not what we find.
Weightlifting, compared to other explosive, power sports, is different. The beginning of the lifts take explosive power, but during the ascent of the lift, the lifter moves the weight slower, which is due to biomechanics and a heavy load. Ben-Zaken et al (2018) studied 47 weightlifters (38 male, 9 female) and 86 controls. Every athlete that was studied competed in national and international meets on a regular basis. Thirty of the weightlifters were also classified as “elite”, which entails participating in and winning national and international competitions such as the Olympics and the European and World Championships).
Ben-Zaken et al (2018) did find that weightlifters had a higher prevalence of the AGT 235T polymorphism when compared to controls, though there was no difference in the prevalence of this polymorphism when elite and national-level competitors were compared, which “[suggests] that this polymorphism cannot determine or predict elite competitive weightlifting performance” (Ben-Zaken et al, 2018: 38). Of course, a favorable genetic profile is important for sporting success, though, despite the higher prevalence of AGT in weightlifters compared to controls, this could not explain the difference between national and elite-level competitors. Other polymorphisms could, of course, contribute to weightlifting success, variables “such as training experience, superior equipment and facilities, adequate nutrition, greater familial support, and motivational factors, are crucial for top-level sports development as well” (Ben-Zaken et al, 2018: 39).
I should also comment on Anatoly Karlin’s new article The (Physical) Strength of Nations. I don’t disagree with his main overall point; I only disagree that grip strength is a good measure of overall strength—even though it does follow the expected patterns. Racial differences in grip strength exist, as I have covered in the past. Furthermore, there are associations between muscle strength and longevity, with stronger men being more likely to live longer, fuller lives (Ruiz et al, 2008; Volkalis, Haille, and Meisinger, 2015; Garcia-Hermosa, et al, 2018) so, of course, strength training can only be seen as a net positive, especially in regard to living a longer and fuller life. Hand grip strength does have a high correlation with overall strength (Wind et al, 2010; Trosclair et al, 2011). While handgrip strength can tell you a whole lot about your overall health (Lee et al, 2016), of course, there is no better proxy than actually doing the lifts/exercises to ascertain one’s level of strength.
There are replicated genetic associations between explosive, powerful athletic performance, along with even the understanding of the causal mechanisms behind the polymorphisms and their carry-over to power sports. We know that if morphology is right and the individual has the RR ACTN3 genotype, that they will exceed in explosive sports. We know the causal pathways of ACTN3 and how it leads to differences in sprinting competitions. It should be worth noting that, while we do know a lot more about the genomics of sports than we did 20, even 10 years ago, current genetic testing has zero predictive power in regard to talent identification (Pitsladis et al, 2013).
So, of course, for parents and coaches who wonder about the athletic potential of their children and students, the best way to gauge whether or not they will excel in athletics is…to have them compete and compare them to other kids. Even if the genetics aspect of elite power performance is fully unlocked one day (which I doubt it will be), the best way to ascertain whether or not one will excel in a sport is to put them to the test and see what happens. We are in our infancy in understanding the genomics of sporting performance, but when we do understand which genotypes are more prevalent in regard to certain sports (and of course the interactions of the genotype with the environment and genes), then we can better understand how and why others are better in certain sports.
The genomics of elite sporting performance is very interesting; however, the answer that reductionists want to see will not appear: genes are difference makers (Sterelny and Griffith, 1999), not causes, and along with a whole slew of other environmental and mental factors (Lippi, Favaloro, and Guidi 2008), along with a favorable genetic profile with sufficient training (and everything else that comes along with it) are needed for the athlete to reach their maximum athletic potential (see Guth and Roth, 2013). Genetic and environmental differences between individuals and groups most definitely explain differences in elite sporting performance, though elucidating what causes what and the mechanisms that cause the studied trait in question will be tough.
Just because group A has gene or gene networks G and they compete in competition C does not mean that gene or gene networks G contribute in full—or in part—to sporting success. The correlations could be coincidental and non-functional in regard to the sport in question. Athletes should be studied in isolation, meaning just studying a specific athlete in a specific discipline to ascertain how, what, and why works for the specific athlete along with taking anthropomorphic measures, seeing how bad they want “it”, and other environmental factors such as nutrition and training. Looking at the body as a system will take us away from privileging one part over another—while we also do understand that they do play a role but not the role that reductionists believe.
These studies, while they attempt to show us how genetic factors cause differences at the elite level in power sports, they will not tell the whole story, because we must look at the whole system, not reduce it down to the sum of its parts (Shenk, 2011: chapter 5). While blacks are more likely to have these polymorphisms that are associated with elite power athlete performance, this does not obviously carry over to strongman and powerlifting competition.
Somatotyping, Constitutional Psychology, and Sports
1600 words
In the 1940s, psychologist William Sheldon created a system of body measures known as “somatotyping”, then took his somatotypes and attempted to classify each soma (endomorph, ectomorph, or mesomorph) to differing personality types. It was even said that “constitutional psychology can guide a eugenics program and save the modern world from itself.”
Sheldon attempted to correlate different personality dimensions to different somas. But his somas fell out of favor before being revived by two of his disciples—without the “we-can-guess-your-personality-from-your-body-type” canard that Sheldon used. Somatotyping, while of course being put to use in a different way today compared to what it was originally created for, it gives us reliable dimensions for human appendages and from there we can ascertain what a given individual would excel at in regard to sporting events (obviously this is just on the basis of physical measures and does not measure the mind one needs to excel in sports).
The somatotyping system is straightforward: You have three values, say at 1-1-7; the first refers to endomorphy, the second refers to mesomorphy and the third refers to ectomorphy, therefore a 1-1-7 would be an extreme ectomorph. However, few people are at the extreme end of each soma, and most people have a combination of two or even all three of the somas.
According to Carter (2002): “The somatotype is defined as the quantification of the present shape and composition of the human body.” So, obviously, somas can change over time. However, it should be noted that the somatotype is, largely, based on one’s musculoskeletal system. This is where the appendages come in, along with body fat, wide and narrow clavicles and chest etc. This is why the typing system, although it began as a now-discredited method, should still be used today since we do not use the pseudoscientific personality measures with somatotyping.
Ectomorphs are long and lean, lanky, you could say. They have a smaller, narrower chest and shoulders, along with longer arms and legs, and have a hard time gaining weight, and a short upper body (I’d say they have a harder time gaining weight due to a slightly faster metabolism, in the variation of the normal range of metabolism, of course). Put simply, ectomorphs are just skinny and lanky with less body fat than mesos and endos. Human races that fit this soma are East Africans and South Asians (see Dutton and Lynn, 2015; one of my favorite papers from Lynn for obvious reasons).
Endomorphs are stockier, shorter and have wider hips, along with short limbs, a wider trunk, more body fat and can gain muscular strength easier than the other somas. Thus, endos, being shorter than ectos and mesos, have a lower center of gravity, along with shorter arms. Thus, we should see that these somas dominate strongman competitions and this is what we see. Pure strength competitions are perfect for this type, such as Strongman competitions and powerlifting. Races that generally conform to this type are East Asians, Europeans, and Pacific Islanders (see Dutton and Lynn, 2015).
Finally, we have mesomorphs (the “king” of all of the types). Mesos are more muscular on average than the two others, they have less body fat than endos but more body fat than ectos; they have wider shoulders, chest and hips, a short trunk and long limbs. The most mesomorphic races are West Africans (Malina, 1969), and due to their somatotype they can dominate sprinting competitions; they also have thinner skin folds (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000), and so they would have an easier time excelling at running competitions but not at weightlifting, powerlifting, or Strongman (see Dutton and Lynn, 2015).
These anatomic differences between the races of man are due to climatic adaptations. The somatypic differences Neanderthals and Homo sapiens mirror the somatotype difference between blacks and whites; since Neanderthals were cold-adapted, they were shorter, had wider pelves and could thusly generate more power than the heat-adapted Homo sapiens who had long limbs and narrow pelvis to better dissipate heat. Either way, we can look at the differences in somatotype between races that evolved in Europe and Africa to ascertain the somatotype of Neanderthals—and we also have fossil evidence for these claims, too (see e.g., Weaver and Hublin, 2009; Gruss and Schmitt, 2016)
Now, just because somatotyping, during its conception, was mixed with pseudoscientific views about differing somas having differing psychological types, does not mean that these differences in body type do not have any bearing on sporting performance. We can chuck the “constitutional psychology” aspect of somatotyping and just keep the anthropometric measures, and, along with the knowledge of human biomechanics, we can then discuss, in a scientific manner, why one soma would excel in sport X or why one soma would not excel in sport X. Attempting to argue that since somatotyping began as some crank psuedoscience does not mean that it is not useful today, since we do not ascribe inherent psychological differences to these somas (I’d claim that saying that this soma has a harder time gaining weight compared to that soma is not ascribing a psychological difference to the soma; it is taking physiologically and on average we can see that different somas have different propensities for weight gain).
In her book Straightening the Bell Curve: How Stereotypes about Black Masculinity Drive Research about Race and Intelligence, Hilliard (2012: 21) discusses the pitfalls of somatotyping and how Sheldon attempted to correlate personality measures with his newfound somatotypes:
As a young graduate student, he [Richard Herrnstein] had fallen under the spell of Harvard professor S. S. Stevens, who had coauthored with William Sheldon a book called The Varieties of Temperament: A Psychology of Constitutional Differences, which popularized the concept of “somatotyping,” first articulated by William Sheldon. This theory sought, through the precise measurement and analysis of human body types, to establish correlations comparing intelligence, temperament, sexual proclivities, and the moral worth of individuals. Thus, criminals were perceived to be shorter and heavier and more muscular than morally upstanding citizens. Black males were reported to rank higher on the “masculine component” scale than white males did, but lower in intelligence. Somatotyping lured the impressionable young Herrnstein into a world promising precision and human predictability based on the measuring of body parts.
Though constitutional psychology is now discredited, there may have been something to some of Sheldon’s theories. Ikeda et al (2018: 3) conclude in their paper, Re-evaluating classical body type theories: genetic correlation between psychiatric disorders and body mass index, that “a trans-ancestry meta-analysis of the genetic correlation between psychiatric disorders and BMI indicated that the negative correlation with SCZ supported classical body type theories proposed in the last century, but found a negative correlation between BD and BMI, opposite to what would have been predicted.” (Though it should be noted that SCZ is a, largely if not fully, environmentally-induced disorder, see Joseph, 2017.)
These different types (i.e., the differing limb lengths/body proportions) have implications for sporting performance. Asfaw and A (2018) found that Ethiopian women high jumpers had the highest ectomorph values whereas long and triple jumpers were found to be more mesomorphic. Sports good for ectos are distance running due to their light frame, tennis etc—anything that the individual can use their light frame as an advantage. Since they have longer limbs and a lighter frame, they can gain more speed in the run up to the jump, compared to endos and mesos (who are heavier). This shows why ectos have a biomechanical advantage when it comes to high jumping.
As for mesomorphs, the sports they excel at are weightlifting, powerlifting, strongman, football, rugby etc. Any sport where the individual can use their power and heavier bone mass will they excel in. Gutnik et al (2017) even concluded that “These results suggest with high probability that there is a developmental tendency of change in different aspects of morphometric phenotypes of selected kinds of sport athletes. These phenomena may be explained by the effects of continuous intensive training and achievement of highly sport-defined shapes.” While also writing that mesomorphy could be used to predict sporting ability.
Finally, for endomorphs, they too would excel in weightlifting, powerlifting, and strongman, but do on average better since they have different levers (i.e., shorter appendages so they can more weight and a shorter amount of time in comparison to those with longer limbs like ectos).
Thus, different somatotypes excel in different sports. Different races and ethnies have differing somatotypes (Dutton and Lynn, 2015), so these different bodies that the races have, on average, is part of the cause for differences in sporting ability. That somatotyping began as a pseudoscientific endeavor 70 years ago does not mean that it does not have a use in today’s world—because it clearly does due to the sheer amount of papers on the usefulness of somatotyping and relating differences in sporting performance due to somatotyping. For example, blacks have thinner skin folds (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000) which is due to their somatotype, which is then due to the climate their ancestors evolved in.
Somatotyping can show us the anthropometric reasons for how and why certain individuals, ethnies, and races far-and-away dominate certain sporting events. It is completely irrelevant that somatotyping began as a psychological pseudoscience (what isn’t in psychology, am I right?). Understanding anthropometric differences between individuals and groups will help us better understand the evolution of these somas along with how and why these somas lead to increased sporting performance in certain domains. Somatotyping has absolutely nothing to do with “intelligence” nor how morally upstanding one is. I would claim that somatotyping does have an effect on one’s perception of masculinity, and thus more masculine people/races would tend to be more mesomorphic, which would explain what Hilliard (2012) discussed when talking about somatotyping and the attempts to correlate differing psychological tendencies to each type.
