
Home » Race Realism (Page 2)
Category Archives: Race Realism
Strategies for Achieving Racial Health Equity: An Argument for When Health Inequalities are Health Inequities
2100 words
Introduction
Health can be defined as “a relative state in which one is able to function well physically, mentally, socially, and spiritually to express the full range of one’s unique potentialities within the environment in which one lives” (Svalastog et al, 2017). Health, clearly, is a multi-dimensional concept (Barr, 2014). Since there are many kinds of referents to the word “health”, it is therefore essential to understand and consider the context and perspective of each person and group when discussing health-related issues and also while implementing healthcare policies and practices.
Inequality exists everywhere on earth and it manifests in numerous forms like in income, health, education, and healthcare access. So the existence of inequality is undeniable, since we can see it with our own eyes, but understanding the mechanisms that lead to inequality should be multifaceted. Central to the understanding of inequality is the relationship between inequality, unfairness and the role of empirical investigations in uncovering not only the implications for societal outcomes, but also in discerning what is an inequity (which is a kind of inequality that is avoidable, unfair and unjust). According to Braveman, (2003: 182):
Health inequities are disparities in health or its social determinants that favour the social groups that were already more advantaged. Inequity does not refer generically to just any inequalities between any population groups, but very specifically to disparities between groups of people categorized a priori according to some important features of their underlying social position.
Talking about health is the best way to understand what inequity actually is. The issue is, true equality of health is impossible, but what is possible is addressing the actual social determinants of health (SDoH). What is also important is understanding what equity is and what equity isn’t, as I have argued in the past. Grifters like James Lindsay and Chris Rufo (along with well-meaning but still wrong institutions) believe that equity is ensuring equal outcomes. This is incorrect. What equity means—in the health sphere—is when “the opportunity to ‘attain their full health potential’ and no one is ‘disadvantaged from achieving this potential because of their social position or other socially determined circumstance’” (Braveman, quoted by the CDC).Therefore, health equity is when everyone has the chance to reach their fill potential, unabated by social determinants. Social conditions and policies strongly influence the health of both individuals and groups and it’s the result of unequal distribution of resources and opportunities. Empirical investigation is pivotal in understanding if a certain inequality is an inequity. And although inequities are a kind of inequality, “inequality” and “inequity” are conceptually distinct (Braveman, 2003).
In this article I will discuss the SDoH, give my argument that we can identify inequity (a kind of inequality) through empirical investigations (meaning that they are avoidable, unfair and unjust). I will then pivot to a real-world example of my argument—that of low birth weight in black American newborns and argue that racism and historical injustices can explain that since non-American black women have children with higher mean birth weights. I will then discuss how blacks who have doctors of of the same race report better care and have higher life expectancies. I will then discuss what can be done about this—and the answer is to educate people on genetic essentialism which leads to racism and racist attitudes.
On SDoH
SDoH include the lack of education, racism, lack of access to health care and poverty. Barr (2011: 64) had a helpful flow chart to understand this issue.
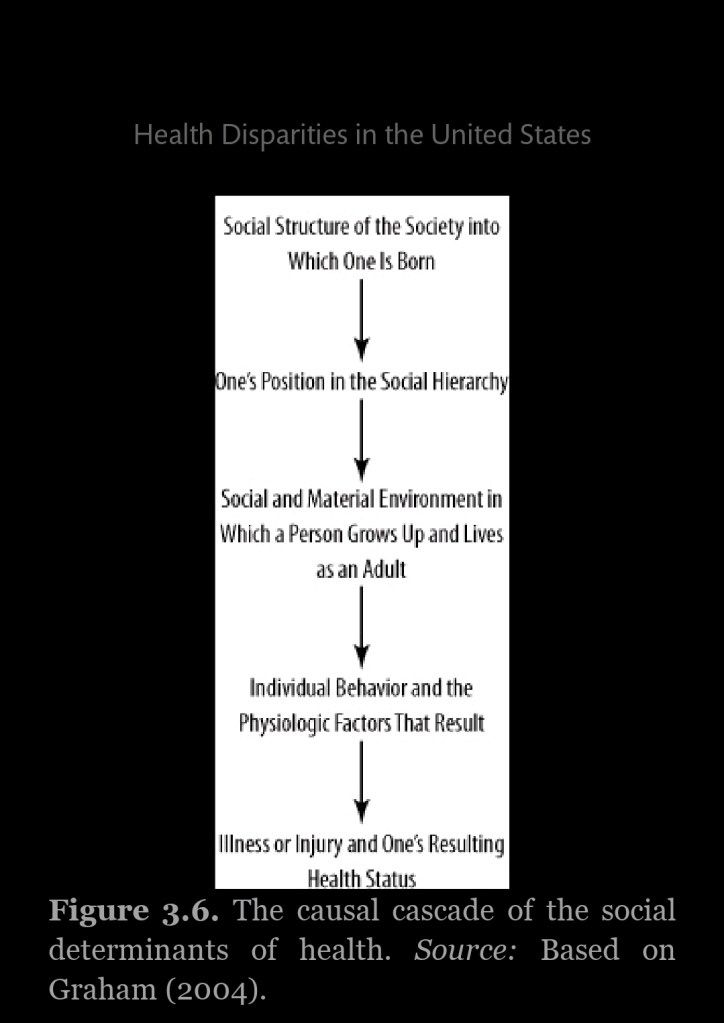
So when it comes to variation in health outcomes, we know that only 20 percent can be attributed to access to medical care, while a whooping 80 percent is attributable to the SDoH:
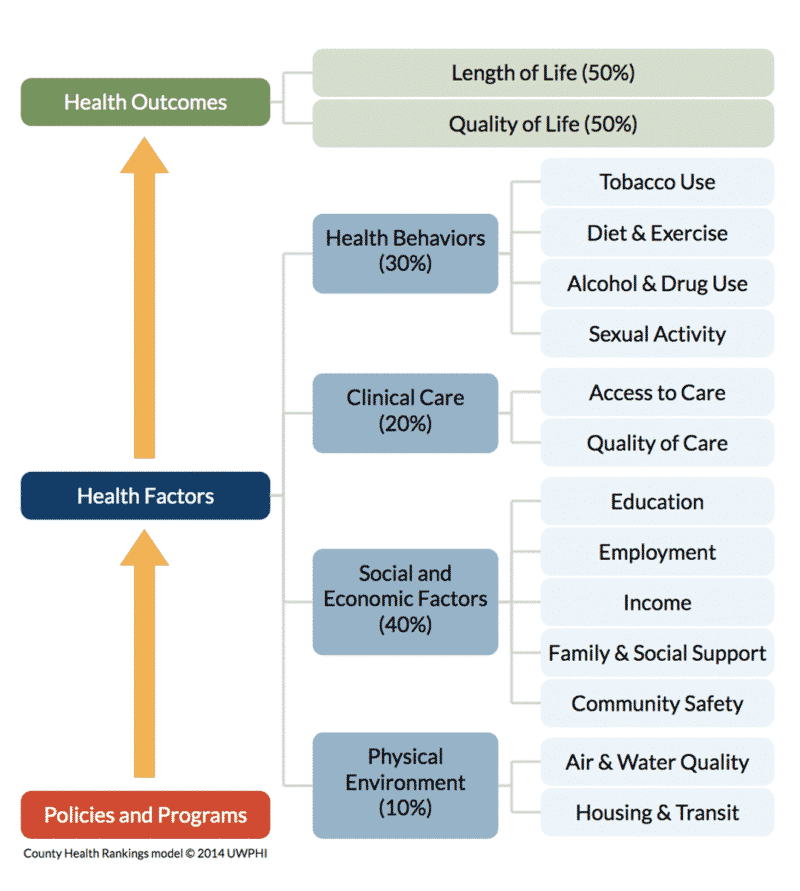
(Ratcliffe, 2017 also states that about 20 percent of the health of nation is attributed to medical care, 5 percent the result of biology and genes, 20 percent the result of individual action, and 50 percent due to the SDoH.)
The WHO (2010) also has a Commission on the Social Determinants of Health (CSDH) with a conceptual framework:
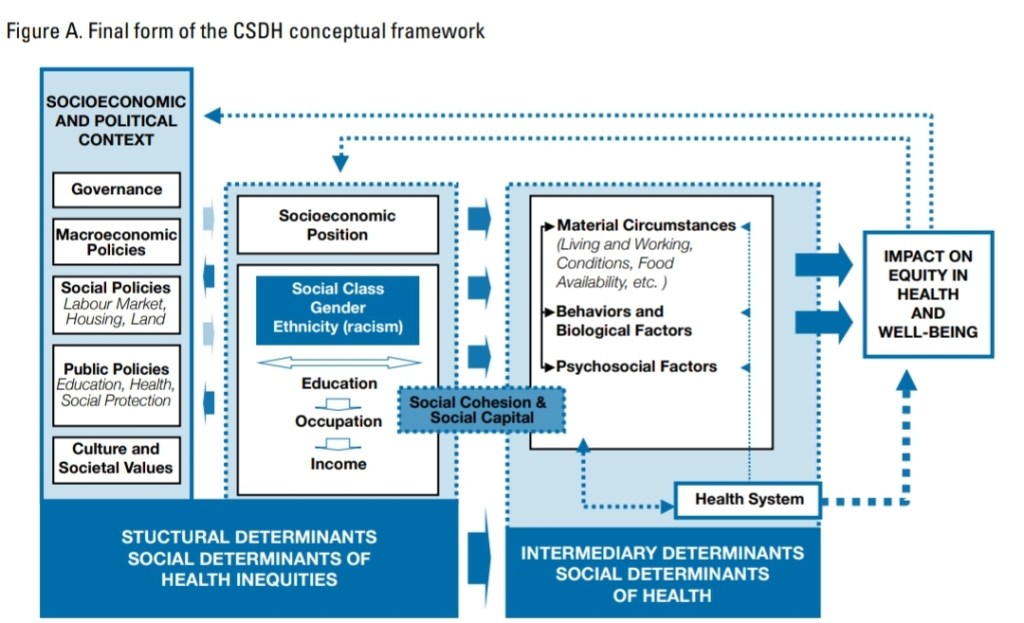
This is staggering. For if the social determinants of health are causal for health outcomes, then it comes down to how society is structured along with how we treat certain people and groups. This also could come down to environmental racism—which is the disproportionate exposure of minority groups to environmental hazards. One pertinent example is lead in paint uses in houses in the 80s, where groups actually used blacks as a kind of experiment in seeing the effects of lead. The issue was further exacerbated by Big Lead in trying to argue that the families had a “history of low intelligence” and that it couldn’t be proven that lead had the damaging effects on the children. Not only does environmental racism cause negative health effects, but so does individual racism which is known to have a negative effect on black women. (Racism and stereotypes which lead to self-fulfilling prophecies also cause the black-white crime gap.)
So empirical research—grounded in data and evidence—can help us in understanding whether a given inequality is an inequity. Certain disparities could reflect historical disadvantages which then perpetuate cycles of disadvantage which then reinforce existing power structures and further continue to marginalize certain communities.
The argument
I have constructed an argument that shows what I am talking about:
UO: Unequal outcome
I: Inequality
A: Avoidable
F: Unfair
J: Unjust
E: Empirical investigation
I involves A, F, J. E may reveal instances where I leads to UO and I is associated with F, J, and A.
Premise 1: E -> (A^F^J)
Premise 2: I -> (A^F^J)
Conclusion: E -> ((I -> UO) ^ (A^F^J))
(P1) If empirical investigations (E) reveal instances where avoidable factors (A), unfairness (F) and injustice (J) are present, and (P2) if inequality (I) leads to conditions involving avoidability (A), unfairness (F) and injustice (J), then (Conclusion) empirical investigations (E) could reveal instances where inequality leads to unequal outcomes (UO), and whether or not they are avoidable (A), unfair (F) or unjust (J). Effectively, since inequities are a kind of inequality, then this can identify inequities where they are, and then we can work to fix them.
For instance, birth weight has decreased recently and the effect is more pronounced for black women (Catov et al, 2016) and racism could as well be a culprit (Collins Jr., et al, 2004). There is also evidence that structural racism in the workplace can and has attributed to this (Chantarat et al, 2022). It’s quite clear that racism can explain birth outcome disparities (Dominguez et al, 2010; Alhusen et al, 2016; Dreyer, 2021). Not only does racism contribute to adverse birth outcomes but so too do factors related to environmental racism (Burris and Hacker, 2018). This also has a historical precedent: slavery (Jasienska, 2009). Hereditarians may try to argue that (as always) this difference has a genetic basis. But we know that African women born in Africa are heavier than African American women; black women born in Africa have children with higher mean birth weights than African American women (David and Collins, 1997). Cabral et al (1991) also found the same—non-American black women birthed children that weighed 135 more grams than American black women. Thus, the difference isn’t genetic in nature—it is environmentally caused and it partly stems from slavery. Clearly this discussion shows that my argument has a real-world basis.
How do we reverse these inequities?
Clearly, racism has societal consequences not only for crime and mental illness, but also low birth weight in black American women. The difference can’t be genetic in nature, so it’s obviously environmental/social in nature due to racism, environmental racism. So how can we alleviate this? There are a few ways.
We can improve access to pre-natal care. By ensuring equitable access to pre-natal care, and by expanding Medicaid coverage, we can the begin to address the issue of low black birth weight. We know that when black newborns are cared for by black doctors, they have a better survival rate (Greenwood et al, 2020). We also have an RCT showing that black doctors could reduce the black-white cardiovascular mortality rate by 19 percent (Alsan, Garrick, and Grasiani, 2019). We also know that a higher percentage of black doctors leads to lower mortality rate and better life expectancy (Peek, 2023; Snyder et al, 2023). This isn’t a new finding—we’ve known this since the 90s (Komaramy et al, 1996; Saha et al, 1999). We also know that people who have same-race doctors are more likely to accept much-needed preventative care (LaVeist, Nuru-Jeter, and Jones, 2003). This could then lead to less systemic bias in healthcare, since we know that some of the difference is systemic in nature (Reschovsky and O’Malley 2008: 229, 230). We also know that bias, stereotyping, and prejudice also play a part (Smedley et al, 2003). Such stereotypes are also sometimes unconscious (Williams and Rucker, 2000). The medical system contributes to said disparities (Bird and Clinton, 2001). Blacks who perceived more racism in healthcare felt more comfortable with a black doctor (Chen et al, 2005)—minorities also trust the healthcare system less than whites (Boulware et al, 2003). Lastly, black and white doctors agree that race is a medically relevant data point, but they don’t agree on why (Bonham et al, 2009).
We know that systemic and structural racism exists and that it impacts health outcomes (Braveman et al, 2022). Some may say that systemic and structural racism don’t exist, but this claim is clearly false. They are “are forms of racism that are pervasively and deeply embedded in and throughout systems, laws, written or unwritten policies, entrenched practices, and established beliefs and attitudes that produce, condone, and perpetuate widespread unfair treatment of people of color. They reflect both ongoing and historical injustices” (Braveman et al, 2022). Perhaps the most important way that systemic racism can harm health is through placing people at an economic disadvantage and stress. Environmental racism then compounds this, and then unfair treatment then leads to higher levels of stress which then leads to negative health outcomes.
Lastly a key issue here is the prevalence of racism. We know that it has a slew of negative health effects and that it affects the incidence of the black-white crime gap. But what can be done to alleviate racist attitudes?
Since many racist ideas have a genetically essentialist tilt, then we can use education to ameliorate racist attitudes (Donovan, 2022). We also know that racial essentialist attitudes are related to the belief that evolution has an intentional tilt and that it’s negatively correlated with biology grades (Donovan, 2015). Much of Donovan’s work shows that education can ameliorate racist attitudes which are due to genetic essentialism. We also know that such essentialist thinking is related to misconceptions about heredity and evolution and is correlated with low grades at the end of the semester in beginner biology course (Donovan, 2016). Thus, by providing accurate and understandable education on race, genetics, and evolution, people may be less likely to hold racial essentialist attitudes and more likely to reject racist ideologies. So there are actionable things we can do to combat racism which leads to crime and negative health outcomes for minority groups.
Conclusion
The SDoH play a pivotal role in shaping the health outcomes while perpetuating health inequities. We can, through empirical investigations, ascertain when an inequality is avoidable, unfair and unjust (meaning, when it is an inequity). We can then understand how historical injustices like racism impact marginalized communities which then contribute to negative health outcomes like low birth weight of black American babies. We know that it’s not a genetic difference since non-American black women have children with higher mean birth weights than black American women, and this suggests thar historical injustices and racism are a cause (as Jasienska argues). Further, studies show that when black patients have black doctors, they report better care and have higher life expectancies. Research has also shown that education can play a role in ameliorating genetic essentialist and racist attitudes which then, as I’ve shown, lead to negative health outcomes. The argument I’ve made here has a real-world basis in the case of low birth weight of black American babies.
In sum, committing to social and racial justice can help to change these inequities, and for that, we will have a better and more inclusive society where people’s negative health outcomes aren’t caused by social goings-on. To achieve racial health equity, we must address the avoidable, unfair and unjust factors that contribute to these inequities.
The “Great Replacement Theory”
2550 words
Introduction
The “Great Replacement Theory” (GRT hereafter) is a white nationalist conspiracy theory (conceptualized by French philosopher Renaud Camus) where there is an intentional effort by some shadowy group (i.e., Jews and global elites) to bring mass amounts of immigrants with high TFRs to countries with whites where whites have low TFRs in order to displace and replace whites in those countries (Beirich, 2021). Vague statements have been made about their “IQs” in that they would be easier to “control” and that they would then intermix with whites to further decrease the IQ of the nation and then be more controllable, all the while the main goal of the GRT—the destruction of the white race—would come to fruition. Here, I will go through the logic of what I think the two premises of the GRT are, and then I will show how the two premises (which I hold to obviously be true) don’t guarantee the conclusion that the GRT is true and that there is an intentional demographic replacement. I will discuss precursors of this that are or almost are 100 years old. I will then discuss what “theory” and “conspiracy theory” means and how, by definition, the GRT is both a theory (an attempted explanation of observed facts) and a conspiracy theory (suggesting a secret plan for the destruction and replacement of the white race).
The genesis of the GRT
The idea of the GRT is older than what spurred it’s discussion in the new millennium, but it can be traced in its modern usage to French political commentator Renaud Camus in his book Le Grand Remplacement.
But one of the earliest iterations of the GRT is the so-called “Kalergi plan.” Kalergi was also one of the founders of the pan-European union (Wiedemer, 1993). Kalergi, in his 1925 book Practical Idealism, wrote that “The man of the future will be of mixed race. Today’s races and classes will gradually disappear owing to the vanishing of space, time, and prejudice. The Eurasian-Negroid race of the future will replace the diversity of peoples with a diversity of individuals.” Which is similar to what Grant (1922: 110) wrote in The Passing of the Great Race:
All historians are familiar with the phenomenon of a rise and decline in civilization such as has oc- curred time and again in the history of the world but we have here in the disappearance of the Cro-Magnon race the earliest example of the replacement of a very superior race by an inferior one. There is great danger of a similar replacement of a higher by a lower type here in America unless the native American uses his superior intelligence to protect himself and his children from competition with intrusive peoples drained from the lowest races of eastern Europe and western Asia.
The idea of a great replacement is obviously much older than what spurred it on today. Movement was much tougher back then as the technology for mass migrations was just beginning to become more mainstream (think of the mass migrations from the 1860s up until the 1930s in America from European groups). Even the migration of other whites from Europe was used as a kind of “replacement” of protestant Anglo-Saxon ways of life. Nonetheless, these ideas of a great replacement are not new, and these two men (one of which—Kalergi—wasn’t using the quote in a nefarious way, contra the white nationalists who use this quote as evidence of the GRT and the plan for it in the modern day) are used as evidence that it is occurring.
Kalergi envisioned a positive blending of the races, whereas Grant expressed concerns of replacement by so-called “inferior” groups replacing so-called “superior” groups. Grant—in trying to argue that Cro-Magnon man was the superior race, replaced by the inferior one—expressed worry of intentional demographic replacement, which is the basis of the GRT today and what the GRT essentially reduces to. The combination of these opposing perspectives of the mixing of races (the positive one from Kalergi and the negative one from Grant) show that the idea of a great replacement is much older than Camus’ worry in his book. (And, as I will argue, the fact that the 2 below premises are true doesn’t guarantee the conclusion of the GRT.)
The concept of the GRT
The GRT has two premises:
(1) Whites have fewer children below TFR
(2) Immigrants have more children above TFR
Which then should get us to:
(C) Therefore, the GRT is true.
But how does (C) follow from (1) and (2)? The GRT suggests not only a demographic shift in which the majority (whites) are replaced and displaced by minorities (in this case mostly “Hispanics” in America), but that this is intentional—that is, it is one man or group’s intention for this to occur. The two premises above refer to factual, verifiable instances: Whites have fewer children; immigrants coming into America have more children. BUT just because those two premises are true, this does NOT mean that the conclusion—GRT is true—follows from the two premises. The two premises focus on the fertility rates of two groups (American whites and immigrants to America), but acceptance of both of those premises does not mean that there is an act of intentional displacement occurring. We can allow the truth of both premises, but that doesn’t lead to the truth of the GRT. Because that change is intentionally driven by some super secret, shadowy and sinister group (the Jews or some other kind of amalgamation of elites who want easy “slave labor”).
The GRT was even endorsed by the Buffalo shooter who heniously shot and killed people in a Tops supermarket. He was driven by claims of the GRT. (The US Congress condemned the GRT as a “White supremacist conspiracy theory“, and I will show how it is a theory and even a conspiracy theory below.) The shooter even plagiarized the “rationale section” of his manifesto (Peterka-Benton and Benton, 2023). This shows that such conspiracy theories like the GRT can indeed lead to radicalization of people.
Even ex-presidential hopeful Vivek Ramaswamy made reference to the GRT, stating that “great replacement theory is not some grand right-wing conspiracy theory, but a basic statement of the Democratic Party’s platform.” Even former Fox News political commentator Tucker Carlson has espoused these beliefs on his former show on Fox News. The belief in such conspiratorial thinking can quite obviously—as seen with the Buffalo shooter—have devestating negative consequences (Adam-Troian et al, 2023). Thus, these views have hit the mainstream as something that’s “plausible” on the minds of many Americans.
Such thinking obviously can be used for both Europe and America—where the Islamization/Africanization of Europe and the browning of America with “Hispanics” and other groups—where there is a nefarious plot to replace the white population of both locations, and these mostly derive on places like 4chan where they try to “meme” what they want into reality (Aguilar, 2023).
On theories and conspiracy theories
Some may say that the GRT isn’t a theory nor is it even a conspiracy theory—it’s a mere observation. I’ve already allowed that both premises of the argument—whites have fewer children below TFR while immigrants have more children above TFR—is true. But that doesn’t mean that the conclusion follows that the GRT is true. Because, as argued above, it is intentional demographic replacement. Intentional by whom? Well the Jews and other global elites who want a “dumb” slave population that just listens, produces and has more children so as to continue the so-called enslavement of the lower populations.
But, by definition, the GRT is a theory and even a conspiracy theory. The GRT is a theory in virtue of it being an explanation for observed demographic changes and the 2 premises I stated above. It is a conspiracy theory because it suggests a deliberate, intentional plan by the so-called global elite to replace whites with immigrants. Of course labeling something as a conspiracy theory doesn’t imply that it’s inaccurate nor invalid, but I would say that the acceptance of both premises DO NOT guarantee the conclusion that those who push the GRT want it to.
The acceptance of both premises doesn’t mean that the GRT is true. The differential fertility of two groups, where one group (the high fertility group) is migrating into the country of another group (the low fertility group) doesn’t mean that there is some nefarious plot by some group to spur race mixing and the destruction and replacement of one group over another.
As shown above, people may interpret and respond to the GRT in different ways. Some may use it in a way to interpret and understand demographic changes while not committing henious actions, while others—like the Buffalo shooter—may use the information in a negative way and take many innocent lives on the basis of belief in the theory. Extreme interpretations of the GRT can lead to the shaping of beliefs which then contribute to negative actions based on the belief that their group is being replaced (Obaidi et al, 2021). Conspiracy theories also rely on the intent to certain events, of which the proponents of the GRT do.
Some white nationalists who hold to the GRT state that the Jews are behind this for a few reasons—one of which I stated above (that they want dumber people to come in who have higher TFRs to replace the native white population in the country)—and another reason which has even less support (if that’s even possible) which is that the Jews are orchestrating the great migration of non-whites into European countries as revenge and retaliation for Europeans expelling Jews from European countries during the middle ages (109 countries). This is the so-called “white genocide” conspiracy theory. This is the kind of hate that Trump ran with in his presidential run and in his time in office as president of the United States (Wilson, 2018). This can also be seen with the phrase “Jews/You will not replace us!” during the Charlottesville protests of 2017 (Wilson, 2021). “You” in the phrase “You will not replace us!” could refer to Jews, or it could refer to the people that the Jews are having migrate into white countries to replace the white population. Beliefs in such baseless conspiracy theories gave led to mass murder in America, Australia, and Norway (Davis, 2024).
One of the main actors in shaping the view that Jews are planning to replace (that is, genocide) Whites is white nationalist and evolutionary psychologist Kevin MacDonald, more specifically in his book series on the origin of Jewish evolutionary group strategies, with A People that Shall Dwell Alone (1994), Separation and it’s Discontents (1998a), and The Culture of Critique (1998b). It is a main argument in this book series that the Jews have an evolved evolutionary group strategy that has them try to undermine and destroy white societies (see Blutinger, 2021 and also Nathan Cofnas’ responses to MacDonald ‘s theory). MacDonald’s theory of a group evolutionary strategy is nothing more than a just-so story. Such baseless views have been the “rationale” of many mass killings in the 2010s (eg Fekete, 2011; Nilsson, 2022). Basically it’s “white genocide is happening and the Jews are behind it so we need to kill those who the Jews are using to enact their plan and we need to kill Jews.” (Note that this isn’t a call for any kind of violence it’s just a simplified version of what many of these mass killers imply in their writings and motivations for carrying out their henious attacks.) One thing driving these beliefs and that jd the GRT is that of anti-Semitism (Allington, Buarque, and Flores, 2020). Overall, such claims of a GRT or “white genocide” flourish online (Keulennar and Reuters, 2023). In this instance, it is claimed that Jews are using their ethnic genetic interests and nepotism to spur these events.
Conclusion
I have discussed the GRT argument and with it so-called “white genocide” (since the two are linked). The 2 premises of the GRT are tru—that American whites have low TFR and those who are emigrating have high TFR—but but that the premises are true doesn’t guarantee the conclusion that there is some great replacement occurring, since it reduces to a kind of intentional demographic replacement by some group (say, the Jews and other elites in society who want cheap, dumb, easily controllable labor who have more children). The GRT is happening, it is claimed, since the Jews want revenge on whites for kicking them out of so many countries. That is, the GRT is an intentional demographic replacement. Those who push the GRT take the two true premises and then incorrectly conclude that there is some kind of plan to eradicate whites through both the mixing of races and bringing in groups of people who have more children than whites do.
I have scrutinized what I take to be the main argument of GRT proponents and have shown that the conclusion they want doesn’t logically follow. Inherent in this is a hasty generalization fallacy and fallacy of composition (in the argument as I have formalized it). This shows the disconnect between both premises and the desired conclusion. Further, the classification of the GRT as a conspiracy theory comes from the attribution of intention to eliminate and eradicate white through the mass migration of non-white immigrant groups who have more children than whites along with racial mixing.
The Buffalo shooting in a Tops supermarket in 2022 shows the impact of these beliefs on people who want there to be some kind of plan or theory for the GRT. Even mainstream pundits and a political candidate have pushed the GRT to a wider audience. And as can be seen, belief in such a false theory can, does, and has led to the harm and murder of innocent people.
Lastly, I showed how the GRT is a theory (since it is an attempt at an explanation for an observed trend) and a conspiracy theory (since the GRT holds that there is a secret plan, with people behind the scenes in the shadows orchestrating the events of the GRT). Such a shift in demographics need not be the result of some conspiracy theory with the intention to wipe out one race of people. Of course some may use the GRT to try to understand how and why the demographics are changing in the West, but it is mostly used as a way to pin blame on why whites aren’t having more children and why mass immigration is occurring.
All in all, my goal here was to show that the GRT has true premises but the conclusion doesn’t follow, and that it is indeed a theory and a conspiracy theory. I have also shown how such beliefs can and have led to despicable actions. Clearly the impact of beliefs on society can have negative effects. But by rationally thinking about and analyzing such claims, we can show that not only are they baseless, but that it’s not merely an observation of observed trends. Evidence and logic should be valued here, while we reject unwanted, centuries-old stereotypes of the purported plan of racial domination of certain groups.
Race, Racism, Stereotypes, and Crime: An Argument for Why Racism is Morally Wrong
2300 words
Introduction
(1) Crime is bad. (2) Racism causes crime. (C) Thus, racism is morally wrong. (1) is self-evident based on people not wanting to be harmed. (2) is known upon empirical examination, like the TAAO and it’s successful novel predictions. (C) then logically follows. In this article, I will give the argument in formal notation and show its validity while defending the premises and then show how the conclusion follows from the premises. I will then discuss two possible counter arguments and then show how they would fail. I will show that you can derive normative conclusions from ethical and factual statements (which then bypasses the naturalistic fallacy), and then I will give the general argument I am giving here. I will discuss other reasons why racism is bad (since it leads to negative physiological and mental health outcomes), and then conclude that the argument is valid and sound and I will discuss how stereotypes and self-fulfilling prophecies also contribute to black crime.
Defending the argument
This argument is obviously valid and I will show how.
B stands for “crime is bad”, C stands for “racism causes crime”, D stands for racism is objectively incorrect, so from B and C we derive D (if C causes B and B is bad, then D is morally wrong). So the argument is “(B ^ C) -> D”. B and C lead to D, proving validity.
Saying “crime is bad” is an ethical judgement. The term “bad” is used as a moral or ethical judgment. “Bad” implies a negative ethical assessment which suggests that engaging in criminal actions is morally undesirable or ethically wrong. The premise asserts a moral viewpoint, claiming that actions that cause harm—including crime—are inherently bad. It implies a normative stance which implies that criminal behavior is wrong or morally undesirable. So it aligns with the idea that causing harm, violating laws or infringing upon others is morally undesirable.
When it comes to the premise “racism causes crime”, this needs to be centered on the theory of African American offending (TAAO). It’s been established that blacks experiencing racism is causal for crime. So the premise implies that racism is a factor in or contributes to criminal behavior amongst blacks who experience racism. Discriminatory practices based on race (racism) could lead to social inequalities, marginalization and frustration which would then contribute to criminal behavior among the affected person. This could also highlight systemic issues where racist policies or structures create an environment conducive to crime. And on the individual level, experiences of racism could influence certain individuals to engage in criminal activity as a response or coping mechanism (Unnever, 2014; Unnever, Cullen, and Barnes, 2016). Perceived racial discrimination “indirectly predicted arrest, and directly predicted both illegal behavior and jail” (Gibbons et al, 2021). Racists propose that what causes the gap is a slew of psychological traits, genetic factors, and physiological variables, but even in the 1960s, criminologists and geneticists rejected the genetic hypothesis of crime (Wolfgang,1964). However we do know there is a protective effect when parents prepare their children for bias (Burt, Simons, and Gibbons, 2013). Even the role of institutions exacerbates the issue (Hetey and Eberhardt, 2014). And in my article on the Unnever-Gabbidon theory of African American offending, I wrote about one of the predictions that follows from the theory which was borne out when it was tested.
So it’s quite obvious that the premise “racism causes crime” has empirical support.
So if B and C are true then D follows. The logical connection between B and C leads to the conclusion that “racism is morally wrong”, expressed by (B ^ C) -> D. Now I can express this argument using modus ponens.
(1) If (B ^ C) then D. (Expressed as (B ^ C) -> D).
(2) (B ^ C) is true.
(3) Thus, D is true.
When it comes to the argument as a whole it can be generalized to harm is bad and racism causes harm so racism is bad.
Furthermore, I can generalize the argument further and state that not only that crime is bad, but that racism leads to psychological harm and harm is bad, so racism is morally wrong. We know that racism can lead to “weathering” (Geronimus et al, 2006, 2011; Simons, 2021) and increased allostatic load (Barr 2014: 71-72). So racism leads to a slew of unwanted physiological issues (of which microaggressions are a species of; Williams, 2021).
Racism leads to negative physiological and mental health outcomes (P), and negative physiological and mental health outcomes are undesirable (Q), so racism is morally objectionable (R). So the factual statement (P) establishes a link between negative health outcomes, providing evidence that racism leads to these negative health outcomes. The ethical statement (Q) asserts that negative health outcomes are morally undesirable which aligns with a common ethical principle that causing harm is morally objectionable. Then the logical connection (Q ^ P) combines the factual observation of harm caused by racism with the ethical judgment that harm is morally undesirable. Then the normative conclusion (R) follows, which asserts that racial is morally objectionable since it leads to negative health outcomes. So this argument is (Q ^ P) -> R.
Racism can lead to stereotyping of certain groups as more prone to criminal behavior, and this stereotype can be internalized and perpetuated which would then contribute to biased law enforcement and along with it unjust profiling. It can also lead to systemic inequalities like in education, employment and housing which are then linked to higher crime rates (in this instance, racism and stereotyping causes the black-white crime gap, as predicted by Unnever and Gabbidon, 2011 and then verified by numerous authors). Further, as I’ve shown, racism can negatively affect mental health leading to stress, anxiety and trauma and people facing these challenges would be more vulnerable to engage in criminal acts.
Stereotypes and self-fulfilling prophecies
In his book Concepts and Theories of Human Development, Lerner (2018: 298) discusses how stereotyping and self-fulfilling prophecies would arise from said stereotyping. He says that people, based on their skin color, are placed into an unfavorable category. Then negative behaviors were attributed to the group. Then these behaviors were associated with different experience in comparison to other skin color groups. These different behaviors then delimit the range of possible behaviors that could develop. So the group was forced into a limited number of possible behaviors, the same behaviors they were stereotyped to have. So the group finally develops the behavior due to being “channeled” (to use Lerner’s word) which is then “the end result of the physically cued social stereotype was a self-fulfilling prophecy” (Lerner, 2018: 298).
From the analysis of the example I provided and, as well, from empirical literature in support of it (e.g., Spencer, 2006; Spencer et al., 2015), a strong argument can be made that the people of color in the United States have perhaps experienced the most unfortunate effects of this most indirect type of hereditary contribution to behavior–social stereotypes. Thus, it may be that African Americans for many years have been involved in an educational and intellectual self-fulfilling prophecy in the United States. (Lerner, 2018: 299)
This is an argument about how social stereotypes can spur behavioral development, and it has empirical support. Lerner’s claim that perception influences behavior is backed by Spencer, Swanson and Harpalani’s (2015) article on the development of the self and Spencer, Dupree, and Hartman’s (1997) phenomenological variant of ecological systems theory (PVEST). (Also see Cunningham et al, 2023). Spencer, Swanson and Harpalani (2015: 764) write:
Whether it is with images of the super-athlete, criminal, gangster, or hypersexed male, it seems that most of society’s views of African Americans are defined by these stereotypes. The Black male has, in one way or another, captured the imagination of the media to such a wide extent that media representations create his image far more than reality does. Most of the images of the Black male denote physical prowess or aggression and downplay other characteristics. For example, stereotypes of Black athletic prowess can be used to promote the notion that Blacks are unintelligent (Harpalani, 2005). These societal stereotypes, in conjunction with numerous social, political, and economic forces, interact to place African American males at extreme risk for adverse outcomes and behaviors.
A -> B—So stereotypes can lead to self-fulfilling prophecies (if there are stereotypes, then they can result in self-fulfilling prophecies). B -> C—Self-fulfilling prophecies can increase the chance of crime for blacks (if there are self-fulfilling prophecies, then they can increase the chance of crime for blacks. So A -> C—Stereotypes can increase the chance of crime for blacks (if there are stereotypes, then they can increase the chance of crime for blacks). Going back to the empirical studies on the TAAO, we know that racism and stereotypes cause the black-white crime gap (Unnever, 2014; Unnever, Cullen, and Barnes, 2016; Herda, 2016, 2018; Scott and Seal, 2019), and so the argument by Spencer et al and Lerner is yet more evidence that racism and stereotypes lead to self-fulfilling prophecies which then cause black crime. Behavior can quite clearly be shaped by stereotypes and self-fulfilling prophecies.
Responses to possible counters
I think there are 3 ways that one could try to refute the argument—(1) Argue that B is false, (2) argue that C is false, or (3) argue that the argument commits the is-ought fallacy.
(1) Counter premise: B’: “Not all crimes are morally bad, some may be morally justifiable or necessary in certain contexts. So if not all crimes are morally bad, then the conclusion that racism is morally wrong based on the premises (B ^ C) isn’t universally valid.”
Premise B reflects a broad ethical judgment which is based on social norms that generally view actions that cause harm morally undesirable. My argument is based on consequences—that racism causes crime. The legal systems of numerous societies categorize certain actions as crimes since they are deemed morally reprehensible and harmful to individuals and communities. Thus, there is a broad moral stance against actions that cause harm which is reflected in the societal normative stance against actions which cause harm.
(2) Counter premise: C’: “Racism does not necessarily cause crime. Since racism does not necessarily cause crime, then the conclusion that racism is objectively wrong isn’t valid.”
Premise C states that racism causes crime. When I say that, it doesn’t mean that every instance of racism leads to an instance of crime. Numerous social factors contribute to criminal actions, but there is a relationship between racial discrimination (racism) and crime:
Experiencing racial discrimination increases the likelihood of black Americans engaging in criminal actions. How does this follow from the theory? TAAO posits that racial discrimination can lead to feelings of frustration and marginalization, and to cope with these stressors, some individuals may resort to commuting criminal acts as a way to exert power or control in response to their experiences of racial discrimination. (Unnever, 2014; Unnever, Cullen, and Barnes, 2016; Herda, 2016, 2018; Scott and Seal, 2019)
(3) “The argument commits the naturalistic fallacy by inferring an “ought” from an “is.” It appears to derive a normative conclusion from factual and ethical statements. So the transition from descriptive premises to moral judgments lacks a clear ethical justification which violates the naturalistic fallacy.” So this possible counter contends that normative statement B and the ethical statement C isn’t enough to justify the normative conclusion D. Therefore it questions whether the argument has good justification for an ethical transition to the conclusion D.”
I can simply show this. Observe X causing Y (C). Y is morally undesirable (B). Y is morally undesirable and X causes Y (B ^ C). So X is morally objectionable (D). So C begins with an empirical finding. B then is the ethical premise. The logical connection is then established with B ^ C (which can be reduced to “Harm is morally objectionable and racism causes harm”). This then allows me to infer the normative conclusion—D—allowing me to bypass the charge of committing the naturalistic fallacy. Thus, the ethical principle that harm is morally undesirable and that racism causes harm allows me to derive the conclusion that racism is objectively wrong. So factual statements can be combined with ethical statements to derive ethical conclusions, bypassing the naturalistic fallacy.
Conclusion
This discussion centered on my argument (B ^ C) -> D. The argument was:
(P1) Crime is bad (whatever causes harm is bad). (B)
(P2) Racism causes crime. (C)
(C) Racism is morally wrong. (D)
I defended the truth of both premises, and then I answered two possible objections, both rejecting B and C. I then defended my argument against the charge of it committing the naturalistic fallacy by stating that ethical statements can be combined with factual statements to derive normative conclusions. Addressing possible counters (C’ and B’), I argued that there is evidence that racism leads to crime (and other negative health outcomes, generalized as “harm”) in black Americans, and that harm is generally seen as bad, so it then follows that C’ and B’ fail. Spencer’s and Lerner’s arguments, furthermore, show how stereotypes can spur behavioral development, meaning that social stereotypes increase the chance of adverse behavior—meaning crime. It is quite obvious that the TAAO has strong empirical support, and so since crime is bad and racism causes crime then racism is morally wrong. So to decrease the rate of black crime we—as a society—need to change our negative attitudes toward certain groups of people.
Thus, my argument builds a logical connection between harm being bad, racism causing harm and moral undesirability. In addressing potential objections and clarifying the ethical framework I ren, So the general argument is: Harm is bad, racism causes harm, so racism is morally wrong.
Jensen’s Default Hypothesis is False: A Theory of Knowledge Acquisition
2000 words
Introduction
Jensen’s default hypothesis proposes that individual and group differences in IQ are primarily explained genetic factors. But Fagan and Holland (2002) question this hypothesis. For if differences in experience lead to differences in knowledge, and differences in knowledge lead to differences in IQ scores, then Jensen’s assumption that blacks and whites have the same opportunity to learn the content is questionable, and I’d think it false. It is obvious that there are differences in opportunity to acquire knowledge which would then lead to differences in IQ scores. I will argue that Jensen’s default hypothesis is false due to this very fact.
In fact, there is no good reason to accept Jensen’s default hypothesis and the assumptions that come with it. Of course different cultural groups are exposed to different kinds of knowledge, so this—and not genes—would explain why different groups score differently on IQ tests (tests of knowledge, even so-called culture-fair tests are biased; Richardson, 2002). I will argue that we need to reject Jensen’s default hypothesis on these grounds, because it is clear that groups aren’t exposed to the same kinds of knowledge, and so, Jensen’s assumption is false.
Jensen’s default hypothesis is false due to the nature of knowledge acquisition
Jensen (1998: 444) (cf Rushton and Jensen, 2005: 335) claimed that what he called the “default hypothesis” should be the null that needs to be disproved. He also claimed that individual and group differences are “composed of the same stuff“, in that they are “controlled by differences in allele frequencies” and that these differences in allele frequencies also exist for all “heritable” characters, and that we would find such differences within populations too. So if the default hypothesis is true, then it would suggest that differences in IQ between blacks and whites are primarily attributed to the same genetic and environmental influences that account for individual differences within each group. So this implies that genetic and environmental variances that contribute to IQ are therefore the same for blacks and whites, which supposedly supports the idea that group differences are a reflection of individual differences within each group.
But if the default hypothesis were false, then it would challenge the assumption that genetic and environmental influences in IQ between blacks and whites are proportionally the same as seen in each group. Thus, this allows us to talk about other causes of variance in IQ between blacks and whites—factors other than what is accounted for by the default hypothesis—like socioeconomic, cultural, and historical influences that play a more substantial role in explaining IQ differences between blacks and whites.
Fagan and Holland (2002) explain their study:
In the present study, we ensured that Blacks and Whites were given equal opportunity to learn the meanings of relatively novel words and we conducted tests to determine how much knowledge had been acquired. If, as Jensen suggests, the differences in IQ between Blacks and Whites are due to differences in intellectual ability per se, then knowledge for word meanings learned under exactly the same conditions should differ between Blacks and Whites. In contrast to Jensen, we assume that an IQ score depends on information provided to the learner as well as on intellectual ability. Thus, if differences in IQ between Blacks and Whites are due to unequal opportunity for exposure to information, rather than to differences in intellectual ability, no differences in knowledge should obtain between Blacks and Whites given equal opportunity to learn new information. Moreover, if equal training produces equal knowledge across racial groups, than the search for racial differences in IQ should not be aimed at the genetic bases of IQ but at differences in the information to which people from different racial groups have been exposed.
There are reasons to think that Jensen’s default hypothesis is false. For instance, since IQ tests are culture-bound—that is, culturally biased—then they are biased against a group so they therefore are biased for a group. Thus, this introduces a confounding factor which challenges the assumption of equal genetic and environmental influences between blacks and whites. And since we know that cultural differences in the acquisition of information and knowledge vary by race, then what explains the black-white IQ gap is exposure to information (Fagan and Holland, 2002, 2007).
The Default Hypothesis of Jensen (1998) assumes that differences in IQ between races are the result of the same environmental and genetic factors, in the same ratio, that underlie individual differences in intelligence test performance among the members of each racial group. If Jensen is correct, higher and lower IQ individuals within each racial group in the present series of experiments should differ in the same manner as had the African-Americans and the Whites. That is, in our initial experiment, individuals within a racial group who differed in word knowledge should not differ in recognition memory. In the second, third, and fourth experiments individuals within a racial group who differed in knowledge based on specific information should not differ in knowledge based on general information. The present results are not consistent with the default hypothesis.(Fagan and Holland, 2007: 326)
Historical and systematic inequalities could also lead to differences in knowledge acquisition. The existence of cultural biases in educational systems and materials can create disparities in knowledge acquisition. Thus, if IQ tests—which reflect this bias—are culture-bound, it also questions the assumption that the same genetic and environmental factors account for IQ differences between blacks and whites. The default hypothesis assumes that genetic and environmental influences are essentially the same for all groups. But SES/class differences significantly affect knowledge acquisition, so if challenges the default hypothesis.
For years I have been saying, what if all humans have the same potential but it just crystallizes differently due to differences in knowledge acquisition/exposure and motivation? There is a new study that shows that although some children appeared to learn faster than others, they merely had a head start in learning. So it seems that students have the same ability to learn and that so-called “high achievers” had a head start in learning (Koedinger et al, 2023). They found that students vary significantly in their initial knowledge. So although the students had different starting points (which showed the illusion of “natural” talents), they had more of a knowledge base but all of the students had a similar rate of learning. They also state that “Recent research providing human tutoring to increase student motivation to engage in difficult deliberate practice opportunities suggests promise in reducing achievement gaps by reducing opportunity gaps (63, 64).”
So we know that different experiences lead to differences in knowledge (it’s type and content), and we also know that racial groups for example have different experiences, of course, in virtue of their being different social groups. So these different experiences lead to differences in knowledge which are then reflected in the group IQ score. This, then, leads to one raising questions about the truth of Jensen’s default hypothesis described above. Thus, if individuals from different racial groups have unequal opportunities to be exposed to information, then Jensen’s default hypothesis is questionable (and I’d say it’s false).
Intelligence/knowledge crystalization is a dynamic process shaped by extensive practice and consistent learning opportunities. So the journey towards expertise involves iterative refinement with each practice opportunity contribute to the crystallization of knowledge. So if intelligence/knowledge crystallizes through extensive practice, and if students don’t show substantial differences in their rates of learning, then it follows that the crystalization of intelligence/knowledge is more reliant on the frequency and quality of learning opportunities than on inherent differences in individual learning rates. It’s clear that my position enjoys some substantial support. “It’s completely possible that we all have the same potential but it crystallizes differently based on motivation and experience.” The Fagan and Holland papers show exactly that in the context of the black-white IQ gap, showing that Jensen’s default hypothesis is false.
I recently proposed a non-IQ-ist definition of intelligence where I said:
So a comprehensive definition of intelligence in my view—informed by Richardson and Vygotsky—is that of a socially embedded cognitive capacity—characterized by intentionality—that encompasses diverse abilities and is continually shaped by an individual’s cultural and social interactions.
So I think that IQ is the same way. It is obvious that IQ tests are culture-bound and tests of a certain kind of knowledge (middle-class knowledge). So we need to understand how social and cultural factors shape opportunities for exposure to information. And per my definition, the idea that intelligence is socially embedded aligns with the notion that varying sociocultural contexts do influence the development of knowledge and cognitive abilities. We also know that summer vacation increases educational inequality, and that IQ decreases during the summer months. This is due to the nature of IQ and achievement tests—they’re different versions of the same test. So higher class children will return to school with an advantage over lower class children. This is yet more evidence in how knowledge exposure and acquisition can affect test scores and motivation, and how such differences crystallize, even though we all have the same potential (for learning ability).
Conclusion
So intelligence is a dynamic cognitive capacity characterized by intentionality, cultural context and social interactions. It isn’t a fixed trait as IQ-ists would like you to believe but it evolves over time due to the types of knowledge one is exposed to. Knowledge acquisition occurs through repeated exposure to information and intentional learning. This, then, challenges Jensen’s default hypothesis which attributes the black-white IQ gap primarily to genetics.Since diverse experiences lead to varied knowledge, and there is a certain type of knowledge in IQ tests, individuals with a broad range of life experiences varying performance on these tests which then reflect the types of knowledge one is exposed to during the course of their lives. So knowing what we know about blacks and whites being different cultural groups, and what we know about different cultures having different knowledge bases, then we can rightly state that disparities in IQ scores between blacks and whites are suggested to be due to environmental factors.
Unequal exposure to information creates divergent knowledge bases which then influence the score on the test of knowledge (IQ test). And since we now know that despite initial differences in initial performance that students have a surprising regularity in learning rates, this suggests that once exposed to information, the rate of knowledge acquisition remains consistent across individuals which then challenges the assumption of innate disparities in learning abilities. So the sociocultural context becomes pivotal in shaping the kinds of knowledge that people are exposed to. Cultural tools environmental factors and social interactions contribute to diverse cognitive abilities and knowledge domains which then emphasize the contextual nature of not only intelligence but performance in IQ tests. So what this shows is that test scores are reflective of the kinds of experience the testee was exposed to. So disparities in test scores therefore indicate differences in learning opportunities and cultural contexts
So a conclusive rejection of Jensen’s default hypothesis asserts that the black-white IQ gap is due to exposure to different types of knowledge. Thus, what explains disparities in not only blacks and whites but between groups is unequal opportunities to exposure of information—most importantly the type of information found on IQ tests. My sociocultural theory of knowledge acquisition and crystalization offers a compelling counter to hereditarian perspectives, and asserts that diverse experiences and intentionality learning efforts contribute to cognitive development. The claim that all groups or individuals are exposed to similar types of knowledge as Jensen assumes is false. By virtue of being different groups, they are exposed to different knowledge bases. Since this is true, and IQ tests are culture-bound and tests of a certain kind of knowledge, then it follows that what explains group differences in IQ and knowledge would therefore be differences in exposure to information.
Rushton, Race, and Twinning
2500 words
As is the case with the other lines of evidence that intend to provide sociobiological evidence in support of the genetic basis of human behavior and development (relating to homology, heritability, and adaptation), Rushton’s work reduces to no evidence at all. (Lerner, 2018)
Introduction
From 1985 until his death in 2012, J. P. Rushton attempted to marshal all of the data and support he could for a theory called r-K selection theory or Differential K theory (Rushton, 1985). The theory posited that while humans were the most K species of all, some human races were more K than others, so it then followed that some human races were more r than others. Rushton then collated mass amounts of data and wrote what would become his magnum opus, Race, Evolution and Behavior (Rushton, 1997). So in the r/K theory first proposed by MacArthur and Wilson, unstable, unpredictable environments favored an r strategy whereas a stable, predictable environments favored a K strategy. (See here for my response to Rushton’s r/K.)
So knowing this, one of the suite of traits Rushton put on his r/K matrix was twinning rates. Rushton (1997: 6) stated:
the rate of dizygotic twinning, a direct index of egg production, is less than 4 per 1,000 births among Mongoloids, 8 per 1,000 among Caucasoids, and 16 or greater per 1,000 among Negroids.
I won’t contest the claim that the rate in DZ twinning is higher by race—because it’s pretty well-established with recent data that blacks are more likely to have twins than whites (that is, blacks have a slightly higher chance of having twins than whites, who have a slightly higher chance of having twins than Asians) (Santana, Surita, and Cecatti, 2018; Wang, Dongarwar, and Salihu, 2020; Monden, Pison, and Smits, 2021)—I’m merely going to contest the causes of DZ twinning. Because it’s clear that Rushton was presuming this to be a deeply evolutionary trait since a highs rate of twins—in an evolutionary context—would mean that there would be a higher chance for children of a particular family to survive and therefore spread their genes and thusly would, in his eyes, lend credence to his claim that Africans were more r compared to whites who were more r compared to Asians.
But to the best of my knowledge, Rushton didn’t explain why, biologically, blacks would have more twins than whites—he merely said “This race has more twins than this race, so this lends credence to my theory.” That is, he didn’t posit a biological mechanism that would instantiate a higher rate of twinning in blacks compared to whites and Asians and then explain how environmental effects wouldn’t have any say in the rate of twinning between the races. However, I am privy to environmental factors that would lead to higher rates of twinning and I am also privy to the mechanisms of action that allow twinning to occur (eg phytoestrogens, FSH, LH, and IGF). And while these are of course biological factors, I will show that there are considerable effects of environmental interactions like diet on the levels of these hormones which are associated with twinning. I will also explain how these hormones are related to twinning.
While the claim that there is a difference in rate of DZ twinning by race seems to be true, I don’t think it’s a biological trait, nevermind an evolutionary one as Rushton proposed (because even if Rushton’s r/K were valid, “Negroids” would be K and “Mongoloids” would be r, Anderson, 1991). Nonetheless, Rushton’s r/K theory is long-refuted, though he did call attention to some interesting observations (which other researchers never ignored, they just didn’t attempt some grand theory of racial differences).
Follicle stimulating hormone, leutinizing hormone, and insulin-like growth factor
We know that older women are more likely to have twins while younger women are less likely (Oleszczuk et al, 2001), so maternal age is a factor. As women age, a hormone called follicle stimulating hormone (FSH) increases due to a decline in estrogen, and it is one of the earliest signs of female reproductive aging (McTavish et al, 2007), being one of the main biomarkers of ovarian reserve tested on day 3 of the menstrual cycle (Roudebush, Kivens, and Mattke, 2008). It is well established that twinning is different in different geographic locations, that the rate of MZ twins is constant at around 3.5 to 4 per 1,000 births (so what is driving the differences is the birth of DZ twins), and that it increases due to an increase in FSH (Santana, Surita, and Cecatti, 2018). We also know that pre-menopausal women who have given birth to DZ twins have higher levels of FSH on the third day of their menstrual cycle (Lambalk et al, 1998).
So if FSH levels stay too high for too long then multiple eggs are released, which could lead to an increase in DZ twinning. FSH stimulates the maturation and growth of ovarian follicles, each of which contains an immature egg called an oocyte. FSH acts on the ovaries to promote the development of multiple ovarian follicles during pregnancy, a process which is called recruitment. In a normal menstrual cycle, only one follicle is stimulated to release one egg; but when FSH levels are elevated, this results in the development and maturation of more than one follicle which is known as polyovulation. Polyovulation then increases the chance of the release of multiple eggs during ovulation. Thus, if more than one egg is released during a menstrual cycle, and they both are fertilized, it can then lead to the development of DZ twins.
Along with FSH, we also have luetenizing hormone (LH). So FSH and LH act synergistically (Raju et al, 2013). LH, like FSH, isn’t directly responsible for the increase in twinning, but the process that it allows (playing a role in ovulation) is a crucial factor in twinning. So LH is responsible for triggering ovulation, which is the release of a mature egg from the ovarian follicle. (Ovulation occurs typically 24 to 36 hours after LH increases.) In a typical menstrual cycle, only one follicle is stimulated to release one egg, which is triggered by the surge in LH. But if there are multiple mature follies in the ovaries (which could be influenced by FSH), then a surge in LH can lead to the release of more than one egg. So the interaction of LH with other hormone like FSH, along with the presence of multiple mature follicles, can be associated with having a higher chance of having DZ twins. FSH therapies are also used in assisted reproduction (eg Munoz et al, 1995 in mice; Ferraretti et al, 2004; Pang, 2005; Pouwer, Farquhar, and Kremer, 2015; Fatemi et al, 2021).
So when it comes to FSH, we know that malnutrition may play a role in twinning, and also that wild yams—a staple food in Nigeria—increases phytoestrogens which increase FSH in the body of women (Bartolus, et al, 1999). Wild yams have been used to increase estrogen in women’s bodies (due to the phytoestrogens they contain), and it enhances estradiol through the mechanism of binding to estrogen receptor sites (Hywood, 2008). And since Nigeria has the highest rate of twinning in the world (Santana, Surita, and Cecatti, 2018), and their diet is wild yam-heavy (Bartolus, et al, 1999), it seems that this fact would go a long way in explaining why they have higher rates of twinning. Mount Sinai says that “Although it does not seem to act like a hormone in the body, there is a slight risk that wild yam could produce similar effects to estrogen.” It acts as a weak phytoestrogen (Park et al, 2009). (But see Beckham, 2002.) But when phytoestrogens are consumed, they can then bind to estrogen receptors in the body and trigger estrogenic effects which could then lead to the potential stimulation and release of multiple eggs which would increase the chance of DZ twinning.
One study showed that black women, in comparison to white women, had “lower follicular phase LH:FSH ratios” (Reuttman et al, 2002; cf Marsh et al, 2011), while Randolph et al (2004) showed that black women had higher FSH than Asian and white women. So the lower LH:FSH ratio could affect the timing and regulation of ovulation, and a lower LH:FSH level could reduce the chances of premature ovulation and could affect the release of multiple eggs.
Lastly, when it comes to insulin-like growth factor (IGF), this could be influenced by a high protein diet or a high carb diet. Diets high in high glycemic carbs can lead to increase insulin production which would then lead to increased IGF levels. Just like with FSH and LH, increased levels of IGF could also in concert with the other two hormones influence the maturation and release of multiple eggs during a menstrual cycle which would then increase the chance of twinning (Yoshimura, 1998). IGF can also stimulate the growth and development of multiple follicles (Stubbs et al, 2013) and have them mature early if IGF levels are high enough (Mazerbourgh and Monget, 2018). This could then also lead to polyovulation, triggering the release of more than one egg during ovulation. IGF can also influence the sensitivity of the ovaries to hormonal signals, like those from the pituitary gland, which then leads to enhanced ovarian sensitivity to hormones like FSH and LH which then, of course, would act synergistically increasing the rate of dizygotic twinning. (See Mazerbourgh and Monget, 2018 for a review of this.)
So we know that black women have higher levels of IGF-1 and free IGF-1—but lower IGF-2 and IGFBP-3—than white women (Berrigan et al, 2010; Fowke et al, 2011). The higher IGF-1 levels in black women could lead to increase ovarian sensitivity to FSH and LH, and thus enhanced ovarian sensitivity could lead to the promotion and release of multiple eggs during ovulation. The lower IGF-2 levels could contribute to the balance of IGF-1 and IGF-2, which would then further influence the ovarian sensitivity to other hormones. IGFBP-3 is a binding protein which regulated the bioavailability of IGF-1, so lower levels of IGFBP-3 could lead to higher concentrations of free IGF-1, which would then further stimulate the ovarian follicles and could lead to polyovulation, leading to increased twinning. Though there is some evidence that this difference does have a “genetic basis” (Higgins et al, 2005), we know that dietary factors do have an effect on IGF levels (Heald et al, 2003).
Rushton’s misinterpretations
Rushton got a ton wrong, but he was right about some things too (which is to be expected if you’re looking to create some grand theory of racial differences). I’m not too worried about that. But what I AM worried about, is Rushton’s outright refusal to address his most serious critics in the literature, most importantly Anderson (1991) and Graves (2002 a, b). If you check his book (Rushton, 1997: 246-248), his responses are hardly sufficient to address the devestating critiques of his theory. (Note how Rushton never responded to Graves, 2002—ever.) Gorey and Cryns (1995) showed how Rushton cherry-picked what he liked for his theory while stating that “any behavioral differences which do exist between blacks, whites and Asian Americans for example, can be explained in toto by environmental differences which exist between them” while Ember, Ember, and Peregrine (2003) concluded similarly. (Rushton did respond to Gorey and Cryns, but not Ember, Ember, and Peregrine.) Cernovsky and Littman (2019) also showed how Rushton cherry-picked his INTERPOL crime data.
Now that I have set the stage for Rushton’s “great” scholarship, let’s talk about the response he got to his twinning theory.
Allen et al (1992) have a masterful critique of Rushton’s twinning theory. They review twinning stats in other countries across different time periods and come to conclude that “With such a wide overlap between races, and such great variation within races, twinning rate is probably no better than intelligence as an index of genetic status for racial groups.” They also showed that the twinning mechanism didn’t seem to be a relevant factor in survival, until the modern day with the advancement of our medical technologies, that is. So since twinning increases the risk for death in the mother (Steer, 2007; Santana et al, 2018). Rushton also misinterpreted numerous traits associated with twinning:
individual twin proneness and its correlates do not provide Rushton’s desired picture of a many-faceted r- strategy (even if such individual variation could have evolutionary meaning). With the exception of shorter menstrual cycles found in one study, the traits Rushton cites as r-selected in association with twinning are either statistical artifacts of no reproductive value or figments of misinterpretation.
Conclusion
I have discussed a few biological variables that lead to higher rates of twinning and I have cited some research which shows that black women have higher rates of some of the hormones that are related to higher rates of twinning. But I have also shown that it’s not so simple to jump to a genetic conclusion, since these hormones are of course mediated by environmental factors like diet.
Rushton quite clearly takes these twinning rate differences to be “genetic” in nature, but we are in the 2020s now, not the 1980s, and we now know that genes are necessary, but passive players in the formation of phenotypes (Noble, 2011, 2012, 2016; Richardson, 2017, 2021; Baverstock, 2021; McKenna, Gawne, and Nijhout, 2022). These new ways of looking at genes—as passive, not active causes, and as not special from any other developmental resources—shows how the reductionist thinking of Rushton and his contemporaries were straight out false. Nonetheless, while Rushton did get it right that there is a racial difference in twinning, the difference, I think, isn’t a genetic difference and I certainly don’t think they it lends credence to his Differential K theory, since Anderson showed that if we were to accept Rushton’s premises, then African would be K and Asians would be r. So while there also are differences in menarche between blacks and whites, this too also seems to be environmentally driven.
Rushton’s twinning thesis was his “best bet” at attempting to show that his r/K theory was “right” about racial differences. But the numerous devestating critiques of not only Rushton’s thesis on twinning but his r/K Differential K theory itself shows that Rushton was merely a motivated reasoner (David Duke also consulted with Rushton when Duke wrote his book My Awakening, where Duke describes how psychologists led to his “racial awakening”), so “The claim that Rushton was acting only as a scientist is not credible given this context” (Winston, 2020). Even the usefulness of psychometric life history theory has been recently questioned (this derives from Rushton’s Differential K, Sear, 2020).
But it is now generally accepted that Rushton’s r/K and the current psychometric life history theory that rose from the ashes of Rushton’s theory just isn’t a good way to conceptualize how humans live in the numerous biomes we live in.
Racial Differences in Motor Development: A Bio-Cultural View of Motor Development
3050 words
Introduction
Psychologist J. P. Rushton was perhaps most famous for attempting to formulate a grand theory of racial differences. He tried to argue that, on a matrix of different traits, the “hierarchy” was basically Mongoloids > Caucasoids > Negroids. But Rushton’s theory was met with much force, and many authors in many of the different disciplines in which he derived his data to formulate his theory attacked his r/K selection theory also known as Differential K theory (where all humans are K, but some humans are more K than others, so some humans are more r than others). Nonetheless, although his theory has been falsified for many decades, did he get some things right about race? Well, a stopped clock is right twice a day, so it wouldn’t be that outlandish to believe that Rushton got some things right about racial differences, especially when it comes to physical differences. While we can be certain that there are physical differences in groups we term “racial groups” and designate “white”, “black”, “Asian”, “Native American”, and “Pacific Islander” (the five races in American racetalk), this doesn’t lend credence to Rushton’s r/K theory.
In this article, I will discuss Rushton’s claims on motor development between blacks and whites. I will argue that he basically got this right, but it is of no consequence to the overall truth of his grand theory of racial differences. We know that there are physical differences between racial groups. But that there are physical differences between racial groups doesn’t entail that Rushton’s grand theory is true. The only entailment, I think, that can be drawn from that is there is a possibility that physical differences between races could exist between them, but it is a leap to attribute these differences to Rushton’s r/K theory, since it is a falsified theory on logical, empirical and methodological grounds. So I will argue that while Rushton got this right, a stopped clock is right twice a day but this doesn’t mean that his r/K theory is true for human races.
Was Rushton right? Evaluating newer studies on black-white motor development
Imagine three newborns: one white, one black and the third Asian and you observe the first few weeks of their lives. Upon observing the beginnings of their lives, you begin to notice differences in motor development between them. The black infant is more motorically advanced than the white infant who is more motorically advanced than the Asian infant. The black infant begins to master movement, coordination and dexterity showing a remarkable level of motoric dexterity, while the white infant shows less motoric dexterity than the black infant, and the Asian infant still shows lower motoric dexterity than the white infant.
These disparities in motor development are evidence in the early stages of life, so is it genetic? Cultural? Bio-cultural? I will argue that what explains this is a bio-cultural view, and so it will of course eschew reductionism, but of course as infants grow and navigate through their cultural milieu and family lives, this will have a significant effect on their experiences and along with it their motoric development.
Although Rushton got a lot wrong, it seems that he got this issue right—there does seem to be differences in precocity of motor development between the races, and the references he cites below in his 2000 edition of Race, Evolution, and Behavior—although most are ancient compared to today’s standards—hold to scrutiny today, where blacks walk earlier than whites who walk earlier than Asians.
Rushton (2000: 148-149) writes:
Revised forms of Bayley’s Scales of Mental and Motor Development administered in 12 metropolitan areas of the United States to 1,409 representative infants aged 1-15 months showed black babies scored consistently above whites on the Motor Scale (Bayley, 1965). This difference was not limited to any one class of behavior, but included: coordination (arm and hand); muscular strength and tonus (holds head steady, balances head when carried, sits alone steadily, and stands alone); and locomotion (turns from side to back, raises self to sitting, makes stepping movements, walks with help, and walks alone).
Similar results have been found for children up to about age 3 elsewhere in the United States, in Jamaica, and in sub-Saharan Africa (Curti, Marshall, Steggerda, & Henderson, 1935; Knobloch & Pasamanik, 1953; Williams & Scott, 1953; Walters, 1967). In a review critical of the literature Warren (1972) nonetheless reported evidence for African motor precocity in 10 out of 12 studies. For example, Geber (1958:186) had examined 308 children in Uganda and reported an “all-round advance of development over European standards which was greater the younger the child.” Freedman (1974, 1979) found similar results in studies of newboms in Nigeria using the Cambridge Neonatal Scales (Brazelton & Freedman, 1971).
Mongoloid children are motorically delayed relative to Caucasoids. In a series of studies carried out on second- through fifth-generation Chinese-Americans in San Francisco, on third- and fourth-generation Japanese-Americans in Hawaii, and on Navajo Amerindians in New Mexico and Arizona, consistent differences were found between these groups and second- to fourth-generation European-Americans using the Cambridge Neonatal Scales (Freedman, 1974, 1979; Freedman & Freedman, 1969). One measure involved pressing the baby’s nose with a cloth, forcing it to breathe with its mouth. Whereas the average Chinese baby fails to exhibit a coordinated “defense reaction,” most Caucasian babies turn away or swipe at the cloth with the hands, a response reported in Western pediatric textbooks as the normal one.
On other measures including “automatic walk,” “head turning,” and “walking alone,” Mongoloid children are more delayed than Caucasoid children. Mongoloid samples, including the Navajo Amerindians, typically do not walk until 13 months, compared to the Caucasian 12 months and Negro 11 months (Freedman, 1979). In a standardization of the Denver Developmental Screening Test in Japan, Ueda (1978) found slower rates of motoric maturation in Japanese as compared with Caucasoid norms derived from the United States, with tests made from birth to 2 months in coordination and head lifting, from 3 to 5 months in muscular strength and rolling over, at 6 to 13 months in locomotion, and at 15 to 20 months in removing garments.
Regarding newer studies on this matter, there are differences between European and Asian children in the direction that Rushton claimed. Infants from Hong Kong displayed a difference sequence of rolling compared to Canadian children. There does seem to be a disparity in motoric development between Asian and white children (Mayson, Harris, and Bachman, 2007). These authors do cite some of the same studies like the DDST (which is currently outdated) which showed how Asian children were motorically delayed compared to white children. And although they put caution on their findings of their literature review, it’s quite clear that this pattern exists and it is a bio-cultural one. So they conclude their literature review writing “the literature reviewed suggests differences in rate of motor development among children of various ethnic origins, including those of Asian and European descent” and that “Limited support suggests also that certain developmental milestones, such as rolling, may differ between infants of Asian and European origin.” Further, cultural practices in northern China—for example, lying them on their backs on sandbags—stall the onset of walking in babies sitting, crawling, and walking by a few months (Karasik et al, 2011).
This is related to the muscles that are used to roll from a supine to prone position and vice versa. Since some Asian children spend a longer time in apparatuses that aren’t conducive to growing a strong muscular base to be able to roll from the supine to prone position, to crawl and eventually walk, this is the “cultural” in the “bio-cultural” approach I will argue for.
One study on Norwegian children found that half of the children were waking by 13 months (the median) while 25 percent were walking by 12 months and 75 percent were walking by 14 months (Storvold, Aarethun, and Bratberg, 2013). One reason for the delayed response time could be supine sleeping, which was put into effect during the Back to Sleep program to mitigate causes of death from SIDS. Although it obviously saved tens of thousands of infant lives, it came at a cost of slightly stunted motoric development. It also seems that there is poor predictive value for infant milestones such as walking when it comes to health (Jenni et al, 2012).
Black Caribbean, black African and Indian infants were less likely to show delays in gross motor milestones compared to white infants. But Pakistani and Bangladeshi infants were more likely to be delayed in motoric development and communicative gestures, which was partly attributed to socio-cultural factors (Kelly et al, 2006). Kelly et al (2006: 828) also warn against genetic conclusions based on their large findings of difference between white and African and Caribbean infants:
The differences we observed between Black African and Black Caribbean compared with White infants are large and remain unaffected after adjusting for important covariates. This makes it tempting to conclude that the remaining effect must be a consequence of genetic differences. However, such a conclusion would be prematurely drawn. First, we have not included the measurement of genetic factors in our analysis, and, therefore, the presence of such effects cannot be demonstrated. Second, speculating on such effects should only be done alongside recognition that the model we have been able to test contains imperfect measurement.
It has also been observed that black and white children achieved greater mastery of motoric ability (locomotor skills) compared to Asian children but there was no difference by age group (Adeyemi-Walker et al, 2018). It was also found that infants with higher motor development scores had a lower weight weight relative to their length as they grew. So it was found that delayed motor development was associated with higher weight relative to length (Shoaibi et al, 2018). Black infants are also more motorically advanced and this is seen at up to two years of age (Malina, 1988) while black children perform better on tests of motor ability than white children (Okano et al, 2001). Kilbride et al (1970) also found that Baganda infants in Uganda showed better motoric ability than white American children. Campbell and Heddeker (2001) also showed that black infants were more motorically advanced than infants of other races.
It is clear that research like this blows up the claim that there should be a “one-size fits all” chart for motoric development in infants and that there should be race-specific milestones. This means that we should throw out the WEIRD assumptions when it comes to motoric development of infants (Karasik et al, 2011). They discuss research in other cultures where African, Caribbean and Indian caregivers massage the muscles of babies, stretch their limbs, toss them in their air, sit them up, and walk with them while helping them which then shapes their muscles and has them learn the mind-muscle connections needed to be able to learn how to eventually walk. And it also seems that random assignment to exercise excelerates how quickly an infant walks. White infants also sit at 6 months while black infants sit at 4 months. Nonetheless, it is clear that culture and context can indeed shape motoric development in groups around the world.
A bio-cultural view of motor development
When it comes to biological influences on motor development, sex and age are two important variables (Escolano-Perez, Sanchez-Lopez, and Herrero-Nivela, 2021). Important to this, of course, is that the individual must be normal, and they must have a normal brain with normal vision and spatial skills. They must be able to hear (to eventually follow commands and hear what is going on in their environment to change their course of action if need be). Further, the child’s home environment and gestational age influence different portions of motoral development (Darcy, 2022). After infants begin crawling, their whole world changes and they process visual motion better and faster, being able to differentiate between different speeds and directions, so a stimulating environment for the infant can spur the development of the brain (Van der Meer and Van der Weel, 2022). Biological maturation and body weight also affect motor development. Walking develops naturally, but walking and motor competence need to be nurtured for the child to reach their full potential; lower motor competence is related to higher body weight (Drenowatz and Greier, 2019).
One study on Dutch and Israeli infants even found—using developmental niche construction—that “infant motor development indeed is at least partly culturally constructed [which] emphasizes the importance of placing infant motor development studies into their ‘cultural cradle‘ (Oudgeneong, Atun-Eni, and Schaik, 2020). Gross motor development—rolling over, crawling, alternating kicks, moving from lying to sitting, and having tummy time—is recognized by the WHO. Further, children from different cultures have different experiences, which also could lead to, for example, not doing things that are conducive to the development of gross motor development (Angulo-Barroso et al, 2010). Moreover, motor development is embodied, enculturated, embedded, and enabling (Adolph and Hoch, 2020). It is also known that differences in the cultural environment “have a non-negligible effect on motor development” (Bril, 1986). Motor development also takes place in physical environments and is purposive and goal-directed (Hallemans, Verbeque, and de Walle, 2020).
So putting this all together, we have conceptualized motor development as a dynamic process which is influenced by a complex interplay of biological and cultural factors (Barnes, Zieff, and Anderson, 1999). Biological factors like sex, age, health, sensory abilities, and socio-cultural factors like home environment and developmental niches explain motor development and differences in them between individuals. The cultural differences, though, can impede motoral development, and not allow one to reach milestones they would have otherwise reached in a different cultural environment, just like if one couldn’t hear or see would have trouble reaching developmental milestones.
Children of course grow up in cultural environments and contexts and so they are culturally situated. So what this means is that both the cultural and social environment the child finds themselves in will of course then influence their physical and mental development and lead them to their milestones they hit which is dictated by the normal biology they have which then is allowed by the socio-cultural environment they are born into. So we have the bio-cultural view on motor development, and beyond the cultural environment the child finds themselves in, the interactions they have between parents and caregivers—more knowledgeable others—can be pertinent to their motor development and reaching of developmental milestones. Cultural practices and expectations could emphasize certain milestones over others and then guide the child towards the trajectory. So the framework recognizes that normal biology and sensory perceptions are needed for the development of normal motor development, but that cultural and social differences in that context will spur motor development in the child who finds themselves in different cultures.
Conclusion
Was Rushton right about this? Yes, I think he was. The recent literature on the matter speaks to this. But that doesn’t mean that his r/K selection theory is true. There are differences in motor development between races. But what is interesting is the interaction between biological and cultural factors that spur motor development. The question of black motor precocity, however, is a socio-political question, since science is a social convention influenced by the values of the scientist in question. Now, to the best of my knowledge, Rushton himself never carried out studies on this, he just collated them to use them for his racial trait matrix. However, it’s quite clear that Rushton was politically politically and socially motivated to prove that his theory was true.
But physical differences between the races are easy enough to prove, and of course they are due to biological and cultural interactions. There are differences in skin color and their properties between blacks and whites (Campiche et al, 2019). There is a 3 percent center of mass difference between blacks and whites which explains why each race excels at running and swimming (Bejan, Jones, and Charles, 2010). There are differences in body composition between Asians and whites which means, at the same BMI, Asians would have thicker skin folds and higher body fat than whites (Wang et al, 1994; WHO expert consultation, 2004; Wang et al, 2011). Just like at the same BMI, blacks have lower body fat and thinner skin folds than whites (Vickery, Cureton, and Collins, 1988; Wagner and Heyward, 2000; Flegal et al, 2010). There are differences in menarche and thelarche between blacks and whites (Wagner and Heyward, 2000; Kaplowitz, 2008; Reagan et al, 2013; Cabrera et al, 2014; Deardorff et al, 2014; ). There are differences in anatomy and physiology and somatotype between blacks and whites and these differences would explain how the races would perform on the big four lifts. There are interesting and real physical differences between races.
So obviously, what is considered “normal” is different in different cultures, and motor development is no different. So just like I think we should have different BMI and skin fold charts for different races, so too should we have different developmental milestones for different races and cultures. The discussion here is clear, since what is “average” and “normal” is different based on race and culture. Like for instance, black babies begin walking around 11 months, white babies around 12 months and Native American babies at 13 months. So while parents may be worried that their child didn’t hit a certain developmental milestone like walking, sitting, rolling, taking a bio-cultural approach will assuage these worries.
Nonetheless, while Rushton was right about race and motor development, we need to center his research project in context. He was clearly motivated, despite the numerous and forceful critiques of his framework, to prove that he was right. But the continuance of Rushton pushing his theory up until his death shows me that he was quite obviously socially and politically motivated, contrary to what he may have said.
We have approached this paper from the stance that science is a social activity, with all observations influenced by, as well as reflective of, the values of scientists and the political leanings of the sociocultural context within which research is conducted. We suggest that when questions of group difference are pursued in science, awareness of how the categories themselves have been shaped by social and historical forces, as well as of the potential effects on society, is important. (Barnes, Zieff, and Anderson, 1999)
“Missing Heritability” and Missing Children: On the Issues of Heritability and Hereditarian Interpretations
3100 words
“Biological systems are complex, non-linear, and non-additive. Heritability estimates are attempts to impose a simplistic and reified dichotomy (nature/nurture) on non-dichotomous processes.” (Rose, 2006)
“Heritability estimates do not help identify particular genes or ascertain their functions in development or physiology, and thus, by this way of thinking, they yield no causal information.” (Panofsky, 2016: 167)
“What is being reported as ‘genetic’, with high heritability, can be explained by difference-making interactions between real people. In other words, parents and children are sensitive, reactive, living beings, not hollow mechanical or statistical units.” (Richardson, 2022: 52)
Introduction
In the world of behavioral genetics, it is claimed that studies of twins, adoptees and families can point us to the interplay between genetic and environmental influences on complex behavioral traits. To study this, they use a concept called “heritability”—taken from animal breeding—which estimates the the degree of variation in a phenotypic trait that is due to genetic variation amongst individuals in the studied population. But upon the advent of molecular genetic analysis after the human genome project, something happened that troubled behavioral genetic researchers: The heritability estimates gleaned from twin, family and adoption studies did not match the estimates gleaned from the molecular genetic studies. This then creates a conundrum—why do the estimates from one way of gleaning heritability don’t match to other ways? I think it’s because biological models represent a simplistic (and false) model of biological causation (Burt and Simon, 2015; Lala, 2023). This is what is termed “missing heritability.” This raises questions that aren’t dissimilar to when a child dissappears.
Imagine a missing child. Imagine the fervor a family and authorities go through in order to find the child and bring them home. The initial fervor, the relentless pursuit, and the agonizing uncertainty constitute a parallel narrative in behavioral genetics, where behavioral geneticists—like the family of a missing child and the authorities—find themselves grappling with unforseen troubles. In this discussion, I will argue that the additivity assumption is false, that this kind of thinking is a holdover from the neo-Darwinian Modern Synthesis, that hereditarians have been told for decades that heritability just isn’t useful for what they want to do, and finally “missing heritability” and missing children are in some ways analogous, but that there is a key difference: The missing children actually existed, while the “missing heritability” never existed at all.
The additivity assumption
Behavioral geneticists pay lip service to “interactions”, but then conceptualize these interactions as due to additive heritability (Richardson, 2017a: 48-49). But the fact of the matter is, genetic interactions create phantom heritability (Zuk et al, 2012). When it comes to the additive claim of heritability, that claim is straight up false.
The additive claim is one of the most important things for the utility of the concept of heritability for the behavioral geneticist. The claim that heritability estimates for a trait are additive means that the contribution of each gene variant is independent and they all sum up to explain the overall heritability (Richardson 2017a: 44 states that “all genes associated with a trait (including intelligence) are like positive or negative charges“). But in reality, gene variants aren’t independent effects, they interact with other genes, the environment and other developmental resources. In fact, violations of the additivity assumption are large (Daw, Guo, and Harris, 2015).
Gene-gene, gene-environment, and environmental factors can lead to overestimates of heritability, and they are non-additive. So after the 2000s with the completion of the human genome project, these researchers realized that the genetic variants that heritability they identified using molecular genetics did not jive with the heritability they computed from twin studies from the 1920s until the late 1990s and then even into the 2020s. So the expected additive contribution of heritability fell short in actually explaining the heritability gleaned from twin studies using molecular genetic data.
Thinking of heritability as a complex jigsaw puzzle may better help to explain the issue. The traditional view of heritability assumes that each genetic piece fits neatly into the puzzle to then complete the overall genetic picture. But in reality, these pieces may not be additive. They can interact in unexpected ways which then creates gaps in our understanding, like a missing puzzle piece. So the non-additive effects of gene variants which includes interactions and their complexities, can be likened to missing pieces in the heritability puzzle. The unaccounted-for genetic interactions and nuances then contribute to what is called “missing heritability.” So just as one may search and search for missing puzzle pieces, so to do behavioral geneticists search and search for the “missing heritability”.
So heritability assumes no gene-gene and gene-environment interaction, no gene-environment correlation, among other false or questionable assumptions. But the main issue, I think, is that of the additivity assumption—it’s outright false and since it’s outright false, then it cannot accurately represent the intricate ways in which genes and other developmental resources interact to form the phenotype.
If heritability estimates assume that genetic influences on a trait are additive and independent, then heritability estimates oversimplify genetic complexity. If heritability estimates oversimplify genetic complexity, then heritability estimates do not adequately account for gene-environment interactions. If heritability does not account for gene-environment interactions, then heritability fails to capture the complexity of trait inheritance. Thus, if heritability assumes that genetic influences on a trait are additive and independent, then heritability fails to capture the complexity of trait inheritance due to its oversimplified treatment of genetic complexity and omission of gene-environment interactions.
One more issue, is that of the “heritability fallacy” (Moore and Shenk, 2016). One commits a heritability fallacy when they assume that heritability is an index of genetic influence on traits and that heritability can tell us anything about the relative contribution of trait inheritance and ontogeny. Moore and Shenk (2016) then make a valid conclusion based on the false belief that heritability us anything about the “genetic strength” on a trait:
In light of this, numerous theorists have concluded that ‘the term “heritability,” which carries a strong conviction or connotation of something “[in]heritable” in the everyday sense, is no longer suitable for use in human genetics, and its use should be discontinued.’31 Reviewing the evidence, we come to the same conclusion. Continued use of the term with respect to human traits spreads the demonstrably false notion that genes have some direct and isolated influence on traits. Instead, scientists need to help the public understand that all complex traits are a consequence of developmental processes.
“Missing heritability”, missing children
Twin studies traditionally find heritability to be estimated between 50 and 80 percent for numerous traits (eg Polderman et al, 2015; see Joseph’s critique). But as alluded to earlier, molecular studies have found heritabilities of 10 percent or lower (eg, Sniekers et al, 2017; Savage et al, 2018; Zabaneh et al, 2018). This discrepancy between different heritability estimates using different tools is what is termed “missing heritability” (Mathhews and Turkheimer, 2022). But the issue is, increasing the sample sizes will merely increase the chance of spurious correlations (Calude and Longo, 2018), which is all these studies show (Richardson, 2017b; Richardson and Jones, 2019).
This tells me one important thing—behavioral geneticists have so much faith in the heritability estimates gleaned from twin studies that they assume that the heritability is “missing” in the newer molecular genetic studies. But if something is “missing”, then that implies that it can be found. They have so much faith that eventually, as samples get higher and higher in GWAS and similar studies, that we will find the heritability that is missing and eventually, be able to identify genetic variants responsible for traits of interest such as IQ. However I think this is confused and a simple analogy will show why.
When a child goes missing, it is implied that they will be found by authorities, whether dead or alive. Now I can liken this to heritability. The term “missing heritability” comes from the disconnect between heritability estimates gleaned from twin studies and heritability estimates gleaned from molecular genetic studies like GWAS. So the implication here is, since twin studies show X percent heritability (high heritability), and molecular genetic studies show Y percent heritability (low heritability) – which is a huge difference between estimates between different tools – then the implication is that there is “missing heritability” that must be explained by rare variants or other factors.
So just like parents and authorities try so hard to find their missing children, so to do behavioral geneticists try so hard to find their “missing heritability.” As families endure anguish as they try to find their children, this is then mirrored in the efforts of behavioral geneticists to try and close the gap between two different kinds of tools that glean heritability.
But there is an important issue at play here—namely the fact that missing children actually exist, but “missing heritability” doesn’t, and that’s why we haven’t found it. Although some parents, sadly, may never find their missing children, the analogy here is that behavioral geneticists will never find their own “children” (their missing heritability) because it simply does not exist.
Spurious correlations
Even increasing the sample sizes won’t do anything, since the larger the sample size, the bigger chance for spurious correlations, and that’s all GWAS studies for IQ are (Richardson and Jones, 2019), while correlations with GWAS are inevitable and meaningless (Richardson, 2017b). Denis Noble (2018) puts this well:
As with the results of GWAS (genome-wide association studies) generally, the associations at the genome sequence level are remarkably weak and, with the exception of certain rare genetic diseases, may even be meaningless (13, 21). The reason is that if you gather a sufficiently large data set, it is a mathematical necessity that you will find correlations, even if the data set was generated randomly so that the correlations must be spurious. The bigger the data set, the more spurious correlations will be found (3). The current rush to gather sequence data from ever larger cohorts therefore runs the risk that it may simply prove a mathematical necessity rather than finding causal correlations. It cannot be emphasized enough that finding correlations does not prove causality. Investigating causation is the role of physiology.
Nor does finding higher overall correlations by summing correlations with larger numbers of genes showing individually tiny correlations solve the problem, even when the correlations are not spurious, since we have no way to find the drugs that can target so many gene products with the correct profile of action.
The Darwinian model
But the claim that there is a line that goes from G (genes) to P (phenotype) is just a mere holdover from the neo-Darwinian modern synthesis. The fact of the matter is, “HBD” and hereditarianism are based on reductionistic models of genes and how they work. But the reality is, genes don’t work how they think they do, reality is much more complex than they assume. Feldman and Ramachandran (2018) ask “Missing compared to what?”, effectively challenging the “missing heritability” claim. As Feldman and Ramachandran (2018) ask, would Herrnstein and Murray have written The Bell Curve if they believed that the heritability of IQ were 0.30? I don’t think they would have. In any case, such a belief in the heritability of IQ being between 0.4 and 0.8 shows the genetic determinist assumptions which are inherent in this type of “HBD” genetic determinist thinking.
Amusingly, as Ned Block (1995) noted, Murray said in an interview that “60 percent of the intelligence comes from heredity” and that that heritability is “not 60 percent of the variation. It is 60 percent of the IQ in any given person.” Such a major blunder from one of the “intellectual spearheads” of the “HBD race realist” movement…
Behavioral geneticists claim that the heritability is missing only because sample sizes are low, and as sample sizes increase, the missing heritability based on associated genes will be found. But this doesn’t follow at all since increasing sample sizes will just increase spurious hits of genes correlated with the trait in question but it says absolutely nothing about causation. Nevertheless, only a developmental perspective can provide us mechanistic knowledge and so-called heritability of a phenotype cannot give us such information because heritability isn’t a mechanistic variable and doesn’t show causation.
Importantly, a developmental perspective provides mechanistic knowledge that can yield practical treatments for pathologies. In contrast, information about the “heritability” of a phenotype—the kind of information generated by twin studies—can never be as useful as information about the development of a phenotype, because only developmental information produces the kind of thorough understanding of a trait’s emergence that can allow for successful interventions. (Moore 2015: 286)
The Darwinian model and it’s assumptions are inherent in thinking about heritability and genetic causation as a whole and are antithetical to developmental, EES-type thinking. Since hereditarianism and HBD-type thinking are neo-Darwinist, it then follows that such thinking is inherent in their beliefs, assumptions, and arguments.
Conclusion
Assumptions of heritability simply do not hold. Heritability, quite simply, isn’t a characteristic of traits but it is a characteristic of “relationships in a population observed in a particular setting” (Oyama, 1985/2000). Heritability estimates tell us absolutely nothing about development, nor the causes of development. Heritability is a mere breeding statistic and tells us nothing at all about the causes of development or whether or not genes are “causal” for a trait in question (Robette, Genin, and Clerget-Darpoux, 2022). It is key to understand that heritability along with the so-called “missing heritability” are based on reductive models of genetics that just do not hold, especially with newer knowledge that we have from systems biology (eg, Noble, 2012).
The assumption that heritability estimates tell us anything useful about genetics, traits, and causes along with a reductive belief in genetic causation for the ontogeny of traits has wasted millions of dollars. Now we need to grapple with the fact that heritability just doesn’t tell us anything about genetic causes of traits, but that genes are necessary, not sufficient, causes for traits because no genes (along with other developmental resources) means no organism. Also coming from twin, family and adoption studies are Turkheimer’s (2000) so-called “laws of behavioral genetics.” Further, the falsity of the EEA (equal environments assumption) is paramount here, and since the EEA is false, genetic conclusions from such studies are invalid (Joseph et al, 2015). There is also the fact that heritability is based on a false biological model. The issue is that heritability rests on a “conceptual model is unsound and the goal of heritability studies is biologically nonsensical given what we now know about the way genes work” (Burt and Simons, 2015: 107). What Richardson (2022) terms “the agricultural model of heritability” is known as false. In fact, the heritability of “IQ” is higher than any heritability found in the animal kingdom (Schonemann, 1997). Why this doesn’t give any researcher pause is beyond me.
Nonetheless, the Darwinian assumptions that are inherent in behavioral genetic, HBD “race realist” thinking are false. And the fact of the matter is, increasing the sample size of molecular genetic studies will only increase the chances of spurious correlations and picking up population stratification. So, it seems that using heritability to show genetic and environmental causes is a bust and has been a bust ever since Jensen revived the race and IQ debate in 1969, along with the subsequent responses that Jensen received against his argument which then led to the 1970s as being a decade in which numerous arguments were made against the concept of heritability (eg, Layzer, 1974).
It has also been pointed out to racial hereditarians for literally decades that heritability is is a flawed metric (Layzer, 1974; Taylor, 1980; Bailey, 1997; Schonemann, 1997; Guo, 2000; Moore, 2002; Rose, 2006; Schneider, 2007; Charney, 2012, 2013; Burt and Simons, 2015; Panofsky, 2014; Joseph et al, 2015; Moore and Shenk, 2016; Panofsky, 2016; Richardson, 2017; Lerner, 2018). These issues—among many more—lead Lerner to conclude:
However, the theory and research discussed across this chapter and previous ones afford the conclusion that no psychological attribute is pre-organized in the genes and unavailable to environmental influence. That is, any alleged genetic difference (or “inferiority”) of African Americans based on the high heritability of intelligence would seem to be an attribution built on a misunderstanding of concepts basic to an appropriate conceptualization of the nature–nurture controversy. An appreciation of the coaction of genes and context—of genes↔context relations—within the relational developmental system, and of the meaning, implications, and limitations of the heritability concept, should lead to the conclusion that the genetic-differences hypothesis of racial differences in IQ makes no scientific sense. (Lerner, 2018: 636)
That heritability doesn’t address mechanisms and ignores genetic factors, along with being inherently reductionist means that there is little to no utility of heritability for humans. And the complex, non-additive, non-linear aspects of biological systems are attempts at reducing biological systems to their component parts, (Rose, 2006), making heritability, again, inherently reductionist. We have to attempt to analyzed causes, not variances (Lewontin, 1974), which heritability cannot do. So it’s very obvious that the hereditarian programme which was revived by Jensen (1969)—and based on twin studies which were first undertaken in the 1920s—is based on a seriously flawed model of genes and how they work. But, of course, hereditarians have an ideological agenda to uphold, so that’s why they continue to pursue “heritability” in order to “prove” that “in part”, racial differences in many socio-behavioral traits—IQ included—are due to genes. But this type of argumentation quite clearly fails.
The fact of the matter is, “there are very good reasons to believe gene variations are at best irrelevant to common disorders and at worst a distraction from the social and political roots of major public health problems generally and of their unequal distribution in particular” (Chaufan and Joseph 2013: 284). (Also see Joseph’s, 2015 The Trouble with Twin Studies for more argumentation against the use of heritability and it’s inflation due to false assumptions along with arguments against “missing heritability.”) In fact, claims of “missing heritability” rest on “genetic determinist beliefs, a reliance on twin research, the use of heritability estimates, and the failure to seriously consider the possibility that presumed genes do not exist” (Joseph, 2012). Although it has been claimed that so-called rare variants explain the “missing heritability” (Genin, 2020), this is nothing but cope. So the heritability was never missing, it never existed at all.
Hypertension, Brain Volume, and Race: Hypotheses, Predictions and Actionable Strategies
2300 words
Introduction
Hypertension (HT, also known as high blood pressure, BP) is defined as a BP of 140/90. But more recently, the guidelines were changed making HT being defend as a BP over 130/90 (Carey et al, 2022; Iqbal and Jamal, 2022). One 2019 study showed that in a sample with an age range of 20-79, 24 percent of men and 23 percent of women could be classified as hypertensive based on the old guidelines (140/90) (Deguire et al, 2019). Having consistent high BP could lead to devestating consequences like (from the patient’s perspective) hot flushes, dizziness, and mood disorders (Goodhart, 2016). However, one serious problem with HT is the issue that consistently high BP is associated with a decrease in brain volume (BV). This has been seen in two systematic reviews and meta-analyses (Alosco et al, 2013; Beauchet et al, 2013; Lane et al, 2019; Alateeq, Walsh and Cherbuin, 2021; Newby et al, 2022) while we know that long-standing hypertension has deleterious effects on brain health (Salerno et al, 1992). However, it’s not only high BP that’s related to this, it’s also lower BP in conjuction with lower pulse pressure (Muller et al, 2010; Foster-Dingley, 2015). So what this says to me is that too much or too little blood flow to the brain is deleterious for brain health.I will state the hypothesis and then I will state the predictions that follow from it. I will then provide three reasons why I think this relationship occurs.
The hypothesis
The hypothesis is simple: high BP (hypertension, HT) is associated with a reduced brain volume. This relationship is dose-dependent, meaning that the extent and duration of HT correlates with the degree of BV changes. So the hypothesis suggests that there is a relationship—an association—between HT and brain volume, where people with HT will be more likely to have decreased BVs than those who lack HT—that is, those with BP in the normal range.
The dose-dependent relationship that has been observed (Alateeq, Walsh and Cherbuin, 2021), and this shows that as HT increases and persists over time, the effects of decreased BV become more pronounced. This relationship suggests that it’s not a binary, either-or situation, present or absent situation, but that it varies across a continuum. So people with shorter-lasting HT will have fewer effects than those with constant and consistent elevated BP and they will then show subsequent higher decreases in BV. This dose-dependent relationship also suggests that as BP continues to elevate, the decrease in BV will worsen.
This dose-dependent relationship implies a few things. The consequences of HT on BV aren’t binary (either or), but are related to the severity of HT, how long one has HT, and at what age they have HT and that it varies on a continuum. For instance, people with mild or short-lasting HT would experience smaller reductions in BV than those that have severe or long-standing HT. The dose-dependent relationship also suggests that the longer one has HT without treatment, the more severe and worse the reduction in BV will be if it is uncontrolled. So as BP continues to elevate, it may lead to a gradual reduction in BV. So the relationship between HT and BV isn’t uniform, but it varies based on the intensity and duration of high BP.
So the hypothesis suggests that HT isn’t just a risk factor for cardiovascular disease, but it’s also a risk factor for decreased BV. This seems intuitive, since the higher one’s BP, the more likely it is that there is the beginnings of a blockage somewhere in the intricate system of blood vessels in the body. And since the brain is a vascular organ, then by decreasing the amount of blood flowing to it, this then would lead to cell death, white matter lesions which would lead to a smaller BV. One newer study showed, with a sample of Asians, whites, blacks, and “Latinos” that, compared to those with normal BP, those who were transitioning to higher BP or already had higher BP had lower brain connectivity, decreased cerebral gray matter and frontal cortex volume, while this change was worse for men (George et al, 2023). Shang et al (2021) showed that HT diagnosed in early and middle life but not late life was associated with decreased BV and increased risk of dimentia. This, of course, is due to the slow cumulative effects of HT and it’s effects on the brain. While Power et al (2016) “The pattern of hypertension ~15 years prior and hypotension concurrent with neuroimaging was associated with smaller volumes in regions preferentially affected by Alzheimer’s disease.” But not only is BP relevant here, so is the variability of BP at night (Gutteridge et al, 2022; Yu et al, 2022). Alateeq, Walsh and Cherbuin (2021) conclude that:
Although reviews have been previously published in this area, they only investigated the effects of hypertension on brain volume [86]. To the best of our knowledge, this study’s the first systematic review with meta-analysis providing quantitative evidence on the negative association between continuous BP and global and regional brain volumes. Our results suggest that heightened BP across its whole range is associated with poorer cerebral health which may place individuals at increased risk of premature cognitive decline and dementia. It is therefore important that more prevention efforts be directed at younger populations with a greater focus on achieving optimal BP rather than remaining below clinical or pre-clinical thresholds[5].
One would think that a high BP would actually increase blood flow to the brain, but HT actually causes alterations in the flow of blood to the brain which leads to ischaemia and it causes the blood-brain barrier to break down (Pires et al, 2013). Essentially, HT has devestating effects on the brain which could lead to dimentia and Alzheimer’s (Iadecola and Davisson, 2009).
So the association between HT and decreased BV means that individuals with HT can experience alterations in BV in comparison to those with normal BP. The hypothesis also suggests that there are several mechanisms (detailed below), which may lead to various physiological and anatomic changes in the brain, such as vascular damage, inflammation and tissue atrophy.
The mechanisms
(1) High BP can damage blood vessels in the brain, which leads to reduced blood flow. This is called “cerebral hypoperfusion.” The reduced blood flow can deprive the cells in the brain of oxygen and nutrients, which cause them to shrink or die which leads to decreased brain volume (BV). Over time, high BP can damage the arteries, making them less elastic
(2) Over a long period of time having high BP, this can cause hypertensive encephalopathy, which is basically brain swelling. A rapid increase in BP could over the short term increase BV, but left untreated it could lead to brain damage and atrophy over time.
And (3) Chronically high BP can lead to the creation of white matter lesions on the brain, and the lesions are areas of damaged brain tissue which could result in microvascular changes caused by high BP (hypertension, HT). Thus, over time, the accumulation of white matter lesions could lead to a decrease in brain volume. HT can contribute to white matter lesions in the brain, which are then associated with cognitive changes and decreased BV, and these lesions increase with BP severity.
So we have (1) cerebral hypoperfusion, (2) hypertensive encephalopathy, and (3) white matter lesions. I need to think/read more on which of these could lead to decreased BV, or if they all actually work together to decrease BV. We know that HT damages blood vessels, and of course there are blood vessels in the brain, so it then follows that HT would decrease BV.
I can also detail a step-by-step mechanism. The process beings with consistently elevated BP, which could be due to various factors like genetics, diet/lifestyle, and underlying medical conditions. High BP then places increased strain on the blood vessels in the body, including those in the brain. This higher pressure could then lead to structural change of the blood vessels over time. Then, chronic HT over time can lead to endothelial dysfunction, which could impair the ability of blood vessels to regulate blood flow and maintain vessel integrity. The dysfunction can result in oxidative stress and inflammation.
Then as a response to prolonged elevated BP, blood vessels in the brain could undergo vascular remodeling, which involves changes im blood vessel structure and thickness, which can then affect blood flow dynamics. Furthermore, in some cases, this could lead to something called cerebral small vessel disease which involves damage to the small blood vessels in the brain including capillaries and arterioles. This could impair delivery of oxygen and nutrients to brain tissue which could lead to cell death and consequently a decrease in BV. Then reduced blood flow along compromised blood vessel integrity could lead to cerebral ischaemia—reduced blood supply—and hypoxia—reduced oxygen supply—in certain parts of the brain. This can then result in neural damage and eventually cell death.
Then HT-related vascular changes and cerebral small vessel disease can trigger brain inflammation. Prolonged exposure to neural inflammation, hypoxia and ischemia can lead to neuronal atrophy, where neurons shrink and lose their functional integrity. HT can also increase the incidence of white matter lesions in the brain which can be seen in neuroimages, which involve areas of white matter tissue which become damaged. Finally, over time, the cumulative effects of the aforementioned processes—vascular changes, inflammation, neural atrophy, and white matter changes could lead to a decrease in BV. This reduction can manifest as brain atrophy which is then observed in parts of the brain which are susceptible and vulnerable to the effects of HT.
So the step-by-step mechanism goes like this: elevated BP —> increased vascular strain —> endothelial dysfunction —> vascular remodeling —> cerebral small vessel disease —> ischemia and hypoxia —> inflammation and neuroinflammation —> neuronal atrophy —> white matter changes —> reduction in BV.
Hypotheses and predictions
H1: The severity of HT directly correlates with the extent of BV reduction. One prediction would be that people with more severe HT would exhibit greater BV decreases than those with moderate (less severe) HT, which is where the dose-dependent relationship comes in.
H2: The duration of HT is a critical factor in BV reduction. One prediction would be that people with long-standing HT will show more significant BV changes than those with recent onset HT.
H3: Effective BP management can mitigate BV reduction in people with HT. One prediction would be that people with more controlled HT would show less significant BV reduction than those with uncontrolled HT.
H4: Certain subpopulations may be more susceptible to BV decreases due to HT. One prediction is that certain factors like age of onset (HT at younger age), genetic factors (some may have certain gene variants that make them more susceptible and vulnerable to damage caused by elevated BP), comorbities (people with diabetes, obesity and heart problems could be at higher risk of decreased BV due to the interaction of these factors), ethnic/racial factors (some populations—like blacks—could be at higher risk of having HT and they could be more at risk due to experiencing disparities in healthcare and treatment.
The hypotheses and predictions generated from the main proposition that HT is associated with a reduction in BV and that the relationship is dose-dependent can be considered risky, novel predictions. They are risky in the sense that they are testable and falsifiable. Thus, if the predictions don’t hold, then it could falsify the initial hypothesis.
Blacks and blood pressure
Due to this, for populations like black Americans, this is significant. About 33 percent of blacks have hypertension (Peters, Arojan, and Flack, 2006), while urban blacks are more likely to have elevated BP than whites (Lindhorst et al, 2007). Though Non, Gravlee, and Mulligan (2012) showed that racial differences in education—not genetic ancestry—explained differences in BP in blacks compared to whites. Further, Victor et al (2018) showed that in black male barbershop attendees who had uncontrolled BP, that along with medication and outreach, this lead to a decrease in BP. Williams (1992) cited stress, socioecologic stress, social support, coping patterns, health behavior, sodium, calcium, and potassium consumption, alcohol consumption, and obesity as social factors which lead to increased BP.
Moreover, consistent with the hypothesis discussed here (that chronic elevated BP leads to reductions in BV which lead to a higher chance of dementia and Alzheimer’s), it’s been shown that vulnerability to HT is a major determinate in the risk of acquiring Alzheimer’s (Clark et al, 2020; Akushevic et al, 2022). It has also been shown that “a lifetime of racism makes Alzheimer’s more common in black Americans” and consistent with the discussion here since racism is associated with stress which is associated with elevated BP, then consistent events of racial discrimination would lead to consistent and elevated BP which would then lead to decreased BV and then a higher chance of acquitting Alzheimer’s. But, there is evidence that blood pressure drugs (in this case telmisartan) reduce the incidence of Alzheimer’s in black Americans (Zhang et al, 2022) while the same result was also seen using antihyperintensive medications in blacks which led to a reduction in incidence of dementia (Murray et al, 2018), which lends credence to the discussed hypothesis. Stress and poverty—experiences—and not ancestry could explain higher rates of dementia in black Americans as well. Thus, since blood pressure could explain higher rates of dementia in black populations, this then lends credence to the discussed hypothesis.
Conclusion
The evidence that chronic elevated BP leads to reductions in BV are well-studied and the mechanisms are well-known. I discussed the hypothesis that chronically elevated BP leads to reduced blood flow to the brain which decreases BV. I then discussed the mechanisms behind the relationship, and then hypotheses and predictions that follow from them. Lastly, I discussed the well-known fact that blacks have higher rates of BP, and also higher rates of dementia and Alzheimer’s, and linked the fact that they have higher rates of BP to those maladies.
So by catching chronically elevated BP in the early ages, since the earlier one has high BP the more likely they are to have reduced brain volume and the associated maladies, we can then begin to fight the associated issues before they coalesce, since we know the mechanisms behind them, along with the fact that blood pressure drugs and antihypertensive medications decrease incidences of dementia and Alzheimer’s in black Americans.
The Theory of African American Offending versus Hereditarian Explanations of Crime: Exploring the Roots of the Black-White Crime Disparity
3450 words
Why do blacks commit more crime? Biological theories (racial differences in testosterone and testosterone-aggression, AR gene, MAOA) are bunk. So how can we explain it? The Unnever-Gabbidon theory of African American offending (TAAO) (Unnever and Gabbidon, 2011)—where blacks’ experience of racial discrimination and stereotypes increases criminal offenses—has substantial empirical support. To understand black crime, we need to understand the unique black American experience. The theory not only explains African American criminal offending, it also makes predictions which were borne out in independent, empirical research. I will compare the TAAO with hereditarian claims of why blacks commit more crime (higher testosterone and higher aggression due to testosterone, the AR gene and MAOA). I will show that hereditarian theories make no novel predictions and that the TAAO does make novel predictions. Then I will discuss recent research which shows that the predictions that Unnever and Gabbidon have made were verified. Then I will discuss research which has borne out the predictions made by Unnever and Gabbidon’s TAAO. I will conclude by offering suggestions on how to combat black crime.
The folly of hereditarianism in explaining black American offending
Hereditarians have three main explanations of black crime: (1) higher levels of testosterone and high levels of testosterone leading to aggressive behavior which leads to crime; (2) low activity MAOA—also known in the popular press as “the warrior gene”—could be more prevalent in some populations which would then lead to more aggressive, impulsive behavior; and (3) the AR gene and AR-CAG repeats with lower CAG repeats being associated with higher rates of criminal activity.
When it comes to (1), the evidence is mixed on which race has higher levels of testosterone (due to low-quality studies that hereditarians cite for their claim). In fact, two recent studies showed that non-Hispanic blacks didn’t have higher levels of testosterone than other races (Rohrmann et al, 2007; Lopez et al, 2013). Contrast this with the classical hereditarian response that blacks indeed do have higher rates of testosterone than whites (Rushton, 1995)—using Ross et al (1986) to make the claim. (See here for my response on why Ross et al is not evidence for the hereditarian position.) Although Nyante et al (2012) showed a small increase in testosterone in blacks compared to whites and Mexican Americans using longitudinal data, the body of evidence shows that there is no to small differences in testosterone between blacks and whites (Richard et al, 2014). So despite claims that “African-American men have repeatedly demonstrated serum total and free testosterone levels that are significantly higher than all other ethnic groups” (Alvarado, 2013: 125), claims like this are derived from flawed studies, and newer more representative analyses show that there is a small difference in testosterone between blacks and whites to no difference.
Nevertheless, even if blacks have higher levels of testosterone than other races, then this would still not explain racial differences in crime, since heightened aggression explains T increases, high T doesn’t explain heightened aggression. HBDers seem to have cause and effect backwards for this relationship. Injecting individuals with supraphysiological doses of testosterone as high as 200 and 600 mg per week does not cause heightened anger or aggression (Tricker et al, 1996; O’Connor et, 2002). If the hereditarian hypothesis on the relationship between testosterone and aggression were true, then we would see the opposite finding from what Tricker et al and O’Connor et al found. Thus this discussion shows that hereditarians are wrong about racial differences in testosterone and that they are wrong about causality when it comes to the T-aggression relationship. (The actual relationship is aggression causing increases in testosterone.) So this argument shows that the hereditarian simplification on the T-aggression relationship is false. (But see Pope, Kouri and Hudson, 2000 where they show that a 600 mg dose of testosterone caused increased manic symptoms in some men, although in most men there was little to no change; there were 8 “responders” and 42 “non-responders.”)
When it comes to (2), MAOA is said to explain why those who carry low frequency version of the gene have higher rates of aggression and violent behavior (Sohrabi, 2015; McSwiggin, 2017). Sohrabi shows that while the low frequency version of MAOA is related to higher rates of aggression and violent behavior, it is mediated by environmental effects. But MAOA, to quote Heine (2017), can be seen as the “Everything but the kitchen sink gene“, since MAOA is correlated with so many different things. But at the and of the day, we can’t blame “warrior genes” for violent, criminal behavior. Thus, the relationship isn’t so simple, so this doesn’t work for hereditarians either.
Lastly when it comes to (3), due to the failure of (1), hereditarians tried looking to the AR gene. Researchers tried to relate CAG repeat length with criminal behaviors. For instance, Geniole et al (2019) tried to argue that “Testosterone thus appears to promote human aggression through an AR-related mechanism.” Ah, the last gasps to explain crime through testosterone. But there is no relationship between CAG repeats, adolescent risk-taking, depression, dominance or self-esteem (Vermeer, 2010) and the number of CAG repeats in men and women (Valenzuela et al, 2022). So this, too, fails. (Also take look at the just-so story on why African slave descendants are more sensitive to androgens; Aiken, 2011.)
Now that I have shown that the three main hereditarian explanations for higher black crime are false, now I will show why blacks have higher rates of criminal offending than other races, and the answer isn’t to be found in biology, but sociology and criminology.
The Unnever-Gabbidon theory of African American criminal offending and novel predictions
In 2011, criminologists Unnever and Gabbidon published their book A Theory of African American Offending: Race, Racism, and Crime. In the book, they explain why they formulated the theory and why it doesn’t have any explanatory or predictive power for other races. That’s because it centers on the lived experiences of black Americans. In fact, the TAAO “incorporates the finding that African Americans are more likely to offend if they associate with delinquent peers but we argue that their inadequate reinforcement for engaging in conventional behaviors is related to their racial subordination” (Unnever and Gabbidon, 2011: 34). The TAAO focuses on the criminogenic effects of racism.
Our work builds upon the fundamental assumption made by Afrocentists that an understanding of black offending can only be attained if their behavior is situated within the lived experiences of being African American in a conflicted, racially stratified society. We assert that any criminological theory that aims to explain black offending must place the black experience and their unique worldview at the core of its foundation. Our theory places the history and lived experiences of African American people at its center. We also fully embrace the Afrocentric assumption that African American offending is related to racial subordination. Thus, our work does not attempt to create a “general” theory of crime that applies to every American; instead, our theory explains how the unique experiences and worldview of blacks in America are related to their offending. In short, our theory draws on the strengths of both Afrocentricity and the Eurocentric canon. (Unnever and Gabbidon, 2011: 37)
Two kinds of racial injustices highlighted by the theory—racial discrimination and pejorative stereotyping—have empirical support. Blacks are more likely to express anger, exhibit low self-control and become depressed if they believe the racist stereotype that they’re violent. It’s also been studied whether or not a sense of racial injustice is related to offending when controlling for low self control (see below).
The core predictions of the TAAO and how they follow from it with references for empirical tests are as follows:
(Prediction 1) Black Americans with a stronger sense of racial identity are less likely to engage in criminal behavior than black Americans with a weak sense of racial identity. How does this prediction follow from the theory? TAAO suggests that a strong racial identity can act as a protective factor against criminal involvement. Those with a stronger sense of racial identity may be less likely to engage in criminal behavior as a way to cope with racial discrimination and societal marginalization. (Burt, Simons, and Gibbons, 2013; Burt, Lei, and Simons, 2017; Gaston and Doherty, 2018; Scott and Seal, 2019)
(Prediction 2) Experiencing racial discrimination increases the likelihood of black Americans engaging in criminal actions. How does this follow from the theory? TAAO posits that racial discrimination can lead to feelings of frustration and marginalization, and to cope with these stressors, some individuals may resort to committing criminal acts as a way to exert power or control in response to their experiences of racial discrimination. (Unnever, 2014; Unnever, Cullen, and Barnes, 2016; Herda, 2016, 2018; Scott and Seal, 2019)
(Prediction 3) Black Americans who feel socially marginalized and disadvantaged are more prone to committing crime as a coping mechanism and have weakened school bonds. How does this follow from the theory? TAAO suggests that those who experience social exclusion and disadvantage may turn to crime as a way to address their negative life circumstances. and feelings of agency. (Unnever, 2014; Unnever, Cullen, and Barnes, 2016)
The data show that there is a racialized worldview shared by blacks, and that a majority of blacks believe that their fate rests on what generally happens to black people in America. Around 38 percent of blacks report being discriminated against and most blacks are aware of the stereotype of them as violent. (Though a new Pew report states that around 8 in 10—about 80 percent—of blacks have experienced racial discrimination.) Racial discrimination and the belief in the racist stereotype that blacks are more violent are significant predictors of black arrests. It’s been shown that the more blacks are discriminated against and the more they believe that blacks are violent, the more likely they are to be arrested. Unnever and Gabbidon also theorized that the aforementioned isn’t just related to criminal offending but also to substance and alcohol abuse. Unnever and Gabbidon also hypothesized that racial injustices are related to crime since they increase the likelihood of experiencing negative emotions like anger and depression (Simons et al, 2002). It’s been experimentally demonstrated that blacks who perceive racial discrimination and who believe the racist stereotype that blacks are more violent express less self-control. The negative emotions from racial discrimination predict the likelihood of committing crime and similar behavior. It’s also been shown that blacks who have less self-control, who are angrier and are depressed have a higher liklihood of offending. Further, while controlling for self-control, anger and depression and other variables, racial discrimination predicts arrests and substance and alcohol abuse. Lastly the experience of being black in a racialized society predicts offending, even after controlling for other measures. Thus, it is ruled out that the reason why blacks are arrested more and perceive more racial injustice is due to low self-control. (See Unnever, 2014 for the citations and arguments for these predictions.) The TAAO also has more empirical support than racialized general strain theory (RGST) (Isom, 2015).
So the predictions of the theory are: Racial discrimination as a contributing factor; a strong racial identity could be a protective factor while a weak racial identity would be associated with a higher likelihood of engaging in criminal activity; blacks who feel socially marginalized would turn to crime as a response to their disadvantaged social position; poverty, education and neighborhood conditions play a significant role in black American offending rates, and that these factors interact with racial identity and discrimination which then influence criminal behavior; and lastly it predicts that the criminal justice system’s response to black American offenders could be influenced by their racial identity and social perceptions which could then potentially lead to disparities in treatment compared to other racial groups.
Ultimately, the unique experiences of black Americans explain why they commit more crime. Thus, given the unique experiences of black Americans, there needs to be a race-centric theory of crime for black Americans, and this is exactly what the TAAO is. The predictions that Unnever and Gabbidon (2011) made from the TAAO have independent empirical support. This is way more than the hereditarian explanations can say on why blacks commit more crime.
One way, which follows from the theory, to insulate black youth from discrimination and prejudice is racial socialization, where racial socialization is “thoughts, ideas, beliefs, and attitudes regarding race and racism are communicated across generations (Burt, Lei, & Simons, 2017; Hughes, Smith, et al., 2006; Lesane-Brown, 2006) (Said and Feldmeyer, 2022).
But also related to the racial socialization hypothesis is the question “Why don’t more blacks offend?” Gaston and Doherty (2018) set out to answer this question. Gaston and Doherty (2018) found that positive racial socialization buffered the effects of weak school bonds on adolescent substance abuse and criminal offending for males but not females. This is yet again another prediction from the theory that has come to pass—the fact that weak school bonds increase criminal offending.
Doherty and Gaston (2018) argue that black Americans face racial discrimination that whites in general just do not face:
Empirical studies have pointed to potential explanations of racial disparities in violent crimes, often citing that such disparities reflect Black Americans’ disproportionate exposure to criminogenic risk factors. For example, Black Americans uniquely experience racial discrimination—a robust correlate of offending—that White Americans generally do not experience (Burt, Simons, & Gibbons, 2012; Caldwell, Kohn-Wood, Schmeelk-Cone, Chavous, & Zimmerman, 2004; Simons, Chen, Stewart, & Brody, 2003; Unnever, Cullen, Mathers, McClure, & Allison, 2009). Furthermore, Black Americans are more likely to face factors conducive to crime such as experiencing poor economic conditions and living in neighborhoods characterized by concentrated disadvantage.
They conclude that:
The support we found for ethnic-racial socialization as a crime-reducing factor has important implications for broader criminological theorizing and practice. Our findings show the value of race-specific theories that are grounded in the unique experiences of that group and focus on their unique risk and protective factors. African Americans have unique pathways to offending with racial discrimination being a salient source of offending. While it is beyond the scope of this study to determine whether TAAO predicts African American offending better than general theories of crime, the general support for the ethnic-racial socialization hypothesis suggests the value of theories that account for race-specific correlates of Black offending and resilience.
…
TAAO draws from the developmental psychology literature and contends, however, that positive ethnic-racial socialization offers resilience to the criminogenic effect of weak school bonds and is the main reason more Black Americans do not offend (Unnever & Gabbidon, 2011, p. 113, 145).
Thus, combined with the fact that blacks face racial discrimination that whites in general just do not face, and combined with the fact that racial discrimination has been shown to increase criminal offending, it follows that racial discrimination can lead to criminal offending, and therefore, to decrease criminal offending we need to decrease racial discrimination. Since racism is due to low education and borne of ignorance, then it follows that education can decrease racial attitudes and, along with it, decrease crime (Hughes et al, 2007; Kuppens et al, 2014; Donovan, 2019, 2022).
Even partial tests of the TAAO have shown that racial discrimination related to offending and I would say that it is pretty well established that positive ethnic-racial socialization acts as a protective factor for blacks—this also explains why more blacks don’t offend (see Gaston and Doherty, 2018). It is also know that bad (ineffective) parenting also increases the risk for lower self-control (Unnever, Cullen, and Agnew, 2006). Black Americans share a racialized worldview and they view the US as racist, due to their personal lived experiences with racism (Unnever, 2014).
The TAAO and situationism
Looking at what the TAAO is and the predictions it makes, we can see how the TAAO is a situationist theory. Situationism is a psychological-philosophical theory which emphasizes the influence of the situation and its effects on human behavior. It posits that people’s actions and decisions are primarily shaped by the situational context that they find themselves in. It highlights the role of the situation in explaining behavior, suggests that people may act differently based on the context they find themselves in, situational cues which are present in the immediate context of the environment can trigger specific behavioral responses, suggests that understanding the situation one finds themselves in is important in explaining why people act the way they do, and asserts that behavior is more context-dependent and unpredictable and could vary across different situations. Although it seems that situationism conflicts with action theory, it doesn’t. Action theory explains how people form intentions and make decisions within specific situations, basically addressing the how and why. Conversely, situationism actually compliments action theory, since it addresses the where and when of behavior from an external, environmental perspective.
So the TAAO suggests that experiencing racial discrimination can contribute to criminal involvement as a response to social marginalization. So situationism can provide a framework for exploring how specific instances of environmental stressors, discrimination, or situational factors can trigger criminal behavior in context. So while TAAO focuses on historical and structural factors which lead to why blacks commit more crime, adding in situationism could show how the situational context interacts with historical and structural factors to explain black American criminal behavior.
Thus, combining situationism and the TAAO can lead to novel predictions like: predictions of how black Americans when faced with specific discriminatory situations, may be more or less likely to engage in criminal behavior based on their perception of the situation; predictions about the influence of immediate peer dynamics in moderating the relationship between structural factors like discrimination and criminal behavior in the black American community; and predictions about how variations in criminal responses to different types of situational cues—like encounters with law enforcement, experiences of discrimination, and economic stress—within the broader context of the TAAO’s historical-structural framework.
Why we should accept the TAAO over hereditarian explanations of crime
Overall, I’ve explained why hereditarian explanations of crime fail. They fail because when looking at the recent literature, the claims they make just do not hold up. Most importantly, as I’ve shown, hereditarian explanations lack empirical support, and the logic they try to use in defense of them is flawed.
We should accept the TAAO over hereditarianism because there is empirical validity, in that the TAAO is grounded in empirical research and it’s predictions and hypotheses have been subject to empirical tests and they have been found to hold. The TAAO also recognizes that crime is a complex phenomena influenced by factors like historical and contemporary discrimination, socioeconomic conditions, and the overall situational context. It also addresses the broader societal issues related to disparities in crime, which makes it more relevant for policy development and social interventions, acknowledging that to address these disparities, we must address the contemporary and historical factors which lead to crime. The TAAO also doesn’t stigmatize and stereotype, while it does emphasize the situational and contextual factors which lead to criminal activity. On the other hand, hereditarian theories can lead to stereotypes and discrimination, and since hereditarian explanations are false, we should also reject them (as I’ve explained above). Lastly, the TAAO also has the power to generate specific, testable predictions which have clear empirical support. Thus, to claim that hereditarian explanations are true while disregarding the empirical power of the TAAO is irrational, since hereditarian explanations don’t generate novel predictions while the TAAO does.
Conclusion
I have contrasted the TAAO with hereditarian explanations of crime. I showed that the three main hereditarian explanations—racial differences in testosterone and testosterone caused aggression, the AR gene, and MAOA—all fail. I have also shown that the TAAO is grounded in empirical research, and that it generates specific, testable predictions on how we can address racial differences in crime. On fhe other hand, hereditarian explanations lack empirical support, specificity, and causality, which makes it ill-suited for generating testable predictions and informing effective policies. The TAAO’s complexity, empirical support, and potential for addressing real-world issues makes it a more comprehensive framework for understanding and attempting to ameliorate racial crime disparities, in contrast to the genetic determinism from hereditarianism. In fact, I was unable to find any hereditarian response to the TAAO, so that should be telling on its own.
Overall, I have shown that the TAAO’s predictions that Unnever and Gabbidon have generated enjoy empirical support, and I have shown that hereditarian explanations fail, so we should reject hereditarian explanations and accept the TAAO, due to the considerations above. I have also shown that the TAAO makes actionable policy recommendations, and therefore, to decrease criminal offending, we thusly need to educate more, since racism is borne of ignorance and education can decrease racial bias.
Ashkenazi Jews Are White
2700 words
Introduction
Recently, I have been seeing people say that Ashkenazi Jews (AJs) are not white. Some may say that Jews “pretend to be white”, so they can accomplish their “group goals” (like pitting whites and blacks against each other in an attempt to sow racial strife, due to their ethnic nepotism due to their genetic similarity). I have also seen people deriding Jews for saying “I’m white” and then finding an instance of them saying “I’m Jewish” (see here for an example), as if that’s a contradiction, but it’s not. It’s the same thing as saying “I’m Italian… I’m white” or “I’m German… I’m white.” But since pluralism about race is true, there could be some contexts and places that Jews aren’t white, due to the social construction of racial identities. However, in the American context it is quite clear: In both historical and contemporary thought in America, AJs are white.
But a claim like this, then, raises an important question: If AJs are not white, then what race are they? This is a question I will answer in this article, and I will of course show that AJs are indeed white in an American conception of race. Using Quayshawn Spencer’s racial identity argument, I will assume that Ashkenazi Jews aren’t white, and then I will argue that this leads to a contradiction, so Jews must be white. And while there was discussion about the racial status of Jews after they began emigrating to America through Ellis Island, I will show that Jews arrived to America as whites.
White or not?
The question of whether or not AJs are white is a vexing one. Of course, AJs are a religious group. However, this doesn’t mean that they themselves have their own specific racial category. It’s like if one says they are German, or Italian, or British. Those are mere ethnicities which make up the white racial group. One study found that AJs have “White privilege vis-á-vis persons of color. This privilege, however, is limited to Jews who can “pass” as White gentiles” (Blumenfeld, 2009). Jews that can “pass as white” are obviously white, and there is no other race for them to be.
This is due to the social nature of race. Since race is a social construct, then the way people’s racial background is perceived in America is based on how they look (their phenotype). An Ashkenazi Jew saying “I’m Jewish. I’m white” isn’t a contradiction, since AJs aren’t a race. It’s just like saying “I’m Italian. I’m white” or “I’m German. I’m white.” It’s quite obviously an ethnic group which is a part of the white race. Jews are white and whites are a socialrace.
This discussion is similar to the one where it is claimed that “Hispanic/Latino/Spanish” aren’t white. But that, too, is a ridiculous claim. In cluster studies, HLSs don’t have their own cluster, but they cluster near the group where their majority ancestry derives (Risch et al, 2002). Saying that AJs aren’t white is similar to this.
But during WWII, Jews were persecuted in Nazi German and eventually some 6 million Jews were killed. Jews, in this instance, were seen as a socialrace in Germany, and so they were themselves racialized. It has been shown that Germans who grew up under their Nazi regime are much more anti-Semitic than Germans who were born before or after the Nazi regime, and it was Nazi schooling which contributed to this the most (Voigtlander and Voth, 2015). This shows how one’s beliefs—and that of a whole society’s—are malleable along with how effective propaganda is. The Nuremberg laws of 1935 established anti-Jewish sentiment in the Nazi racial state, and so they had to have a way to identify Jews. They settled on the religious affiliation of one’s 4 grandparents as a way to identify Jews. But when one’s origins were in doubt, the Reich Kinship Office was deployed in order to ascertain one’s genealogy. But in the event this could not be done, one’s physical attributes would be assessed and compared to 120 physical measures between the individual and their parents (Rupnow, 2020: 373-374).
This can now be centered on Whoopi Goldberg’s divisive comment from February, 2022, where she states that the attempted genocide of Jews in Nazi Germany “wasn’t about race“, but it was about “man’s inhumanity to man; [it involved] two groups of white people.” Of course Goldberg is operating under an American conception of race, so I could see why she would say that. However, at the time in Nazi Germany, Jews were Racialized Others, and so they were a socialrace in Germany.
Per Pew, most Jews in America identify as white:
92% of U.S. Jews describe themselves as White and non-Hispanic, while 8% say they belong to another racial or ethnic group. This includes 1% who identify as Black and non-Hispanic; 4% who identify as Hispanic; and 3% who identify with another race or ethnicity – such as Asian, American Indian or Hawaiian/Pacific Islander – or with more than one race.
A super majority (94%) of American Jews are (and identify as) white and non-“Hispanic” in Pew’s 2013 research, which is down slightly from the 2020 research (Lugo et al, 2013):

AJs were viewed as white even as early as 1790 when the Naturalization Act was put into law, which stated that only free white persons were allowed to emigrate to America (Tanner, 2021). Even in 1965, Srole (1965) stated that “Jews are white.” But the perception that all Jews are white came after WWII (Levine-Rasky, 2020) and this claim is of course false. All Jews certainly aren’t white, but some Jews are white. Thus, even historically in the history of America, AJs were seen as white. Yang and Koshy (2016) write:
We found no evidence from U.S. censuses, naturalization legislation, and court cases that the racial categorization of some non-Anglo-Saxon European immigrant groups such as the Irish, Italians, and Jews changed to white. They were legally white and always white, and there was no need for them to switch to white.
…
White ethnics could be considered ethnically inferior and discriminated against because of their ethnic distinctions, but in terms of race or color, they were all white and had access to resources not available to nonwhites.
It was precisely because of the changing meanings of race that “the Irish race,” “the German race,” “the Dutch race,” “the Jewish race,” “the Italian race,” and so on changed their races and became white. In today’s terminology, it should be read that these European groups changed their ethnicities to become part of whites, or more precisely they were racialized to become white.
…
Our findings help resolve the controversy over whether certain U.S. non-Anglo-Saxon European immigrant groups became white in historical America. Our analysis suggests that “becoming white” carries different meanings: change in racial classification, and change in majority/minority status. In terms of the former, “becoming white” for non-Anglo-Saxon European immigrant groups is bogus. Hence, the argument of Eric Arnesen (2001), Aldoph Reed (2001), Barbara Fields (2001), and Thomas Guglielmo (2003) that the Irish, Italians, and Jews were white on arrival in America is vindicated.
But one article in The Forward argued that “Ashkenazi Jews are not functionally white.” The author (Danzig) attempts to make an analogy between the founder of the NAACP Walter White who was “white-passing” (both of his parents were born into slavery) and Jews who are “white-passing”, “due to years of colonialism, expulsion and exile in European lands.” The author then claims that as along as Jews maintain their unique Jewish identity, they therefore are a racial group. This article is a response to another which claims that Ashkenazi Jews are” functionally white” (Burton). Danzig discusses Button’s claim that a “white-passing ‘Latinx'” person could be deported if their immigration status is discovered. This of course implies that “Hispanics” are themselves a racial group (they aren’t). Danzig discusses the discrimination that his family went through in the 1920s, stating that they could do certain things because they were Jewish. The argument in Danzig’s article, I think, is confused. It’s confused because just because Jews were discriminated against in the past doesn’t mean they weren’t white. In fact, Jews, Italians, and the Irish were white on arrival to the United States (Steward, 1964; Yang and Koshy, 2016). But this doesn’t mean that they didn’t face discrimination. That is, Jews, Italians and the Irish didn’t change to white they were always legally white in America. (But see Gardaphe, 2002, Bisesi, 2017, Baddorf, 2020, and Rubin, 2021. Italians didn’t become white as those authors claim, they were white upon arrival). So Danzig’s claim fails—Jews are functionally white because they are white and they arrived in America as white. Claims to the contrary that AJs (and Italians and the Irish) became white are clearly false.
So despite claims that Jews became white after WWII, Jews are in fact white in America (Pearson and Geronimus, 2011). Of course in the early 1900s as immigrants were arriving to Ellis Island, the question of whether or not Jews (“Hebrews” in this instance) were white or even if they were their own racial group had a decent amount of discussion at the time (Goldstein, 2005; Pearlman, 2018). The fact that there was ethnic strife between new-wave immigrants to Ellis Island doesn’t entail that they were racial groups or that those European immigrants weren’t white. It’s quite clear that Jews—like italians and the Irish—were considered white upon arrival.
Now that I have established the fact that Jews AJs are indeed white (and arrived to America as white) despite the confused protestations of some authors, now I will formalize the argument that AJs are white, since if they aren’t white, then they would need to fit into one of the other 4 racial categories.
Many may know that I push Quayshawn Spencer’s OMB race theory, and that I am a pluralist about race. In the volume What is Race?: Four Philosophical Views, philosopher or race Quayshawn Spencer (2019: 98) writes:
After all, in OMB race talk, White is not a narrow group limited to Europeans, European Americans, and the like. Rather, White is a broad group that includes Arabs, Persians, Jews, and other ethnic groups originating from the Middle East and North Africa.
Although there is some research on the racial identity of MENA (Middle Eastern/North African people) and how they may not perceive themselves as white or be perceived as white (Maghbouleh, Schachter, and Flores, 2022), the OMB is quite clear that the social group designated “white” doesn’t refer only to Europeans (Spencer, 2019).
So, if AJs aren’t white, then they must be part of another of the 4 OMB races (black, Native American, East Asian or Pacific Islander). Part of this racial scheme is K=5—where when K is set to 5 in STRUCTURE, 5 clusters are spit out and these map onto the OMB races. But of those 5 clusters, there is no Jewish cluster. Note that I am not denying that there is some kind of genetic structure to AJs, I’m just denying that this would entail that they are a racial group. If they were, then they would appear in these runs. AJs are merely an ethno-religious in the white socialrace. So let’s assume this is true: Ashkenazi Jews are not white.
When we consider the complexities of racial classification, it becomes apparent that societies tend to organize individuals on numerous traits into distinct categories based on physical traits, cultural background, and ancestry. If AJs aren’t white in an American context, then they would have to fall into one of the four other racial groups in a Spencerian OMB race theory.
But there is one important aspect to consider here—that of the phenotype of Ashkenazi Jews. Many Ashkenazi Jews exhibit physical traits which are more likely associated with “white” populations. This simple observation shows that AJs don’t fit into the established categories of East Asian, Pacific Islander, black or Native American. AJs’ typical phenotype aligns more closely with that of white populations.
So examining the racial landscape in America, we can see that how social perceptions and classifications can significantly impact how individuals are positioned in a broader framework. AJs have historically been classified and perceived as white in the American racial context, as can be seen above. So within American racetalk, AJs are predominantly classified in the white racial grouping.
So taking all of this together, I can rightly state that Jews are white. Since we assumed at the outset that if they weren’t white they would belong to some other racial group, but they don’t look like any other racial group but look and are treated as white (both in contemporary thought and historically), then AJs are most definitely seen as white in American racetalk. Here’s the formalized argument:
P1: If AJs aren’t white, then they must belong to one of the other 4 racial categories (black, Native American, East Asian or Pacific Islander).
P2: AJs do not belong to any of the four racial categories mentioned (based on their phenotype typical of white people).
P3: In the American racial context, AJs are predominantly classified and perceived as white.
Conclusion: from P1, if AJs aren’t white then they must belong to one of the other 4 racial groups. But from P2, AJs do not belong to any of those categories, because from P3, AJs are perceived and classified as white. These premises, then, lead to a contradiction, since they all cannot be simultaneously true.
So we must reject the assumption that AJs aren’t white, and the logical conclusion is that AJs are considered white in the American context, based on their phenotype (and the fact that they arrived to America as white). Jews didn’t “become white” like some claim (eg, Brodkin, 2004). American Jews even benefit from white privilege (Schraub, 2019). MacDonald-Dennis’ (2005, 2006) qualitative research (although small not generalizable) shows that some Ashkenazi Jews think of themselves as white. AJs are legally and politically white.
All Jews aren’t white, but some (most) Jews are white (in America).
Conclusion
Thus, AJs are white. Although many authors have claimed that Jews became white after arrival to America (or even after WWII), this claim is false. It is false even as far back as 1790. If we accept the assumption that AJs aren’t white, then it leads to a contradiction, since they would have to be one of the other 4 racial groups, but since they look white, they cannot be a part of those racial groups.
There are white Jews and there are non-white Jews. But when it comes to AJs, the question “When did they become white?” is nonsense since they were always perceived and treated as white in America from it’s founding. Some AJs are white, some aren’t; some Mizrahi Jews are white, some aren’t. However in the context of this discussion, it is quite clear that AJs are white, and there is no other race for them to be, based on the OMB race theory. In fact, in the minds of most Americans, Jews aren’t a racialized group, but they are perceived as outsiders (Levin, Filindra, and Kopstein, 2022). But there were some instances in history where sometimes Jews were racialized, and sometimes they weren’t (Hochman, 2017). But what I have decisively shown here, in the American context ever since its inception, AJs are most definitely white. Saying that AJs are white is like saying that Italians or Germans are white. There is no contradiction. Jews get treated as white in the American social context, they look white, and have been considered white since they have arrived to America in the early 1900s (like the Irish and Italians).
The evidence and reasoning presented in this article points to one conclusion: That AJs are indeed white. This of course doesn’t mean that all AJs are white, it merely means that some (and I would say most) are white. AJs have been historically, legally, and politically white. Mere claims that they aren’t white are irrelevant.
