
Home » Posts tagged 'Racial Differences' (Page 2)
Tag Archives: Racial Differences
Somatotyping, Constitutional Psychology, and Sports
1600 words
In the 1940s, psychologist William Sheldon created a system of body measures known as “somatotyping”, then took his somatotypes and attempted to classify each soma (endomorph, ectomorph, or mesomorph) to differing personality types. It was even said that “constitutional psychology can guide a eugenics program and save the modern world from itself.”
Sheldon attempted to correlate different personality dimensions to different somas. But his somas fell out of favor before being revived by two of his disciples—without the “we-can-guess-your-personality-from-your-body-type” canard that Sheldon used. Somatotyping, while of course being put to use in a different way today compared to what it was originally created for, it gives us reliable dimensions for human appendages and from there we can ascertain what a given individual would excel at in regard to sporting events (obviously this is just on the basis of physical measures and does not measure the mind one needs to excel in sports).
The somatotyping system is straightforward: You have three values, say at 1-1-7; the first refers to endomorphy, the second refers to mesomorphy and the third refers to ectomorphy, therefore a 1-1-7 would be an extreme ectomorph. However, few people are at the extreme end of each soma, and most people have a combination of two or even all three of the somas.
According to Carter (2002): “The somatotype is defined as the quantification of the present shape and composition of the human body.” So, obviously, somas can change over time. However, it should be noted that the somatotype is, largely, based on one’s musculoskeletal system. This is where the appendages come in, along with body fat, wide and narrow clavicles and chest etc. This is why the typing system, although it began as a now-discredited method, should still be used today since we do not use the pseudoscientific personality measures with somatotyping.
Ectomorphs are long and lean, lanky, you could say. They have a smaller, narrower chest and shoulders, along with longer arms and legs, and have a hard time gaining weight, and a short upper body (I’d say they have a harder time gaining weight due to a slightly faster metabolism, in the variation of the normal range of metabolism, of course). Put simply, ectomorphs are just skinny and lanky with less body fat than mesos and endos. Human races that fit this soma are East Africans and South Asians (see Dutton and Lynn, 2015; one of my favorite papers from Lynn for obvious reasons).
Endomorphs are stockier, shorter and have wider hips, along with short limbs, a wider trunk, more body fat and can gain muscular strength easier than the other somas. Thus, endos, being shorter than ectos and mesos, have a lower center of gravity, along with shorter arms. Thus, we should see that these somas dominate strongman competitions and this is what we see. Pure strength competitions are perfect for this type, such as Strongman competitions and powerlifting. Races that generally conform to this type are East Asians, Europeans, and Pacific Islanders (see Dutton and Lynn, 2015).
Finally, we have mesomorphs (the “king” of all of the types). Mesos are more muscular on average than the two others, they have less body fat than endos but more body fat than ectos; they have wider shoulders, chest and hips, a short trunk and long limbs. The most mesomorphic races are West Africans (Malina, 1969), and due to their somatotype they can dominate sprinting competitions; they also have thinner skin folds (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000), and so they would have an easier time excelling at running competitions but not at weightlifting, powerlifting, or Strongman (see Dutton and Lynn, 2015).
These anatomic differences between the races of man are due to climatic adaptations. The somatypic differences Neanderthals and Homo sapiens mirror the somatotype difference between blacks and whites; since Neanderthals were cold-adapted, they were shorter, had wider pelves and could thusly generate more power than the heat-adapted Homo sapiens who had long limbs and narrow pelvis to better dissipate heat. Either way, we can look at the differences in somatotype between races that evolved in Europe and Africa to ascertain the somatotype of Neanderthals—and we also have fossil evidence for these claims, too (see e.g., Weaver and Hublin, 2009; Gruss and Schmitt, 2016)
Now, just because somatotyping, during its conception, was mixed with pseudoscientific views about differing somas having differing psychological types, does not mean that these differences in body type do not have any bearing on sporting performance. We can chuck the “constitutional psychology” aspect of somatotyping and just keep the anthropometric measures, and, along with the knowledge of human biomechanics, we can then discuss, in a scientific manner, why one soma would excel in sport X or why one soma would not excel in sport X. Attempting to argue that since somatotyping began as some crank psuedoscience does not mean that it is not useful today, since we do not ascribe inherent psychological differences to these somas (I’d claim that saying that this soma has a harder time gaining weight compared to that soma is not ascribing a psychological difference to the soma; it is taking physiologically and on average we can see that different somas have different propensities for weight gain).
In her book Straightening the Bell Curve: How Stereotypes about Black Masculinity Drive Research about Race and Intelligence, Hilliard (2012: 21) discusses the pitfalls of somatotyping and how Sheldon attempted to correlate personality measures with his newfound somatotypes:
As a young graduate student, he [Richard Herrnstein] had fallen under the spell of Harvard professor S. S. Stevens, who had coauthored with William Sheldon a book called The Varieties of Temperament: A Psychology of Constitutional Differences, which popularized the concept of “somatotyping,” first articulated by William Sheldon. This theory sought, through the precise measurement and analysis of human body types, to establish correlations comparing intelligence, temperament, sexual proclivities, and the moral worth of individuals. Thus, criminals were perceived to be shorter and heavier and more muscular than morally upstanding citizens. Black males were reported to rank higher on the “masculine component” scale than white males did, but lower in intelligence. Somatotyping lured the impressionable young Herrnstein into a world promising precision and human predictability based on the measuring of body parts.
Though constitutional psychology is now discredited, there may have been something to some of Sheldon’s theories. Ikeda et al (2018: 3) conclude in their paper, Re-evaluating classical body type theories: genetic correlation between psychiatric disorders and body mass index, that “a trans-ancestry meta-analysis of the genetic correlation between psychiatric disorders and BMI indicated that the negative correlation with SCZ supported classical body type theories proposed in the last century, but found a negative correlation between BD and BMI, opposite to what would have been predicted.” (Though it should be noted that SCZ is a, largely if not fully, environmentally-induced disorder, see Joseph, 2017.)
These different types (i.e., the differing limb lengths/body proportions) have implications for sporting performance. Asfaw and A (2018) found that Ethiopian women high jumpers had the highest ectomorph values whereas long and triple jumpers were found to be more mesomorphic. Sports good for ectos are distance running due to their light frame, tennis etc—anything that the individual can use their light frame as an advantage. Since they have longer limbs and a lighter frame, they can gain more speed in the run up to the jump, compared to endos and mesos (who are heavier). This shows why ectos have a biomechanical advantage when it comes to high jumping.
As for mesomorphs, the sports they excel at are weightlifting, powerlifting, strongman, football, rugby etc. Any sport where the individual can use their power and heavier bone mass will they excel in. Gutnik et al (2017) even concluded that “These results suggest with high probability that there is a developmental tendency of change in different aspects of morphometric phenotypes of selected kinds of sport athletes. These phenomena may be explained by the effects of continuous intensive training and achievement of highly sport-defined shapes.” While also writing that mesomorphy could be used to predict sporting ability.
Finally, for endomorphs, they too would excel in weightlifting, powerlifting, and strongman, but do on average better since they have different levers (i.e., shorter appendages so they can more weight and a shorter amount of time in comparison to those with longer limbs like ectos).
Thus, different somatotypes excel in different sports. Different races and ethnies have differing somatotypes (Dutton and Lynn, 2015), so these different bodies that the races have, on average, is part of the cause for differences in sporting ability. That somatotyping began as a pseudoscientific endeavor 70 years ago does not mean that it does not have a use in today’s world—because it clearly does due to the sheer amount of papers on the usefulness of somatotyping and relating differences in sporting performance due to somatotyping. For example, blacks have thinner skin folds (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000) which is due to their somatotype, which is then due to the climate their ancestors evolved in.
Somatotyping can show us the anthropometric reasons for how and why certain individuals, ethnies, and races far-and-away dominate certain sporting events. It is completely irrelevant that somatotyping began as a psychological pseudoscience (what isn’t in psychology, am I right?). Understanding anthropometric differences between individuals and groups will help us better understand the evolution of these somas along with how and why these somas lead to increased sporting performance in certain domains. Somatotyping has absolutely nothing to do with “intelligence” nor how morally upstanding one is. I would claim that somatotyping does have an effect on one’s perception of masculinity, and thus more masculine people/races would tend to be more mesomorphic, which would explain what Hilliard (2012) discussed when talking about somatotyping and the attempts to correlate differing psychological tendencies to each type.
Blumenbachian Partitions and Mimimalist Races
2100 words
Race in the US is tricky. On one hand, we socially construct races. On the other, these socially constructed races have biological underpinnings. Racial constructivists, though, argue that even though biological races are false, races have come into existence—and continue to exist—due to human culture and human decisions (see the SEP). Sound arguments exist for the existence of biological races. Biological races exist, and they are real. One extremely strong view is from philosopher of science Quayshawn Spencer. In his paper A Radical Solution to the Race Problem, Spencer (2014) argues that biological races are real; that the term “race” directly refers; that race denotes proper names, not kinds; and these sets of human populations denoted by Americans can be denoted as a partition of human populations which Spencer (2014) calls “the Blumenbach partition”.
To begin, Spencer (2014) defines “referent”: “If, by using appropriate evidential methods (e.g., controlled experiments), one finds that a term t has a logically inconsistent set of identifying conditions but a robust extension, then it is appropriate to identify the meaning
of t as just its referent.” What he means is that the word “race” is just a referent, which means that the term “race” lies in what points out in the world. So, what “race” points out in the world becomes clear if we look at how Americans define “race”.
Spencer (2014) assumes that “race” in America is the “national meaning” of race. That is, the US meaning of race is just the referent to the Census definitions of race, since race-talk in America is tied to the US Census. But the US Census Bureau defers to the Office of Management and Budget (OMB). Therefore, since the US Census Bureau defers to the OMB on matters of race, and since Americans defer to the US Census Bureau, then Americans use the OMB definitions of race.
The OMB describes a “comprehensive set” of categories (according to the OMB) which lead Spencer (2014) to believe that the OMB statements on race are pinpointing Caucasians, Africans, Pacific Islanders, East Asians, and Amerindians. Spencer (2014: 1028-29) thusly claims that race in America “is a term that rigidly designates a particular set of “population groups.” Now, of course, the question is this: are these population groups socially constructed? Do they really exist? Are the populations identified arbitrary? Of course, the answer is that they identify a biologically real set of population groups.
To prove the existence of his Blumenbachian populations, Spencer (2014) invokes populational genetic analyses. Population geneticists first must make the assumption at how many local populations exist in the target species. According to Spencer, “The current estimate for humans is 7,105 ethnic groups, half of which are in Africa and New Guinea.” After the assumptions are made, the next step is to sample the species’ estimated local populations. Then they must test noncoding DNA sequences. Finally, they must attempt to partition the sample so that each partition at each level is unique which then minimizes genetic differences in parts and maximizes genetic differences among parts. There are two ways of doing this: using structure and PCA. For the purposes of this argument, Spencer (2014) chooses structure, invoking a 5-population racial model, (see e.g., Rosenberg et al, 2002).
K = 5 corresponds to 5 populational clusters which denote Africans, Oceanians, East Asians, Amerindians, and Caucasians (Spencer, 2014; Hardimon, 2017b). K = 5 shows that the populations in question are genetically structured—that is, meaningfully demarcated on the basis of genetic markers and only genetic markers. Thus, that the populations in question are meaningfully demarcated on the basis of genetic markers, this is evidence that Hardimon’s (2017b) minimalist races are a biological reality. Furthermore, since Rosenberg et al (2002) used microsatellite markers in their analysis, this is a nonarbitrary way of constructing genetic clusters which then demarcate the continental-level minimalist races (Hardimon, 2017b: 90).
Thus, Spencer (2014) argues to call the partition identified in K = 5 “the Blumenbachian partition” in honor of Johann Blumenbach, anthropologist, physician, physiologist, and naturalist. (Though it should be noted that one of his races “Malays” was not a race, but Oceaninans are, so he “roughly discovered” the population partition.) So we can say that “the Blumenbach partition” is just the US meaning of “race”, the partitions identified by K = 5 (Rosenberg et al, 2002).
Furthermore, like Lewontin (1972), Rosenberg et al (2002) found that a majority of human genetic variation is between individuals, not races. That is, Rosenberg et al (2002) found that only 4.3 percent of human genetic variation was found to lie between the continental-level minimalist races. Thus, minimalist races are a biological kind, “if only a modest one” (Hardimon, 2017b: 91). Thus, Rosenberg et al (2002) support the contention that minimalist races exist and are a biological reality since a fraction of human population variation is due to differences among continental-level minimalist races (Africans, Caucasians, East Asians, Oceanians, and Amerindians). The old canard is true, there really is more genetic variation within races than between them, but, as can be seen, that does not rail against the reality of race, since that small amount of genetic variation shows that humanity is meaningfully clustered in a genetic sense.
Spencer (2014: 1032) then argues why Blumenbachian populations are “race” in the American sense:
It is not hard to generate accessible possible worlds that support the claim that US race terms are just aliases for Blumenbachian populations. For example, imagine a possible world τ where human history unfolded exactly how it did in our world except that every Caucasian in τ was killed by an infectious disease in the year 2013. Presumably, we have access to τ, since it violates no logical, metaphysical, or scientific principles. Then, given that we use ‘white’ in its national American meaning in our world, and given that we use ‘Caucasian’ in its Blumenbachian meaning in our world, it is fair to say that both ‘Caucasian’ and ‘white’ are empty terms in τ in 2014—which makes perfect sense if ‘white’ is just an alias for Caucasians. It is counterfactual evidence like this that strongly suggests that the US meaning of ‘race’ is just the Blumenbach partition.
Contrary to critics, this partition is biologically real and demarcates the five genetically structured populations of the human race. Rosenberg et al (2005) found that if sufficient data are used, “the geographic distribution of the sampled individuals has little effect on the analysis“, while their results verify that genetic clusters “arise from genuine features of the underlying pattern of human genetic variation, rather than as artifacts of uneven sampling along continuous gradients of allele frequencies.”
Some may claim that K = 5 is “arbitrary”, however, constructing genetic clusters using microsatellites is nonarbitrary (Hardimon, 2017b: 90):
Constructing genetic clusters using microsatellites constitutes a nonarbitrary way of demarcating the boundaries of continental-level minimalist races. And the fact that it is possible to construct genetic clusters corresponding to continental-level minimalist races in a nonarbitrary way is itself a reason for thinking that minimalist race is biologically real 62.
It should also be noted that Hardimon writes in note 62 (2017b: 197):
Just to be perfectly clear, I don’t think that the results of the 2002 Rosenberg article bear on the question: Do minimalist races exist? That’s a question that has to be answered separately. In my view, the fundamental question in the philosophy of race on which the results of this study bear is whether minimalist race is biologically real. My contention is that they indicate that minimalist race (or more precisely, continental-level minimalist race) is biologically real if sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races.
Sub-Saharan Africans, Caucasians, East Asians, Amerindians, and Oceanians constitute minimalist races, therefore race is a biological reality. We can pinpoint them on the basis of patterns of visible physical features; these visible physical features correspond to geographic ancestry; this satisfies the criteria for minimalist races; therefore race exists. Race exists as a biological kind.
Furthermore, if these five populations that Rosenberg et al (2002) identified (the Blumenbachian populations) are minimalist races, then minimalist race is “a minor principle of human genetic structure” (Hardimon, 2017b: 92). Since minimalist races constitute a dimension within the small amount of human genetic variation that is captured between the continental-level minimalist races (4.3 percent), then it is completely possible to talk meaningfully about the racial structure of human genetic variation which consists of the human genetic variation which corresponds to continental-level minimalist races.
Thus, the US meaning of race is just a referent; the US meaning of race refers to a particular set of human populations; races in the US are classically-defined races (Amerindian, Caucasian, African, East Asian, and Oceanians; the Blumenbach partition); and race is both a biological reality as well as socially constructed. These populations are biologically real; if these populations are biologically real, then it stands to reason that biological racial realism is true (Hardimon, 2012 2013, 2017a; 2017b; Spencer, 2014, 2015).
Human races exist, in a minimalist biological sense, and there are 5 human races. Defenders of Rushton’s work—who believed there are only 3 primary races: Caucasoids, Mongoloids, and Negroids (while Amerindians and others were thrown into the “Mongoloid race” and Pacific Islanders being grouped with the “Negroid race” (Rushton, 1988, 1997; see also Liberman, 2001 for a critique of Rushton’s tri-racial views)—are forced into a tri-racial theory, since he used this tri-racial theory as the basis for his, now defunct, r/K selection theory. The tri-racial theory, that there are three primary races of man—Caucasoid, Mongoloid, and Negroid—has fallen out of favor with anthropologists for decades. But what we can see from new findings in population genetics since the sequencing of the human genome, however, is that human populations cluster into five populations and these five populations are races, therefore biological racial realism is true.
Biological racial realism (the fact that race exists as a biological reality) is true, however, just like with Hardimon’s minimalist races, they do not denote “superiority”, “inferiority” for one race over another. Most importantly, Blumenbachian populations do not denote those terms because the genetic evidence that is used to support the Blumenbachian partition use noncoding DNA. (It should also be noted that the terms “superior” and “inferior” are nonsensical, when used outside of their anatomic contexts. The head is the most superior part of the human body, the feet are the most inferior part of the human body. This is the only time these terms make sense, thus, using the terms outside of this context makes no sense.)
It is worth noting that, while Hardimon’s and Spencer’s views on race are similar, there are some differences between their views. Spencer sees “race” as a referent, while Hardimon argues that race has a set descriptive meaning on the basis of C (1)-(3); (C1) that, as a group, is distinguished from other groups of human beings by patterns of visible physical features, (C2) whose members are linked be a common ancestry peculiar to members of that group, and (C3) that originates from a distinctive geographic location” (Hardimon, 2017b: 31). Whether or not one prefers Blumenbachian partitions or minimalist races depends on whether or not one prefers race in a descriptive sense (i.e., Hardimon’s minimalist races) or if the term race in America is a referent to the US Census discourse, which means that “race” refers to the OMB definitions which then denote Blumenbachian partitions.
Hardimon also takes minimalist races to be a biological kind, while Spencer takes them to be a proper name for a set of population groups. Both of these differing viewpoints regarding race, while similar, are different in that one is describing a kind, while the other describes a proper name for a population group; these two views regarding population genetics from these two philosophers are similar, they are talking about the same things and hold the same deflationary views regarding race. They are talking about how race is seen in everyday life and where people get their definitions of “race” from and how they then integrate it into their everyday lives.
“Race” in America is a proper name for a set of human population groups, the five population groups identified by K = 5. Americans defer to the US Census Bureau on race, who defers to the Office of Management and Budget to define race. They hold that races are a “set”, and these “sets” are Oceanians, Caucasians, East Asians, Amerindians, and Africans. Race, thusly, refers to a set of population groups; “race” is not a “kind”, but a proper name for known populational groups. K = 5 then shows us that the demarcated clusters correspond to continental-level minimalist races, what is termed “the Blumenbach partition.” This partition is “race” in the US sense of the term, and it is a biological reality, therefore, like Hardimon’s minimalist races, the Blumenbach partition identifies what we in America know to be race. (It’s worth noting that, obviously, the Blumenbach partition/minimalist races are one in the same, Spencer is a deflationary realist regarding race, just like Hardimon.)
Black-White Differences in Anatomy and Physiology: Black Athletic Superiority
3000 words
Due to evolving in different climates, the different races of Man have differing anatomy and physiology. This, then, leads to differences in sports performance—certain races do better than others in certain bouts of athletic prowess, and this is due to, in large part, heritable biological/physical differences between blacks and whites. Some of these differences are differences in somatotype, which bring a considerable advantage for, say, runners (an ecto-meso, for instance, would do very well in sprinting or distance running depending on fiber typing). This article will discuss differences in racial anatomy and physiology (again) and how it leads to disparities in certain sports performance.
Kerr (2010) argues that racial superiority in sport is a myth. (Read my rebuttal here.) In his article, Kerr (2010) attempts to rebut Entine’s (2000) book Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It. In a nutshell, Kerr (2010) argues that race is not a valid category; that other, nongenetic factors play a role other than genetics (I don’t know if anyone has ever argued if it was just genetics). Race is a legitimate biological category, contrary to Kerr’s assertions. Kerr, in my view, strawman’s Entine (2002) by saying he’s a “genetic determinist”, but while he does discuss biological/genetic factors more than environmental ones, Entine is in no way a genetic determinist (at least that’s what I get from my reading of his book, other opinions may differ). Average physical differences between races are enough to delineate racial categories and then it’s only logical to infer that these average physical/physiological differences between the races (that will be reviewed below) would infer an advantage in certain sports over others, while the ultimate cause was the environment that said race’s ancestors evolved in (causing differences in somatotype and physiology).
Black athletic superiority has been discussed for decades. The reasons are numerous and of course, this has even been noticed by the general public. In 1991, half of the respondents of a poll on black vs. whites in sports “agreed with the idea that “blacks have more natural physical ability,“” (Hoberman, 1997: 207). Hoberman (1997) of course denies that there is any evidence that blacks have an advantage over whites in certain sports that come down to heritable biological factors (which he spends the whole book arguing). However, many blacks and whites do, in fact, believe in black athletic superiority and that physiologic and anatomic differences between the races do indeed cause racial differences in sporting performance (Wiggins, 1989). Though Wiggins (1989: 184) writes:
The anthropometric differences found between racial groups are usually nothing more than central tendencies and, in addition, do not take into account wide variations within these groups or the overlap among members of different races. This fact not only negates any reliable physiological comparisons of athletes along racial lines, but makes the whole notion of racially distinctive physiological abilities a moot point.
This is horribly wrong, as will be seen throughout this article.
The different races have, on average, differing somatotypes which means that they have different anatomic proportions (Malina, 1969):
| Data from Malina, (1969: 438) | n | Mesomorph | Ectomorph | Endomorph |
| Blacks | 65 | 5.14 | 2.99 | 2.92 |
| Whites | 199 | 4.29 | 2.89 | 3.86 |
| Data from Malina (1969: 438) | Blacks | Whites |
| Thin-build body type | 8.93 | 5.90 |
| Submedium fatty development | 48.31 | 29.39 |
| Medium fleshiness | 33.69 | 43.63 |
| Fat and very fat categories | 9.09 | 21.06 |
This was in blacks and whites aged 6 to 11. Even at these young ages, it is clear that there are considerable anatomic differences between blacks and whites which then lead to differences in sports performance, contra Wiggins (1989). A basic understanding of anatomy and how the human body works is needed in order to understand how and why blacks dominate certain sports over whites (and vice versa). Somatotype is, of course, predicated on lean mass, fat mass, bone density, stature, etc, which are heritable biological traits, thus, contrary to popular belief that somatotyping holds no explanatory power in sports today (see Hilliard, 2012).
One variable that makes up somatotype is fat-free body mass. There are, of course, racial differences in fat mass, too (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000). Lower fat mass would, of course, impede black excellence in swimming, and this is what we see (Rushton, 1997; Entine, 2000). Wagner and Heyward (2000) write:
Our review unequivocally shows that the FFB of blacks and whites differs significantly. It has been shown from cadaver and in vivo analyses that blacks have a greater BMC and BMD than do whites. These racial differences could substantially affect measures of body density and %BF. According to Lohman (63), a 2% change in the BMC of the body at a given body density could, theoretically, result in an 8% error in the estimation of %BF. Thus, the BMC and BMD of blacks must be considered when %BF is estimated.
While Vickery, Cureton, and Collins (1988) found that blacks had thinner skin folds than whites, however, in this sample, somatotype did not explain racial differences in bone density, like other studies (Malina, 1969), Vickery, Cureton, and Collins (1988) found that blacks were also more likely to be mesomorphic (which would then express itself in racial differences in sports).
Hallinan (1994) surveyed 32 sports science, exercise physiology, biomechanics, motor development, motor learning, and measurement evaluation textbooks to see what they said racial differences in sporting performance and how they explained them. Out of these 32 textbooks, according to Wikipedia, these “textbooks found that seven [textbooks] suggested that there are biophysical differences due to race that might explain differences in sports performance, one [textbook] expressed caution with the idea, and the other 24 [textbooks] did not mention the issue.” Furthermore, Strklaj and Solyali (2010), in their paper “Human Biological Variation in Anatomy Textbooks: The Role of Ancestry” write that their “results suggest that this type of human variation is either not accounted for or approached only superficially and in an outdated manner.”
It’s patently ridiculous that most textbooks on the anatomy and physiology of the human body do not talk about the anatomic and physiologic differences between racial and ethnic groups. Hoberman (1997) also argues the same, that there is no evidence to confirm the existence of black athletic superiority. Of course, many hypotheses have been proposed to explain how and why blacks are at an inherent advantage in sport. Hoberman (1997: 269) discusses one, writing (quoting world record Olympian in the 400-meter dash, Lee Evans):
“We were bred for it [athletic dominance] … Certainly the black people who survived in the slave ships must have contained the highest proportion of the strongest. Then, on the plantations, a strong black man was mated with a strong black woman. We were simply bred for physical qualities.”
While Hoberman (1997: 270-1) also notes:
Finally, by arguing for a cultural rather than a biological interpretation of “race,” Edwards proposed that black athletic superiority results from “a complex of societal conditions” that channels a disproporitionate number of talented blacks into athletic careers.
The fact that blacks were “bred for” athletic dominance is something that gets brought up often but has little (if any) empirical support (aside from just-so stories about white slavemasters breeding their best, biggest and strongest black slaves). The notion that “a complex of societal conditions” (Edwards, 1971: 39) explains black dominance in sports, while it has some explanatory power in regard to how well blacks do in sporting competition, it, of course, does not tell the whole story. Edwards (1978: 39) argues that these complex societal conditions “instill a heightened motivation among black male youths to achieve success in sports; thus, they channel a proportionately greater number of talented black people than whites into sports participation.” While this may, in fact, be true, this does nothing to rebut the point that differences in anatomic and physiologic factors are a driving force in racial differences in sporting performance. However, while these types of environmental/sociological arguments do show us why blacks are over-represented in some sports (because of course motivation to do well in the sport of choice does matter), they do not even discuss differences in anatomy or physiology which would also be affecting the relationship.
For example, one can have all of the athletic gifts in the world, one can be endowed with the best body type and physiology to do well in any type of sport you can imagine. However, if he does not have a strong mind, he will not succeed in the sport. Lippi, Favaloro, and Guidi (2008) write:
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.
Any athlete—no matter their race—needs a strong mental background, for if they don’t, they can have all of the physical gifts in the world, they will not become top-tier athletes in the sport of their choice; advantageous physical factors are imperative for success in differing sports, though myriad variables work in concert to produce the desired effect so you cannot have one without the other. On the other side, one can have a strong mental background and not have the requisite anatomy or physiology needed to succeed in the sport in question, but if he has a stronger mind than the individual with the requisite morphology, then he probably will win in a head-to-head competition. Either way, a strong mind is needed for strong performance in anything we do in life, and sport is no different.
Echoing what Hoberman (1997) writes, that “racist” thoughts of black superiority in part cause their success in sport, Sheldon, Jayaratne, and Petty (2007) predicted that white Americans’ beliefs in black athletic superiority would coincide with prejudice and negative stereotyping of black’s “intelligence” and work ethic. They studied 600 white men and women to ascertain their beliefs on black athletic superiority and the causes for it. Sheldon, Jayaratne, and Petty (2007: 45) discuss how it was believed by many, that there is a “ perceived inverse relationship between athleticism and intelligence (and hard work).” (JP Rushton was a big proponent of this hypothesis; see Rushton, 1997. It should also be noted that both Rushton, 1997 and Entine, 2000 believe that blacks’ higher rate of testosterone—3 to 15 percent— [Ross et al, 1986; Ellis and Nyborg, 1992; see rebuttal of both papers] causes their superior athletic performance, I have convincingly shown that they do not have higher levels of testosterone than other races, and if they do the difference is negligible.) However, in his book The Sports Gene: Inside the Science of Extraordinary Athletic Performance, Epstein (2014) writes:
With that stigma in mind [that there is an inverse relationship between “intelligence” and athletic performance], perhaps the most important writing Cooper did in Black Superman was his methodological evisceration of any supposed inverse link between physical and mental prowess. “The concept that physical superiority could somehow be a symptom of intellectual superiority became associated with African Americans … That association did not begin until about 1936.”
What Cooper (2004) implied is that there was no “inverse relationship” with intelligence and athletic ability until Jesse Owens blew away the competition at the 1936 Olympics in Berlin, Germany. In fact, the relationship between “intelligence” and athletic ability is positive (Heppe et al, 2016). Cooper is also a co-author of a paper Some Bio-Medical Mechanisms in Athletic Prowess with Morrison (Morrison and Cooper, 2006) where they argue—convincingly—that the “mutation appears to have triggered a series of physiological adjustments, which have had favourable athletic consequences.”
Thus, the hypothesis claims that differences in glucose conversion rates between West African blacks and her descendants began, but did not end with the sickling of the hemoglobin molecule, where valine is substituted for glutamic acid, which is the sixth amino acid of the beta chain of the hemoglobin molecule. Marlin et al (2007: 624) showed that male athletes who were inflicted with the sickle cell trait (SCT) “are able to perform sprints and brief exercises at the highest levels.” This is more evidence for Morrison and Cooper’s (2006) hypothesis on the evolution of muscle fiber typing in West African blacks.
Bejan, Jones, and Charles (2010) explain that the phenomenon of whites being faster swimmers in comparison to blacks being faster runners can be accounted for by physics. Since locomotion is a “falling-forward cycle“, body mass falls forward and then rises again, so mass that falls from a higher altitude falls faster and forward. The altitude is set by the position of center of mass above the ground for running, while for swimming it is set by the body rising out of the water. Blacks have a center of gravity that is about 3 percent higher than whites, which implies that blacks have a 1.5 percent speed advantage in running whereas whites have a 1.5 percent speed advantage in swimming. In the case of Asians, when all races were matched for height, Asians fared even better, than whites in swimming, but they do not set world records because they are not as tall as whites (Bejan, Jones, and Charles, 2010).
It has been proposed that stereotype threat is part of the reasons for East African running success (Baker and Horton, 2003). They state that many theories have been proposed to explain black African running success—from genetic theories to environmental determinism (the notion that physiologic adaptations to climate, too, drive differences in sporting competition). Baker and Horton (2003) note that “that young athletes have internalised these stereotypes and are choosing sport participation accordingly. He speculates that this is the reason why white running times in certain events have actually decreased over the past few years; whites are opting out of some sports based on perceived genetic inferiority.” While this may be true, this wouldn’t matter, as people gravitate toward what they are naturally good at—and what dictates that is their mind, anatomy, and physiology. They pretty much argue that stereotype threat is a cause of East African running performance on the basis of two assertions: (1) that East African runners are so good that it’s pointless to attempt to win if you are not East African and (2) since East Africans are so good, fewer people will try out and will continue the illusion that East Africans would dominate in middle- and long-distance running. However, while this view is plausible, there is little data to back the arguments.
To explain African running success, we must do it through a systems view—not one of reductionism (i.e., gene-finding). We need to see how the systems in question interact with every part. So while Jamaicans, Kenyans, and Ethiopians (and American blacks) do dominate in running competitions, attempting to “find genes” that account for success n these sports seems like a moot point—since the whole system is what matters, not what we can reduce the system in question to.
However, there are some competitions that blacks do not do so well in, and it is hardly discussed—if at all—by any author that I have read on this matter. Blacks are highly under-represented in strength sports and strongman competitions. Why? My explanation is simple: the causes for their superiority in sprinting and distance running (along with what makes them successful at baseball, football, and basketball) impedes them from doing well in strength and strongman competitions. It’s worth noting that no black man has ever won the World’s Strongest Man competition (indeed the only African country to even place—Rhodesia—was won by a white man) and the causes for these disparities come down to racial differences in anatomy and physiology.
I discussed racial differences in the big four lifts and how racial differences in anatomy and physiology would contribute to how well said race performed on the lift in question. I concluded that Europeans and Asians had more of an advantage over blacks in these lifts, and the reasons were due to inherent differences in anatomy and physiology. One major cause is also the differing muscle fiber typing distribution between the races (Alma et al, 1986; Tanner et al, 2002; Caesar and Henry, 2015 while blacks’ fiber typing helps them in short-distance sprinting (Zierath and Hawley, 2003). Muscle fiber typing is a huge cause of black athletic dominance (and non-dominance). Blacks are not stronger than whites, contrary to popular belief.
I also argued that Neanderthals were stronger than Homo sapiens, which then had implications for racial differences in strength (and sports). Neanderthals had a wider pelvis than our species since they evolved in colder climes (at the time) (Gruss and Schmidt, 2016). With a wider pelvis and shorter body than Homo sapiens, they were able to generate more power. I then implied that the current differences in strength and running we see between blacks and whites can be used for Neanderthals and Homo sapiens, thusly, evolution in differing climates lead to differences in somatotype, which eventually then lead to differences in sporting competition (what Baker and Horton, 2003 term “environmental determinism” which I will discuss in the context of racial differences in sports in the future).
Finally, blacks dominate the sport of bodybuilding, with Phil Heath dominating the competition for the past 7 years. Blacks dominate bodybuilding because, as noted above, blacks have thinner skin folds than whites, so their striations in their muscles would be more prevalent, on average, at the same exact %BF. Bodybuilders and weightlifters were similar in mesomorphy, but the bodybuilders showed more musculature than the bodybuilders whereas the weightlifters showed higher levels of body fat with a significant difference observed between bodybuilders and weightlifters in regard to endomorphy and ectomorphy (weightlifters skewing endo, bodybuilders skewing ecto, as I have argued in the past; Imran et al, 2011).
To conclude, blacks do dominate American sporting competition, and while much ink has been spilled arguing that cultural and social—not genetic or biologic—factors can explain black athletic superiority, they clearly work in concert with a strong mind to produce the athletic phenotype, no one factor has prominence over the other; though, above all, if one does not have the right mindset for the sport in question, they will not succeed. A complex array of factors is the cause of black athletic dominance, including muscle fibers, the type of mindset, anatomy, overall physiology and fat mass (among other variables) explain the hows and whys of black athletic superiority. Cultural and social explanations—on their own—do not tell the whole story, just as genetic/biologic explanations on their own would not either. Every aspect—including the historical—needs to be looked at when discussing the dominance (or lack thereof) in certain sports along with genetic and nongenetic factors to see how and why certain races and ethnies excel in certain sports.
Nina Jablonski on Race
1550 words
Nina Jablonski’s work on vitamin D and the implications that lighter skin had not only on our evolution but our health are extremely important for understanding how we evolved after the out of Africa migration. However, Jablonski then takes what she has written about skin color over the past few decades and concludes that race doesn’t exist. Jablonski believes that the term “race” should be discontinued from our lexicon, but as most may know, the term “race” does not need to disappear from our lexicon. (Watch her TED Talk Skin Color is an Illusion.)
In 2014, Nina Jablonski stated that the term “race” was ready for scientific retirement. In the article—and her book (Jablonski, 2012: chapters 9 and 10)—she states that race was a “vague and slippery concept”, eschewing the views of Kant and Hume as “racist”. She talks about how Kant was really one of the first people to recognize and categorize groups of people as “races”, stating that skin color, hair type, skull type etc—along with differing mores, aptitudes, and capacity for civilization—arranged in a hierarchical manner with Europeans at the top. A climatic theory was held, which stated that the original humans were light and became darker since “the transformation from light to dark was a form of degeneration, a departure from the norm” (Jablonski, 2012: 143).
She then discusses how, in Biblical history, skin color was meaningful, meaningful because it was believed that darker-skinned races were descendants of Ham:
And the sons of Noah, that went forth of the ark, were Shem, and Ham, and Japheth: and Ham is the father of Canaan. These are the three sons of Noah: and of them was the whole earth overspread. And Noah began to be an husbandman, and he planted a vineyard: And he drank of the wine, and was drunken; and he was uncovered within his tent. And Ham, the father of Canaan, saw the nakedness of his father, and told his two brethren without. And Shem and Japheth took a garment, and laid it upon both their shoulders, and went backward, and covered the nakedness of their father; and their faces were backward, and they saw not their father’s nakedness. And Noah awoke from his wine, and knew what his younger son had done unto him. And he said, Cursed be Canaan; a servant of servants shall he be unto his brethren. And he said, Blessed be the Lord God of Shem; and Canaan shall be his servant. (Genesis, 9: 18-26)
So Noah’s three sons—Ham, Japheth and Shem—were seen to be the three modern-day races of man—Africans, Europeans, and Asians, respectively. The term “servant of servants” was taken to mean that the descendants of Ham would serve the descendants of Shem and Japheth. This, according to those who believed the authority of the Bible, was enough to justify chattel slavery.
Jablonski—in an interview with the magazine Nautilus—stated that there “are no clean breaks between human populations. Individuals have different groups of genes” and that “Only a tiny fraction of alleles, and a small fraction of allelic combinations, is restricted to a single geographic region, and even less to a single population” which “is why attempts to identify races in humans have failed.” She commits the continuum fallacy, and the argument form is thus: “One extreme is X, at another is Y. There is no definable point where X becomes Y. Therefore, there is no difference between X and Y.” This has also been called the “Argument of the Beard”: at what point does a man not become clean shaven?
The use of the continuum fallacy, that there “are no clean breaks between human populations” shows how far the “race is a social construct” line has come (it is, but that race is a social construct does not also mean that it cannot also be a significant biological reality). The continuum fallacy is one of the most-used fallacies by those who deny race. Though, those who use the continuum fallacy are only attempting to argue that the claim is “too vague” because it is not as precise as they would like it to be. It does not matter that there “are no clean breaks between human populations“; what matters is that patterns of visible physical features correspond to geographic ancestry, and this is what we find.
Her second problem arises when she says that “Only a tiny fraction of alleles, and a small fraction od allelic combinations, is restricted to a single geographic region, and even less to s single population“. That there are no “race genes” or “genes for race” does not mean that race does not exist as a biological reality; these rigid “either this or that” definitions that some people have for race, such as race-specific genes are strawmen: people who believe that race is a significant biological reality do not believe in race-specific genes. That there are no race-specific genes does not mean that race doesn’t exist, as we know that genes are expressed differently in different races.
Finally, she claims that this “is why attempts to identify races in humans have failed“, though these attempts have not failed, of course. So-called races are distinguished by patterns of visible physical features; these patterns are observed between real, existing groups; these real existing groups that share these patterns of visible physical features satisfy the requisites of minimalist race; therefore race exists. Of course, Jablonski has reservations about acknowledging the reality of race due to how the transatlantic slave trade was promogulated through so-called differences that stemmed from Noah and Ham’s curse, but I fail to see why she would discard the argument just provided for the existence of race since differences in mores, intelligence, physical and mental abilities, are not discussed in the argument. ONLY the observable differences between populations are observed, with no value-judgment put onto each race, such as having lower “intelligence” or differing mores compared to another race.
She also states, in an interview with the New York Times, that skin color is not about race, “it’s about sun and how close our ancestors lived to the Equator. Skin color is what regulates our body’s reaction to the sun and its rays. … That shows that color is not a permanent trait.” That the differences in skin color observed in human populations can change over time does not mean that skin color “is not about race” as Jablonski claims. Skin color is one physical trait to delineate races, along with hair type, physiognomy, and anatomy, that groups peoples into groups we call “races”. This is not a good argument against the existence of race; of course anatomy, physiology, and physiognomy can change over time: but this does not mean that race does not exist!
Michael Hardimon’s race concepts (Hardimon, 2017) show that one does not need to believe that races differ in “intelligence”, mores, etc to believe in the existence of race. The concept takes everything from the racialist concept and “minimizes it”, taking the aspect of visible differences in physical features, while leaving the so-called mental differences (“intelligence”, mores) alone. This is enough to recognize that race exists and, as Jablonski has noted for decades in her career, being displaced from the environment where your skin color evolved causes an environmental mismatch which then—in the case of black Americans—may lead to vitamin D deficiency. This is one significant aspect that shows that race has an impact on health policy.
The minimalist concept of race is “deflationary” in that it does not discuss what we “can’t see” with our own eyes; it only discusses physical traits which should be enough for Jablonski to say that race is real and exists as a biological reality. Combined with the known health effects of, for instance, living in differing climates with differing amounts of UV radiation that is not “for” your skin color has further consequences and is why, in some cases, race-based medicine should stick around (though I am aware that, first and foremost, the individual matters first in a medical context, racial membership is secondary).
In sum, Jablonski refers to old and outdated individuals when speaking about the biological reality of race. She does a good job chronicling how and why the concept of race arose, especially through Biblical history and the curse of Ham. However, she takes it too far and claims that race does not exist, nor is it a significant biological reality since there are no “race-specific genes” (also remember that you do not need genes to delineate race, using differences in physical traits and then correlating them to geography is sufficient) and there “are no clean breaks between human populations“. These fallacies aside, it is possible, as I have noted before, to denote racial classifications sans the use of “intelligence” or “mores” in the concept. Skin color is just one of many observable traits that differ by geography that make the basis for separating groups on the basis of race.
The minimalist race concept from Hardimon is non-hierarchical: meaning that it doesn’t discuss anything that would put races in a hierarchy like the racialist concept does (with mores and “intelligence”). If anything, this strictly physical definition of races (and the simple argument for it) should be enough to sway race-deniers to become race-believers.
Cold Winter Theory, the Vitamin D Hypothesis and the Prediction of Novel Facts
2400 words
HBDers purport that as one moves further north from Africa that IQ raises as a function of how the population in question needed to survive. The explanation is that as our species migrated out of Africa, more “intelligence” was needed and this is what explains the current IQ disparities across the world: the ancestors of populations evolving in different areas with different demands then changed their “IQs” and this then is responsible for differential national development between nations. Cold winter theory (CWT) explains these disparities.
On the other hand is the vitamin D hypothesis (VDH). The VDH purports to explain why populations have light skin at northern latitudes. As the migration north out of Africa occurred, peoples needed to get progressively lighter in order to synthesize vitamin D. The observation here is that as light skin is selected for in locations where UVB is absent, seasonal or more variable whereas dark skin is selected for where UVB is stronger. So we have two hypotheses: but there is a problem. Only one of these hypotheses makes novel predictions. Predictions of novel predictions are what science truly is. A predicted fact is a novel fact for a hypothesis if it wasn’t used in the construction of the hypothesis (Musgrave, 1988). In this article, I will cover both the CWT and VDH, predictions of facts that each made (or didn’t make) and which can be called “science”.
Cold winter theory
The cold winter theory, formulated by Lynn and Rushton, purports to give an evolutionary explanation for differences in national IQs: certain populations evolved in areas with deathly cold winters in the north, while those who lived in tropical climes had, in comparison to those who evolved in the north, an “easier time to live”. Over time as populations adapted to their environments, differences in ‘intelligence’ (whatever that is) evolved due to the different demands of each environment, or so the HBDers say.
Put simply, the CWT states that IQ differences exist due to different evolutionary pressures. Since our species migrated into cold, novel environments, this was the selective pressure needed for higher levels of ‘intelligence’. On the other hand, humans who remained in Africa and other tropical locations did experience these novel, cold environments and so their ‘intelligence’ stayed at around the same level as it was 70,000 years ago. Many authors hold this theory, including Rushton (1997), Lynn (2006), Hart, (2007) Kanazawa (2008), Rushton and Templer (2012; see my thoughts on their hypothesis here) and Wade (2014). Lynn (2013) even spoke of a “widespreadonsensus” on the CWT, writing:
“There is widespread consensus on this thesis, e.g. Kanazawa (2008), Lynn (1991, 2006), and Templer and Arikawa (2006).”
So this “consensus” seems to be a group of his friends and his own publications. We can change this sentence to ““There is widespread consensus on this thesis, including two of my publications, a paper where the author assumes that the earth is flat: “First, Kanazawa’s (2008) computations of geographic distance used Pythagoras’ theorem and so the paper assumed that the earth is flat (Gelade, 2008).” (Wicherts et al, 2012) and another publication where the authors assume hot weather leads to lower intelligence. Oh yea, they’re all PF members. Weird.” That Lynn (2013) calls this “consensus” is a joke.
What caused higher levels of ‘intelligence’ in those that migrated out of Africa? Well, according to those who push the CWT, finding food and shelter. Kanazawa, Lynn, and Rushton all argue that finding food, making shelter and hunting animals were all harder in Eurasia than in Africa.
One explanation for high IQs of people who evolved recently in northern climes is their brain size. Lynn (2006: 139) cites data showing the average brain sizes of populations, along with the temperatures in that location:
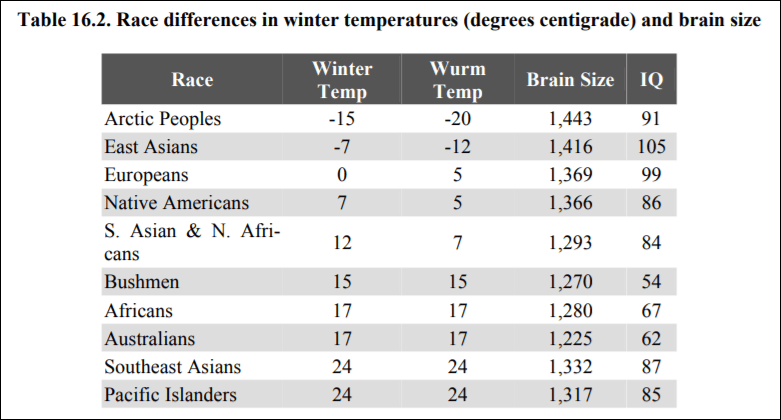
Do note the anomaly with the Arctic peoples. To explain this away in an ad-hoc manner, Lynn (2006: 156-7) writes:
These severe winters would be expected to have acted as a strong selection for increased intelligence, but this evidently failed to occur because their IQ is only 91. The explanation for this must lie in the small numbers of the Arctic Peoples whose population at the end of the twentieth century was only approximately 56,000 as compared with approximately 1.4 billion East Asians.
This is completely ad-hoc. There is no independent verifier for the claim. That the Arcitic don’t have the highest IQs but experienced the harshest temperatures and therefore have the biggest brain size is a huge anomaly, which Lynn (2006) attempts to explain away by population size.
He does not explain why natural selection among Arctic peoples would result in larger brain sizes or enhanced visual memory yet the same evolutionary pressures associated with a cold environment would not also produce higher intelligence. Arctic peoples have clear physical adaptations to the cold, such as short, stocky bodies well-suited to conserving heat.
Furthermore, the argument that Lynn attempts is on the mutations/population size is special pleading—he is ignoring anomalies in his theory that don’t fit it. However, “evolution is not necessary for temperature and IQ to co-vary across geographic space” (Pesta and Poznanski, 2014).
If high ‘intelligence’ is supposedly an adaptation to cold temperatures, then what is the observation that disconfirms a byproduct hypothesis? On the other hand, if ‘intelligence’ is a byproduct, which observation would disconfirm an adaptationist hypothesis? No possible observation can confirm or disconfirm either hypothesis, therefore they are just-so stories. Since a byproduct explanation would explain the same phenomena since byproducts are also inherited, then just saying that ‘intelligence’ is a byproduct of, say, needing larger heads to dissipate heat (Lieberman, 2015). One can make any story they want to fit the data, but if there is no prediction of novel facts then how useful is the hypothesis if it explains the data it purports to explain and only the data it purports to explain?
It is indeed possible to argue that hotter climates need higher levels of intelligence than colder climates, which has been argued in the past (see Anderson, 1991; Graves, 2002; Sternberg, Grigorenko, and Kidd, 2005). Indeed, Sternberg, Grigorenko, and Kidd (2005: 50) write: “post hoc evolutionary arguments … can have the character of ad hoc “just so” stories designed to support, in retrospect, whatever point the author wishes to make about present-day people.” One can think up any “just-so” story to explain any data. But if the “just-so” story doesn’t make any risky predictions of novel facts, then it’s not science, but pseudoscience.
Vitamin D hypothesis
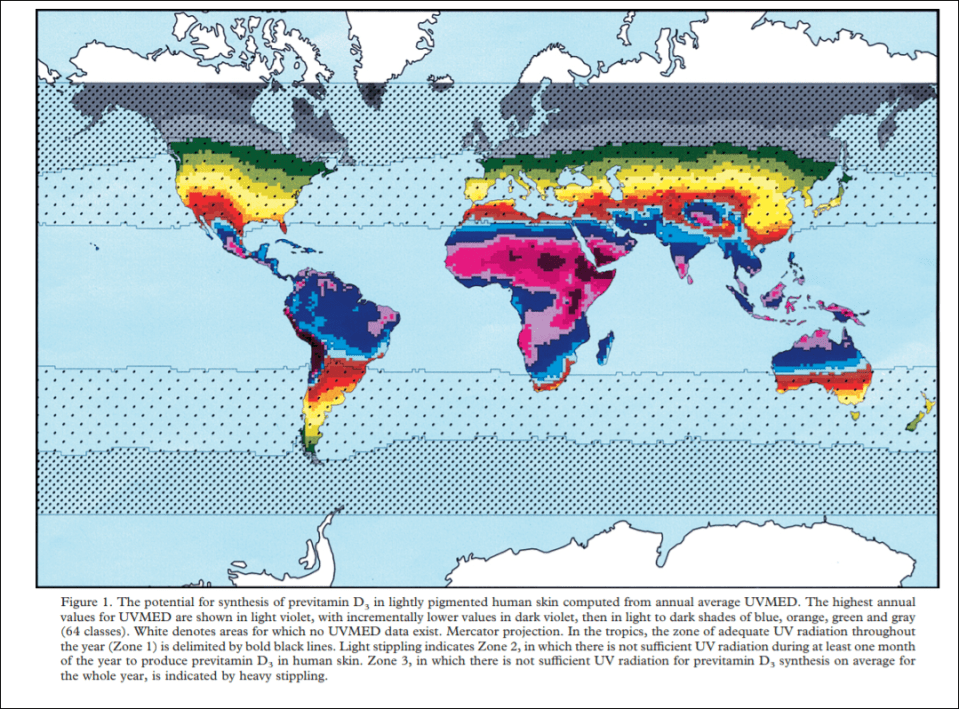
The VDH is simple: those populations that evolved in areas with seasonal, absent, or more variable levels of UVB have lighter skin than populations that evolved in areas with strong UVB levels year-round (Chaplan and Jablonksi, 2009: 458). Robins (2009) is a huge critic of the VDH, though her objections to the VDH have been answered (and will be discussed below).
The VDH is similar to the CWT in that it postulates that the adaptations in question only arose due to migrations out of our ancestral lands. We can see a very strong relationship between high UVB rays and dark skin and conversely with low UVB rays and light skin. Like with the CWT, the VDH has an anomaly and, coincidentally, the anomaly has to do with the same population involved in the CWT anomaly.
Arctic people have dark-ish skin for living in the climate that they do. But since they live in very cold climates then we have a strange anomaly here that needs explaining. We only need to look at the environment around them. They are surrounded by ice. Ice reflects UVB rays. UVB rays hit the skin. Arctic people consume a diet high in vitamin D (from fish). Therefore what explains Arctic skin color is UVB rays bouncing off the ice along with their high vitamin D diet. The sun’s rays are, actually, more dangerous in the snow than on the beach, with UVB rays being 2.5 more times dangerous in the snow than beach.
Evolution in different geographic locations over tens of thousands of years caused skin color differences. Thus, we can expect that, if peoples are out of the conditions where their ancestors evolved their skin color, that there would then be expected complications. For example, if human skin pigmentation is an adaptation to UV rays (Jablonski and Chaplan, 2010), we should expect that, when populations are removed from their ancestral lands and are in new locations with differing levels of UV rays, that there would be a subsequent uptick in diseases caused by vitamin D deficiencies.
This is what we find. We find significant differences in circulating serum vitamin D levels, and these circulating serum vitamin D levels then predict health outcomes in certain populations. This would only be true if sunlight influenced vitamin D production and that skin progressively gets lighter as one moves away from Africa and other tropical locations.
Skin pigmentation regulates vitamin D production (Neer, 1975). This is due to the fact that when UVB rays strike the skin, we synthesize vitamin D, and the lighter one’s skin is, the more vitamin D can be synthesized in areas with fewer UVB rays. (Also see Daraghmeh et al, 2016 for more evidence for the vitamin D hypothesis.)
P1) UV rays generate vitamin D in human skin
P2) Human populations that migrate to climates with less sunlight get fewer UV rays
P3) To produce more vitamin D, the skin needs to get progressively lighter
C) Therefore, what explains human skin variation is climate and UV rays linked to vitamin D production in the skin.
Novel predictions
Science is the generation of novel facts from risky predictions (Musgrave, 1988; Winther, 2009). And so, hypotheses that predict novel facts from risky predictions are scientific hypotheses, whereas those hypotheses that need to continuously backtrack and think up ad-hoc hypotheses are then pseudoscientific. Pseudoscience is simple enough to define. The Stanford Encyclopedia of Philosophy defines it as:
“A pretended or spurious science; a collection of related beliefs about the world mistakenly regarded as being based on scientific method or as having the status that scientific truths now have.”
All theories have a protective belt of ad hoc hypotheses. Theories become pseudoscientific when they fail to make new predictions and must take on more and more ad-hoc hypotheses that have no predictive value. If the ad-hoc hypotheses that are added to the main hypothesis have no predictive value then the new explanations for whichever hypothesis that is in danger of being falsified are just used to save the hypothesis from being refuted and it thus becomes pseudoscience.
In the case of CWT, it makes no prediction of novel facts; it only explains the data that it purports to explain. What is so great about the CWT if it makes no predictions of novel facts and only explains what it purports to explain? One may attempt to argue that it has made some ‘novel’ predictions but the ‘predictions’ that are proposed are not risky at all.
For example, Hart (2007: 417) makes a few “predictions”, but whether or not they’re “risky” or “novel” I’ll let you decide (I think they’re neither, of course). He writes that very few accomplishments will be made by Africans, or Australian or New Guinean Aborigines; members of those groups will not be highly represented in chess; and that major advances in scientific fields will come from those of European ancestry or the “Monglids”, Koreans, Chinese or Japanese.
On the other hand, Hart (2007: 417) makes two more “predictions”: he says that IQ data for Congoid Pygmies, Andaman Islanders, and Bantu-speaking people are few and far between and he believes that when enough IQ testing is undertaken there he expects IQ values between 60 and 85. Conversely, for the Lapps, Siberians, Eskimoes, Mongols and Tibetans, he predicts that IQ values should be between 85-105. He then states that if these “predictions” turn out to be wrong then he would have to admit that his hypothesis is wrong. But the thing is, he chose “predictions” that he knew would come to pass and therefore these are not novel, risky predictions but are predictions that Hart (2007) knows would come to pass.
What novel predictions has the VDH made? This is very simple. The convergent evolution of light skin was predicted in all hominids that trekked out of Africa and into colder lands. This occurred “because of the importance of maintaining the potential for producing pre-vitamin D3 in the skin under conditions of low annual UVB (Jablonski and Chaplin, 2000; Jablonski, 2004)” while these predictions “have been borne out by recent genetic studies, which have demonstrated that depigmented skin evolved independently by different molecular mechanisms multiple times in the history of the human lineage” (Chaplan and Jablonksi, 2009: 452). This was successfully predicted by Chaplan and Jablonski (2000).
The VDH still holds explanatory scope and predictive success; no other agent other than vitamin D can explain the observation that light skin is selected for in areas where there is low, absent or seasonal UVB. Conversely, in areas where there is a strong, year-round presence of UVB rays, dark skin is selected for.
Conclusion
Scientific hypotheses predict novel facts not known before the formulation of the hypothesis. The VDT has successfully predicted novel facts, whereas I am at a loss thinking of a novel fact that the CWT predicted.
In order to push an adaptationist hypothesis for CWT and ‘intelligence’, one must propose an observation that would confirm the adaptationist hypothesis while at the same time disconfirming the byproduct hypothesis. Since byproducts are inherited to, the byproduct hypothesis would predict the same things that an adaptationist hypothesis would. Thus, the CWT is a just-so story since no observation would confirm or disconfirm either hypothesis. On the other hand, the CWT doesn’t make predictions of novel facts, it makes “predictions” that are already known and would not undermine the hypothesis if disproved (but there would always be a proponent of the CWT waiting in the wings to propose an ad-hoc hypothesis in order to save the CWT, but I have already established that it isn’t science).
On the other hand, the VDT has successfully predicted that hominins that trekked out of Africa would have light skin which was then subsequently confirmed by genomic evidence. The fact that strong UVB rays year-round predict dark skin whereas seasonal, absent, or low levels of UVB predict light skin has been proved to be true. With the advent of genomic testing, it has been shown that hominids that migrated out of Africa did indeed have lighter skin. This is independent verification for the VDH; the VDH has predicted a novel fact whereas the CWT has not.
Race and Vitamin D Deficiency
1600 words
Vitamin D is an important “vitamin” (it is really a steroid hormone). It is produced when the skin (the largest organ in the body) is exposed to the sun’s UVB rays (Nair and Maseeh, 2012). So this is one of the only ways to get natural levels of UVB. We can then think that, if a population is outside of its natural evolutionary habitat (the habitat where that skin color evolved), then we should note numerous problems caused by the lack of vitamin D in whichever population is studied outside of a location that doesn’t get the correct amount of UVB rays from the sun.
Black Americans are more likely than other ethnies to be deficient in vitamin D (Harris, 2006; Cosman et al, 2007; Nair, 2012; Forest and Stuhldreher, 2014; Taksler et al, 2014). But, paradoxically, low vitamin D levels don’t cause weaker bones in black Americans (O’Conner et al, 2014). However, like with all hypotheses, there are naysayers. For example. Powe et al (2013) argue that vitamin D tests misdiagnose blacks, that blacks have a form of the vitamin that cells can use called 25-hydroxyvitamin D. They conclude: “Community-dwelling black Americans, as compared with whites, had low levels of total 25-hydroxyvitamin D and vitamin D–binding protein, resulting in similar concentrations of estimated bioavailable 25-hydroxyvitamin D. Racial differences in the prevalence of common genetic polymorphisms provide a likely explanation for this observation.” Though there are a whole host of problems here.
The limitations of Powe et al (2013) striking: it was cross-sectional and observational (like most nutrition studies) so they were unable to predict effects of vitamin-D binding protein on bone fractures; no data on the consumption of vitamin D supplements; measurement of bone turnover markers, urinary calcium excretion and levels of 1,25-dihydroxyvitamin D may explain the effect of VDBP (vitamin D-binding protein) on mineral metabolism; and they relied on a calculation, rather than a measurement of 25-hydroxyvitamin D levels.
Powe et al’s (2013) findings, though, have been disputed. Using different measurement tools from Powe et al (2013), Henderson et al (2015) conclude that “Counter to prior observations by immunoassay, VDBG concentrations did not vary by race.” While Bouillon (2014) writes: In our view, black Americans, as compared with white Americans, have lower levels of not only total 25-hydroxyvitamin D but also free or bioavailable 25-hydroxyvitamin D.” And finally, Hollis and Bikle (2014) write: “Specifically, for any given physically measured level of bio-available 25-hydroxyvitamin D, the authors are overestimating bio-available 25-hydroxyvitamin D by 2 to 2.5 times owing to underestimation of vitamin D–binding protein in blacks.”
Either way, even if what Powe et al (2013) conclude is true, that would not mean that black Americans should not supplement with vitamin D, since many diseases and health problems are associated with low vitamin D intake in blacks, including osteoporosis, cardiovascular disease, cancer, diabetes, and other serious conditions (Harris, 2006). An indirect relationship between low levels of vitamin D and hypertension is also noted (Mehta and Agarwal, 2017). Since there is an indirect relationship between vitamin D levels and hypertension, then we should keep an eye on this because black Americans have some of the highest levels of hypertension in the world (Ferdinand and Armani, 2007; see also Fuchs, 2011).
Vitamin D is, of course, important for skeletal and nonskeletal health (Kennel et al, 2010). So if vitamin D is important for skeletal and nonskeletal health, we should see more diseases in black Americans that imply a lack of this steroid in the body. Although blacks have stronger bones even when deficient in vitamin D, it is still observed that black children who break their forearms have less vitamin D circulating in their blood (Ryan et al, 2011). This observation is borne out by the data, since black children are more likely to be deficient in vitamin D compared to other ethnies (Moore, Murphy, and Hollick, 2005). Since black skin predicts vitamin D deficiency (Thomas and Demay, 2000), it seems logical to give vitamin D supplements to children, especially black children, on the basis that it would help lower incidences of bone fractures, even though blacks have stronger bones than whites.
Furthermore, physiologically “normal” levels of vitamin D differ in blacks compared to whites (Wright et al, 2012). They showed that it is indeed a strong possibility that both whites and blacks have different levels of optimum vitamin D. Wright et al (2012) showed that there is a relationship between 25(OH)D levels and intact parathyroid hormone (iPth); for blacks, the threshold in which there was no change was 20 ng/ml whereas for whites it was 30 ng/ml which suggests that there are different levels of optimal vitamin D for each race, and the cause is due to skin color. Thus, physiologically “normal” levels of vitamin D differ for blacks and whites.
There is also a high prevalence of vitamin D deficiency/insufficiency and asthma in black inner-city youth in Washington DC (Freishtat et al, 2010). We can clearly see that, even though black Americans have stronger bones than white Americans and vitamin D predicts bone strength, the fact that blacks have stronger bones than whites even while being deficient in vitamin D on average does not mean that black Americans should not supplement with vitamin D, since it would ameliorate many other problems they have that are related to vitamin D deficiency.
There are also racial differences in prostate cancer (PCa) acquisition too, and vitamin D deficiency may also explain this disparity (Khan and Partin, 2004; Bhardwaj et al, 2017). I have heavily criticized the explanations that testosterone influences PCa, while having indicated that environmental factors such as diet and vitamin D deficiency may explain a large amount of the gap (Batai et al, 2017; but see Stranaland et al, 2017 for a contrary view). Since low vitamin D is related to prostate cancer, by supplementing with vitamin D, it is possible that levels of PCa may decrease. Kristal et al (2014) show that both high and low levels of vitamin D are associated with PCa.
Evidence also exists that vitamin D levels and hypertension are related. Rostand (2010) proposes a unified hypothesis: an important role exists in vitamin D deficiency and the pathogenesis and maintenance of hypertension in blacks (Rostand, 2010).
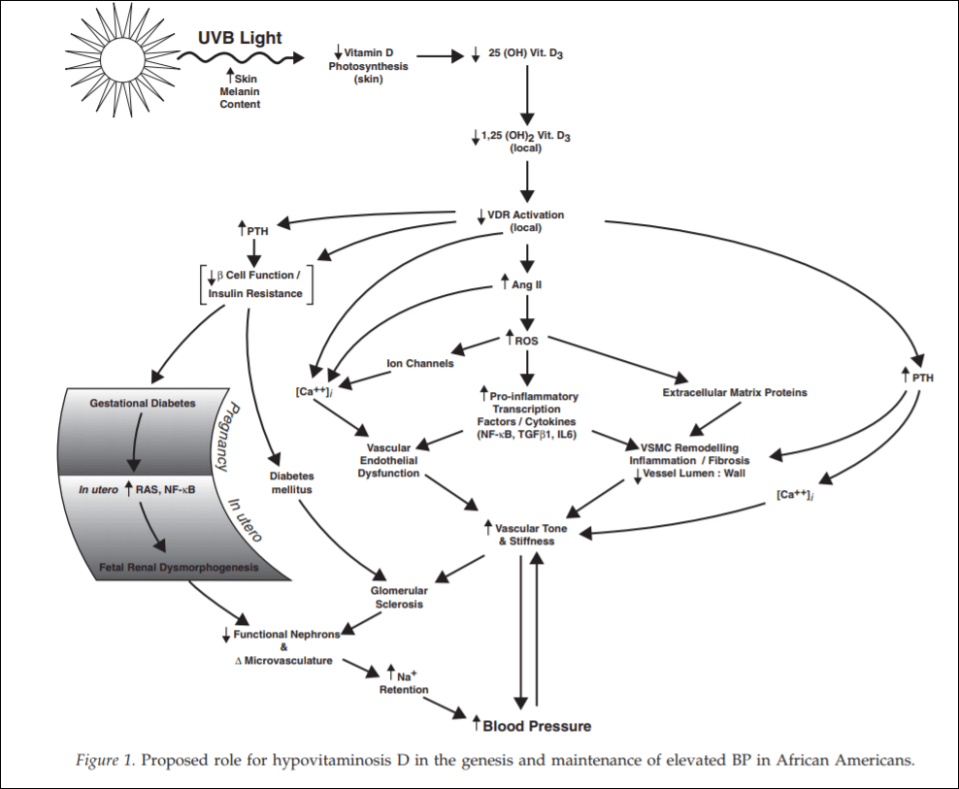
(From Rostand, 2010)
Since black Americans are no longer near the equator, their ability to synthesize vitamin D from UVB rays is diminished. This then probably leads the RAS (renin-angiotensin system) and inflammatory cytokine activation which then leads to vascular endothelial dysfunction along with structural changes to the microvasculature, which have been linked to vascular (arterial) stiffness along with increased vascular resistance, and these changes are shown to precede hypertension, which also occurs early in life. So since blacks are deficient in vitamin D, which even starts in the womb (Bodnar et al, 2007; Dawodu and Wagner, 2007; Lee et al, 2007; Khalessi et al, 2015; Seto et al, 2016), and this vitamin D deficiency most likely produces changes in large and small arteries and arterials, this could be the explanation for higher hypertension in black Americans (Rostand, 2010: 1701).
This would be a large environmental mismatch: since the population is displaced from its ancestral homeland, then this causes problems since it is not the environment where their ancestors evolved. So in this case, since black Americans are concentrated in the southeast corner of the United States, this may explain the high rates of vitamin D deficiency and hypertension in the black American community.
People whose ancestors evolved in locations with fewer UVB rays have lighter skin, whereas people whose ancestors evolved in locations with more UVB rays have darker skin. Thus, by placing populations in their opposite evolutionary environment, we can see how and why deleterious effects would occur in the population that is in the mismatched environment. For whites, skin cancer would occur, whereas for blacks, higher rates of hypertension and low birth weights occur.
Looking at levels of vitamin D deficiency in races is a great way to understand the evolution of certain populations. Because if the vitamin D hypothesis is correct, if skin color is an adaptation to UVB rays, with light skin being an adaptation to low UVB while dark skin is an adaptation to high UVB, then we can safely hypothesize about certain problems that would arise in races that are outside of their natural habitats. We have confirmed these hypotheses—black Americans who are outside of the location that their ancestors evolved in are more likely to have deleterious symptoms, and the symptoms are due to differences in vitamin D production, which come down to differences in skin color and how the skin synthesizes vitamin D in low-light environments.
Even though blacks have stronger bones than whites, this does not mean that they do not experience fractures at a high rate—especially children—and since the association was noticed, then by supplementing with vitamin D, this may lower the disparity of these types of injuries.
Since black Americans, compared to their evolutionary history, live in low-light environments, this then explains the how and why of vitamin D deficiency and why blacks need to supplement with vitamin D; no matter if certain studies show that blacks are ‘healthy’ even though they have low levels of vitamin D. If true (which I strongly doubt), that does not mean that black Americans should not supplement with vitamin D, because numerous other maladies are associated with vitamin D intake. This is one aspect where understanding the evolution of our species and the different races in it would lead to better medical care for individuals and ancestral groups that may need special treatment.
It is clear that race and geography should inform vitamin D intake, for if we do this, many diseases that arise can be ameliorated and quality of life can increase for everyone.
Race Differences in Penis Size Revisited: Is Rushton’s r/K Theory of Race Differences in Penis Length Confirmed?
2050 words
In 1985 JP Rushton, psychology professor at the University of Ontario, published a paper arguing that r/K selection theory (which he termed Differential K theory) explained and predicted outcomes of what he termed the three main races of humanity—Mongoloids, Negroids and Caucasoids (Rushton, 1985; 1997). Since Rushton’s three races differed on a whole suite of traits, he reasoned races that were more K-selected (Caucasoids and Mongoloids) had slower reproduction times, higher time preference, higher IQ etc in comparison to the more r-selected Negroids who had faster reproduction times, lower time preference, lower IQ etc (see Rushton, 1997 for a review; also see Van Lange, Rinderu, and Bushmen, 2017 for a replication of Rushton’s data not theory). Were Rushton’s assertions on race and penis size verified and do they lend credence to his Differential-K claims regarding human races?
Rushton’s so-called r/K continuum has a whole suite of traits on it. Ranging from brain size to speed of maturation to reaction time and IQ, these data points supposedly lend credence to Rushton’s Differential-K theory of human differences. Penis size is, of course, important for Rushton’s theory due to what he’s said about it in interviews.
Rushton’s main reasoning for penis size differences between race is “You can’t have both”, and that if you have a larger brain then you must have a smaller penis; if you have a smaller penis you must have a larger brain. He believed there was a “tradeoff” between brain size and penis size. In the book Darwin’s Athletes: How Sport Has Damaged Black America and Preserved the Myth of Race, Hoberman (1997: 312) quotes Rushton: “Even if you take something like athletic ability or sexuality—not to reinforce stereotypes or some such thing—but, you know, it’s a trade-off: more brain or more penis. You can’t have both.” This, though, is false. There is no type of evidence to imply that this so-called ‘trade-off’ exists. In my readings of Rushton’s work over the years, that’s always something I’ve wondered: was Rushton implying that large penises take more energy to have and therefore the trade-off exists due to this supposed relationship?
Andrew Joyce of the Occidental Observer published an article the other day in defense of Richard Lynn. Near the end of his article he writes:
Another tactic is to belittle an entire area of research by picking out a particularly counter-intuitive example that the public can be depended on to regard as ridiculous. A good example is J. Philippe Rushton’s claim, based on data he compiled for his classic Race, Evolution and Behavior, that average penis size varied between races in accord with the predictions of r/K theory. This claim was held up to ridicule by the likes of Richard Lewontin and other crusaders against race realism, and it is regularly presented in articles hostile to the race realist perspective. Richard Lynn’s response, as always, was to gather more data—from 113 populations. And unsurprisingly for those who keep up with this area of research, he found that indeed the data confirmedRushton’s original claim.
The claim was ridiculed because it was ridiculous. This paper by Lynn (2013) titled Rushton’s r-K life history theory of race differences in penis length and circumference examined in 113 populations is the paper that supposedly verifies Rushton’s theory regarding race differences in penis size, along with one of its correlates in Rushton’s theory (testosterone). Lynn (2013) proclaims that East Asians are the most K-evolved, then come Europeans, while Africans are the least K-evolved. This, then, is the cause of the supposed racial differences in penis size.
Lynn (2013) begins by briefly discussing Rushton’s ‘findings’ on racial differences in penis size while also giving an overview of Rushton’s debunked r/K selection theory. He then discusses some of Rushton’s studies (which I will describe briefly below) along with stories from antiquity of the supposed larger penis size of African males.
Our old friend testosterone also makes an appearance in this paper. Lynn (2013: 262) writes:
Testosterone is a determinant of aggression (Book, Starzyk, & Quinsey, 2001; Brooks & Reddon, 1996; Dabbs, 2000). Hence, a reduction of aggression and sexual competitiveness between men in the colder climates would have been achieved by a reduction of testosterone, entailing the race differences in testosterone (Negroids > Caucasoids > Mongoloids) that are given in Lynn (1990). The reduction of testosterone had the effect of reducing penis length, for which evidence is given by Widodsky and Greene (1940).
Phew, there’s a lot to unpack here. (I discuss Lynn 1990 in this article.) Testosterone does not determine aggression; see my most recent article on testosterone (aggression increases testosterone; testosterone does not increase aggression. Book, Starzyk and Quinsey, 2001 show a .14 correlation between testosterone and aggression, whereas Archer, Graham-Kevan, and Davies 2005 show the correlation is .08). This is just a correlation. Sapolsky (1997: 113) writes:
Okay, suppose you note a correlation between levels of aggression and levels of testosterone among these normal males. This could be because (a) testosterone elevates aggression; (b) aggression elevates testosterone secretion; (c) neither causes the other. There’s a huge bias to assume option a while b is the answer. Study after study has shown that when you examine testosterone when males are first placed together in the social group, testosterone levels predict nothing about who is going to be aggressive. The subsequent behavioral differences drive the hormonal changes, not the other way around.
Brooks and Reddon (1996) also only show relationships with testosterone and aggressive acts; they show no causation. This same relationship was noted by Dabbs (2000; another Lynn 2013 citation) in prisoners. More violent prisoners were seen to have higher testosterone, but there is a caveat here too: being aggressive stimulates testosterone production so of course they had higher levels of testosterone; this is not evidence for testosterone causing aggression.
Another problem with that paragraph quoted from Lynn (2013) is that it’s a just-so story. It’s an ad-hoc explanation. You notice something with data you have today and then you imagine a nice-sounding story to attempt to explain your data in an evolutionary context. Nice-sounding stories are cool and all and I’m sure everyone loves a nicely told story, but when it comes to evolutionary theory I’d like theories that can be independently verified of the data they’re trying to explain.
My last problem with that paragraph from Lynn (2013) is his final citation: he cites it as evidence that the reduction of testosterone affects penis length…..but his citation (Widodsky and Green, 1940) is a study on rats… While these studies can give us a wealth of information regarding our physiologic systems (at least showing us which types of avenues to pursue; see my previous article on myostatin), they don’t really mean anything for humans; especially this study on the application of testosterone to the penis of a rat. See, the fatal flaw in these assertions is this: would a, say, 5 percent difference in testosterone lead to a larger penis as if there is a dose-response relationship between testosterone and penis length? It doesn’t make any sense.
Lynn (2013), though, says that Rushton’s theory doesn’t propose that there is a direct causal relationship between “intelligence”‘ and penis length, but just that they co-evolved together, with testosterone reduction occurring when Homo sapiens migrated north out of Africa they needed to cooperate more so selection for lower levels of testosterone subsequently occurred which then shrunk the penises of Rushton’s Caucasian and Mongoloid races.
Lynn (2013) then discusses two “new datasets”, one of which is apparently in Donald Templer’s book Is Size Important (which is on my to-read list, so many books, so little time). Table 1 below is from Lynn reproducing Templer’s ‘work’ in his book.
The second “dataset” is extremely dubious. Lynn (2013) attempts to dress it up, writing that “The information in this website has been collated from data obtained by research centres and reports worldwide.” Ethnicmuse has a good article on the pitfalls of Lynn’s (2013) article. (Also read Scott McGreal’s rebuttal.)
Rushton attempted to link race and penis size for 30 years. In a paper with Bogaert (Rushton and Bogaert, 1987), they attempt to show that blacks had larger penises than whites who h ad longer penises than Asians which then supposedly verified one dimension of Rushton’s theory. Rushton (1988) also discusses race differences in penis size, citing a previous paper by Rushton and Bogaert, where they use data from Alfred Kinsey, but this data is nonrepresentative and nonrandom (see Zuckermann and Brody, 1988 and Weizmann et al, 1990: 8).
Still others may attempt to use supposed differences in IGF-1 (insulin-like growth factor 1) as evidence that there is, at least, physiological evidence for the claim that black men have larger penises than white men, though I discussed that back in December of 2016 and found it strongly lacking.
Rushton (1997: 182) shows a table of racial differences in penis size which was supposedly collected by the WHO (World Health Organization). Though a closer look shows this is not true. Ethnicmuse writes:
ANALYSIS: The WHO did not study penis sizes. It relied on three separate studies, two of which were not peer-reviewed and the data was included as “Appendix III” (which should have alerted Rushton that this was not an original study). The first study references Africans in the US (not Africa!) and Europeans in the US (not Europe!), the second Europeans in Australia (not Europe!) and the third, Thais.
So it seems to be bullshit all the way down.
Ajmani et al (1985) showed that 385 healthy Nigerians had an average penile length of 3.21 inches (flaccid). Orakwe and Ebuh (2007) show that while Nigerians had longer penises than other ethnies tested, the only statistical difference was between them and Koreans. Though Veale et al (2014: 983) write that “There are no indications of differences in racial variability in our present study, e.g. the study from Nigeria was not a positive outlier.”
Lynn and Dutton have attempted to use androgen differentials between the races as evidence for racial differences in penis size (this is another attempt at a physiological argument to attempt to show the existence of racial differences in penis size). Edward Dutton attempted to revive the debate on racial differences in penis size during a 2015 presentation where he, again, showed that Negroids have higher levels of testosterone than Caucasoids who have higher levels of androgens than Mongoloids. These claims, though, have been rebutted by Scott McGreal who showed that populations differences in androgen levels are meaningless while they subsequently fail to validate Rushton and Lynn’s claims on racial differences in penis size.
Finally, it was reported the other day that condoms from China were too small in Zimbabwe, per Zimbabwe’s health minister. This led Kevin MacDonald to proclaim that this was “More corroboration of race differences in penis size which was part of the data Philippe Rushton used in his theory of r/K selection (along with brain size, maturation rates, IQ, etc.)” This isn’t “more corroboration” for Rushton’s long-dead theory; nor is this evidence that blacks have longer penises. I don’t understand why people make broad and sweeping generalizations. It’s one country in Africa that complained about smaller condoms from a country in East Asia, therefore this is more corroboration for Rushton’s r/K selection theory? The logic doesn’t follow.
Asians have small condoms. Those condoms go to Africa. They complain condoms from China are too small. Therefore Rushton’s r/K selection theory is corroborated. Flawed logic.
In sum, Lynn (2013) didn’t verify Rushton’s theory regarding racial differences in penis size and I find it even funnier that Lynn ends his article talking about “falsification’ stating that this aspect of Rushton’s theory has survived two attempts at falsification, therefore, it can be regarded as a “progressive research program“, though obviously, with the highly flawed “data” that was used, one cannot rationally make that statement. Supposed hormonal differences between the races do not cause penis size differences; even if blacks had levels of testosterone significantly higher than whites (the 19 percent that is claimed by Lynn and Rushton off of one highly flawed study in Ross et al, 1986) they still would not have longer penises.
The study of physical differences between populations is important, but sometimes, stereotypes do not tell you anything, especially in this case. Though in this instance, the claim that blacks have the longest penis lies on shaky ground, and with what evidence we do have for the claim, we cannot logically make the inference (especially not from Lynn’s (2013) flimsy data). Richard Lynn did not “confirm” anything with this paper; the only thing he “confirmed” are his own preconceived notions; he did not ‘prove’ what he set out to.
Is Racial Superiority in Sports a Myth? A Response to Kerr (2010)
2750 words
Racial differences in sporting success are undeniable. The races are somewhat stratified in different sports and we can trace the cause of this to differences in genes and where one’s ancestors were born. We can then say that there is a relationship between them since, they have certain traits which their ancestors also had, which then correlate with geographic ancestry, and we can explain how and why certain populations dominate (or would have the capacity to based on body type and physiology) certain sporting events. Critiques of Taboo: Why Black Athletes Dominate Sports and Why We’re Afraid to Talk About It are few and far between, and the few that I am aware of are alright, but this one I will discuss today is not particularly good, because the author makes a lot of claims he could have easily verified himself.
In 2010, Ian Kerr published The Myth of Racial Superiority in Sports, who states that there is a “dark side” to sports, and specifically sets his sights on Jon Entine’s (2000) book Taboo. In this article, Kerr (2010) makes a lot of, in my opinion, giant claims which provide a lot of evidence and arguments in order to show their validity. I will discuss Kerr’s views on race, biology, the “environment”, “genetic determinism”, and racial dominance in sports (which will have a focus on sprinting/distance running in this article).
Race
Since establishing the reality and validity of the concept of race is central to proving Entine’s (2002) argument on racial differences in sports, then I must prove the reality of race (and rebut what Kerr 2010 writes about race). Kerr (2010: 20) writes:
First, it is important to note that Entine is not working in a vacuum; his assertions about race and sports are part of a larger ongoing argument about folk notions of race. Folk notions of race founded on the idea that deep, mutually exclusive biological categories dividing groups of people have scientific and cultural merit. This type of thinking is rooted in the notion that there are underlying, essential differences among people and that those observable physical differences among people are rooted in biology, in genetics (Ossorio, Duster, 2005: 2).
Dividing groups of people does have scientific, cultural and philosophical merit. The concept of “essences” has long been discarded by philosophers. Though there are differences in both anatomy and physiology in people that differ by geographic location, and this then, at the extreme end, would be enough to cause the differences in elite sporting competition that is seen.
Either way, the argument for the existence of race is simple: 1) populations differ in physical attributes (facial, morphological) which then 2) correlate with geographic ancestry. Therefore, race has a biological basis since the physical differences between these populations are biological in nature. Now that we have established that race exists using only physical features, it should be extremely simple to show how Kerr (2010) is in error with his strong claims regarding race and the so-called “mythology” of racial superiority in sports. Race is biological; the biological argument for race is sound (read here and here, and also see Hardimon, 2017).
Genetic determinism
True genetic determinism—as is commonly thought—does not have any sound, logical basis (Resnick and Vorhaus, 2006). So Kerr’s (2010) claims in this section need to be dissected here. This next quote, though, is pretty much imperative to the soundness and validity of his whole article, and let’s just say that it’s easy to rebut and invalidates his whole entire argument:
Vinay Harpalani is one of the most outspoken critics of using genetic determinism to validate notions of inferiority or the superiority of certain groups (in this case Black athletes). He argues that in order for any of Entine’s claims to be valid he must prove that: 1) there is a systematic way to define Black and White populations; 2) consistent and plausible genetic differences between the populations can be demonstrated; 3) a link between those genetic differences and athletic performance can be clearly shown (2004).
This is too easy to prove.
1) While I do agree that the terminology of ‘white’ and ‘black’ are extremely broad, as can be seen by looking at Rosenberg et al (2002), population clusters that cluster with what we call ‘white’ and ‘black’ exist (and are a part of continental-level minimalist races). So is there a systematic way to define ‘Black’ and ‘White’ populations? Yes, there is; genetic testing will show where one’s ancestors came from recently, thereby proving point 1.
2) Consistent and plausible genetic differences between populations can be demonstrated. Sure, there is more variation within races than between them (Lewontin, 1972; Rosenberg et al, 2002; Witherspoon et al, 2007; Hunley, Cabana, and Long, 2016). Even these small between-continent/group differences would have huge effects on the tail end of said distribution.
3) I have compiled numerous data on genetic differences between African ethnies and European ethnies and how these genetic differences then cause differences in elite athletic performance. I have shown that Jamaicans, West Africans, Kenyans and Ethiopians (certain subgroups of the two aforementioned countries) have genetic/somatypic differences that then lead to differences in these sporting competitions. So we can say that race can predict traits important for certain athletic competitions.
1) The terminology of ‘White’ and ‘Black’ are broad; but we can still classify individuals along these lines; 2) consistent and plausible genetic differences between races and ethnies do exist; 3) a link between these genetic differences between genes/athletic differences between groups can be found. Therefore Entine’s (2002) arguments—and the validity thereof—are sound.
Kerr (2010) then makes a few comments on the West’s “obsession with superficial physical features such as skin color”, but using Hardimon’s minimalist race concept, skin color is a part of the argument to prove the existence and biological reality of race, therefore skin color is not ‘superficial’, since it is also a tell of where one’s ancestors evolved in the recent past. Kerr (2010: 21) then writes:
Marks writes that Entine is saying one of three things: that the very best Black athletes have an inherent genetic advantage over the very best White athletes; that the average Black athlete has a genetic advantage over the average White athlete; that all Blacks have the genetic potential to be better athletes than all Whites. Clearly these three propositions are both unknowable and scientifically untenable. Marks writes that “the first statement is trivial, the secondly statistically intractable, and the third ridiculous for its racial essentialism” (Marks, 2000: 1077).
The first two, in my opinion (the very best black athletes have an inherent genetic advantage over the very best white athletes and the average black athlete has a genetic advantage over the average white athlete), are true, and I don’t know how you can deny this; especially if you’re talking about AVERAGES. The third statement is ridiculous, because it doesn’t work like that. Kerr (2010), of course, states that race is not a biological reality, but I’ve proven that it is so that statement is a non-factor.
Kerr (2010) then states that “ demonstrating across the board genetic variations between
populations — has in recent years been roundly debunked“, and also says “ Differences in height, skin color, and hair texture are simply the result of climate-related variation.” This is one of the craziest things I’ve read all year! Differences in height would cause differences in elite sporting competition; differences in skin color can be conceptualized as one’s ancestors’ multi-generational adaptation to the climate they evolved in as can hair texture. If only Kerr (2010) knew that this statement here was the beginning of the end of his shitty argument on Entine’s book. Race is a social construct of a biological reality, and there are genetic differences between races—however small (Risch et al, 2002; Tang et al, 2005) but these small differences can mean big differences at the elite level.
The “environment” and biological variability
Kerr (2010) then shifts his focus over to, not genetic differences, but biological differences. He specifically discusses the Kenyans—Kalenjin—stating that “height or weight, which play an instrumental role in helping define an individual’s athletic prowess, have not been proven to be exclusively rooted in biology or genetics.” While estimates of BMI and height are high (both around .8), I think we can disregard the numbers since they came from highly flawed twin studies, since molecular genetic evidence shows lower heritabilities. Either way, surely height is strongly influenced by ‘genes’. Another important caveat is that Kenya has one of the lowest BMIs in the world, 20.7 for Kenyan men, which also is part of the cause of why certain African ethnies dominate running competitions.
I don’t disagree with Kerr (2010) here too much; many papers show that SES/cultural/social factors are very important to Kenyan runners (Onywera et al, 2006; Wilbur and Pistiladis, 2012; Tucker, Onywera, and Santos-Concejero, 2015). You can have all of the ‘physical gifts’ in the world, if it’s not combined with the will to want to do your best, along with cultural and social factors you won’t succeed. But having an advantageous genotype and physique are useless without a strong mind (Lippi, Favaloro, and Guidi, 2008):
An advantageous physical genotype is not enough to build a top-class athlete, a champion capable of breaking Olympic records, if endurance elite performances (maximal rate of oxygen uptake, economy of movement, lactate/ventilatory threshold and, potentially, oxygen uptake kinetics) (Williams & Folland, 2008) are not supported by a strong mental background.”
Dissecting this, though, is tougher. Because being born at certain altitudes will cause certain advantageous traits, such as a larger lung capacity (and you will have an advantage in lung capacity when competing at lower altitudes), but certain subpopulations live in these high-altitude areas, so what is it? Genetic? Cultural? Environmental? All three? Nature vs nurture is a false dichotomy; so it is a mixture of the three.
How does one explain, then, the athlete who trains countless hours a day fine-tuning a jump shot, like LeBron James or shaving seconds off sub-four minute miles like Robert Kipkoech Cheruiyot, a four time Boston Marathon winner?
Literally no one denies that elite athletes put in insane amounts of practice; but if everyone has the same amount of practice they won’t have similar abilities.
He also briefly brings up muscle fibers, stating:
These include studies on African fast twitch muscle fibers and development of motor skills. Entine includes these studies to demonstrate irrevocable proof of embedded genetic differences between populations but refuses to accept the fact that any differences may be due to environmental factors or training.
This, again, shows ignorance of the literature. An individual’s muscle fibers are formed during development from the fusion of several myoblasts, with differentiation being completed before birth. Muscle fiber typoing is also set at age 6, no difference in skeletal muscle tissue was found when comparing 6-year-olds and adults, therefore we can state that muscle fiber typing is set by age 6 (Bell et al, 1980). You can, of course, train type II fibers to have similar aerobic capacity to type I fibers, but they’ll never be fully similar. This is something that Kerr (2010) obviously is ignorant to because he’s not well-read on the literature which causes him to make dumb statements like “any differences [in muscle fiber typing] may be due to environmental factors or training“.
Black domination in sports
Finally, Kerr (2010) discusses the fact that whites dominated certain running competitions in the Olympics and that before the 1960s, a majority of distance-running gold medals went to white athletes. He then states that the 2008 Boston Marathon winner was Kenyan; but the next 4 behind him were not. Now, let’s check out the 2017 Marathon winners: Kenya, USA, Japan for the top 3; while 5 Kenyans/Ethiopians are in the top 15 while the same is also true of women; a Kenyan winner, with Kenyans/Ethiopians taking 5 of the top 15 spots. The fact that whites used to do well in running sports is a non-factor; Jesse Owens blew away the competition in the Games in Germany, which showed how blacks would begin to dominate in the US decades later.
Kerr (2010) then ends the article with a ton of wild claims; the wildest one, in my opinion, being that “Kenyans are no more genetically different from any other African or European population on average“, does anyone believe this? Because I have data to the contrary. They have a higher Vo2 max, which of course is trainable but with a ‘genetic’ component (Larsen, 2003), while other authors argue that genetic differences between populations account for differences in success in running competition between populations (Vancini et al, 2014), while male and female Kenyan and Ethiopian runners are the fastest in the half and full marathon (Knechtle et al, 2016). There is a large amount of data out there that speaks about Kenyan/Ethiopian and others’ dominance in running; it seems Kerr (2010) just ignored the data. I agree with Kerr that Kenyanholos show that humans can adapt to their environment; but his conclusion here:
The fact that runners coming from Kenya do so well in running events attests to the fact the combination of intense high altitude training, consumption of a low-fat, high protein diet, and a social and cultural expectation to succeed have created in recent decades an environment which is highly conducive to producing excellent long-distance runners.
is very strong, and while I don’t disagree at all with anything here, he’s disregarding how somatype and genes differ between Kenyans and other populations that compete in these sports that then lead to differences in elite sporting competitions.
Elite sporting performance is influenced by myriad factors, including psychology, ‘environment’, and genetic factors. Something that Kerr (2010) doesn’t understand—because he’s not well-read on this literature—is that many genetic factors that influence sporting performance are known. The ability to become elite depends on one’s capacity for endurance, muscle performance, the ability of the tendons and ligaments to withstand stress and injury, and the attitude to train and push above and beyond what normal people can do (Lippi, Longo, and Maffulli, 2010). We can then extend this to human races; some are better-equipped to excel in running competitions than others.
On its face, Kerr’s (2010) claim that there are no inherent differences between races is wrong. Races differ in somatype, which is due to evolution in different geographic locations for tens of thousands of years. The human body is perfectly adapted to for long distance running (Murray and Costa, 2012), and since our capabilities for endurance running evolved in Africa and they, theoretically, have a musculoskeletal structure similar to the Homo sapiens that left Africa around 70 kya, then it’s only logical to state that African’s, on average, have an inherent ability in running competitions (West and East Africans, while North Africans fare very well in middle distance running, which, again, comes down to living in higher altitudes like Kenyans and Ethiopians).
Wagner and Heyward (2000) reviewed many studies on the physiological differences between blacks and whites. Blacks skew towards mesomorphy; black youths had smaller billiac and bitrochanteric width (the widest measure of the pelvis at the outer edges and the flat process on the femur, respectively), and black infants had longer extremities than white infants (Wagner and Heyward, 2000). We have anatomic evidence that blacks are superior runners (in an American context). Mesomorphic athletes are more likely to be sprinters (Sands et al, 2005; which is also seen in prepubescent children: Marta et al, 2013) Kenyans are ecto-dominant (Vernillo et al, 2013) which helps to explain their success at long-distance running. So just on only looking at the phenotype (a marker for race with geographic ancestry, proving the biological existence of race) we can confidently state, on average just by looking at an individual or a population, how they will fare in certain competitions.
Conclusion
Kerr’s (2010) arguments leave a ton to be desired. Race exists and is a biological reality. I don’t know why this paper got published since it was so full of errors; his arguments were not sound and much of the literature contradicts his claims. What he states at the end about Kenyans is not wrong at all, but to not even bring up genetic/biologic differences as a factor influencing their performance is dishonest.
Of course, a whole slew of factors, be they biological, cultural, psychological, genetic, socioeconomic, anatomic, physiologic etc influence sporting performance, but certain traits are more likely to be found in certain populations, and in the year 2018 we have a good idea of what influences elite sporting performance and what does not. It just so happens that these traits are unevenly distributed between populations, and the cause is evolution in differing climates in differing geographic locations.
Race exists and is a biological reality. Biological anatomic/physiological differences between these races then manifest themselves in elite sporting competition. The races differ, on average, in traits important for success in certain competitions. Therefore, race explains some of the variance in elite sporting competition.
Lead, Race, and Crime
2500 words
Lead has many known neurological effects on the brain (regarding the development of the brain and nervous system) that lead to many deleterious health outcomes and negative outcomes in general. Including (but not limited to) lower IQ, higher rates of crime, higher blood pressure and higher rates of kidney damage, which have permanent, persistent effects (Stewart et al, 2007). Chronic lead exposure, too, can “also lead to decreased fertility, cataracts, nerve disorders, muscle and joint pain, and memory or concentration problems” (Sanders et al, 2009). Lead exposure in vitro, in infancy, and in childhood can also lead to “neuronal death” (Lidsky and Schneider, 2003). While epigenetic inheritance also plays a part (Sen et al, 2015). How do blacks and whites differ in exposure to lead? How much is the difference between the two races in America, and how much would it contribute to crime? On the other hand, China has high rates of lead exposure, but lower rates of crime, so how does this relationship play out with the lead-crime relationship overall? Are the Chinese an outlier or is there something else going on?
The effects of lead on the brain are well known, and numerous amounts of effort have been put into lowering levels of lead in America (Gould, 2009). Higher exposure to lead is also found in poorer, lower class communities (Hood, 2005). So since higher levels of lead exposure are found more often in lower-class communities, then blacks should have higher blood-lead levels than whites. This is what we find.
Blacks had a 27 percent higher concentration of lead in their tibia, while having significantly higher levels of blood lead, “likely because of sustained higher ongoing lead exposure over the decades” (Theppeang et al, 2008). Other data—coming out of Detroit—shows the same relationships (Haar et al, 1979; Talbot, Murphy, and Kuller, 1982; Lead poisoning in children under 6 jumped 28% in Detroit in 2016; also see Maqsood, Stanbury, and Miller, 2017) while lead levels in the water contribute to high levels of blood-lead in Flint, Michigan (Hanna-Attisha et al, 2016; Laidlaw et al, 2016). Cassidy-Bushrow et al (2017) also show that “The disproportionate burden of lead exposure is vertically transmitted (i.e., mother-to-child) to African-American children before they are born and persists into early childhood.”
Children exposed to lead have lower brain volumes as children, specifically in the ventrolateral prefrontal cortex, which is the same region of the brain that is impaired in antisocial and psychotic persons (Cecil et al, 2008). The community that was tested was well within the ‘safe’ range set by the CDC (Raine, 2014: 224), though the CDC says that there is no safe level of lead exposure. There is a large body of studies which show that there is no safe level of lead exposure (Needleman and Landrigan, 2004; Canfield, Jusko, and Kordas, 2005; Barret, 2008; Rossi, 2008; Abelsohn and Sanborn, 2010; Betts, 2012; Flora, Gupta, and Tiwari, 2012; Gidlow, 2015; Lanphear, 2015; Wani, Ara, and Usmani, 2015; Council on Environmental Health, 2016; Hanna-Attisha et al, 2016; Vorvolakos, Aresniou, and Samakouri, 2016; Lanphear, 2017). So the data is clear that there is absolutely no safe level of lead exposure, and even small effects can lead to deleterious outcomes.
Further, one brain study of 532 men who worked in a lead plant showed that those who had higher levels of lead in their bones had smaller brains, even after controlling for confounds like age and education (Stewart et al, 2008). Raine (2014: 224) writes:
The fact that the frontal cortex was particularly reduced is very interesting, given that this brain region is involved in violence. This lead effect was equivalent to five years of premature aging of the brain.
So we have good data that the parts of the brain that relate to violent tendencies are reduced in people exposed to more lead had the same smaller parts of the brain, indicating a relationship. But what about antisocial disorders? Are people with higher levels of lead in their blood more likely to be antisocial?
Needleman et al (1996) show that boys who had higher levels of lead in their blood had higher teacher ratings of aggressive and delinquent behavior, along with higher self-reported ratings of aggressive behavior. Even high blood-lead levels later in life is related to crime. One study in Yugoslavia showed that blood lead levels at age three had a stronger relationship with destructive behavior than did prenatal blood lead levels (Wasserman et al, 2008); with this same relationship being seen in America with high blood lead levels correlating with antisocial and aggressive behavior at age 7 and not age 2 (Chen et al 2007).
Nevin (2007) showed a strong relationship between preschool lead exposure and subsequent increases in criminal cases in America, Canada, Britain, France, Australia, Finland, West Germany, and New Zealand. Reyes (2007) also shows that crime increased quicker in states that saw a subsequent large decrease in lead levels, while variations in lead levels within cities correlating with variations in crime rates (Mielke and Zahran, 2012). Nevin (2000) showed a strong relationship between environmental lead levels from 1941 to 1986 and corresponding changes to violent crime twenty-three years later in the United States. Raine (2014: 226) writes (emphasis mine):
So, young children who are most vulnerable to lead absorption go on twenty-three years later to perpetrate adult violence. As lead levels rose throughout the 1950s, 1960s, and 1970s, so too did violence correspondingly rise in the 1970s, 1980s and 1990s. When lead levels fell in the late 1970s and early 1980s, so too did violence fall in the 1990s and the first decade of the twenty-first century. Changes in lead levels explained a full 91 percent of the variance in violent offending—an extremely strong relationship.
[…]
From international to national to state to city levels, the lead levels and violence curves match up almost exactly.
But does lead have a causal effect on crime? Due to the deleterious effects it has on the developing brain and nervous system, we should expect to find a relationship, and this relationship should become stronger with higher doses of lead. Fortunately, I am aware of one analysis, a sample that’s 90 percent black, which shows that with every 5 microgram increase in prenatal blood-lead levels, that there was a 40 percent higher risk of arrest (Wright et al, 2008). This makes sense with the deleterious developmental effects of lead; we are aware of how and why people with high levels of lead in their blood show similar brain scans/brain volume in certain parts of the brain in comparison to antisocial/violent people. So this is yet more suggestive evidence for a causal relationship.
Jennifer Doleac discusses three studies that show that blood-lead levels in America need to be addressed, since they are related strongly to negative health outcomes.Aizer and Curry (2017) show that “A one-unit increase in lead increased the probability of suspension from school by 6.4-9.3 percent and the probability of detention by 27-74 percent, though the latter applies only to boys.” They also show that children who live nearer to roads have higher blood-lead levels, since the soil near highways was contaminated decades ago with leaded gasoline. Fiegenbaum and Muller (2016) show that cities’ use of lead pipes increased murder rates between the years o921 and 1936. Finally, Billings and Schnepnel (2017: 4) show that their “results suggest that the effects of high levels of [lead] exposure on antisocial behavior can largely be reversed by intervention—children who test twice over the alert threshold exhibit similar outcomes as children with lower levels of [lead] exposure (BLL<5μg/dL).”
A relationship with lead exposure in vitro and arrests at adulthood. The sample was 90 percent black, with numerous controls. They found that prenatal and post-natal blood-lead exposure was associated with higher arrest rates, along with higher arrest rates for violent acts (Wright et al, 2008). To be specific, for every 5 microgram increase in prenatal blood-lead levels, there was a 40 percent greater risk for arrest. This is direct causal evidence for the lead-causes-crime hypothesis.
One study showed that in post-Katrina New Orleans, decreasing lead levels in the soil caused a subsequent decrease in blood lead levels in children (Mielke, Gonzales, and Powell, 2017). Sean Last argues that, while he believes that lead does contribute to crime, that the racial gaps have closed in the recent decades, therefore blood-lead levels cannot be a source of some of the variance in crime between blacks and whites, and even cites the CDC ‘lowering its “safe” values’ for lead, even though there is no such thing as a safe level of lead exposure (references cited above). White, Bonilha, and Ellis Jr., (2015) also show that minorities—blacks in particular—have higher rates of lead in their blood. Either way, Last seems to downplay large differences in lead exposure between whites and blacks at young ages, even though that’s when critical development of the mind/brain and other important functioning occurs. There is no safe level of lead exposure—pre- or post-natal—nor are there safe levels at adulthood. Even a small difference in blood lead levels would have some pretty large effects on criminal behavior.
Sean Last also writes that “Black children had a mean BLL which was 1 ug/dl higher than White children and that this BLL gap shrank to 0.9 ug/dl in samples taken between 2003 and 2006, and to 0.5 ug/dl in samples taken between 2007 and 2010.” Though, still, there are problems here too: “After adjustment, a 1 microgram per deciliter increase in average childhood blood lead level significantly predicts 0.06 (95% confidence interval [CI] = 0.01, 0.12) and 0.09 (95% CI = 0.03, 0.16) SD increases and a 0.37 (95% CI = 0.11, 0.64) point increase in adolescent impulsivity, anxiety or depression, and body mass index, respectively, following ordinary least squares regression. Results following matching and instrumental variable strategies are very similar” (Winter and Sampson, 2017).
Naysayers may point to China and how they have higher levels of blood-lead levels than America (two times higher), but lower rates of crime, some of the lowest in the world. The Hunan province in China has considerably lowered blood-lead levels in recent years, but they are still higher than developed countries (Qiu et al, 2015). One study even shows ridiculously high levels of lead in Chinese children “Results showed that mean blood lead level was 88.3 micro g/L for 3 – 5 year old children living in the cities in China and mean blood lead level of boys (91.1 micro g/L) was higher than that of girls (87.3 micro g/L). Twenty-nine point nine one per cent of the children’s blood lead level exceeded 100 micro g/L” (Qi et al, 2002), while Li et al (2014) found similar levels. Shanghai also has higher levels of blood lead than the rest of the developed world (Cao et al, 2014). Blood lead levels are also higher in Taizhou, China compared to other parts of the country—and the world (Gao et al, 2017). But blood lead levels are decreasing with time, but still higher than other developed countries (He, Wang, and Zhang, 2009).
Furthermore, Chinese women, compared to American women, had two times higher BLL (Wang et al, 2015). With transgenerational epigenetic inheritance playing a part in the inheritance of methylation DNA passed from mother to daughter then to grandchildren (Sen et al, 2015), this is a public health threat to Chinese women and their children. So just by going off of this data, the claim that China is a safe country should be called into question.
Reality seems to tell a different story. It seems that the true crime rate in China is covered up, especially the murder rate:
In Guangzhou, Dr Bakken’s research team found that 97.5 per cent of crime was not reported in the official statistics.
Of 2.5 million cases of crime, in 2015 the police commissioner reported 59,985 — exactly 15 less than his ‘target’ of 60,000, down from 90,000 at the start of his tenure in 2012.
The murder rate in China is around 10,000 per year according to official statistics, 25 per cent less than the rate in Australia per capita.
“I have the internal numbers from the beginning of the millennium, and in 2002 there were 52,500 murders in China,” he said.Instead of 25 per cent less murder than Australia, Dr Bakken said the real figure was closer to 400 per cent more.”
Guangzhou, for instance, doesn’t keep data for crime committed by migrants, who commit 80 percent of the crime in this province. Out of 2.5 million crimes committed in Guangzhou, only 5,985 crimes were reported in their official statistics, which was 15 crimes away from their target of 6000. Weird… Either way, China doesn’t have a similar murder rate to Switzerland:
The murder rate in China does not equal that of Switzerland, as the Global Times claimed in 2015. It’s higher than anywhere in Europe and similar to that of the US.
China also ranks highly on the corruption index, higher than the US, which is more evidence indicative of a covered up crime rate. So this is good evidence that, contrary to the claims of people who would attempt to downplay the lead-crime relationship, that these effects are real and that they do matter in regard to crime and murder.
So it’s clear that we can’t trust the official Chinese crime stats since there much of their crime is not reported. Why should we trust crime stats from a corrupt government? The evidence is clear that China has a higher crime—and murder rate—than is seen on the Chinese books.
Lastly, effects of epigenetics can and do have a lasting effect on even the grandchildren of mothers exposed to lead while pregnant (Senut et al, 2012; Sen et al, 2015). Sen et al (2015) showed lead exposure during pregnancy affected the DNA methylation status of the fetal germ cells, which then lead to altered DNA methylation on dried blood spots in the grandchildren of the mother exposed to lead while pregnant.—though it’s indirect evidence. If this is true and holds in larger samples, then this could be big for criminological theory and could be a cause for higher rates of black crime (note: I am not claiming that lead exposure could account for all, or even most of the racial crime disparity. It does account for some, as can be seen by the data compiled here).
In conclusion, the relationship between lead exposure and crime is robust and replicated across many countries and cultures. No safe level of blood lead exists, even so-called trace amounts can have horrible developmental and life outcomes, which include higher rates of criminal activity. There is a clear relationship between lead increases/decreases in populations—even within cities—that then predict crime rates. Some may point to the Chinese as evidence against a strong relationship, though there is strong evidence that the Chinese do not report anywhere near all of their crime data. Epigenetic inheritance, too, can play a role here mostly regarding blacks since they’re more likely to be exposed to high levels of lead in the womb, their infancy, and childhood. This could also exacerbate crime rates, too. The evidence is clear that lead exposure leads to increased criminal activity, and that there is a strong relationship between blood lead levels and crime.
Is There Really More Variation Within Races Than Between Them?
1500 words
In 1972 Richard Lewontin, studying the blood groups of different races, came to the conclusion that “Human racial classification is of no social value and is positively destructive of social and human relations. Since such racial classification is now seen to be of virtually no genetic or taxonomic significance either, no justification can be offered for its continuance” (pg 397). He also found that “the difference between populations within a race account for an additional 8.3 percent, so that only 6.3 percent is accounted for by racial classification.” This has lead numerous people to, along with Lewontin, conclude that race is ‘of virtually no genetic or taxonomic significance’ and conclude that, due to this, race does not exist.
Lewontin’s main reasoning was that since there is more variation within races than between them (85 percent of differences were within populations while 15 percent was within them) then since a lion’s share of human diversity is distributed within races, not between them, then race is of no genetic nor taxonomic use. Lewontin is correct that there is more variation within races than between them, but he is incorrect that this means that racial classification ‘is of no social value’, since knowing and understanding the reality of race (even our perceptions of them, whether they are true or not) influence things such as medical outcomes.
Though, like Lewontin, people have cited this paper as evidence against the existence of human races, for if there is way more genetic variation between races, and that most human genetic variation is within races, then race cannot be of any significance for things such as medical outcomes since most genetic variation is within races not between them.
Rosenberg et al (2002) also confirmed and replicated Lewontin’s analysis, showing that within-population genetic variation accounts for 93-95 percent of human genetic variation, while 3 to 5 percent of human genetic variation lies between groups. Philosopher Michael Hardimon (2017) uses these arguments to buttress his point that ‘racialist races’ (as he calls them) do not exist. His criteria being:
(a) The fraction of human genetic diversity between populations must exceed the fraction of diversity between them.
(b) The fraction of human genetic diversity within populations must be small.
(c) The fraction of diversity between populations must be large.
(d) Most genes must be highly differentiated by race.
(e) The variation in genes that underlie obvious physical differences must be typical of the genome in general.
(f) There must be several important genetic differences between races apart from the genetic differences that underlie obvious physical differences.
Note: (b) says that racialist races are genetically racially homogeneous groups; (c)-(f) say that racialist races are distinguised by major biological differences.
Call (a)-(f) the racialist concept of race’s genetic profile. (Hardimon, 2017: 21)
He clearly strawmans the racialist position, but I’ll get into that another day. Hardimon writes about how both of these studies lend credence to his above argument on racialist races (pg 24):
Rosenberg and colleagues also confirm Lewontin’s findings that most genes are not highly differentiated by race and that the variation in genes that underlie obvious physical differences is not typical of the variation of the genome in general. They also suggest that it is not the case that there are many important genetic differences between races apart from the genetic differences that underlie the obvious physical differences. These considerations further buttress the case against the existence of racialist races.
[…]
The results of Lewontin’s 1972 study and Rosenberg and colleagues’ 2002 study strongly suggest that it is extremely unlikely that there are many important genetic differences between races apart from the genetic differences that underlie the obvious physical differences.
(Hardimon also writes on page 124 that Rosenberg et al’s 2002 study could also be used as evidence for his populationist concept of race, which I will return to in the future.)
Though, my reasoning for writing this article is to show that the findings by Lewontin and Rosenberg et al regarding more variation within races than between them are indeed true, despite claims to the contrary. There is one article, though, that people cite as evidence against the conclusions by Lewontin and Rosenberg et al, though it’s clear that they only read the abstract and not the full paper.
Witherspoon et al (2007) write that “sufficient genetic data can permit accurate classification of individuals into populations“, which is what the individuals who cite this study as evidence for their contention mean, though they conclude (emphasis mine):
The fact that, given enough genetic data, individuals can be correctly assigned to their populations of origin is compatible with the observation that most human genetic variation is found within populations, not between them. It is also compatible with our finding that, even when the most distinct populations are considered and hundreds of loci are used, individuals are frequently more similar to members of other populations than to members of their own population. Thus, caution should be used when using geographic or genetic ancestry to make inferences about individual phenotypes.
Witherspoon et al (2007) analyzed the three classical races (Europeans, Africans and East Asians) over thousands of loci and came to the conclusion when genetic similarity is measured over thousands of loci, the answer to the question “How often is a pair of individuals from one population genetically more dissimilar than two individuals chosen from two different populations?” is “never“.
Hunley, Cabana, and Long (2016: 7) also confirm Lewontin’s analysis, writing “In sum, we concur with Lewontin’s conclusion that Western-based racial classifications have no taxonomic significance, and we hope that this research, which takes into account our current understanding of the structure of human diversity, places his seminal finding on firmer evolutionary footing.” But the claim that “racial classifications have no taxonomic significance” is FALSE.
This is a point that Edwards (2003) rebutted in depth. While he did agree with Lewontin’s (1972) analysis that there was more variation within races than between them (which was confirmed through subsequent analysis), he strongly disagreed with Lewontin’s conclusion that race is of no taxonomic significance. Richard Dawkins, too disagreed with Lewontin, though as Dawkins writes in his book The Ancestors Tale: “Most of the variation among humans can be found within races as well as between them. Only a small admixture of extra variation distinguishes races from each other. That is all correct. What is not correct is the inferene that race is therefore a meaningless concept.” The fact that there is more variation within races than between them is irrelevant to taxonomic classification, and classifying races by phenotypic differences (morphology, and facial features) along with geographic ancestry shows that just by looking at the average phenotype that race exists, though these concepts make no value-based judgements on anything you can’t ‘see’, such as mental and personality differences between populations.
Though while some agree with Edwards’ analysis of Lewontin’s argument about race’s taxonomic significance, they don’t believe that he successfully refuted Lewontin. For instance, Hardimon (2017: 22-23) writes that Lewontin’s argument against—what Hardimon (2017) calls ‘racialist race’ (his strawman quoted above)—the existence of race because the within-race component of genetic variation is greater than the genetic variation between races “is untouched by Edwards’ objections.”
Though Sesardic (2010: 152) argues that “Therefore, contra Lewontin, the racial classification that is based on a number of genetic differences between populations may well be extremely reliable and robust, despite the fact that any single of those genetic between-population differences\ remains, in itself, a very poor predictor of racial membership.” He also states that the 7 to 10 percent difference between populations “actually refers to the inter-racial portion of variation that is averaged over the separate contributions of a number of individual genetic indicators that were sampled in different studies” (pg 150).
I personally avoid all of this talk about genes/allele frequencies between populations and jump straight to using Hardimon’s minimalist race concept—a concept that, according to Hardimon is “stripped down to its barest bones” since it captures enough of the racialist concept of race to be considered a race concept.
In sum, variation within races is greater than variation between races, but this does not mean anything for the reality of race since race can still be delineated based on peculiar physical features and peculiar geographic ancestry to that group. Using a few indicators (morphology, facial features such as nose, lips, cheekbones, facial structure, and hair along with geographic ancestry), we can group races based on these criteria and we can show that race does indeed exist in a physical—not social—sense and that these categories are meaningful in a medical context (Hardimon, 2013, 2017). So even though genetic variation is greater within races than between them, this does not mean that there is no taxonomic significance to race, as other authors have argued. Hardimon (2017: 23) agrees, writing (emphasis his) “… Lewontin’s data do not preclude the possibility that racial classification might have taxonomic significance, but they do preclude the possibility that racialist races exist.”
Hardimon’s strawman of the racialist concept notwithstanding (which I will cover in the future), his other three race concepts (minimalist, populationist and socialrace concepts) are logically sound and stand up to a lot of criticism. Either way, race does exist, and it does not matter if the apportionment of human genetic diversity is greatest within races than between them.
